
1
Wapń
BIOCHEMIA
WYKŁAD 14

2
- Jest pierwiastkiem występującym w największym
stężeniu w organizmie człowieka:
99% zawarte jest w kośćcu
0,01% w osoczu
pozostały w przestrzeni międzykomórkowej i
komórkowej
- Dzienne zapotrzebowanie (z diety) dla dorosłego
człowieka – ok. 25 mmol/dobę
- Występuje w osoczu w 3 formach:
a) wolny (zjonizowany, jedyna aktywna
biologicznie forma) –
47%
b) związany z białkami –
46%
( z tego 80% z
albuminą i 20% z globulinami)
c) związany z innymi jonami (cytrynianem,
fosforanem, siarczanem lub mleczanem ) –
7%

3
Rola jonów Ca
2+
- Kofaktor wielu enzymów
- Bierze czynny udział w procesach krzepnięcia krwi
- Regulator aktywności ATP-az
- Bierze udział w regulacji przepuszczalności błon
komórkowych
- Wymianie jonów
- Powstawaniu potencjałów czynnościowych
- W przewodzeniu bodźców nerwowych
- Warunkuje prawidłowy przebieg skurczu i
rozkurczu mięśni
- Regulator uwalniania hormonów
- Tworzenie hydroksyapatytów w kości

4
Stężenie
Ca
2+
w przestrzeniach
międzykomórkowych i otoczeniu komórki
• Poziom Ca
2+
w komórce jest bardzo niski,
rzędu 10
-7
mol/l
• W płynie międzykomórkowym 10
-3
mol/l

5
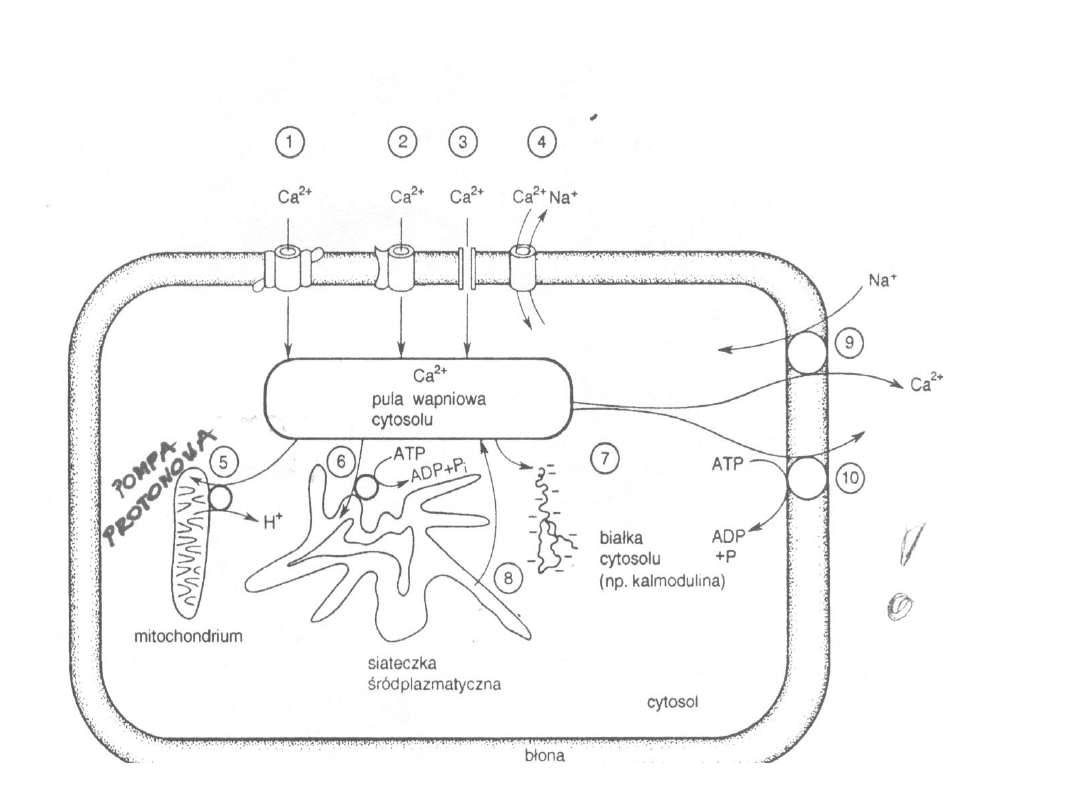
Mechanizmy kontroli stężenia
Ca
2+
w
komórce
Drogi wnikania Ca
2+
do komórki:
kanały wapniowe
regulowane przez:
- potencjał błonowy
- neurotransmitery
- receptory nikotynowe (w płytce
neuromięśniowej)
- receptory ATP (w mięśniach gładkich)
* ok. 1% Ca
2+
przedostaje się do komórki poprzez
kanał sodowy

6
Kanały wapniowe regulowane przez
potencjał błonowy
- wyst. w mięśniach, neuronach i komórkach
wydzielniczych
- Jest bezwzględnie potrzebny do sprzężenia
między pobudzeniem a skurczem w mięśniach
szkieletowych, gładkich i mięśniu sercowym
- Antagoniści kanałów L – leki przeciw nadciśnieniu

7
Kanały wapniowe sterowane przez
neurotransmitery
- Najbardziej znane są aktywowane przez kwas
glutaminowy, a także przez jego analog – kwas
metylo-asparaginowy (NMDA)
- jego aktywacja związana jest z
uczeniem się i śmiercią neuronalną w wyniku
niedotlenienia
8
Regulowan
e
napięciem
Regulowane przez
cząstkę sygnałową
(np.
neurotransmiter
)
przeciekanie
Działający niefizjologicznie (odwrotnie)
Wymiennik Na
+
/Ca
2+
Wymiennik
Na
+
/Ca
2+
Pompa
wapienna

9
Usuwanie Ca
2+
do przestrzeni
międzykomórkowej
1) Pompa wapniowa (Ca
2+
-ATPaza)
-
Układ wykorzystujący swobodną energię
uwolnioną w procesie hydrolizy ATP do transportu
jonów wbrew gradientowi stężeń
-
Wysokie powinowactwo do Ca
2+
, ale stosunkowo
małej pojemności (pracuje dokładnie, ale powoli)
-
Wymaga obecności Mg
2+
-
Hydroliza 1 ATP pozwala na przesunięcie 2 Ca
2+
-
W błonie erytrocytów jest związana z kalmoduliną
-
Zahamowany przez inhibitory metabolizmu
tkankowego
-
Bardzo duża rola w siateczce sarkoplazmatycznej

10
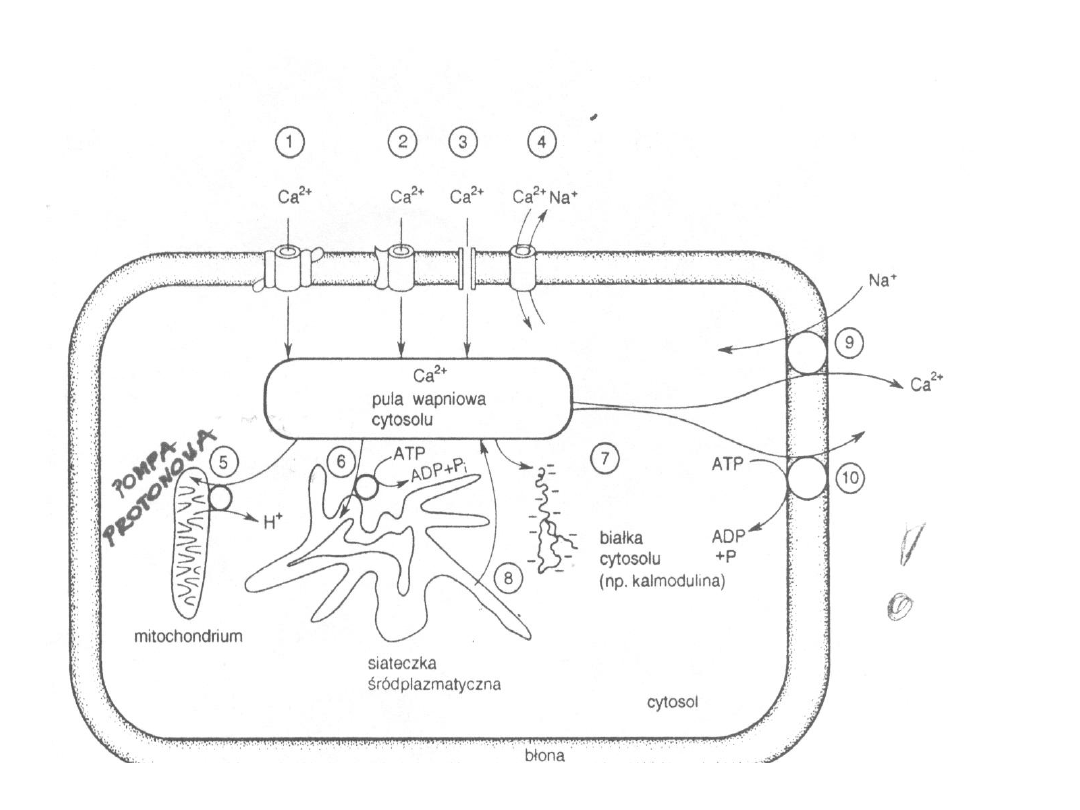
2) Wymiennik sodowo-wapniowy
- Odpowiedzialny głównie za szybkie usuwanie Ca
2+
- Ma niższe powinowactwo do Ca
2+
, ale znacznie
większą pojemność
- Nie występuje w erytrocytach
- Dominującą rolę w usuwaniu Ca
2+
pełni w:
miocytach serca, neuronach, komórkach nabłonka
jelitowego i nerkowego
- Na 3 Na
+
przenoszone do wnętrza komórki
usuwany jest 1 Ca
2+
- W warunkach spoczynkowych w mięśniu sercowym
jest głównym mechanizmem usuwającym Ca
2+
- Umiejscowiony w błonie plazmatycznej i
wewnętrznej błonie mitochondrialnej
11
Regulowan
e
napięciem
Regulowane przez
cząstkę sygnałową
(np.
neurotransmiter
)
przeciekanie
Działający niefizjologicznie (odwrotnie)
Wymiennik Na
+
/Ca
2+
Wymiennik
Na
+
/Ca
2+
Pompa
wapienna

12
Magazynowanie Ca
2+
w organellach
Magazyny płytkie
– o dużym powinowactwie, ale małej
pojemności
- siateczka śródplazmatyczna (w komórkach
mięśniowych jest ładowana wapniem przy udziale
ATPazy zależnej od Ca
2+
i Mg
2+
; wewnatrz siateczki
stężenie Ca
2+
może dochodzić do 10 mmol/l, rola
kalsekwestryny)
- elementy cytoszkieletu i lizosomy
Magazyny głębokie
– małe powinowactwo, duża pojemność
- mitochondria (ładowane przy pomocy wymiennika
protonowego, zaczynają akumulować Ca
2+
gdy stężenie
w cytozolu przekroczy 5-10 umol/l, a więc w stanach
silnego pobudzenia lub patologii)
- jądro
- białka wiążące wapń

13
Białka wiążące wapń i ich rola
- rodzina białek z motywem dłoni EF: kalmodulina,
parwalbumina, bialka z rodziny S-100, troponina C
- białka organelli: kalsekwestryna, kalretikulina
- Białka wiążące fosfolipidy zależne od Ca
2+
: kinaza
proteinowa C, aneksyny
- pozakomórkowe: fibrynogen, α-laktalbumina,
proteinazy serynowe

14
I.
Specyficzne białka receptorowe
a) Rozpuszczone w cytoplazmie
1. kalmodulina (wszystkie tkanki)
2. troponina C (mięsień szkieletowy i sercowy)
3. parwalbuminy
4. leki łańcuch miozyny
5. białko indukowane wit.D – CaBP (głównie jelito)
b)
Związane z błoną komórkową
1. kalmodulina
II. Enzymy aktywowane wapniem nie posiadajace specyficznej
podjednostki regulatorowej
a)
W błonie mitochondrialnej
1. dehydrogenaza glicerolo-3-fosforanowa
b)
W macierzy mitochondrialnej
1. dehydrogenaza pirogronianowa
2. dehydrogenaza α-ketoglutaranowa
c) W cytoplazmie
1. aktywowana wapniem, zależna od fosfolipidów kinaza
proteinowa (C-kinaza)
d) W błonie komórkowej
1. fosfolipaza C
2. fosfolipaza A
2

15
Kalmodulina:
- Białko występujące we wszystkich organizmach
eukariotycznych
- Stężenie w cytoplazmie komórek wynosi aż 10 umol/l
- Posiada 4 miejsca wiązania jony wapnia (1 mol
kalmoduliny wiąże 4 mole Ca
2+
)
- Po związaniu co najmniej 3 jonów Ca
2+
następuje jej
aktywacja
- Kompleks kalmodulina – Ca
2+
reguluje:
* enzymy metabolizmu cyklicznych nukleotydów
(fosfodiesterazy, cyklazy)
* enzymy transportu Ca
2+
* kinazy (np. kinaza lekkich łańcuchów miozyny) i
fosfataza białkowe
* białka wchodzące w skład cytoszkieletu
* fosfolipaza A
2
* uwalnianie neuroprzekaźników
16
Kalsekwestryna i kalretikulina
• Występuje wewnątrz
cystern ER
• Posiada niezmiernie
wysoka zdolność
wiązania Ca
2+
(1 mol
białka – 45 moli Ca
2+
)
• Dominuje w błonach
ER
• Posiada dwa regiony: o
duzej pojemności i
małym powinowactwie
(1mol białka – 20 -25
moli Ca
2+
) oraz o małej
pojemności (1 mol
bialka – 1 mol Ca
2+
)

17
Kinazy proteinowe C:
- Aktywność jest regulowana przez : Ca
2+
, fosfolipidy i
diacyloglicerol (b. znacznie zwiększa powinowactwo
kinazy do Ca
2+
)
- Uczestniczy w regulacji cytoplazmatycznego poziomu
Ca
2+
Aneksyny:
- białka, które w sposób zależny od Ca
2+
wiążą się z
fosfolipidami
- Mają zdolność hamowania aktywności fosfolipazy A
2
- Mogą w zależności od typu : stymulować zależny od
Ca
2+
proces endocytozy, regulować kanały wapniowe
18
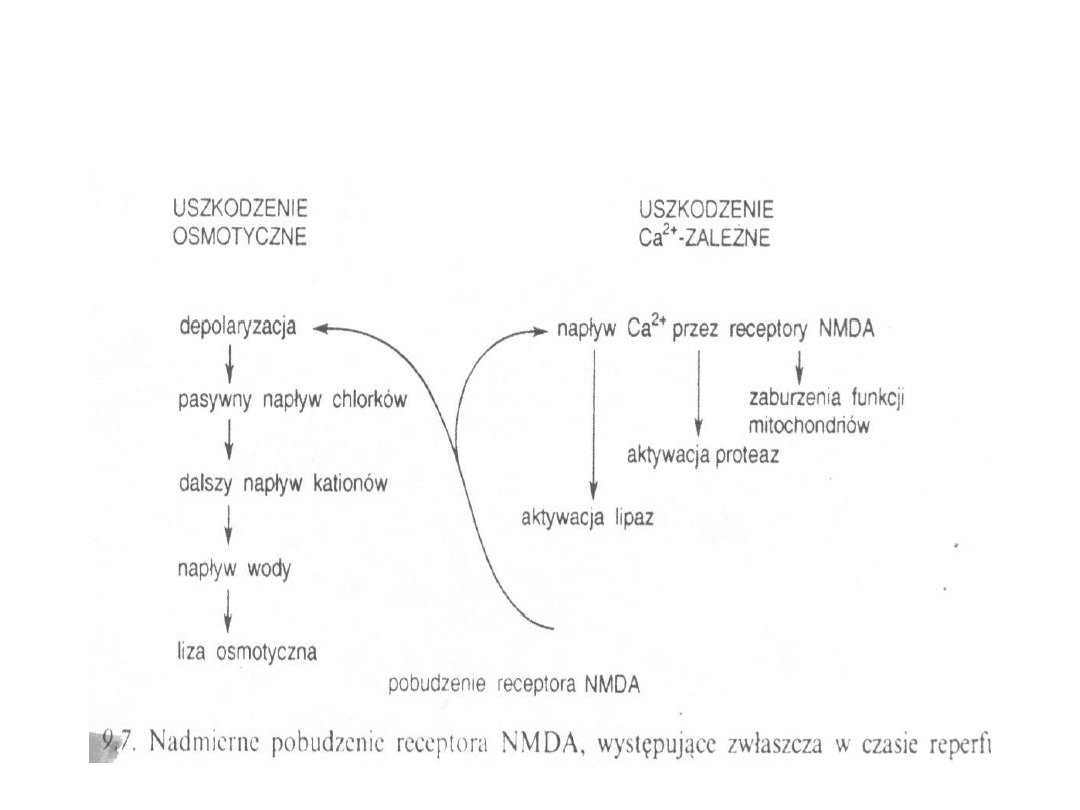
Cytotoksyczne efekty wywołane wysokimi
stężeniami Ca
2+
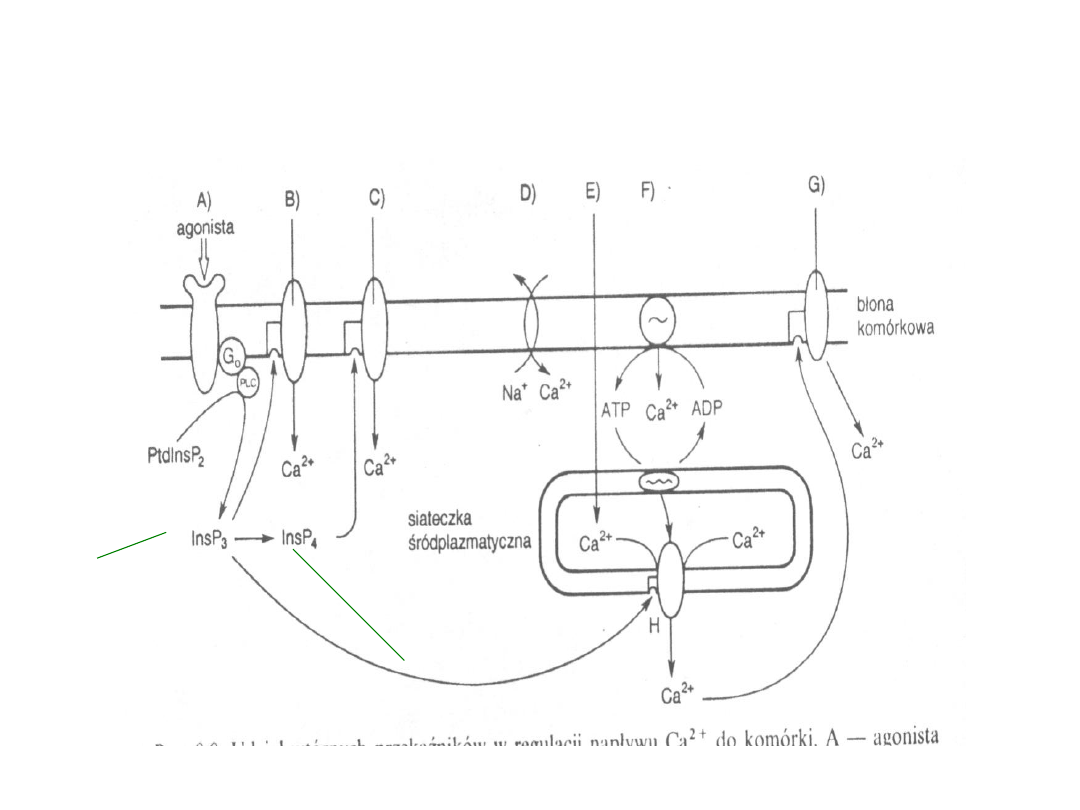
19
Udział wtórnych przekźników w regulacji
napływu Ca
2+
do komórek
Uwalnia jony Ca2+
Ze zbiorniczkow
siateczki
śródplazmatycznej
Ma zdolność aktywacji
Blonowych kanałów wapniowych

20
Wapń osocza
• Stężenie wapnia w osoczu wynosi 2,3 – 2,74 mmol/l
• Stężenie wapnia w osoczu zależy od wieku –
większe jest u noworodków
• Dorosły człowiek spożywa ok. 1g/24h
• Dobowe zapotrzebowanie 10-15mg/kg masy ciała
• Fosforany i kwasy tłuszczowe wiążą wapń w jelicie i
uniemożliwiają jego wchłanianie
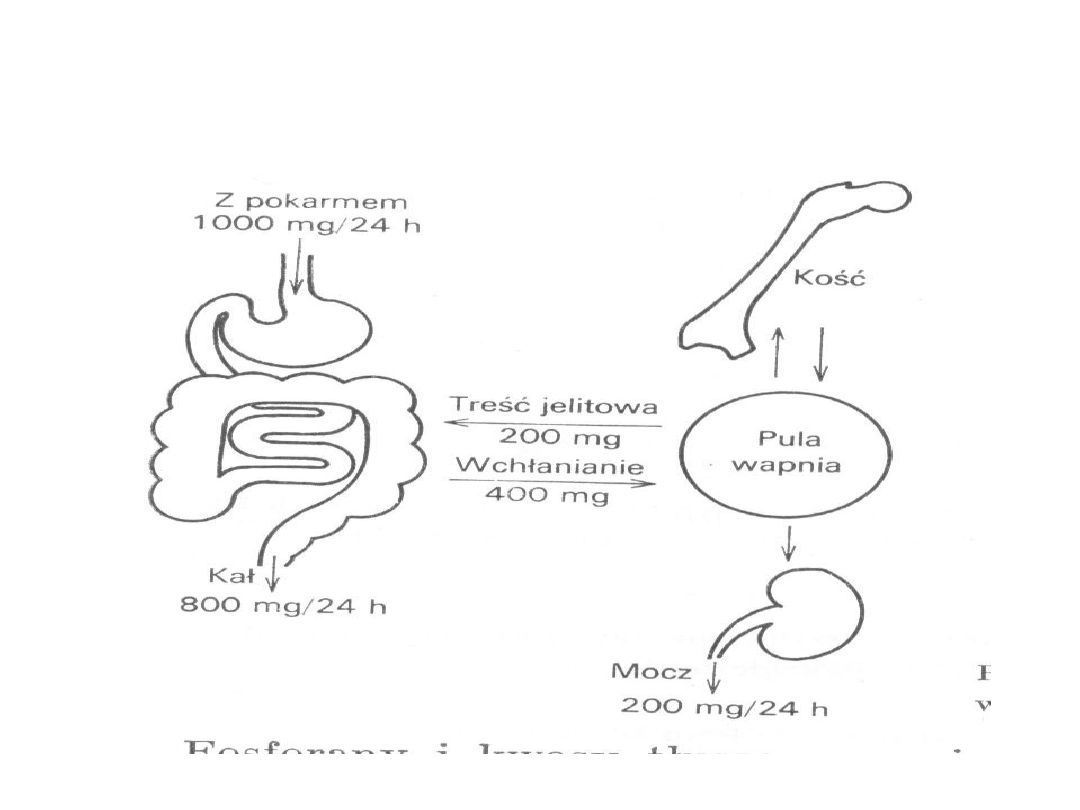
21
Wchłanianie wapnia

22
Za regulację stężenia wapnia
odpowiedzialne są:
1) narządy:
- kości
- jelito cienkie
- nerki
2) Substancje regulacyjne:
- parathormon
- witamina D
3
- kalcytonina

23
W wyniku zmniejszonej podaży wapnia w diecie
dochodzi do umiarkowanej
hipokalcemii
, co
powoduje:
- Uwalnianie parathormonu z przytarczyc
- Zwiększonego przeksztalcania 25(OH)
2
D
3
w
1,25(OH)
2
D
3
w mitochondriach kanalików
proksymalnych nerek
* 1,25(OH)
2
D
3
i parathormon zwiększają wchłanianie
wapnia w jelicie, jak i uwalnianie z kości

24
Przy zwiększonej podaży wapnia w diecie –
hiperkalcemia:
- Zmniejsza się wydzielanie parathormonuz
przytarczyc
- Zwiększa się wydzielanie kalcytoniny wytwarzanej
w komórkach C tarczycy
- W nieobecności parathormonu maleje aktywność
1α-hydroksylazy 25(OH)D
3
, a zwiększa się
aktywność 24-hydroksylazy – prowadząc do
zwiększonego wytwarzania 24,25(OH)
2
D
3
(pobudza wyłącznie wchłanianie wapnia w jelicie,
nie ma wpływu na uwalnianie wapnia z kości)
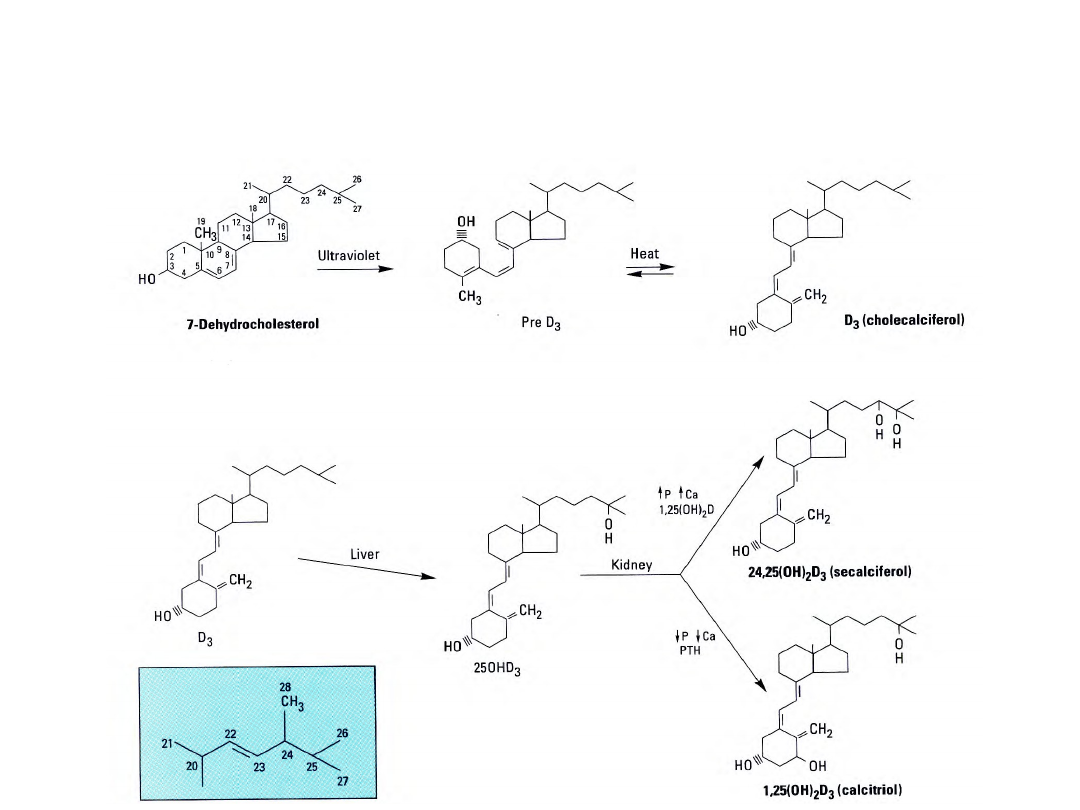
25
Przemiany witaminy D
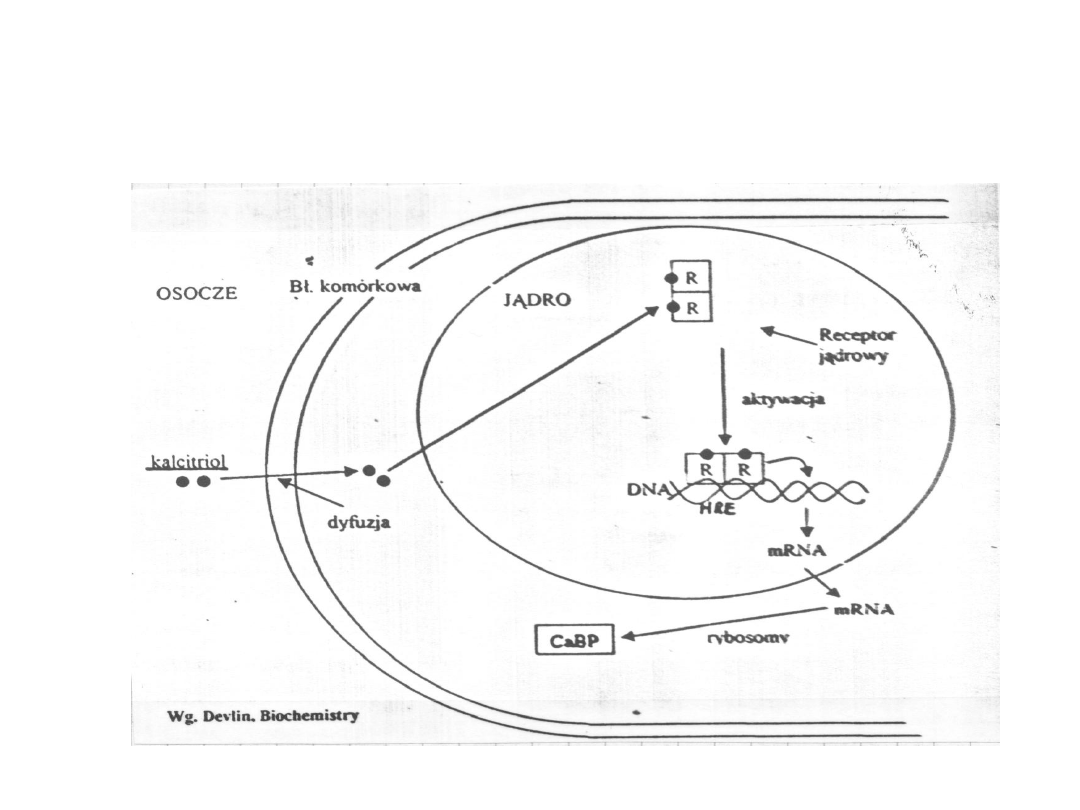
26
Działanie popprzez receptory jądrowe
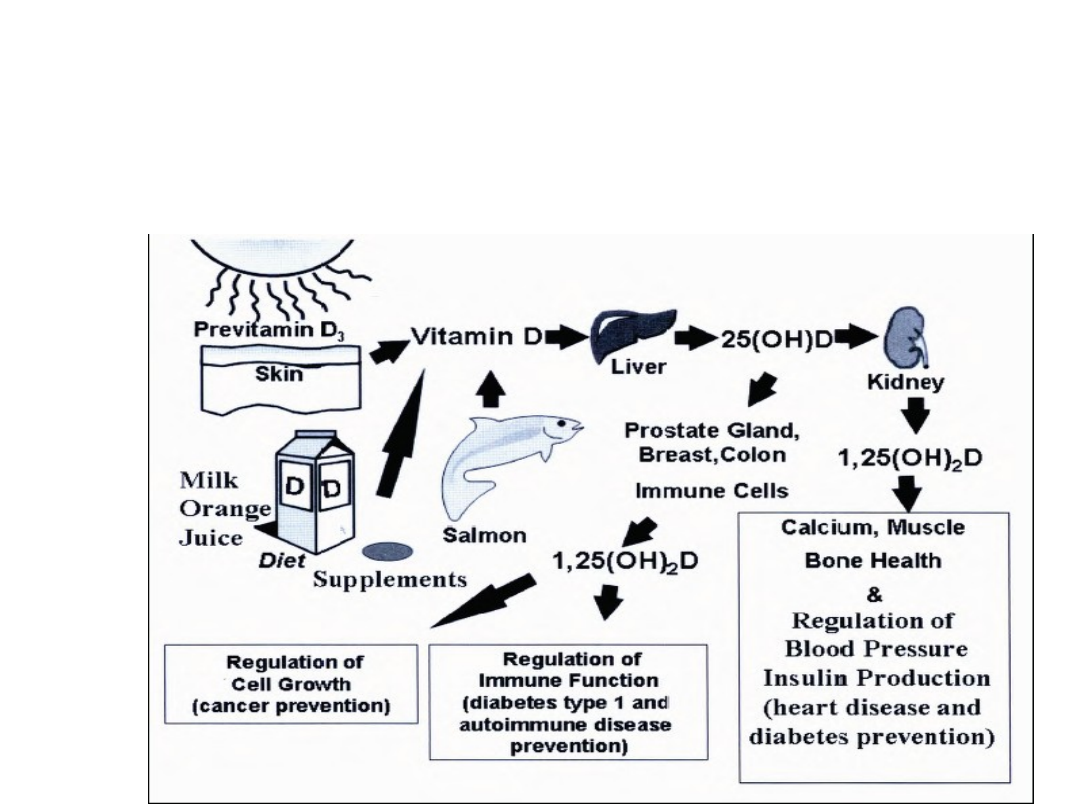
27
Inne działania wit. D

28
Niedobór wit. D
- U dzieci wywołuje krzywicę
- U dorosłych osteomalację (rozmiękanie kości) i
osteoporozę
- Bezpośrednim efektem niedoboru wit. D jest
hipokalcemia, która jest kompensowana przez wzrost
uwalniania parathormonu z przytarczyc, a ten szybko
przywraca normalny poziom Ca
2+
w osoczu
- Prowadzi do ubytku zasobów kostnych wapnia i
fosforanu, powoduje demineralizację tkanki kostnej
* Hiperwitaminoza D prowadzi do hiperkalcemii i
hiperkalciurii i powstawania ognisk zwapnienia w
tkankach miękkich

29
Parathormon (PTH)
-
Polipeptyd składający się z 84 aa
-
Wytwarzany przez komórki przytarczyc w formie
prekursora
-
Główny regulator homeostazy wapniowej
-
Wykazano odwrotną zależność pomiędzy stężeniem
tego hormonu a stężeniem jonów wapnia w osoczu
* wydzielanie PTH ustaje, gdy [Ca] wzrośnie do 3
mmol/l
-
Okres półtrwania 20 min
-
Wpływa na czynność nerki (utrata jonów fosforanowych
z moczem, doprowadza do zmniejszenia ich w osoczu)
-
Pobudza cyklazę adenylanową – rośnie cAMP (wpływ na
transport bierny jonów wapniowych z płynu
pozakomórkowego do komórki)
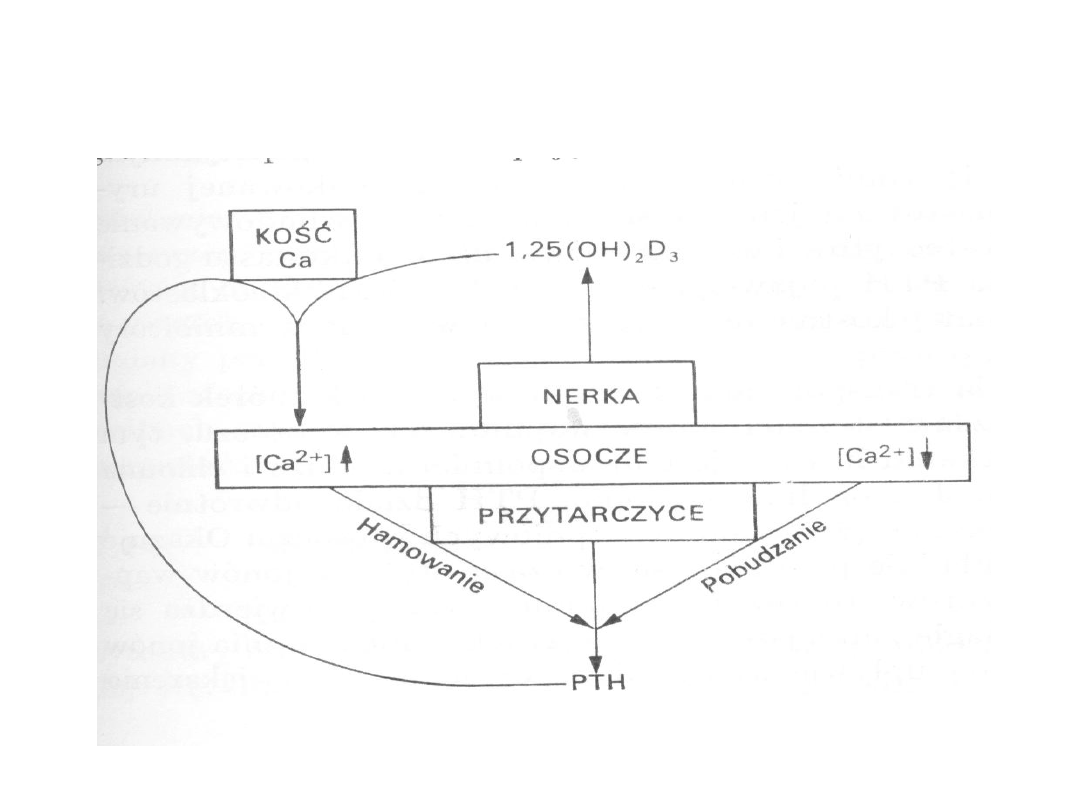
30
Mechanizm działania PTH i
1,25(OH)
2
D
3

31
Główne działania witaminy D i
parathormonu

32
Kalcytonina
- Wytwarzana przez komórki C tarczycy, przytarczycach
i niekiedy w grasicy
- Jej stężenie we krwi jest tego samego rzędu co PTH
(10
-10
mol)
- Okres półtrwania kilka minut
- Miejscem działania jest tkanka kostna i
prawdopodobnie jelito, działanie przeciwstawne do
działania PTH
- Hamuje resorpcję kości pobudzaną przez PTH
- Zapobiega rozpadowi kolagenu kości
- Zmniejsza ilość osteoklastów i ich aktywność
resorpcyjną
- Znosi mobilizujące działanie PTH, blokując
demineralizację kości spowodowaną przez cAMP
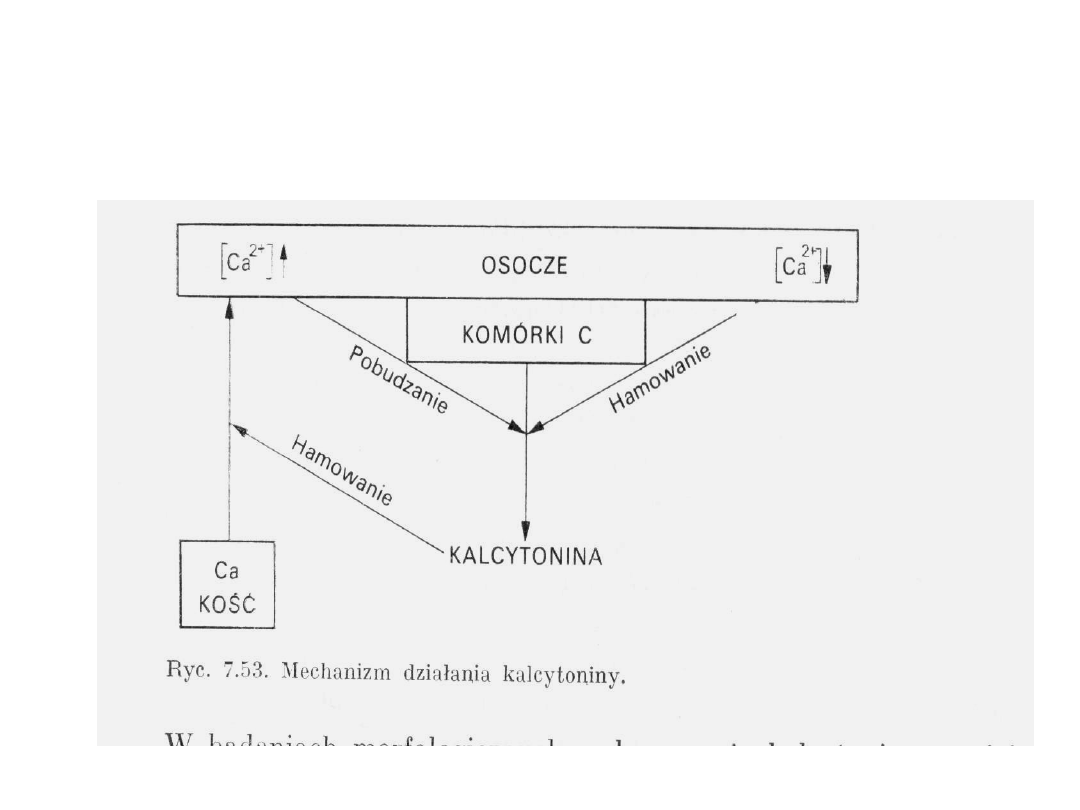
33
Mechanizm działania kalcytoniny
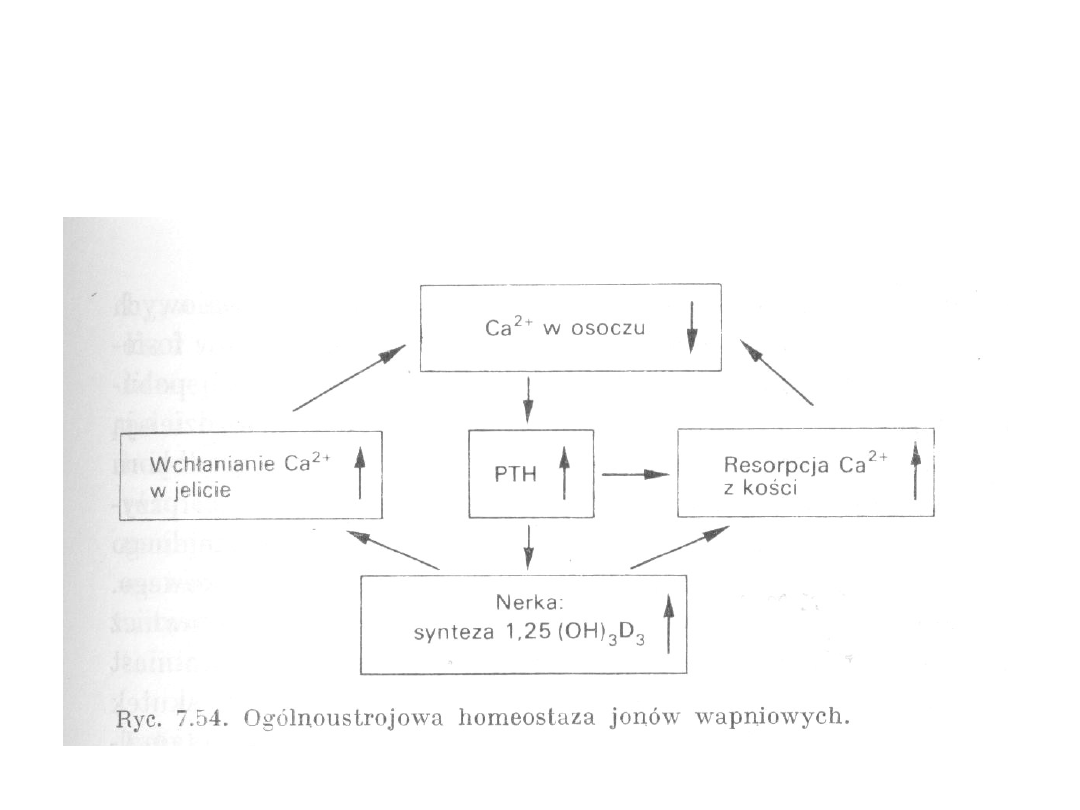
34
Homeostaza jonów wapnia
Zmniejszone stężenie Ca
2+
w
osoczu
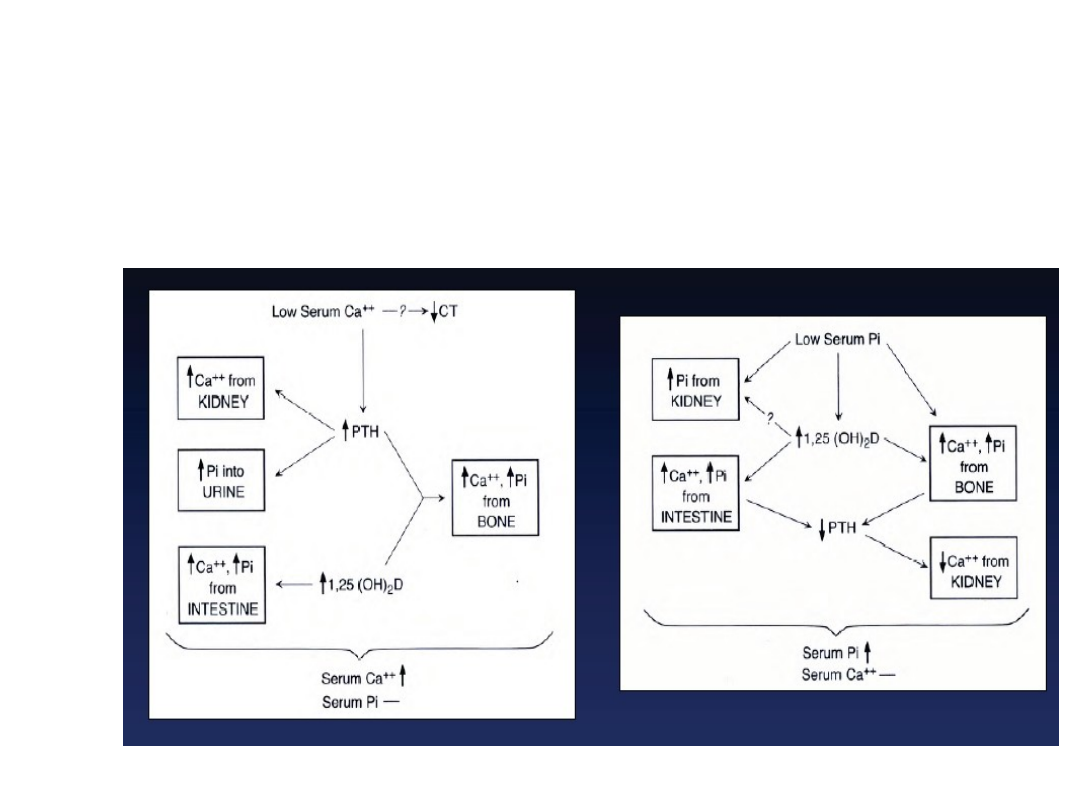
35
Reakcja na niskie stężenie
wapnia i fosforanu we krwi
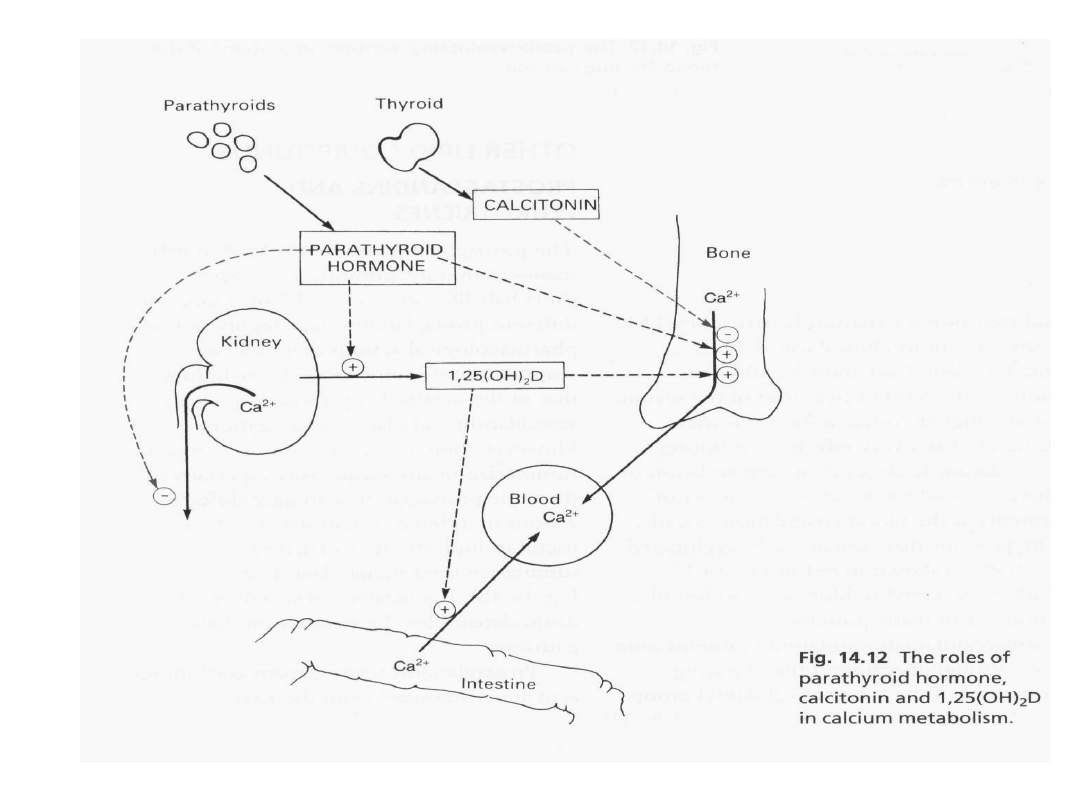
36
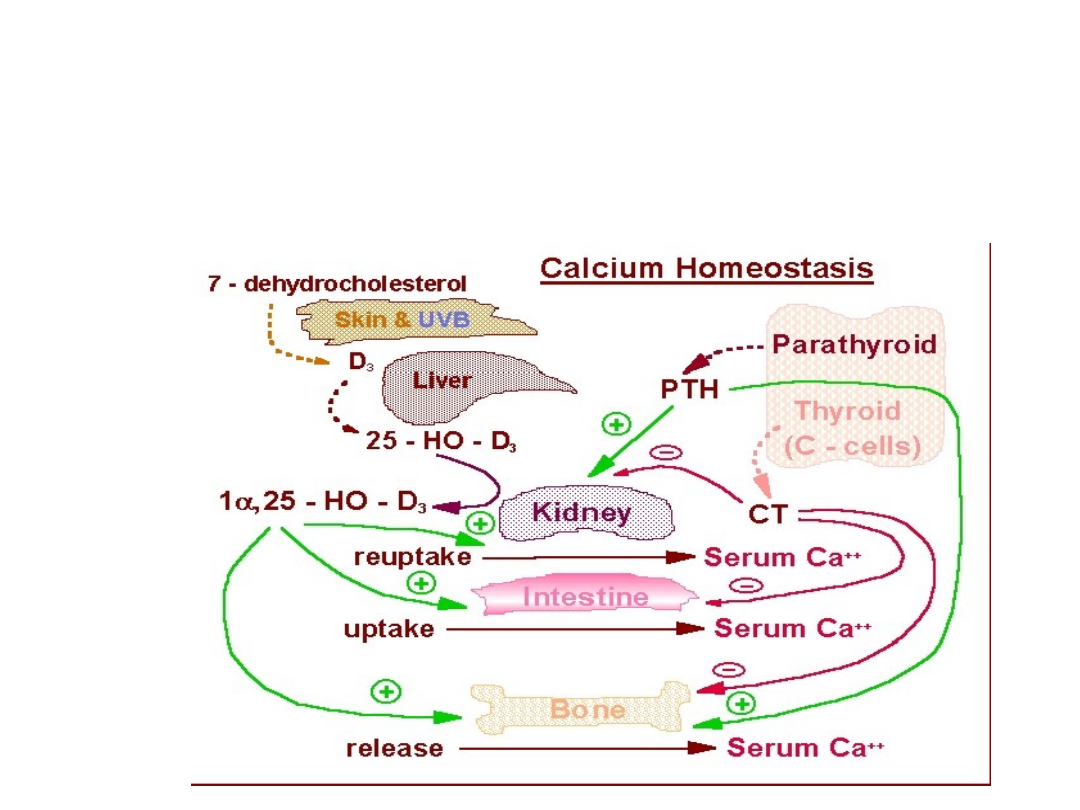
37
Hormony regulujące przemiany wapnia
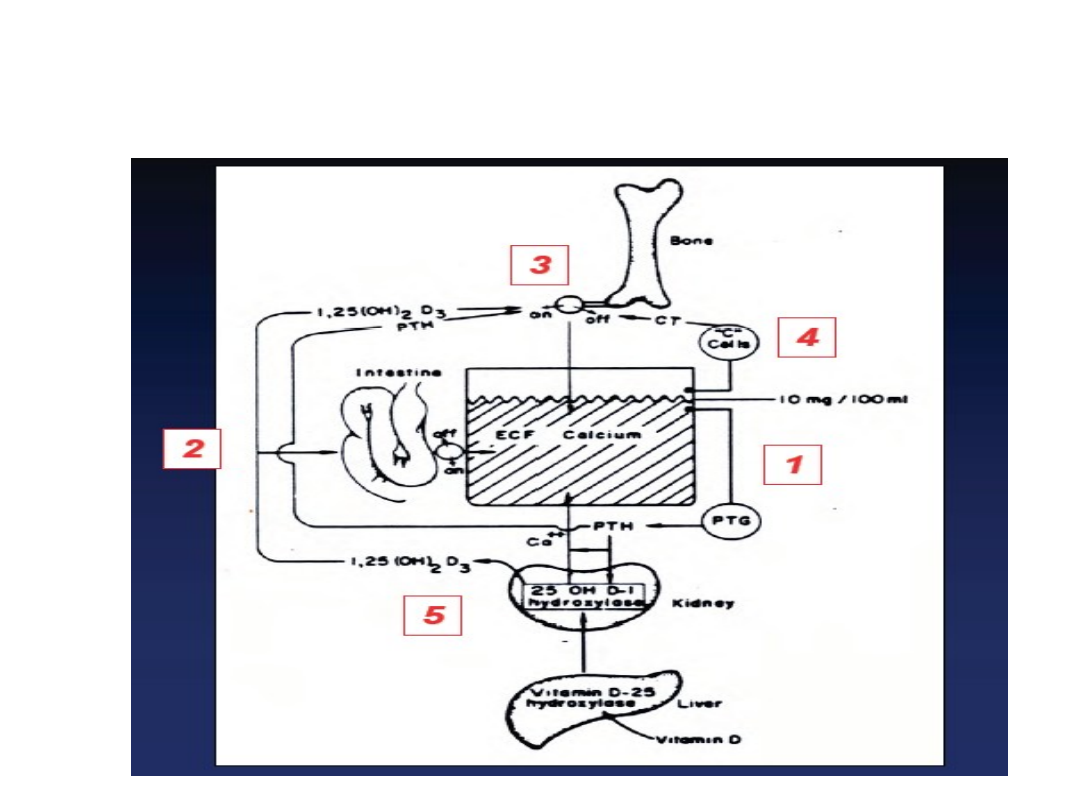
38
Regulacja stężenia wapnia w osoczu
39
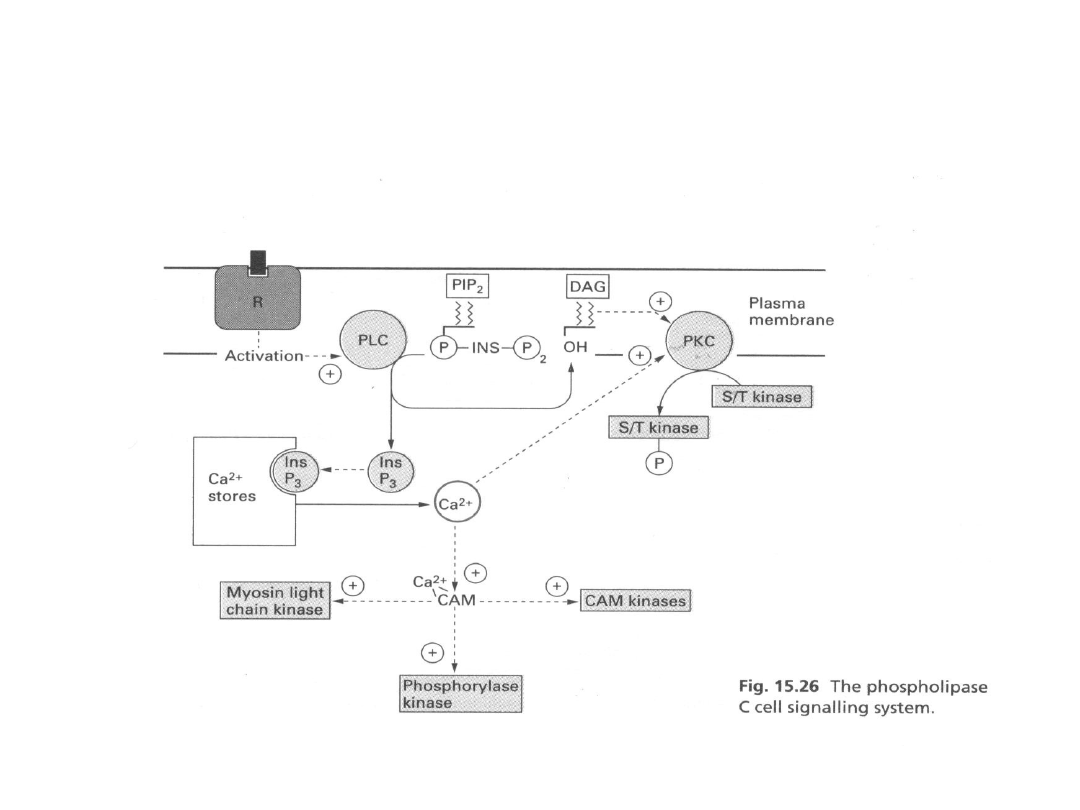
Rola jonów wapnia jako drugi przekaźnik

40
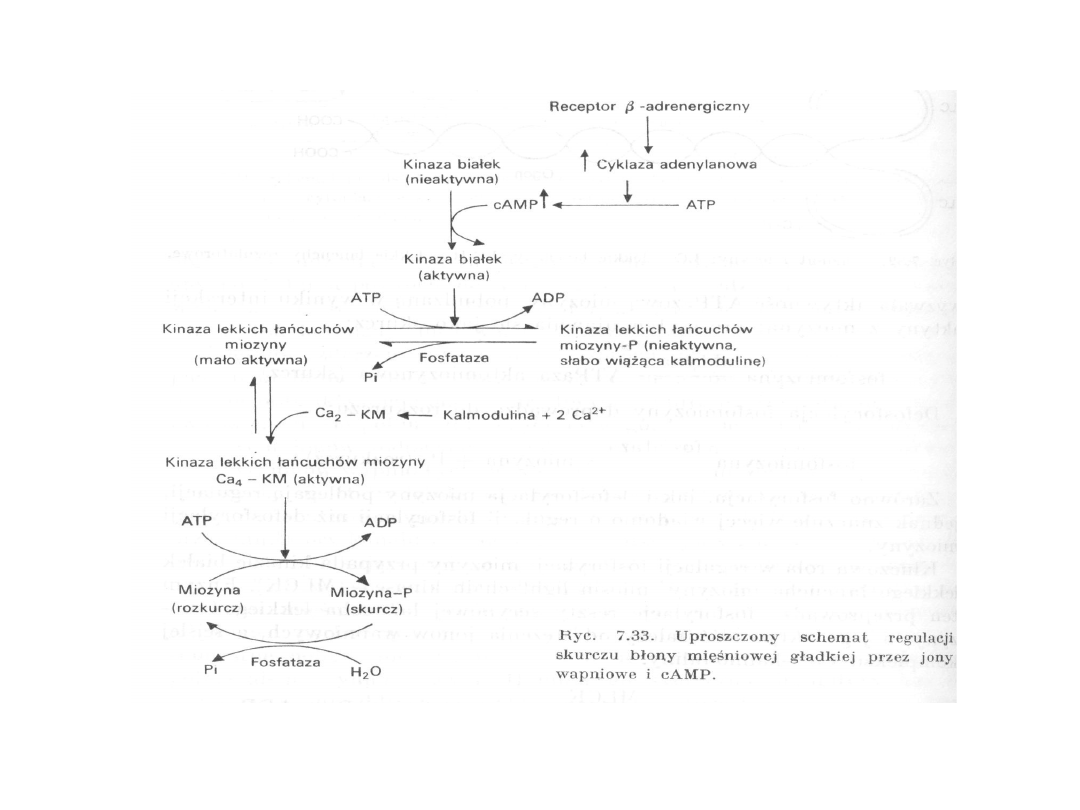
Rola jonów wapnia w skurczu
-skurcz jest uruchamiany przez gwałtowny wzrost
stężenia Ca
2+
-rozkurcz zależy od szybkiego usunięcia Ca
2+
z
cytozolu
- białkiem pośredniczącym w oddziaływaniu Ca
2+
na
proces skurczu mięśni szkieletowych i mięśnia
sercowego
41

42
Rola jonów wapnia w budowie kości
Tkanka kostna składa się z utkania organicznego
(95% kolagen) i podłoża mineralnego (stanowi 65-
75%wagowych)
- W obrębie podłoża mineralnego wyróżnia się 2
pule fosforanu wapniowego
* pulę bezpostaciową (amorficzną)-przeważa
w kości mlodej
* krystaliczną podobną do hydroksyapatytu
Ca
10
(PO
4
)
6
(OH)
2
- W niedoborze wit. D
3
przeważa postać
amorficzna, maleje pula krystaliczna (obecność D
3
warunkuje prawidłowe uwapnienie kości)

43
Resorpcja i tworzenie kości
- Między 20-30 rokiem życia te procesy są
zrównoważone
- Po 40 – resorpcja przeważa nad tworzeniem
- U podstaw przebudowy leży wymiana jonów Ca
2+
i
PO
43-
między osoczem krwi a zmineralizowaną
macierzą tkanki kostnej – szybkość tej wymiany
podlega kontroli PTH, 1,25(OH)
2
D
3
i kalcytoniny
(hamuje ona resorpcję kości zarówno w obecności
jak i niedoborze D3)
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
- Slide 38
- Slide 39
- Slide 40
- Slide 41
- Slide 42
- Slide 43
Wyszukiwarka
Podobne podstrony:
w14 redox, Nernst
W14 3
14 TIOB W14 zelbet i klasyfikacja deskowan
Microsoft Word W14 Szeregi Fouriera
Odp do W14
FChUK W14 Uplynnianie betonow
14 TIORB W14 roboty ciesielskie, TIORB
w14
Nie - Boska komedia, W14, Na podstawie części I i II dramatu zreferuj zasadnicze myśli o poezji i po
W14 Logika modalna
TRB W14 12 01 13 o wszytkim i niczym
FO W14
w14
w14 15 test grupy cwiczeniowe
W14 2Wspolczesnezarzadzanie
więcej podobnych podstron