
96
ÂWIAT NAUKI
Zaktualizowane ze Âwiata Nauki numer 4/2001
Chromosomy determinujàce p∏eç ludzi – Xi Y – bardzo si´ od siebie ró˝nià. Pozosta-
∏e 22 pary chromosomów w naszych komórkach sà dobrze dobrane – jeden chromosom
z ka˝dej pary pochodzi od matki, drugi od ojca, ale obydwa sà tej samej wielkoÊci i ma-
jà te same geny. Natomiast chromosom Y jest znacznie mniejszy od X, wr´cz lilipuci. Za-
wiera zaledwie kilkadziesiàt genów, podczas gdy chromosom Xa˝ 2–3 tys. Wiele genów
znajdujàcych si´ w Y nie ma odpowiedników w X. Ponadto w Y jest niezmiernie du˝o
CHROMOSOM Y
Nasze chromosomy X i Y
to osobliwa para.
X przypomina
inne chromosomy,
ale Y – êród∏o m´skoÊci
– jest niezwyk∏y
Karin Jegalian i Bruce T. Lahn
y
BIOPHOTO ASSOCIA
TES
Photo R
esear
chers, Inc.
C O D A L E J ?
Dlaczego
Êmieciowego DNA: sekwencji nukleoty-
dów nie stanowiàcych instrukcji wytwa-
rzania bia∏ek.
Do niedawna biolodzy nie potrafili
wyjaÊniç powodów odmiennoÊci Y. By-
∏y wprawdzie ró˝ne teorie na ten temat,
nie umiano jednak ich udowodniç. Sy-
tuacja uleg∏a zmianie dzi´ki Projek-
towi Poznania Ludzkiego Genomu i
innym wysi∏kom zmierzajàcym do od-
czytania sekwencji ludzkiego DNA we
wszystkich 24 chromosomach – czyli
X, Y i 22 autosomach (chromosomach
nie determinujàcych p∏ci). Podobnie jak
paleontolodzy, którzy badajàc szkiele-
ty wspó∏czesnych zwierzàt oraz skamie-
nia∏oÊci ich przodków, Êledzà ewolucj´
gatunków, biolodzy molekularni potra-
fià obecnie odtworzyç ewolucj´ chro-
mosomów i genów, rozszyfrowujàc se-
kwencje DNA.
Z najnowszych danych wynika, ˝e hi-
storia chromosomów p∏ci by∏a niezwy-
kle dynamiczna. Cechowa∏a jà seria
nag∏ych zmian w chromosomie Y, kom-
pensowanych zmianami w X. Te prze-
miany i oddzia∏ywania trwajà nadal.
Co wi´cej, chromosom Y – od daw-
na uwa˝any za niedobitka, przydatnego
w zasadzie tylko do rozpocz´cia pro-
gramu rozwoju osobnika p∏ci m´skiej
– okaza∏ si´ znacznie wa˝niejszy, ni˝
podejrzewa∏a wi´kszoÊç biologów. Przez
oko∏o 300 mln lat zachowa∏ niewielkà
liczb´ genów wa˝nych do prze˝ycia
samców i pozyska∏ nowe, potrzebne do
rozmna˝ania. Zamiast staç si´ Rod-
neyem Dangerfieldem* chromosomów
(tak niektórzy nazywali niezbyt szano-
wany chromosom Y), jest bardziej po-
dobny do Woody’ego Allena: mimo nie-
pozornego wyglàdu potrafi tak wiele
dokonaç.
jest taki dziwny
taki
CHROMOSOM X
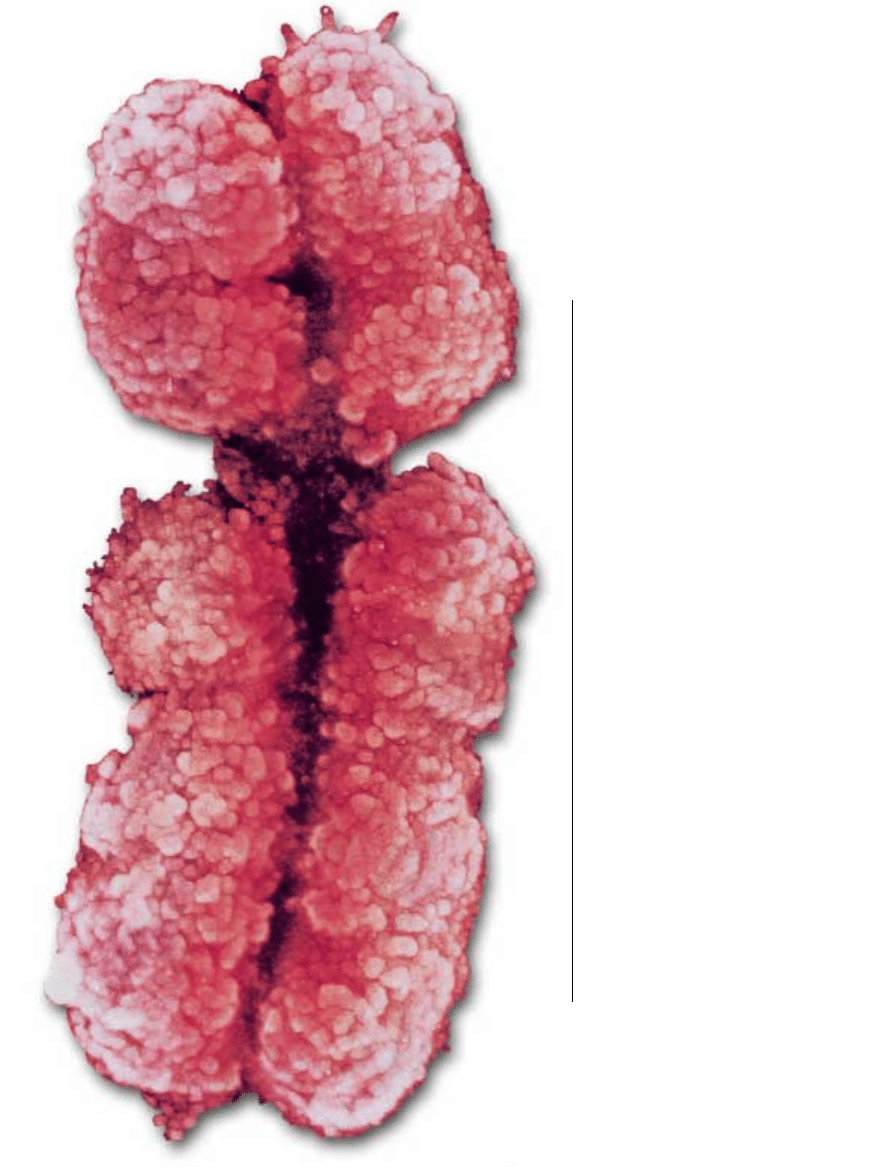
CHROMOSOMY X i Y setki milionów lat temu stanowi∏y dobranà par´.
Jednak Y skurczy∏ si´ znacznie, podczas gdy X pozosta∏ taki sam.
Zaczynamy rozumieç, jak to si´ sta∏o. Mikrofotografie przedstawiajà
chromosomy podczas metafazy – jednej z faz podzia∏u komórkowego.
WYDANIE SPECJALNE
ÂWIAT NAUKI
97
Si∏à nap´dowà wi´kszoÊci badaƒ nad
ewolucjà ludzkich chromosomów p∏ci
by∏a zwyk∏a ciekawoÊç. Ale istnia∏ te˝
inny powód: ch´ç zrozumienia przy-
czyn i leczenia bezp∏odnoÊci m´˝czyzn.
Odkrycie genów Y, majàcych wp∏yw na
zdolnoÊç do rozmna˝ania, mog∏oby za-
owocowaç nowymi terapiami dla osób,
u których brak tych genów lub sà uszko-
dzone [ramka na stronie 101].
Ostatnie osiàgni´cia to efekt ponad-
stuletnich badaƒ. W XIX wieku biolo-
dzy uwa˝ali, ˝e o p∏ci ludzi i innych ssa-
ków decyduje Êrodowisko, podobnie jak
u wspó∏czesnych gadów (temperatura
zarodka we wczesnym stadium rozwo-
ju rozstrzyga o p∏ci gadziego potomka).
Na poczàtku XX wieku naukowcy zrozu-
mieli, ˝e w przypadku niektórych ga-
tunków p∏eç jest determinowana przez
chromosomy. Oko∏o 20 lat póêniej wyka-
zano, ˝e dotyczy to tak˝e ssaków, a za
okreÊlenie p∏ci podczas rozwoju zarod-
kowego odpowiadajà chromosomy Xi Y.
Przybywa dowodów
W CI
ÑGU
nast´pnych kilkudziesi´ciu lat
naukowcy zidentyfikowali Y jako spraw-
c´ m´skoÊci i uznali, ˝e Xi Y wyewo-
luowa∏y z pary autosomów dawnego
przodka. Przypadkowo, tu˝ przed poja-
wieniem si´ ssaków lub wkrótce po tym
wydarzeniu, mutacja w jednej kopii au-
tosomu, który przekszta∏ci∏ si´ w prze-
sz∏oÊci w Y, spowodowa∏a, ˝e z zarod-
ków dziedziczàcych ten zmieniony
chromosom powstawa∏y samce, a z pozo-
sta∏ych – samice.
W 1990 roku genetycy okreÊlili, która
cz´Êç Y jest odpowiedzialna za m´s-
koÊç: gen zwany SRY (sex-determining
region Y – okreÊlajàcy p∏eç obszar Y).
Bia∏ko kodowane przez SRY odpowia-
da za wykszta∏cenie jàder, aktywujàc
zapewne geny na ró˝nych chromoso-
mach. Nast´pnie testosteron i inne sub-
stancje wytwarzane w jàdrach decydu-
jà o powstaniu cech m´skich.
98
ÂWIAT NAUKI
EWOLUCJA CZ¸OWIEKA /NOWE SPOJRZENIE
Regiony pseudoautosomalne,
w których zachodzi wymiana DNA
(rekombinacja) z X
†
Geny metabolizmu podstawowego
‡
Geny majàce odpowiedniki w X,
ale aktywne tylko w jàdrach
SRY
odpowiada za wykszta∏cenie si´
jàder. Pochodzi od genu SOX3
i jest podobny do genu SOX3 w X,
ale pe∏ni innà funkcj´
*
HESED P
ADILL
A-NASH
National Institutes of Health
(mikr
ofotografia
); ALFRED T
. KAMAJIAN i
HEIDI NOL
AND (
ilustracje z
lewej i
na gór
ze
)
1
7
13
19
20
21
22
X
Y
2
8
14
3
9
15
4
10
16
5
11
17
6
12
18
BPY2
PRY
CDY
DAZ
RBMY ‡
PRY
TTY2
RBMY ‡
EIF1AY †
SMCY †
Zmniejszona
zdolnoÊç
do produkcji
plemników
Zmniejszona
zdolnoÊç
do produkcji
plemników
CDY
XKRY
VCY ‡
USP9Y †
DBY †
UTY †
Centromer
PRY
TTY1
TTY2
TSPY
AMELY
RPS4Y †
SRY
(okreÊla
p∏eç m´skà)
TTY1
TSPY
Zmniejszona
zdolnoÊç
do produkcji
plemników
Niski
wzrost
Obszar
nie zawierajàcy
funkcjonalnych genów
Geny Y majàce
odpowiedniki
w X
Geny Y (lub rodziny genów)
nie spotykane w X i czynne
tylko w jàdrach
*
Konsekwencje
zwiàzane
z delecjà
odcinków Y
CHROMOSOM Y
ZFY †
PCDHY
TB4Y †
GADZI PRZODKOWIE SSAKÓW
350 mln
lat temu?
CZAS
Powstaje gen SRY
PARA AUTOSOMÓW
U GADZIEGO PRZODKA
POWSTAJÑCY
Y
SRY
Centromer
Identyczne
chromosomy
zdolne do
rekombinacji
(wymiany
odcinków)
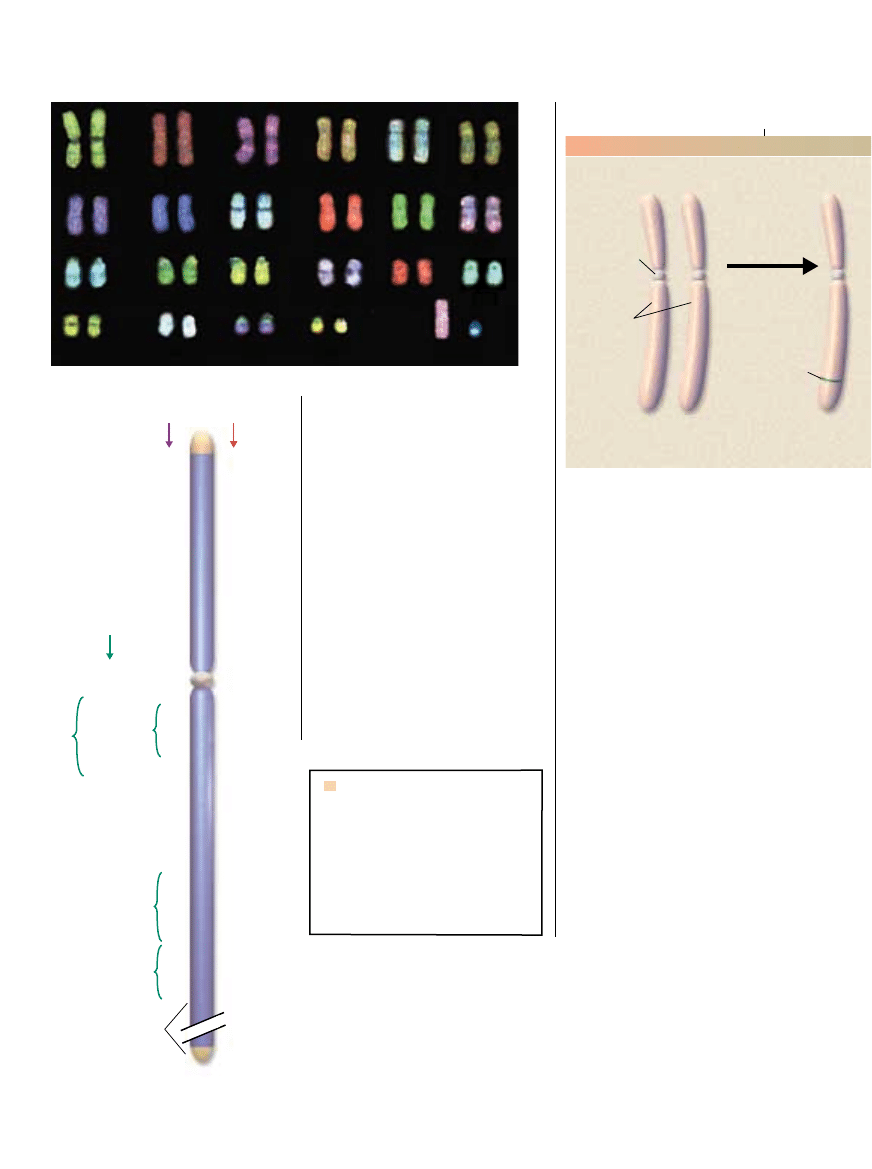
CHROMOSOMY z normalnej komórki m´˝czyzny (zdj´cie) to 22 pary autosomów
(nie determinujàcych p∏ci) oraz X i Y; w ka˝dej parze jeden chromosom pochodzi od matki,
a drugi od ojca. Geny w regionie nierekombinujàcym chromosomu Y (NRY – non-recombining
region Y; niebieski na diagramie) pomog∏y w zrozumieniu ewolucji X i Y. Region ten zosta∏
tak nazwany, gdy˝ w jego obr´bie nie zachodzi rekombinacja (wymiana DNA) z X.
Zestawiono tylko te geny, które sà nadal funkcjonalne. Oko∏o po∏owy ma odpowiedniki
w X (czerwony); niektóre z nich to geny metabolizmu podstawowego, niezb´dne do prze˝ycia
ogromnej wi´kszoÊci komórek. Pewne geny w NRY sà aktywne tylko w jàdrach (fioletowy),
decydujàc prawdopodobnie o m´skiej p∏odnoÊci.
Naukowcy doszli do wniosku, ˝e ludz-
kie chromosomy p∏ci rozpocz´∏y ˝ycie
jako dopasowana para, dlatego ˝e ich
koƒce sà bardzo podobne i zdolne do
rekombinacji. Podczas mejozy (podzia-
∏u komórek rozrodczych) chromosomy
homologiczne ustawiajà si´ w pary
i wymieniajà odcinki, po czym po jed-
nej kopii ka˝dego autosomu oraz po
jednym chromosomie p∏ci przechodzi
do ka˝dej komórki rozrodczej. Choç X
i Y sà obecnie podobne tylko w niewiel-
kim stopniu, podczas mejozy u sam-
ców ich koƒce ustawiajà si´ obok siebie
(jest to konieczne do w∏aÊciwego po-
dzia∏u chromosomów do plemników).
Inne dowody, ˝e Xi Y by∏y kiedyÊ po-
dobne, pochodzà z tej cz´Êci Y, która
z Xnie rekombinuje. Wiele genów znaj-
dujàcych si´ w owym regionie nadal
ma odpowiedniki w X.
Nierekombinujàcy obszar – 95% Y
– Êwiadczy, jak bardzo ten chromosom
sta∏ si´ w∏asnym cieniem. Rekombi-
nacja sprzyja zachowaniu integralnoÊci
chromosomów, jej brak zaÊ prowadzi
do nagromadzenia si´ w genach z niere-
kombinujàcych regionów niszczàcych
mutacji, wskutek czego te fragmenty
chromosomu cz´sto zanikajà. Przypusz-
czano wi´c, ˝e coÊ spowodowa∏o wy-
mian´ DNA mi´dzy du˝ymi cz´Êciami
Xi Y, po czym geny w nierekombinujà-
cym regionie Y przesta∏y funkcjonowaç.
Przez kilkadziesiàt lat nie wiedziano jed-
nak, w jaki sposób dosz∏o do zahamo-
wania rekombinacji po powstaniu Y.
Nowy wizerunek
WYNIKI BADA
¡
przeprowadzonych w cià-
gu ostatnich lat wype∏ni∏y wiele luk. Na
przyk∏ad w 1999 roku Lahn oraz David
C. Page z Whitehead Institute for Bio-
medical Research w Cambridge w Mas-
sachusetts wykazali, ˝e Y nieoczekiwa-
nie straci∏ zdolnoÊç do wymiany DNA
z Xi by∏ to proces wieloetapowy. Po-
czàtkowo zjawisko to dotyczy∏o DNA
otaczajàcego gen SRY, a nast´pnie roz-
przestrzeni∏o si´ w kilku odr´bnych
etapach na nieomal ca∏à d∏ugoÊç chro-
mosomu Y. W rezultacie utraty wymia-
ny X–Y uwsteczni∏ si´ tylko Y, X zaÊ na-
dal podlega∏ rekombinacji, gdy dwie
jego kopie spotyka∏y si´ podczas mejo-
zy u samic.
Czym mo˝na wyt∏umaczyç zaburze-
nie rekombinacji mi´dzy Xi Y? Gdy
wczesne wersje obu tych chromosomów
mia∏y wymieniaç si´ odcinkami pod-
czas mejozy u jakiegoÊ odleg∏ego przod-
ka wspó∏czesnych ssaków, cz´Êç DNA
Y prawdopodobnie si´ odwróci∏a, czy-
li znalaz∏a si´ do góry nogami wzgl´-
dem równowa˝nej cz´Êci X. Poniewa˝
rekombinacja wymaga ustawienia si´
dwóch podobnych sekwencji DNA obok
siebie, taka inwersja mog∏a spowodo-
waç zniesienie interakcji mi´dzy uprzed-
nio dopasowanymi regionami Xi Y.
Gdy zbadaliÊmy sekwencje nukleoty-
dów 19 genów, które pojawiajà si´
w nierekombinujàcym regionie zarów-
no X, jak i Y (niektóre z kopii Y ju˝ nie
dzia∏ajà), odkryliÊmy, ˝e do zaniku re-
kombinacji dosz∏o w serii kolejnych
zdarzeƒ. Na ogó∏ jeÊli para genów traci
t´ zdolnoÊç, ich sekwencje z up∏ywem
czasu stajà si´ coraz bardziej ró˝ne.
Stosunkowo niewielka liczba ró˝nic
wskazuje na niedawne zaprzestanie re-
kombinacji, du˝a zaÊ – ˝e nie zachodzi
ona ju˝ od dawna.
Wi´kszoÊç par X–Y nale˝a∏a do jednej
z czterech grup. W ka˝dej z nich chro-
mosomy te ró˝ni∏y si´ w zasadzie w tym
samym procencie, co wskazywa∏oby, ˝e
rekombinacja usta∏a mniej wi´cej w
tym samym czasie. Ale ró˝nice pomi´-
dzy grupami by∏y jednak wyraêne. Ko-
WYDANIE SPECJALNE
ÂWIAT NAUKI
99
SSAKI
320–240 mln
lat temu
170–130 mln
lat temu
130–80 mln
lat temu
50–30 mln
lat temu
Obecnie
1
przypadek
niezdolnoÊci
do rekombinacji,
prowadzàcy
do degeneracji
i skrócenia
cz´Êci Y
2
przypadek
niezdolnoÊci
do rekombinacji,
powodujàcy
dalsze
uwstecznienie Y
3
przypadek
niezdolnoÊci
do rekombinacji,
wiodàcy
do kolejnego
skrócenia Y
4
przypadek
niezdolnoÊci
do rekombinacji,
prowadzàcy
do obecnego,
silnie
uwstecznionego Y
POWSTAJÑCY
X
Y X
JAK U TORBACZY
1
2
3
4
Y X
JAK U STEKOWCÓW
Y X
JAK U MA¸P
Y X
U LUDZI
Obszary
niezdolne ju˝
do rekombinacji
Obszary
nadal zdolne
do rekombinacji
W jakimÊ
nieznanym
momencie SRY
przemieÊci∏ si´
na krótkie
rami´ Y
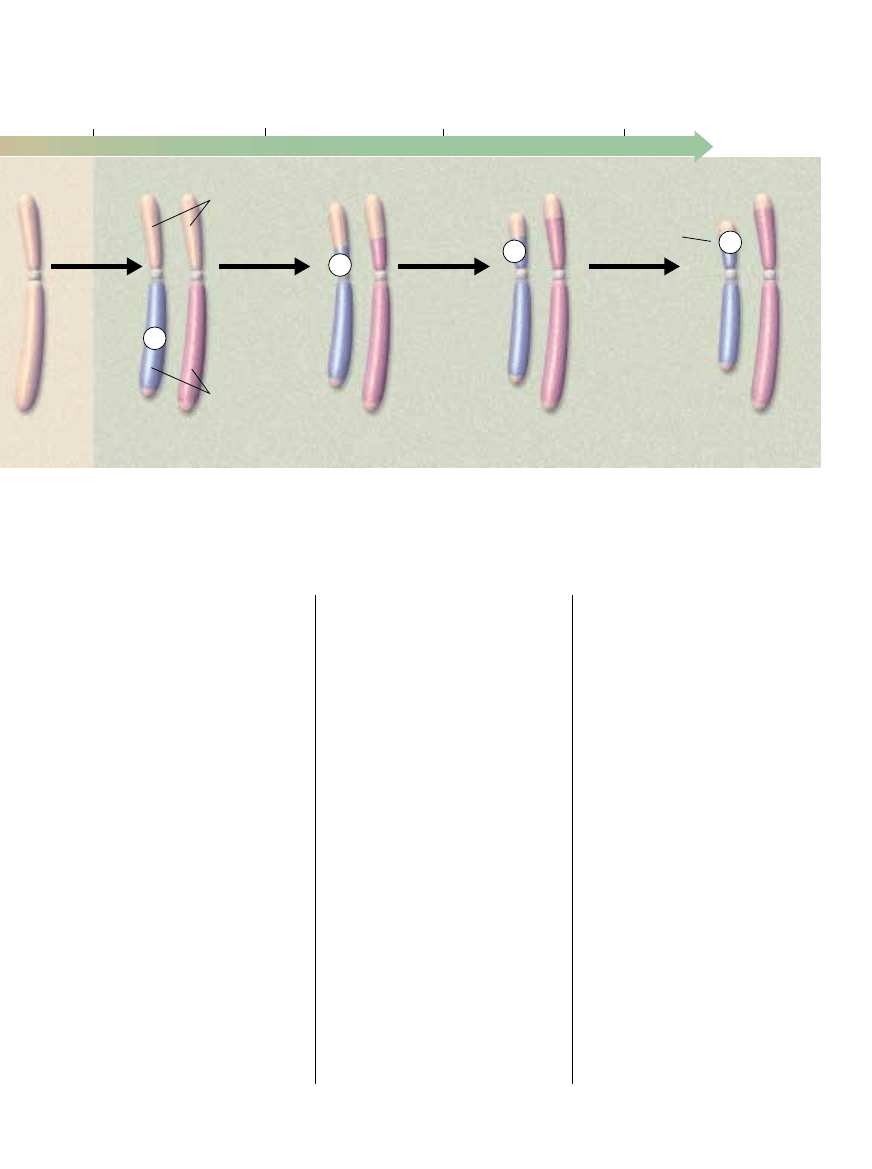
DEGENERACJA Y przebiega∏a w czterech odr´bnych etapach, od oko∏o 300 mln lat, gdy b´dàcy przodkiem ssaków gad ssakokszta∏tny
zyska∏ nowy gen (SRY) na jednym ze swoich autosomów. Ka˝dy z epizodów by∏ nast´pstwem braku rekombinacji (wymiany DNA)
mi´dzy X i Y podczas mejozy, podzia∏u komórki prowadzàcego do powstania komórek jajowych lub plemników. JeÊli rekombinacja nie jest
mo˝liwa, geny z tych obszarów przestajà funkcjonowaç i ulegajà degeneracji. Przedstawiona tu sekwencja zdarzeƒ jest bardzo uproszczona.
Na przyk∏ad w pewnych okresach Y powi´ksza∏ si´ wskutek wprowadzania DNA autosomów do regionów nadal zdolnych
do rekombinacji, zanim jej utrata spowodowa∏a jego kurczenie si´.
pie genów Y, które zacz´∏y ró˝nicowaç
si´ od swoich odpowiedników na X
w chwili powstania genów SRY, wyka-
zywa∏y w stosunku do swoich partne-
rów najwi´kszà odmiennoÊç, natomiast
w obr´bie innych grup ró˝nice te by∏y
coraz mniejsze.
Porównujàc sekwencje DNA mi´dzy
gatunkami, biolodzy mogà w przybli˝e-
niu wyliczyç, kiedy podobne wczeÊniej
geny (a wi´c i obszary, w których si´
znajdujà) zacz´∏y si´ ró˝nicowaç. Wy-
nika stàd, ˝e autosomalne prekursory
Xi Y by∏y nadal podobne i nie zmienio-
ne u gadów ˝yjàcych w okresie poprze-
dzajàcym oddzielenie si´ ewolucyjnej
linii ssaków. Stekowce (jak dziobak i kol-
czatka), które najwczeÊniej oddzieli∏y
si´ od innych grup ssaków, majà zarów-
no gen SRY, jak i przyleg∏y do niego,
nierekombinujàcy region. A zatem do
powstania tego genu i zaniku rekombi-
nacji w jego otoczeniu dosz∏o prawdo-
podobnie wówczas, gdy oddzieli∏a si´
linia ewolucyjna wiodàca do ssaków,
czyli oko∏o 300 mln lat temu.
Stosujàc analiz´ „zegara molekular-
nego”, uzyskaliÊmy wi´cej informacji
na ten temat. Biolodzy sà w stanie
oszacowaç, kiedy mo˝e dojÊç do zmia-
ny sekwencji DNA, o ile nie jest na nie
wywierany szczególny nacisk, by po-
zosta∏y takie same. Mno˝àc stopieƒ roz-
bie˝noÊci sekwencji w parach X–Y
przez szacunkowe tempo zmian, wyli-
czyliÊmy, ˝e pierwsza inwersja, która
zatrzyma∏a rekombinacj´, zasz∏a przed
320–240 mln lat. Podobne analizy
wskazujà, ˝e nast´pna zdarzy∏a si´
170–130 mln lat temu, na krótko przed
oddzieleniem si´ torbaczy od linii pro-
wadzàcej do ssaków ∏o˝yskowych. Trze-
100
ÂWIAT NAUKI
EWOLUCJA CZ¸OWIEKA /NOWE SPOJRZENIE
RAZI SEARLES
Br
uce Coleman Inc.
(kot
); ALFRED T
. KAMAJIAN
ZDARZENIE 1
Kopia genu na Y
degeneruje si´
ze wzgl´du
na brak
rekombinacji
mi´dzy X i Y
ZDARZENIE 2
Podwaja si´
aktywnoÊç
genu na X, by
skompensowaç
niedobór bia∏ka
u samców
ZDARZENIE 3
Samice
inaktywujà
losowo jednà
kopi´ genu
w ka˝dej
komórce
Stan
wyjÊciowy
Komórka
m´ska
Komórka
˝eƒska
EFEKT KO¡COWY
Wyrównanie
poziomu bia∏ek
u samców
i samic
EFEKT
Samce wytwarzajà
dwa razy mniej bia∏ka
ni˝ samice
EFEKT
Brak zmian
u samic
EFEKT
Samice
wytwarzajà teraz
za du˝o bia∏ka
EFEKT
Ogólna produkcja
bia∏ek u samców
powraca do normy
EFEKT
Brak zmian
u samców
EFEKT
Poziom bia∏ka
u samic powraca
do normy
Bia∏ka
kodowane
przez gen
Aktywne
kopie
genu
Wy∏àczony
gen
Utracony
gen
Y
X
X
X
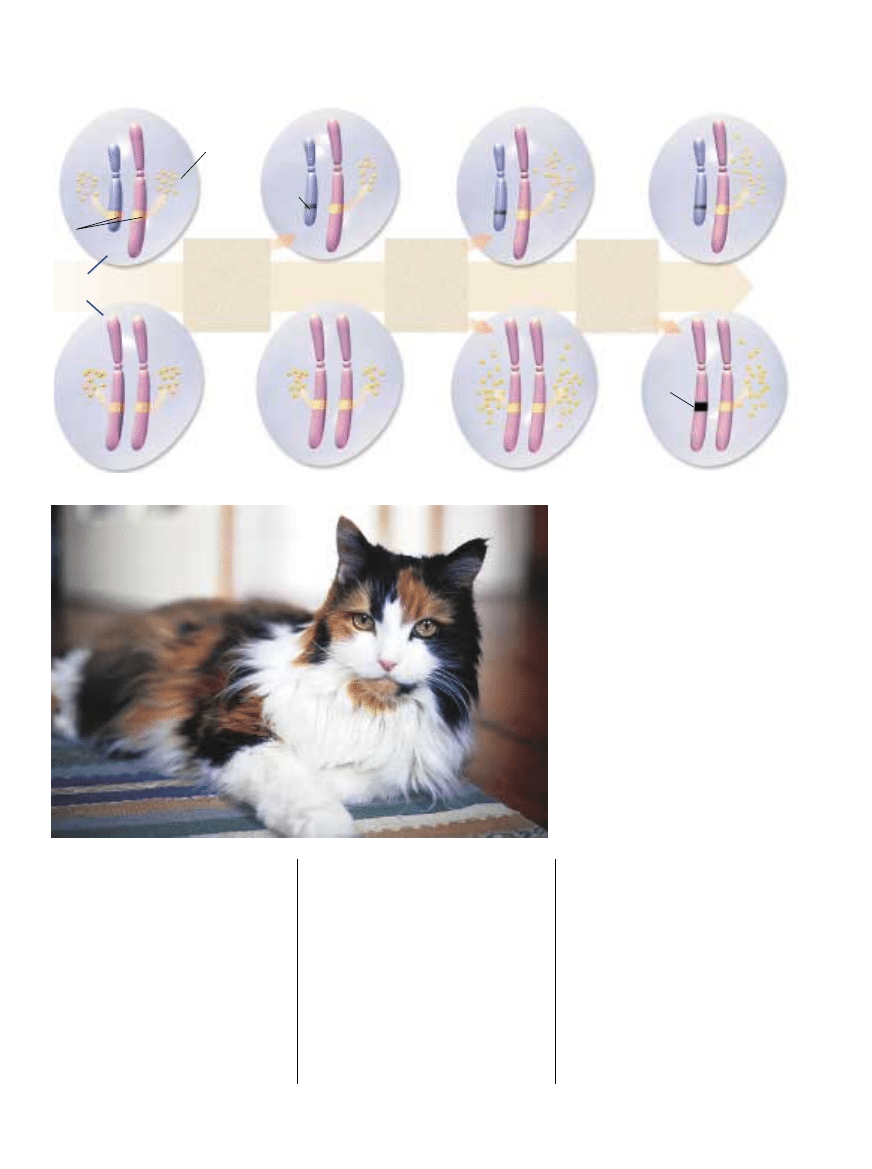
EWOLUCJA INAKTYWACJI X, wy∏àczania
wi´kszoÊci genów na jednym z tych
chromosomów w komórkach samic,
zachodzi∏a niewàtpliwie etapami
– po jednym lub po kilka genów
jednoczeÊnie – by skompensowaç utrat´
genów na chromosomie Y (schemat).
Jeden z efektów inaktywacji X mo˝na
zaobserwowaç u kotów szylkretowych.
Gen okreÊlajàcy, czy barwa sierÊci jest
pomaraƒczowa, czy czarna
(tzn. nie pomaraƒczowa) znajduje si´
w chromosomie X. Futro samic majàcych
pomaraƒczowà wersj´ genu na jednym
chromosomie X, a czarnà na drugim,
b´dzie w czarne i pomaraƒczowe plamy,
zale˝nie od tego, który X i w której komórce
jest wy∏àczony. Inny gen jest odpowiedzialny
za bia∏e fragmenty sierÊci.

cia wystàpi∏a przed 130–80 mln lat, nim
dosz∏o do ró˝nicowania si´ ssaków ∏o-
˝yskowych. Ostatnia inwersja w Y na-
stàpi∏a oko∏o 50–30 mln lat temu, po
wyodr´bnieniu si´ linii ewolucyjnej
ma∏p wàskonosych, ale zanim rozdzie-
li∏y si´ drogi ma∏p cz∏ekokszta∏tnych
i cz∏owieka.
Wbrew ogólnie obowiàzujàcej prawi-
d∏owoÊci dla par X–Y cz´Êç genów w
nierekombinujàcym obszarze drugie-
go z tych chromosomów koduje bia∏ka,
które niewiele ró˝nià si´ od bia∏ek ko-
dowanych przez ich odpowiedniki
na X, nawet w regionach, gdzie naj-
wczeÊniej dosz∏o do inwersji. Wynika
to zapewne z prostego prawa ewolucji:
jeÊli gen jest konieczny do prze˝ycia or-
ganizmu, zazwyczaj zostaje zachowany.
Faktycznie, geny Y, które ma∏o si´ zmie-
ni∏y, sà na ogó∏ tzw. genami metaboli-
zmu podstawowego („housekeeping”
genes) – niezb´dnymi do prawid∏owe-
go funkcjonowania i integralnoÊci nie-
omal wszystkich komórek organizmu.
Nadrabianie strat
Z ZASAD LOGIKI
– i rezultatów wielu ba-
daƒ – wynika, ˝e brak rekombinacji mi´-
dzy Xi Y, a w konsekwencji degenera-
cja wielu genów w tym drugim, musia∏y
wywo∏aç kolejny proces, który by te zjawi-
ska kompensowa∏. Wyglàda to nast´pujà-
co: nie w ka˝dej komórce aktywne sà
wszystkie geny. Ale gdy potrzebne jest jej
dane bia∏ko, na ogó∏ w∏àczona zostaje
zarówno ojcowska, jak i matczyna kopia
odpowiedniego genu. IloÊç bia∏ka wypro-
dukowana dzi´ki ka˝dej kopii jest precy-
zyjnie dopasowana do optymalnego roz-
woju i codziennego funkcjonowania
organizmu. Tak wi´c gdyby geny na Y za-
cz´∏y zanikaç, produkcja kodowanych
przez nie bia∏ek spad∏aby u samców o po-
∏ow´, o ile gatunki, u których ten proces
zachodzi, nie wykszta∏ci∏yby czegoÊ, co
rekompensowa∏oby t´ strat´.
Wiele zwierzàt, na przyk∏ad muszka
owocowa, radzi sobie z tym, podwaja-
jàc aktywnoÊç odpowiedników utra-
conych z Y genów, obecnych na chro-
mosomie X. Pewne organizmy majà
bardziej z∏o˝onà strategi´. Najpierw
dochodzi do zwi´kszenia aktywnoÊci
genów w Xu przedstawicieli obu p∏ci –
w ten sposób uzupe∏niany jest niedo-
bór bia∏ek u samców, ale u samic two-
rzy si´ ich za du˝o, a nast´pnie (np.
w przypadku nicieni) zmniejsza si´ ak-
tywnoÊç genów w Xu samic. U innych,
w tym ssaków, zachodzi proces zwany
inaktywacjà X, podczas którego komór-
ki wczesnych zarodków ˝eƒskich lo-
sowo wy∏àczajà wi´kszoÊç genów w
jednym z chromosomów X. Sàsiednie
komórki mogà wyciszaç ró˝ne kopie
tego chromosomu, ale wszyscy potom-
kowie danej komórki b´dà wykazywa-
li ten sam wzór jego inaktywacji.
Choç jest ona od dawna uwa˝ana za
odpowiedê na degeneracj´ genów w Y,
nie by∏o na to dowodów. JeÊli utrata
funkcji genów w Y powodowa∏a unie-
czynnienie X, to mo˝na przypuszczaç,
˝e geny w Xmajàce funkcjonalne odpo-
wiedniki w nie podlegajàcym rekom-
binacji regionie Y b´dà u samic nadal
dzia∏aç, tak by iloÊç bia∏ek u osobni-
ków obu p∏ci by∏a podobna. Analizu-
WYDANIE SPECJALNE
ÂWIAT NAUKI
101
MARK HARMEL
Stone
OPRÓCZ POZNANIA historii chromosomów p∏ci badania chromosomu Y pomagajà
wyjaÊniç niektóre przypadki bezp∏odnoÊci. Za mniej wi´cej po∏ow´ z ogólnej ich liczby
ca∏kowità lub cz´Êciowà win´ ponosi m´˝czyzna, który produkuje za ma∏o plemników
bàdê nie wytwarza ich wcale. Przyczyny tego stanu cz´sto sà niejasne. Z nowych badaƒ
wynika jednak, ˝e Y zawiera pewnà liczb´ genów p∏odnoÊci i zaburzenia choçby
w jednym sà przyczynà tego typu dolegliwoÊci u oko∏o 10% dotkni´tych nià m´˝czyzn.
Po raz pierwszy na rol´ Y zwrócono uwag´ w latach siedemdziesiàtych, gdy stwierdzono,
˝e u wielu bezp∏odnych m´˝czyzn brakuje ma∏ych odcinków tego chromosomu.
DziÊ wiadomo, ˝e delecje (ubytki) w którymkolwiek z trzech regionów Y mogà powodowaç
bezp∏odnoÊç i ˝e ka˝dy z nich – okreÊlany jako AZF (azoospermia factor – czynnik
azoospermii) a, b i c– zawiera wiele genów.
Wi´kszoÊç z nich jest bardzo aktywna
w jàdrach, gdzie wytwarzane sà
plemniki. (Oznacza to, ˝e powstajà
tam du˝e iloÊci bia∏ek kodowanych
przez te geny). A zatem geny
w regionach AZF sà istotne w produkcji
plemników, choç nie znamy dok∏adnie
ich funkcji ani interakcji z genami
p∏odnoÊci w innych chromosomach.
OkreÊlenie delecji w Y uwa˝ane
jest przez niektórych specjalistów
od bezp∏odnoÊci za element
diagnostyki. JeÊli m´˝czyêni z tego
typu zmianami w ogóle wytwarzajà
plemniki, mo˝na im zaproponowaç
terapi´ zwanà w skrócie ICSI
(intracytoplasmic sperm injection
– wstrzykiwanie plemnika
do cytoplazmy komórki jajowej),
w której uzyskane z jàder m´skie
komórki rozrodcze wprowadza si´
do komórek jajowych w laboratorium.
Niestety, ich synowie odziedziczà
wadliwy chromosom Y i zapewne
b´dà mieli te same problemy
z bezp∏odnoÊcià.
Gdy naukowcy poznajà dok∏adnie rol´ bia∏ek kodowanych przez geny z regionów AZF,
byç mo˝e b´dà w stanie wyleczyç bezp∏odnych m´˝czyzn z delecjami Y, podajàc im
brakujàce bia∏ka, a mo˝e nawet geny. Z kolei takie informacje przyczyni∏yby si´
zapewne do opracowania nowych, zak∏ócajàcych wytwarzanie plemników Êrodków
antykoncepcyjnych dla m´˝czyzn.
DOSTARCZENIE PLEMNIKA (widocznego
w mikroigle) wprost do komórki jajowej to
prawdopodobnie sposób na pokonanie
bezp∏odnoÊci u niektórych m´˝czyzn
majàcych mutacje w chromosomie Y.
TAJEMNICA M¢SKIEJ BEZP¸ODNOÂCI
KARIN JEGALIAN i BRUCE T. LAHN uzy-
skali stopnie doktorskie w laboratorium
genetyka Davida C. Page’a w Whitehead
Institute for Biomedical Research w Cam-
bridge w stanie Massachusetts oraz w Mas-
sachusetts Institute of Technology. Jega-
lian zajmuje si´ obecnie popularyzacjà
nauki w National Institutes of Health.
Lahn, który pochodzi z Chin, jest bada-
czem w Howard Hughes Medical Insti-
tute i profesorem na Wydziale Genetyki
Cz∏owieka University of Chicago.
O
AUTORZE
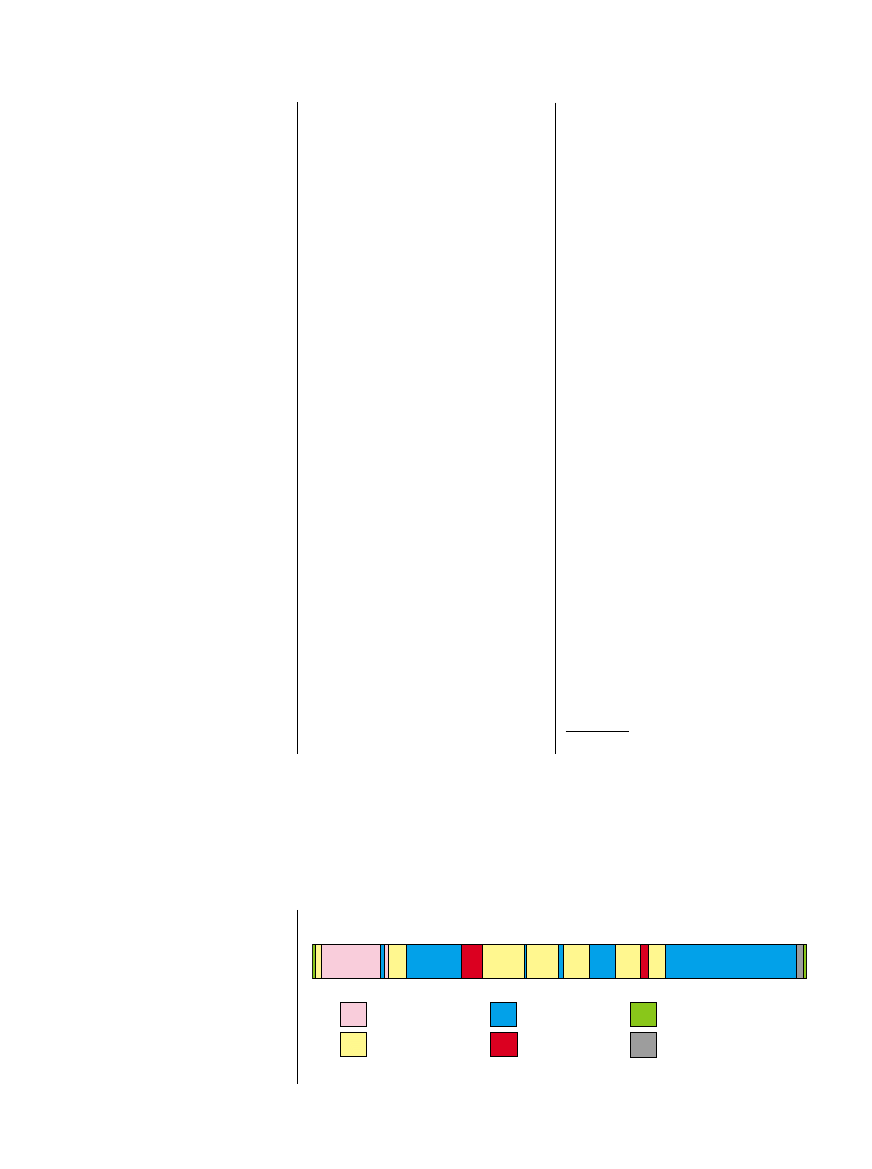
jàc aktywnoÊç utrzymanych par X–Y
u ponad 20 gatunków ssaków, Jegalian
oraz Page stwierdzili kilka lat temu,
˝e kopie funkcjonalnych genów Y w X
unikajà inaktywacji. Badacze ci wyka-
zali tak˝e, ˝e unieczynnienie X, zacho-
dzàce obecnie „na poczekaniu”, nie po-
wsta∏o od razu, ale raczej stopniowo,
odcinek po odcinku lub byç mo˝e gen
po genie.
Nowe motywy
CO DZIWNIEJSZE
, nie podlegajàcy rekom-
binacji region chromosomu Y ma nie tyl-
ko garstk´ cennych genów obecnych
tak˝e w X, ale równie˝ kilkanaÊcie zwià-
zanych z m´skà p∏odnoÊcià, które kodu-
jà bia∏ka wytwarzane wy∏àcznie w jà-
drach (zapewne biorà udzia∏ w produkcji
plemników). Prawdopodobnie niektóre
z nich przeskoczy∏y na Y z innych chro-
mosomów. Pozosta∏e by∏y na tym chro-
mosomie od poczàtku, ale pierwotnie
pe∏ni∏y odmienne funkcje, z up∏ywem
czasu zaÊ naby∏y nowych. Degeneracja
stanowi wi´c tylko jeden z elementów
istotnych w ewolucji chromosomu Y.
Drugi, znany dopiero od niedawna, to
nabycie lub powstanie genów p∏odnoÊci.
Teoretycy nie sà zgodni, jakie si∏y
przekszta∏ci∏y Y w magnes przyciàga-
jàcy takie geny. Byç mo˝e obecnoÊç wy-
∏àcznie u samców genów szkodliwych
lub oboj´tnych dla samic jest dla ga-
tunku korzystna. Niewykluczone, ˝e
Y ochrania m´skie geny p∏odnoÊci, gwa-
rantujàc ich przechodzenie bezpoÊred-
nio od samca do samca, z pomini´ciem
samicy (która mog∏aby je zgubiç bez
szkody dla siebie).
Inna zagadka: w jaki sposób geny
p∏odnoÊci sà w stanie istnieç, jeÊli nie
zachodzi rekombinacja, czyli w warun-
kach, które spowodowa∏y degeneracj´
wi´kszoÊci genów w Y. Prawdopodob-
nie przyczynà tego jest fakt, ˝e niemal
ka˝dy gen m´skiej p∏odnoÊci obecny
w Y ma wiele kopii. Mo˝e to znosiç
efekty szkodliwych mutacji, które w da-
nym momencie zachodzà tylko w jednej
z nich. Gdy wskutek nagromadzenia
mutacji pewne kopie przestajà pe∏niç
swoje funkcje, pozosta∏e podtrzymujà
zdolnoÊç samca do rozmna˝ania i mo-
gà si´ same namna˝aç.
Najdok∏adniej badano ewolucj´ ludz-
kich chromosomów p∏ci. Porównujàc
uzyskane informacje z wynikami ba-
daƒ innych gatunków, okreÊlono ogól-
ne zasady determinacji p∏ci, dzia∏ajà-
ce nie tylko u ssaków. U niektórych
zwierzàt, na przyk∏ad ptaków i motyli,
funkcjonuje system determinacji p∏ci
W–Z. Gdy dziedziczenie jednej kopii
specyficznego chromosomu jest wy-
znacznikiem p∏ci m´skiej, chromosom
jest okreÊlany jako Y, a jego partner ja-
ko X. Natomiast jeÊli dziedziczenie
jednej kopii takiego chromosomu wa-
runkuje rozwój samicy, chromosom na-
zywa si´ W, a jego partner w parze – Z.
Godne uwagi jest to, ˝e chromoso-
my p∏ci pochodzà od autosomów. Jed-
nak autosomy te mogà byç ró˝ne. Pta-
sie chromosomy W i Z powsta∏y na
przyk∏ad z innych chromosomów ni˝
X i Y ssaków oraz muszek owocowych.
W przypadku wi´kszoÊci rozmna˝a-
jàcych si´ p∏ciowo gatunków, u których
powsta∏y chromosomy p∏ci, stawa∏y si´
one coraz mniej podobne do siebie,
przechodzàc jeden lub wi´cej cykli z∏o-
˝onych z trzech kolejnych etapów:
wstrzymania rekombinacji, degeneracji
nierekombinujàcych cz´Êci chromoso-
mu specyficznego dla p∏ci (Y lub W)
oraz kompensacji utraconych genów
przez drugi chromosom. W tym czasie
chromosom specyficzny dla p∏ci móg∏
staç si´ wa˝ny dla p∏odnoÊci organi-
zmu, tak jak Y u ludzi i owadów.
Nale˝y zastanowiç si´ nad przysz∏o-
Êcià naszego gatunku. Czy cykl ten
b´dzie trwaç dalej, a˝ ca∏kowicie unie-
mo˝liwi rekombinacj´ mi´dzy chromo-
somami p∏ci i ostatecznie zniszczy
Y za tysiàce czy miliony lat? Z nowych
odkryç wynika, ˝e samce potrafià chro-
niç te geny z Y, które sà wa˝ne do prze-
˝ycia i rozmna˝ania. Niemniej ca∏ko-
wita degeneracja Y jest teoretycznie
mo˝liwa.
Cz´sto podejmuje si´ badania genów,
by zrozumieç i leczyç choroby. Chromo-
somem Y zaj´to si´ w∏aÊnie z ch´ci zro-
zumienia rozwoju samców i leczenia
bezp∏odnoÊci. Jednak wiele badaƒ nie
koncentrowa∏o si´ na terapii. W miar´
analizy coraz wi´kszej liczby genów
w Xi Y, identyfikowanych za pomocà
testów medycznych i systematycznego
sekwencjonowania, naukowcy zajmujà-
cy si´ ewolucjà zadali podstawowe pyta-
nie – czy geny te powiedzà coÊ nowego
o odleg∏ej przesz∏oÊci dziwnie niedobra-
nej pary chromosomów Xi Y? Opo-
wieÊç okaza∏a si´ bardzo ciekawa.
n
* Rodney „No Respect” Dangerfield – komik ame-
rykaƒski o przydomku „Bez Szacunku” (przyp. red.).
102
ÂWIAT NAUKI
EWOLUCJA CZ¸OWIEKA /NOWE SPOJRZENIE
GDY W LUTYM 2001 ROKU ukaza∏ si´
numer Nature w ca∏oÊci poÊwi´cony
sekwencji genomu ludzkiego, zaledwie
2.5 z jego 216 stron dotyczy∏y chromoso-
mu Y. Autor tego krótkiego artyku∏u Da-
vid C. Page, u którego doktoryzowali si´
Karin Jagelian i Bruce T. Lahn, opisywa∏
przygotowanie mapy chromosomu Y, za-
dania wyjàtkowo trudnego ze wzgl´du
na liczne powtarzajàce si´ sekwencje
obecne w jego DNA. Jeszcze wówczas
STRUKTURA chromosomu Y.
K O M E N TA R Z
A jednak Y si´ trzyma!
X-transponowane
X-zdegenerowane
Amplikonowe
Heterochromatyczne
Pseudoautosomalne
Inne
Wyszukiwarka
Podobne podstrony:
Dlaczego entuzjazm jest taki ważny
Jest taki smutek
JEST TAKI DZIEŃ
Jest taki kwiat malinski
Jest taki dzień, TEKSTY PIOSENEK
JEST TAKI DZIEŃ
Wigilia - JEST TAKI JEDEN DZIEŃ W ROKU, Inscenizacje
Borysewicz, Kukiz Jest taki dzień
1 dlaczego jest dzień i noc, Edukacja społeczno - przyrodnicza
STRES NIE JEST TAKI ZŁY
Jest taki dzień?rdzo ciepły poezja
Dlaczego jest tak wiele nieudanych małżeństw, małżeństwo
Dlaczego jest święto Bożego Narodzenia, Szkoła
Jest taki obraz - Mars, Kultura-sztuka, Jest taki obraz
Jest taki Urząd Etnografia organizacji służby publicznej
więcej podobnych podstron