
FOTOSYNTEZA
I
CHEMOSYNTEZA

METABOLIZM
• To całokształt reakcji chemicznych i związanych
z nimi przemian energii zachodzących w
żywych komórkach, stanowiący podstawę
wszelkich zjawisk biologicznych.
• Procesy te pozwalają komórce na wzrost i
reprodukcję, zarządzanie swoją strukturą
wewnętrzną oraz odpowiadanie na bodźce
zewnętrzne.
• Wyróżniamy procesy:
• kataboliczne
• anaboliczne

KATABOLIZM
• To ogół reakcji chemicznych metabolizmu
prowadzący do rozpadu złożonych związków
chemicznych na prostsze cząsteczki.
• Procesy anaboliczne to m.in.:
• oddychanie tlenowe
• oddychanie beztlenowe (zamiast tlenu mogą
być wykorzystane związki nieorganiczne,
takie jak azotany lub siarczany)
• fermentacja

ANABOLIZM
• To grupa reakcji chemicznych, w wyniku których
z prostych substratów powstają związki złożone,
gromadzące energię. Jest to ta część
metabolizmu, która związana jest ze wzrostem
tkanek organizmu.
• Anabolizm to reakcje syntez związków bardziej
złożonych z prostszych, wymagające
dostarczenia energii.
• Powstający w ten sposób produkt rekcji zawiera
większą ilość energii, niż substraty.
• Dostarczona energia zostaje zmagazynowana w
postaci wiązań chemicznych.

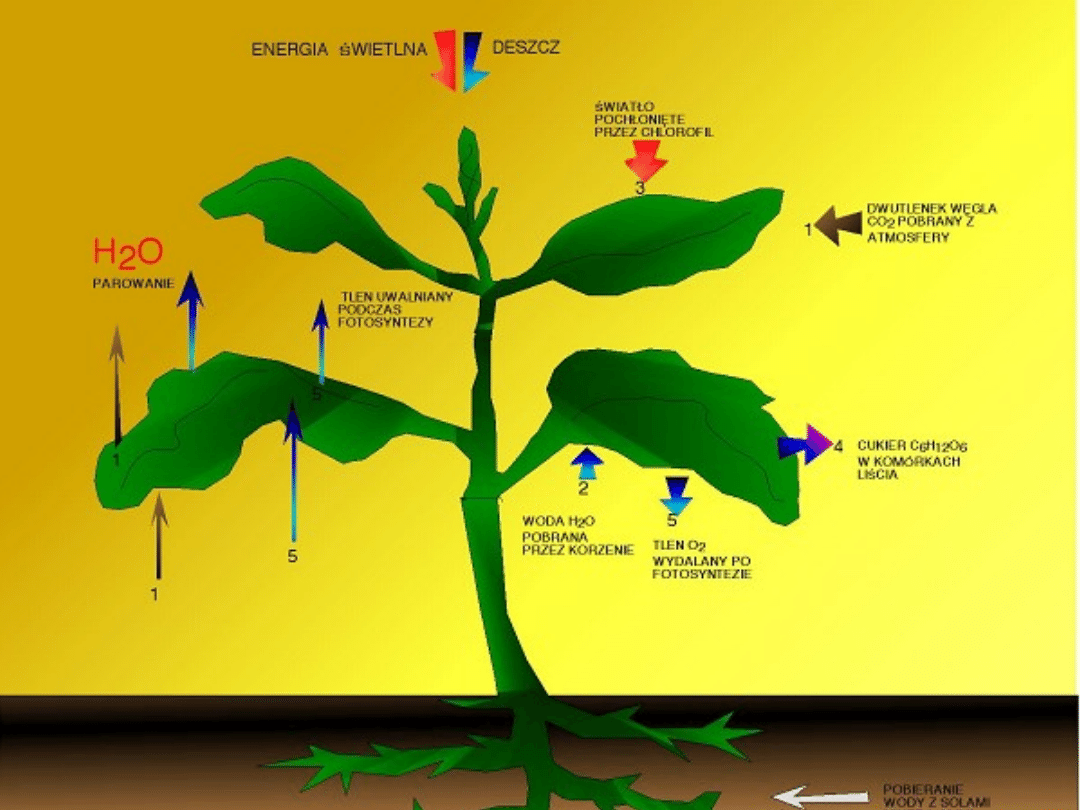
FOTOSYNTEZA
•
Fotosynteza - proces anaboliczny, w trakcie którego z
prostych substancji nieorganicznych z udziałem energii
świetlnej powstają związki organiczne.
•
W formie sumarycznej przebieg fotosyntezy można zapisać
następująco:
6H
2
O + 6CO
2
+ energia świetlna + chlorofil -> C
6
H
12
O
6
+ 6O
2
•
Organizmy produkujące związki organiczne na drodze
fotosyntezy to:
•
prawie wszystkie rośliny (nieliczne wyjątki to rośliny
cudzożywne, saprofityczne i pasożytnicze)
•
niektóre Protista,
•
bakterie purpurowe oraz sinice

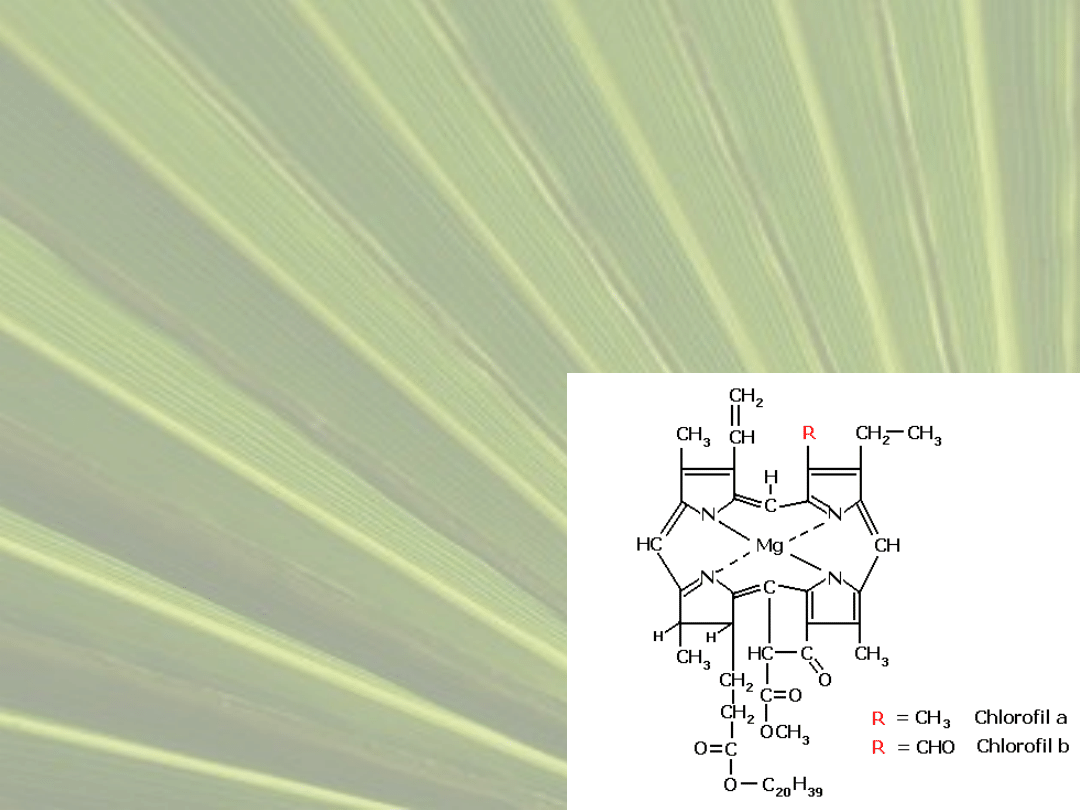
CHLOROFIL
• Barwnik którego zadaniem jest generowanie pod
wpływem światła widzialnego wolnych elektronów,
które są następnie spożytkowane w dalszych
etapach fotosyntezy.
• Wyróżniania się następujące typy chlorofilu
• chlorofil a - występuje powszechnie w
roślinach
i sinicach,
• chlorofil b - rośliny wyższe, zielenice, eugleny
• chlorofil c - okrzemki, brunatnice
• chlorofil d – krasnorosty
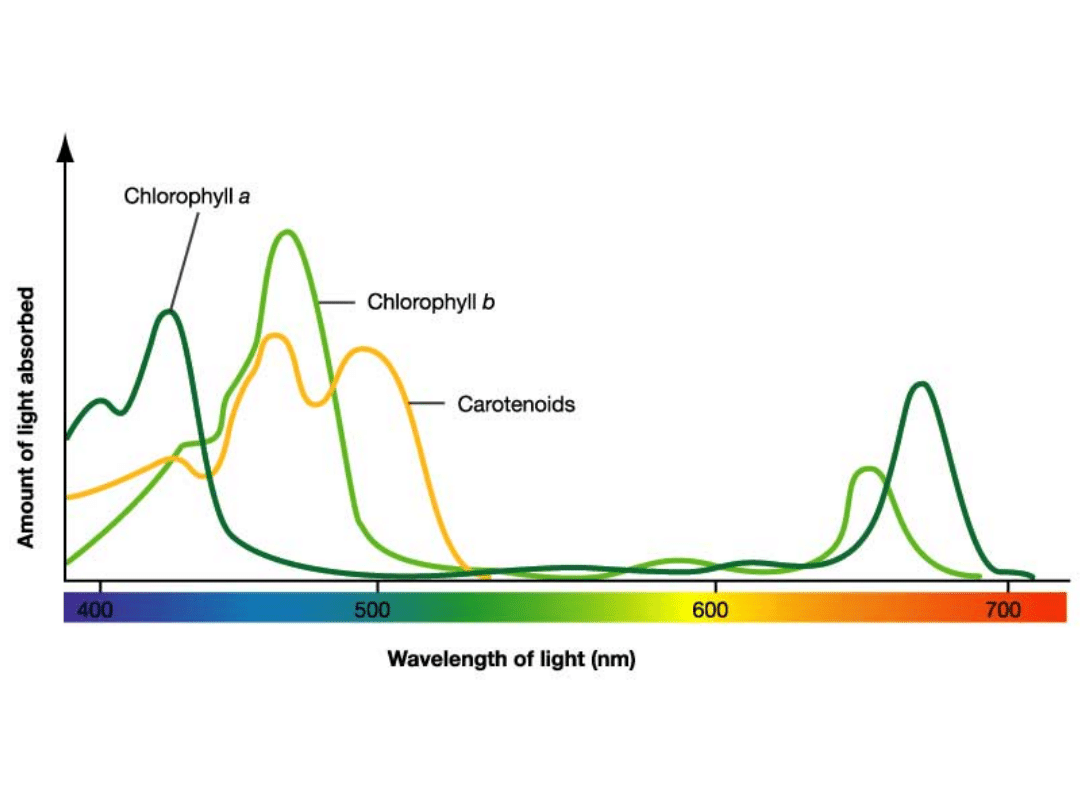
• Stosunki ilościowe chlorofili w roślinach zależą
między innymi od warunków siedliskowych:
rośliny cieniolubne mają więcej chlorofilu b,
światłolubne chlorofilu a.
• Barwniki fotosyntetyczne to, oprócz chlorofili,
również karotenoidy i fikobiliny.
• Chlorofil to chemicznie
kompleks jonu
magnezowego
i pochodnej porfiryny, z
przyłączonym fitolem.

• Fotosynteza przebiega w błonie tylakoidów i stromie
chloroplastów i dzieli się na dwie fazy: jasną, ciemną
• Reakcje fazy świetlnej energię świetlną i wodę
wykorzystują do wytworzenia tzw. siły
asymilacyjnej, to znaczy ATP i NADPH.
• Reakcje fazy ciemnej zużywają z kolei siłę
asymilacyjną w procesie syntezy cukru z dwutlenku
węgla.
• U roślin zielonych i sinic wyróżniamy dwa rodzaje
fotosystemu, określane jako :
• fotosystem I ( PS I) max700nm
• fotosystem II ( PS II) max 680nm.

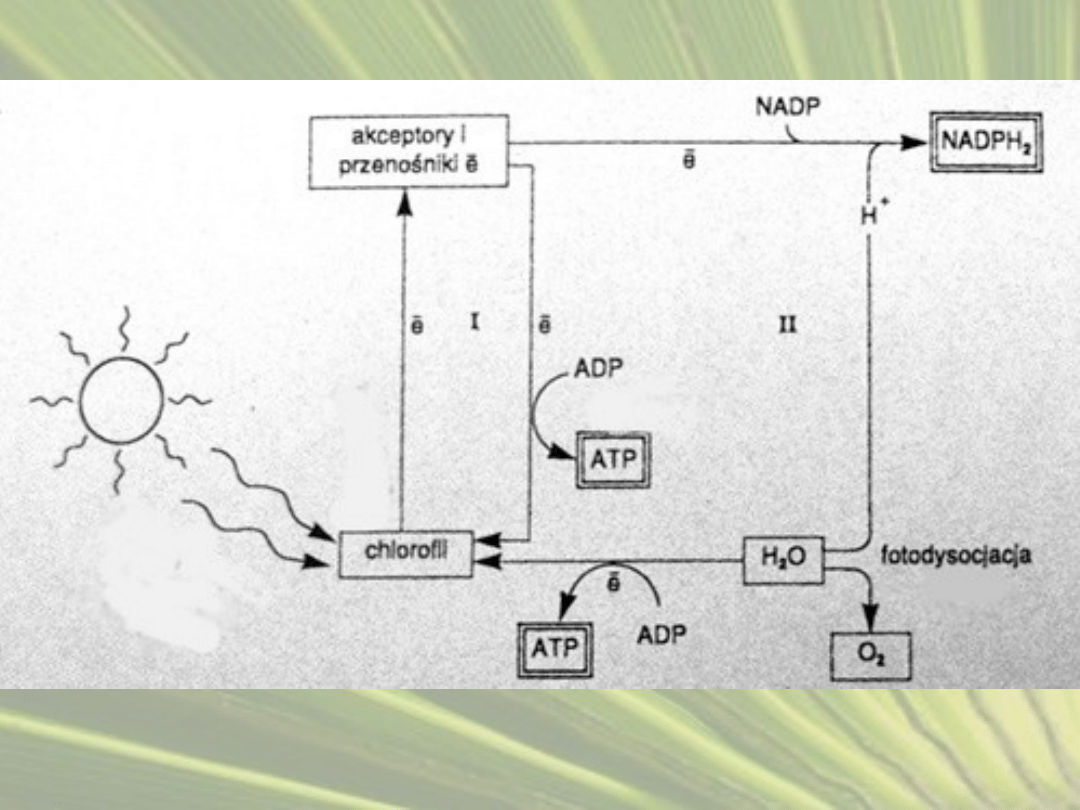
1.
FAZA JASNA
1) Cząsteczka chlorofilu w kompleksie PS II pochłania
światło, a energia jest kierowana do centrum
reakcji.
2) PS II odłącza elektron o wysokiej energii, który
trafia na lipid o nazwie plastochinon
3) Z kolei kation P680
+
odbiera elektron cząsteczce
wody i odzyskuje aktywność.
wzór fotolizy wody : H
2
O → 2e
-
+ 2H
+
+ ½O
2
)
4) Uwolniony tlen trafia do atmosfery, a wodór
gromadzi się w zbiornikach tylakoidów.

5) Kompleks cytochromów b-f przenosi elektron
dalej na białko plastocyjaninę, zawierające
miedź.
6) Światło wzbudza również fotosystem I, którego
energia również wybija elektron z cząsteczki
7) Przekazuje go na białko ferredoksynę.
8) Aktywność przywraca mu przyjęcie elektronu z
plastocyjaniny, dostarczonego przez PS II.
9) Cząsteczki zredukowanej ferredoksyny
przekazują elektron koenzymowi NADP
+
, który
przechodzi w formę NADPH
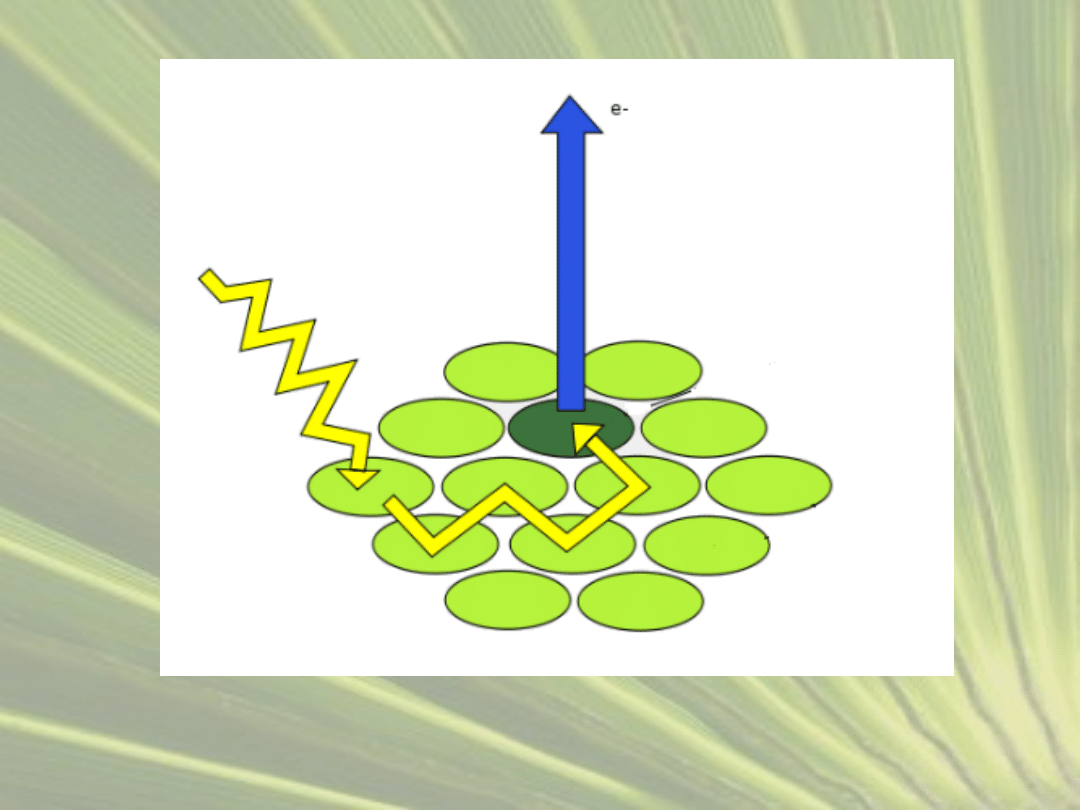

ŚWIATŁO
PS II
FERREDOKSYNA
PLASTOCHINON
NADP
+
NADPH
PLASTOCYJANINA
CYTOCHROMY B-F
ŚWIATŁO
PS I
FOTOLIZA WODY
H
2
O → 2e
-
+ 2H
+
+ ½O
2

ŚWIATŁO
PS II
FERREDOKSYNA
PLASTOCHINON
NADP
+
NADPH
PLASTOCYJANINA
CYTOCHROMY B-F
ŚWIATŁO
PS I
FOTOLIZA WODY
H
2
O → 2e
-
+ 2H
+
+ ½O
2
ATP
ATP

1.
FAZA CIEMNA
• Energia zgromadzona w ATP i NADPH
wykorzystywana jest do związania CO
2
i
wytworzenia prostych cukrów.
• Ze względu na ilość węgli w pierwszym
produkcie asymilacji CO
2
wyróżniamy dwa
typy roślin :
• C3 (pierwszy związek 3 węglowy)
• C4 (pierwszy związek 4 węglowy)

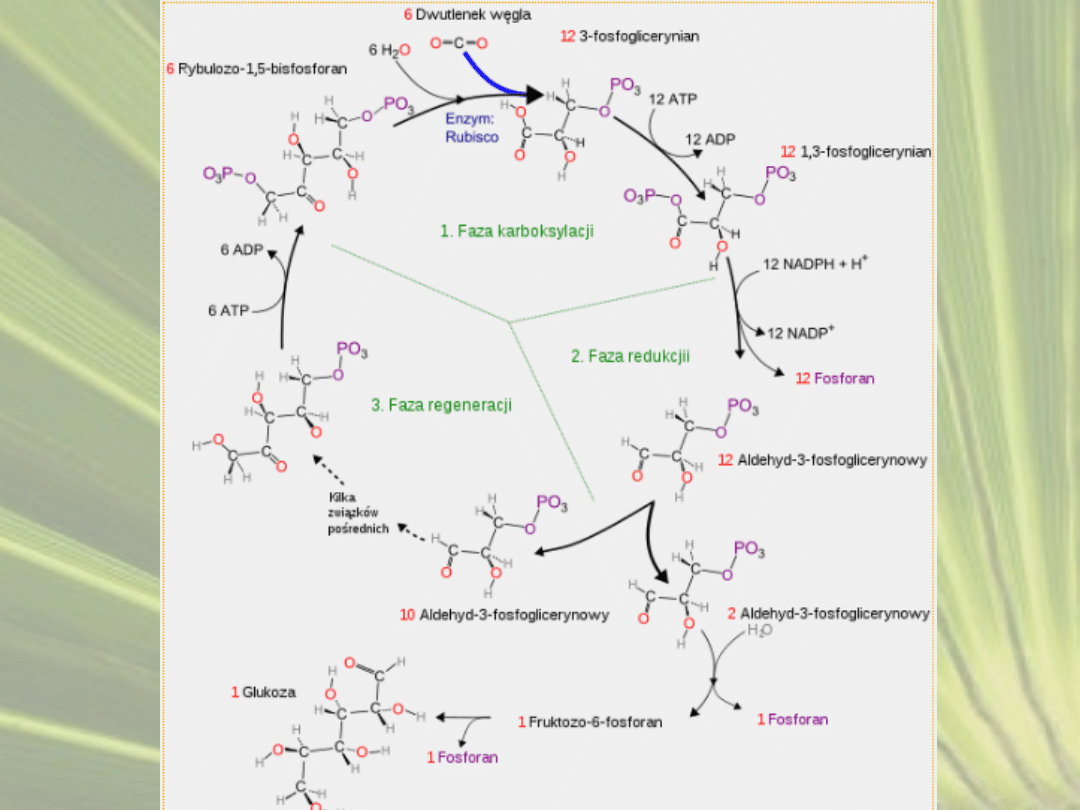
CYKL ROŚLIN C3
Faza karboksylacji
• Dwutlenek węgla przyłączany jest do 1,5-
bisfosforybulozy. Enzymem katalizującym przyłączenie
cząsteczki CO
2
jest karboksylaza 1,5-bisfosforybulozy
• W wyniku przyłączenia cząsteczki CO
2
do 1,5-
bisfosforybulozy powstaje nietrwały związek
sześciowęglowy - 1,5-bisfosfo-2-karboksy-3-ketoarabitol,
• Niemal natychmiast rozpada się na dwie cząsteczki
kwasu 3-fosfoglicerynowego.
Faza redukcji
• Kwas 3-fosfoglicerynowy jest fosforylowany do kwasu
1,3-bisfosfoglicerynowego.
• Redukcja kwasu 1,3-bisfosfoglicerynowego do aldehydu
3-fosfoglicerynowego.

Faza regeneracji
• Z aldehydu 3-fosfoglicerynowego odtwarzany jest
akceptor CO
2
1,5-bisfosforybuloza.
• Po związaniu 6 cząsteczek CO
2
z cyklu może zostać
wyprowadzona 1 cząsteczka heksozy.
• Reakcje te zachodzą w stromie chloroplastów.
• Istotą fazy ciemnej jest przyłączenie dwutlenku
węgla do prostych związków organicznych, przy tym
ten etap fotosyntezy nie wymaga światła.W efekcie
powstaje cukier (glukoza) oraz tlen.

• Węglowodany gromadzone są w organizmach
roślin głównie w postaci skrobi i sacharozy.
• Cząsteczki skrobi mogą być syntetyzowane
bezpośrednio w chloroplastach z glukozy,
która ulega aktywacji poprzez przyłączenie
cząsteczki ATP, co prowadzi do powstania ADP-
glukozy.
• Sacharoza wytwarzana jest w cytozolu. Do
syntezy cząsteczki sacharozy wykorzystywany
jest fruktozo-6-fosforan i UDP-glukoza.

FOTOODDYCHANIE
• Jest to proces biochemiczny zachodzący na świetle w komórkach
roślinnych, objawiający się pobieraniem tlenu i wydzielaniem
dwutlenku węgla.
• Biochemicznie proces fotooddychania związany jest z
dwufunkcyjnością enzymu karboksylazy/oksygenazy rybulozo-
1-5-bisfosforanu (RuBisCO), odpowiedzialnego zarówno za
przyłączenie do rybulozo-1,5-bisfoforanu (RuBP) cząsteczki CO
2
,
jak i cząsteczki O
2
w chloroplastach podczas oświetlania.
• CO
2
i O
2
konkurują o miejsce katalityczne Rubisco.
• W wyniku przyłączenia tlenu do rybulozo-1-5-bisfosforanu
powstaje jedna cząsteczka kwasu fosfoglicerynowego (jak w fazie
ciemne fotosyntezy) oraz jedna cząsteczka fosfoglikolanu,
pierwszego (dwuwęglowego C2) produktu fotooddychania.
• Stąd pochodzi inna nazwa fotooddychania – cykl C2.

• Powstający w chloroplastach fosfoglikolan ulega
defosforylacji i przenoszony jest do peroksysomów.
• Tam przekształcany jest do glicyny.
• Glicyna transportowana jest do mitochondriów i
przekształcana do seryny z wydzieleniem cząsteczki CO
2
,
NH
3
, oraz NADH.
• Powstała w mitochondriach seryna transportowana jest do
peroksysomów i przekształcana do kwasu glicerynowego
• Produkt reakcji przenoszony jest do chloroplastów i może
służyć do odtworzenia cząsteczki rybulozo-1-5-
bisfosforanu.

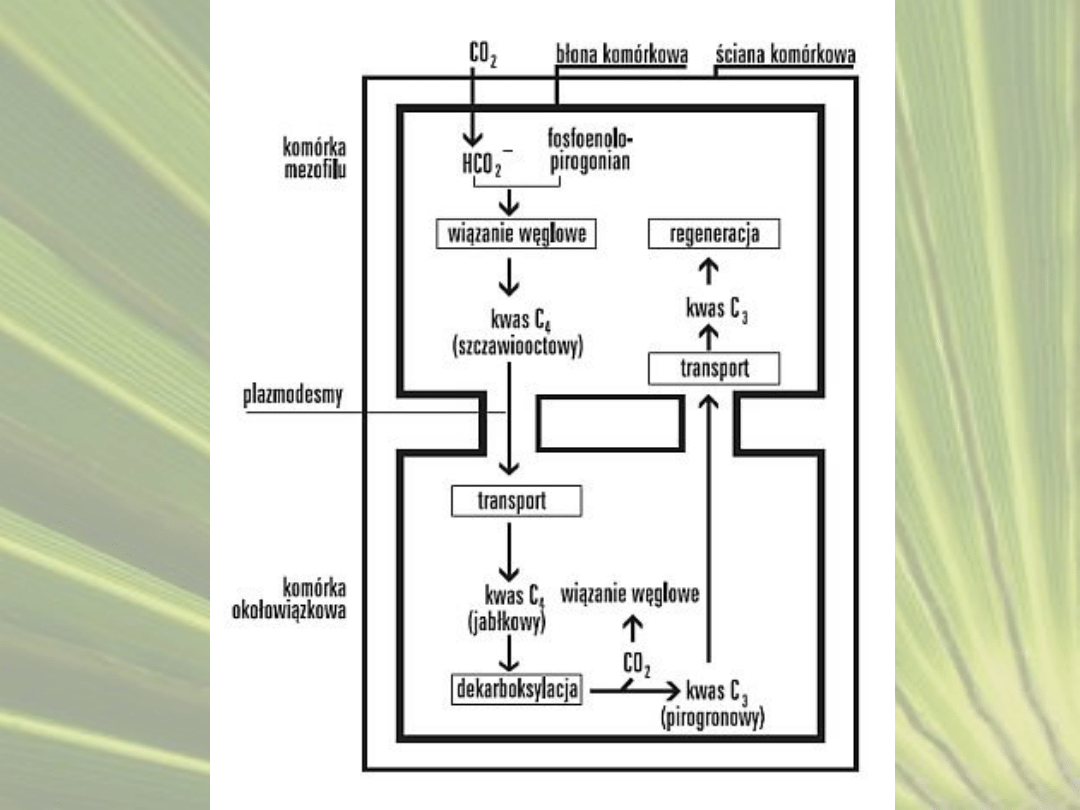
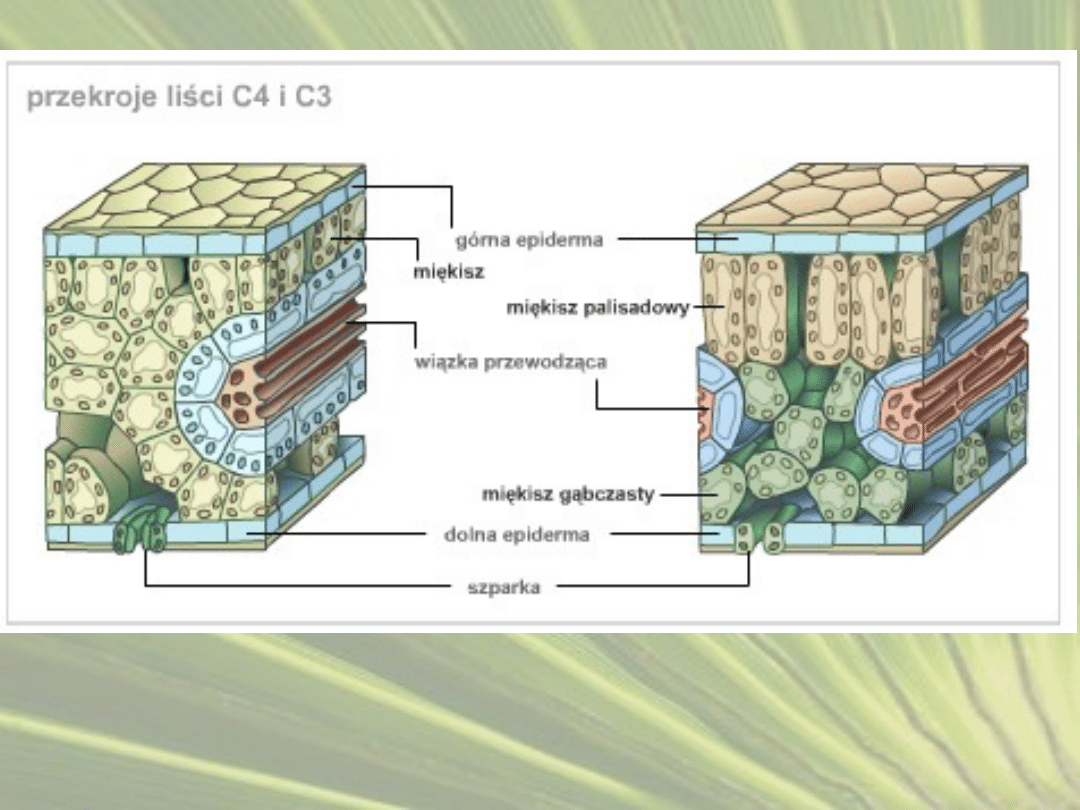
CYKL ROŚLIN C4
• Fotosynteza C
4
, cykl Hatcha-Slacka,– rodzaj fotosyntezy, w
której dodatkowy mechanizm wiązania dwutlenku węgla
poprzedza cykl cykl Calvina.
• Rośliny posiadające zdolność wiązania CO
2
do
fosfoenolopirogronianu określane są nazwą rośliny typu C
4
.
• Pierwszym trwałym produktem wiązania jest związek o
czterech atomach węgla – szczawiooctan
• Rośliny te wykształciły mechanizmy anatomiczne i
fizjologiczne pozwalające na zwiększenie stężenia CO
2
w
komórkach, w których zachodzi cykl Calvina
• W efekcie nie obserwuje się zachodzenia u tych roślin
fotooddychania, które jest przyczyna strat energii u roślin C
3
.
• Rośliny C
4
, pomimo konieczności zużycia dodatkowej energii w
postaci ATP, cechują się większą wydajnością fotosyntezy i
szybszą produkcją biomasy.

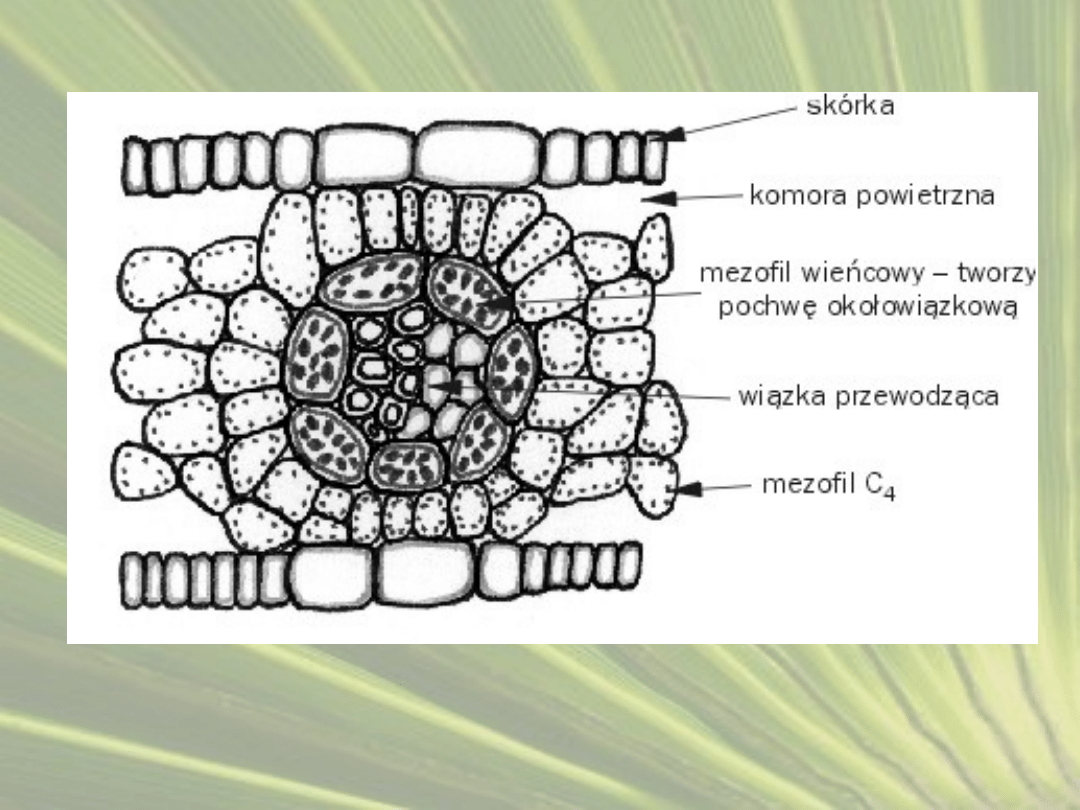
•
Przystosowania anatomiczne polegają na zróżnicowaniu komórek
zaangażowanych w wiązanie CO
2
na komórki mezofilu oraz
komórki pochew okołowiązkowych.
•
Komórki pochew okołowiązkowych posiadają grubą ścianę
komórkową, zwykle wysyconą suberyną, dzięki czemu ściana
komórkowa jest w bardzo małym stopniu przepuszczalna dla
gazów.
•
Proces wiązania CO
2
zachodzi dwukrotnie.
•
Po wniknięciu do komórek mezofilu przez aparaty szparkowe,
dwutlenek węgla przyłączany jest do
fosfoenolopirogronianu. W reakcji tej powstaje związek
czterowęglowy – szczawiooctan. Jest on w zależności od gatunku
rośliny przekształcany do asparaginianu lub jabłczanu i w tej
postaci przenoszony do komórek pochew okołowiązkowych.
•
Tam zachodzi reakcja dekarboksylacji i wydzielenie CO
2
, który
jest włączany do cyklu Calvina. Cykl ten zachodzi tylko w
komórkach pochew okołowiązkowych, gdzie stężenie CO
2
przekracza 10-20 razy stężenie CO
2
w komórkach mezofilu.

• Brak cyklu Calvina w komórkach mezofilowych związany jest z
brakiem enzymu, przyłączającego CO
2
do cząsteczki rybulozo-
1,5-bisfosforanu (RuBP) określanego nazwą karboksylaza
oksygenaza rybulozo-1,5-bisfosforanu (RuBisCO).
• Nie rozstrzygnięto ostatecznie, czy u roślin C
4
zachodzi
fotooddychanie. Wykształcony w czasie ewolucji mechanizm
koncentracji CO
2
niewątpliwie w dużym stopniu ogranicza
reakcję oksygenacji RuBP. Jednak wiele obserwacji wskazuje, że
ten proces u roślin C
4
zachodzi, chociaż w niewielkim stopniu.
• Rośliny o fotosyntezie C
4
szczególnie często występują w
ekosystemach trawiastych i sawannach klimatu gorącego.
Fotosynteza C
4
występuje jedynie u roślin okrytozalążkowych.
Szczególnie często szlak metaboliczny występuje u traw i turzyc
• Obecnie metabolizm C
4
występuje u ok. 1% gatunków roślin.

CYKL CAM
• Jest to szczególny typ fotosyntezy zachodzącej u roślin
określanych jako rośliny CAM
• Do grupy tej należą rośliny, które
ze względu na klimat muszą
prowadzić szczególnie oszczędną
gospodarkę wodną, m.in.
gruboszowate.
• Większość roślin reprezentujących
metabolizm CAM to epifity (np.
storczykowate) lub sukulenty
(np. kaktusy),
• Portulacaria afra jest jedyną rośliną,
która wykazywać zarówno fotosyntezę
CAM, jak i fotosyntezę C4.

• Fotosynteza CAM wiąże się z dwoma etapami wiązania CO
2
:
Noc
• W nocy do fosfoenolopirogronianu (PEP) przyłączany jest
CO
2
. W reakcji tej powstaje szczawiooctan, który następnie
redukowany jest do jabłczanu i w tej postaci CO
2
niezbędny
do przeprowadzania fotosyntezy transportowany jest do
wakuoli i magazynowany.
Dzień
• W dzień jabłczan z wakuoli transportowany jest do
cytozolu, gdzie zachodzi reakcja dekarboksylacji
przeprowadzana przez enzym jabłczanowy.
• Wydzielony CO
2
pokrywa zapotrzebowanie cyklu Calvina w
okresie kiedy intensywnie zachodzi faza jasna fotosyntezy,
a pobieranie CO
2
jest ograniczone w wyniku zamknięcia
aparatów szparkowych.

Faza I
• Noc, w okresie której aparaty szparkowe pozostają otwarte.
• CO
2
atmosferyczny wnika do komórek, gdzie jest wiązany
poprzez przyłączenie do fosfoenolopirogronianu (PEP) przez
enzym karboksylazę fosfoenolopirogronianu (PEPC).
• Fosfoenolopirogronian potrzebny do związania CO
2
wytwarzany jest w procesie glikolizy z cukrów zapasowych i
cukrów rozpuszczalnych powstałych w dniu poprzednim.
• Powstały po przyłączeniu cząsteczki dwutlenku węgla
szczawiooctan (OAA) redukowany jest do jabłczanu,
transportowanego do wakuoli.
• Jabłczan pozostaje w wakuoli aż do wschodu słońca.

Faza II
• Krótki okres początku dnia kiedy aparatu szparkowe
pozostają otwarte a CO
2
wnika.
• Cykl Calvina zachodzi z niewielkim natężeniem.
• Na świetle enzym RuBisCO ulega aktywacji, a
karboksylaza PEP jest dezaktywowana poprzez
defosforylację.
• Jabłczan przestaje być wytwarzany, a zwiększa się
szybkość reakcji cyklu Calvina.

Faza III
• Większą część dnia aparaty szparkowe pozostają zamknięte w
celu ograniczenia utraty wody.
• Cykl Calvina zachodzi dzięki wydzielaniu CO
2
dzięki
dekarboksylacji jabłczanu uwalnianego z wakuoli.
• Proces dekarboksylacji jabłczanu może być przeprowadzany
przez jeden lub kilka enzymów dekarboksylujących: enzym
jabłczanowy zależny od NADP (NADP-ME), enzym jabłczanowy
zależny od NAD (NAD-ME) lub karboksykinazę PEP (PEP-CK).
• Podczas dekarboksylacji wytwarzany jest CO
2
oraz PEP lub
pirogronian.
• CO
2
wykorzystywany jest do wytwarzania węglowodanów, a
związki trójwęglowe przekształcane są powtórnie w cukrów w
procesie glukoneogenezy

Faza IV
• Krótki okres końca dnia kiedy aparaty szparkowe otwierają się i
powtórnie CO
2
jest bezpośrednio włączany w cykl Calvina. W tym
okresie jabłczan z wakuoli jest już wyczerpany.
• Faza II i IV są szczególnie wrażliwe na czynniki środowiskowe.
Dobrze widoczne gdy roślina ma dostęp do odpowiedniej ilości
wody. I nie występują, w okresie suszy, gdy rośliny CAM
maksymalnie oszczędzają wodę.
• W przypadku poważnego ograniczenia ilości wody aparaty
szparkowe roślin CAM pozostają zamknięte zarówno w dzień jak i w
nocy a faza ciemna fotosyntezy zachodzi jedynie dzięki
wewnętrznemu obiegowi CO
2
wytwarzanego w procesie oddychania.
• Także w nocy przy zamkniętych aparatach szparkowych CO
2
pochodzący z oddychania jest przyłączany do PEP i wytwarzane
niewielkie ilości jabłczanu.

CHEMOSYNTEZA
• Proces starszy ewolucyjnie od fotosyntezy i
mniej od niej skomplikowany.
• Przeprowadzają go organizmy nazywane
chemoautotrofami - wyłącznie bakterie.
• Źrodłem enegii do asymilacji dwutlenku węgla
są reakcje utlenienia prostszych związków:
• nieorganicznych (chemolitotrofy)
• organicznych (chemoorganotrofy)

•
Chemosynteza pełni bardzo ważną rolę w obiegach
pierwiastków.
•
Chemosyntezę można podzielić na dwa etapy:
1.
utlenianie związku chemicznego
związek mineralny + O
2
→ związek mineralny +
energia
2.
związanie CO
2
i produkcja glukozy
CO
2
+ H
2
O + energia → związek organiczny + O2
•
Bakterie chemosyntetyzujące.

1. BAKTERIE NITRYFIKACYJNE
• bakterie z rodzaju Nitrosomonas -
wykorzystują utlenianie amoniaku do
azotynów
2NH
3
+ 3O
2
→ 2HNO
2
+ 2H
2
O + ENERGIA
• bakterie z rodzaju Nitrobacter -
wykorzytują utlenianie azotynów do
azotanów
2HNO
2
+ O
2
--> 2HNO
3
+ ENERGIA

2. BAKTERIE SIARKOWE
• utleniają siarkowodór do czystej siarki
2H
2
S + O
2
--> 2H
2
O + 2S + ENERGIA
• utleniają czystą siarke do kwasu
siarkowego(VI)
2S + 2H
2
O + 3O
2
--> 2H
2
SO
4
+ ENERGIA
3.
BAKTERIE WODOROWE
•
utleniają wodór do wody
2H
2
+ O
2
--> 2H
2
O + ENERGIA

4. BAKTERIE ŻELAZOWE
•
(utleniają sole żelaza(II) do soli żelaza(III)
2Fe(HCO
3
)
2
+ 1/2O
2
+ H
2
O -->2Fe(OH)
3
+4CO
2
+
ENERGIA
5. BAKTERIE METANOWE
• bakterie utleniające metan do dwutlenku węgla:
CH
4
+ 2O
2
--> CO
2
+ 2H
2
O + ENERGIA
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
- Slide 38
- Slide 39
- Slide 40
Wyszukiwarka
Podobne podstrony:
Fotosynteza i chemosynteza.Zadania maturalne, BIOLOGIA(1)
Fotosynteza i chemosynteza po Nieznany
fotosynteza i chemosynteza
Fotosynteza fizjolog 05
podrecznik 2 18 03 05
regul praw stan wyjątk 05
05 Badanie diagnostyczneid 5649 ppt
Podstawy zarządzania wykład rozdział 05
05 Odwzorowanie podstawowych obiektów rysunkowych
05 Instrukcje warunkoweid 5533 ppt
05 K5Z7
05 GEOLOGIA jezior iatr morza
05 IG 4id 5703 ppt
05 xml domid 5979 ppt
Świecie 14 05 2005
Wykł 05 Ruch drgający
więcej podobnych podstron