
Hormony roślinne
• Auksyny
• Gibereliny
• Cytokininy
• Brasinosteroidy
• Kwas abscysynowy
• Etylen
• Kwas jasmonowy
• Poliaminy
• Kwas salicylowy
• Związki fenolowe (allelopatiny)
Klasyczne hormony
roślinne
Regulatory wzrostu
stymulator
y
inhibitory
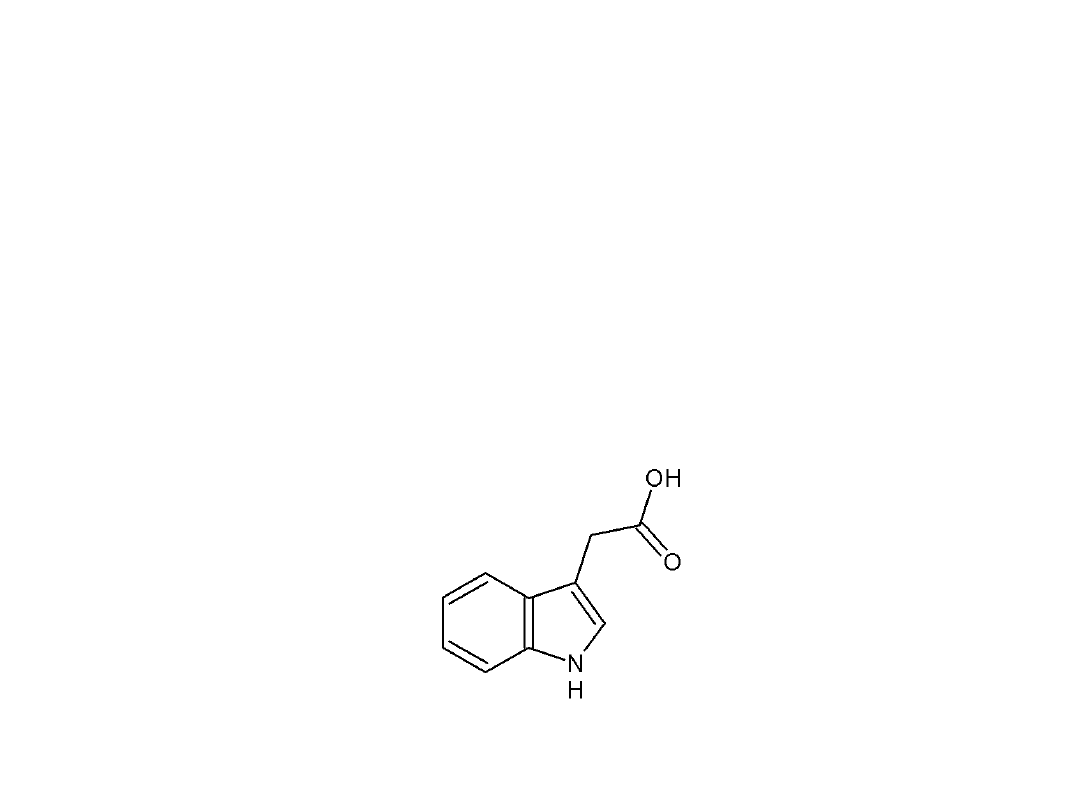
AUKSYNY – hormony wzrostu
Występowanie auksyn
•auksyny występują
we wszystkich roślinach
wyższych
oraz w niektórych gatunkach grzybów;
• są syntetyzowane głównie w merystemach
wierzchołkowych, głównie pędów (także korzeni),
najmłodszych liściach;
• z miejsca swojego powstania przemieszczają się do
innych części rośliny (bazypetalnie);
• różna podatność poszczególnych części roślin na
działanie auksyn;

Auksyny występują w komórkach w
formie wolnej i związanej
Koniugaty
auksyn:
• są to kompleksy auksyny z cukrami (glukoza i myo-
inozytol);
• są nieaktywne biologicznie;
• jest to transportowa lub akumulowana forma
hormonu.

Aktywność biologiczna auksyn
• Auksyny uczestniczą w bardzo różnorodnych procesach
fizjologicznych związanych
ze wzrostem i rozwojem
roślin:
• stymulują wzrost elongacyjny ściany komórkowej
• kontrolują dominację wierzchołkową
• odpowiadają za fototropizm, geotropizm
• stymulują tworzenie zawiązków korzeniowych
• pobudzają rozwoju owoców (partenokarpia)
• hamują opadaniu liści i owoców
• biorą udział w naprawie zranień
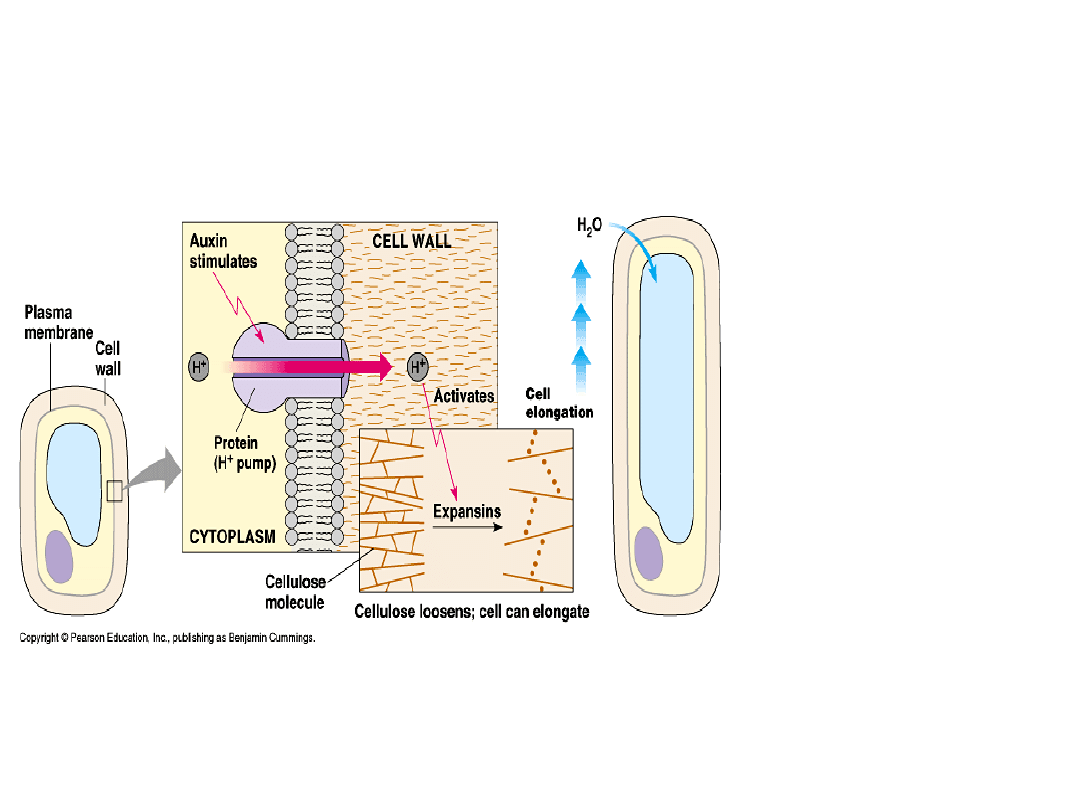
Auksyny stymulują wzrost elongacyjny
ściany komórkowej (tzw. wzrost
kwasowy)
-IAA aktywuje pompę
protonową zależną od ATP
w plazmalemie lub ER
-Protony przemieszczają się
do apoplastu, co powoduje
zakwaszenie apoplastu
-Następuje aktywacja
enzymów hydrolitycznych i
rozerwanie wiązań
wodorowych pomiędy
mikrofibrylami
celulozowymi
- To prowadzi do
rozluźnienia struktury
ściany komórkowej
szybka odpowiedź na
hormon

Dominacja wierzchołkowa
A – dominuje pąk wierzchołkowy (IAA syntetyzowany w
merystemie transportowany
bazypetalnie
powoduje uśpienie
pąków pachwinowych (spoczynek)
B – usunięcie pąka wierzchołkowego powoduje uwolnienie
pąków bocznych
C – po dekapitacji umieszczenie bloczka agarowego z auksyną
( + IAA) powoduje uśpienie pąków bocznych (C=A)
D –po dekapitacji umieszczenie
bloczka agarowego kontrolnego
( - IAA) powoduje uwolnienie pąków
bocznych (D=B)
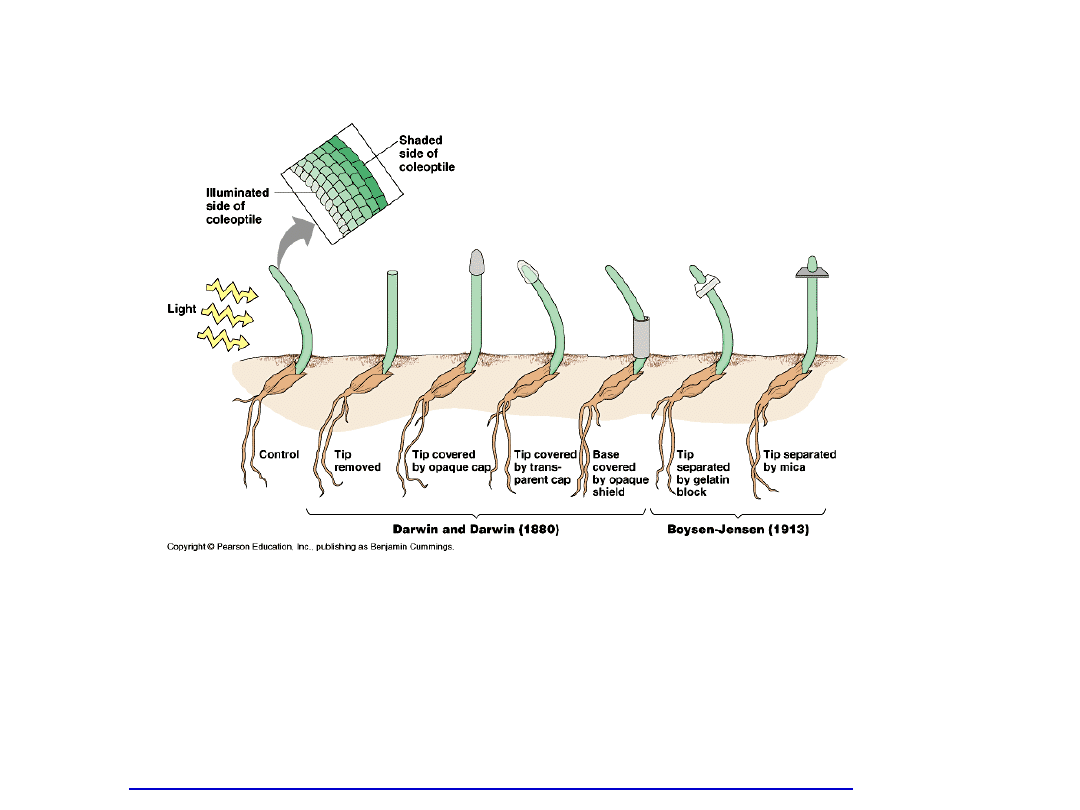
Fototropizm
-
Mechanizm zjawiska fototropizmu tłumaczy się
nierównomiernym
rozmieszczeniem auksyn w strefie wzrostu koleoptyla.
- Na stronie zacienionej występuje większe nagromadzenie
auksyn
.
- Następuje zahamowanie syntezy IAA po stronie
oświetlonej i
poprzeczny transport auksyn na stronę zaciemnioną.

Geotropizm
• Nierównomierny rozkład auksyn w strefie reagującej na bodźce.
• auksyn przemieszczają się w roślinie w kierunku pola
przyciągania ziemskiego.
• pędy ułożone w pozycji poziomej zawierają więcej auksyn po
stronie dolnej,
• większa wrażliwość na auksynę po stronie dolnej.
• nagromadzenie auksyn powoduje geotropizm ujemny pędu i
geotropizm dodatni korzenia
.
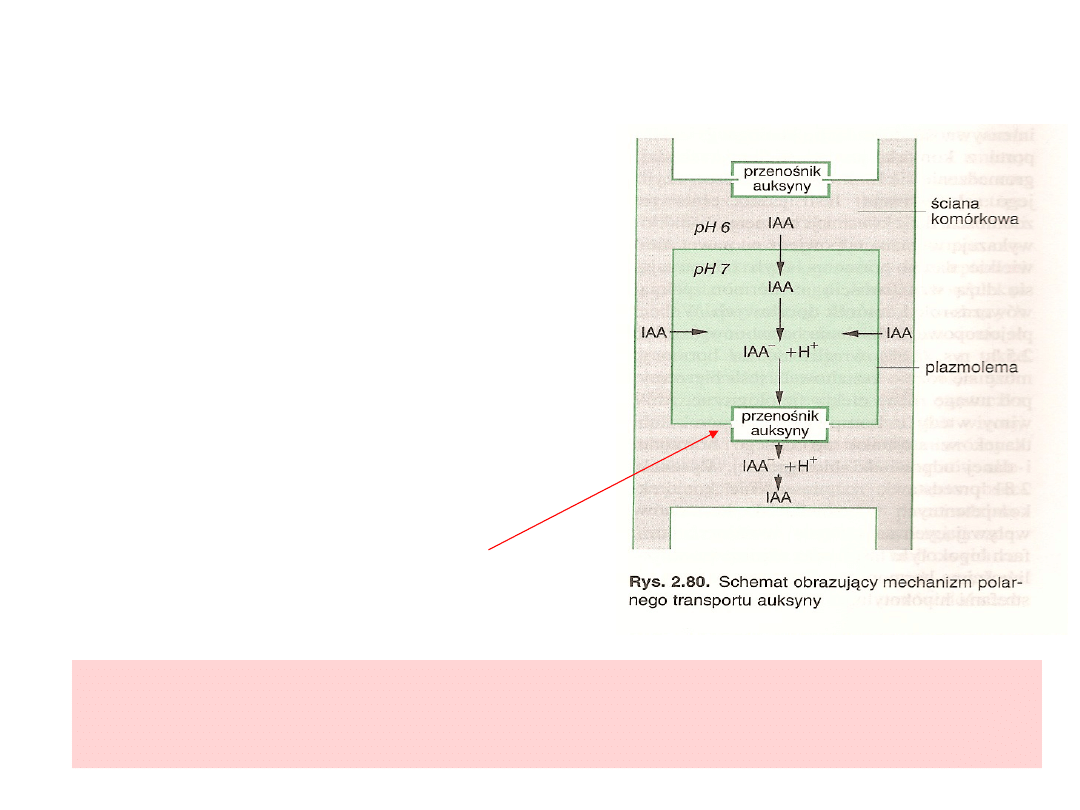
Polarny transport auksyn (bazypetalny)
- Apoplast ma odczyn kwaśny
- IAA w apoplaście występuje w
formie niezdysocjowanej
- Wewnątrz komórki (pH 7) IAA
ulega jonizacji i jako IAA
-
nie może
przeniknąć plazmolemmy
-Przenośniki anionu IAA
-
zlokalizowane w błonie
komórkowej w sposób aktywny
wypompowują IAA
-
z cytoplazmy
do ściany komórkowej
- Przenośniki te są rozmieszczone
niesymetrycznie w cytoplazmie,
znajdują się na bazalnej stronie
komórek, co warunkuje kierunek
transportu.
Polarny transport powoduje stałe utrzymanie w roślinie
przepływu auksyn od wierzchołka pędu do korzenia, co jest
ważnym mechanizmem determinującym polarność
organizmu i jego skoordynowany harmonijny rozwój.
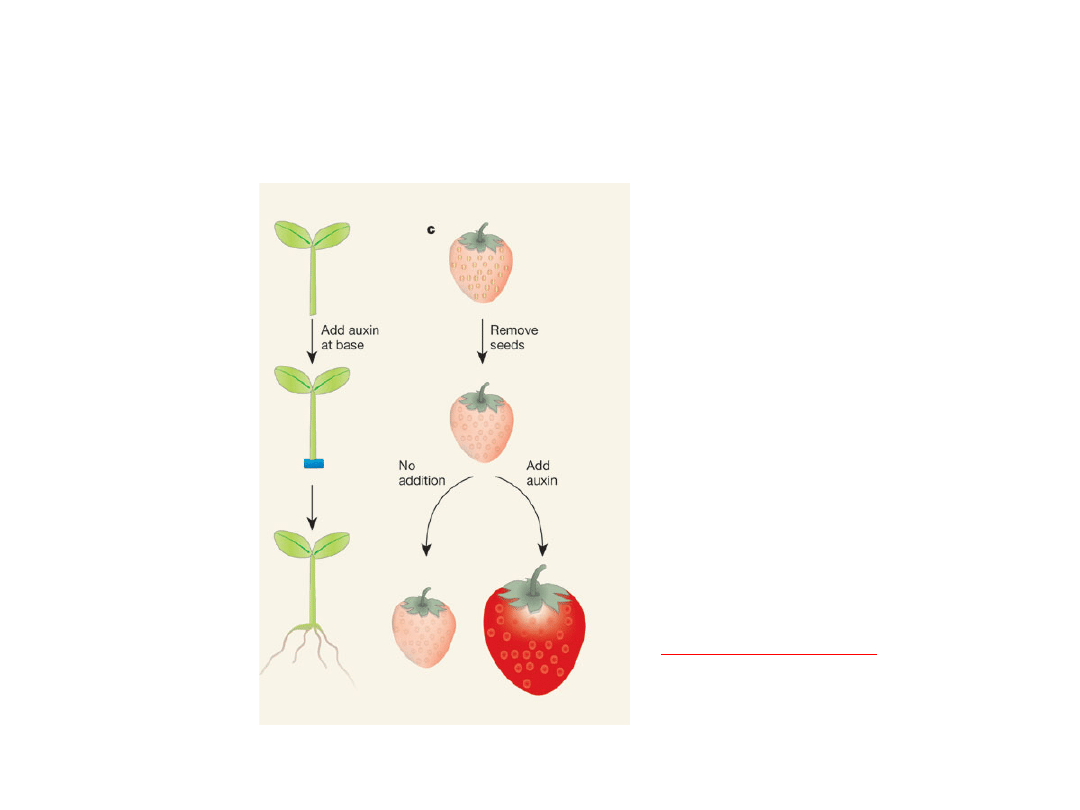
Auksyny stymulują ukorzenianie i
rozwoju owoców
-Usunięcie niełupek
(naturalnego źródła IAA)
-Aplikacja IAA powoduje:
- rozrost dna
kwiatowego
- rozrost zalążni
- produkcja
bezpestkowych
owoców
Partenokarpia -
indukowanie wytwarzania
owoców z pominięciem
zapylenia.
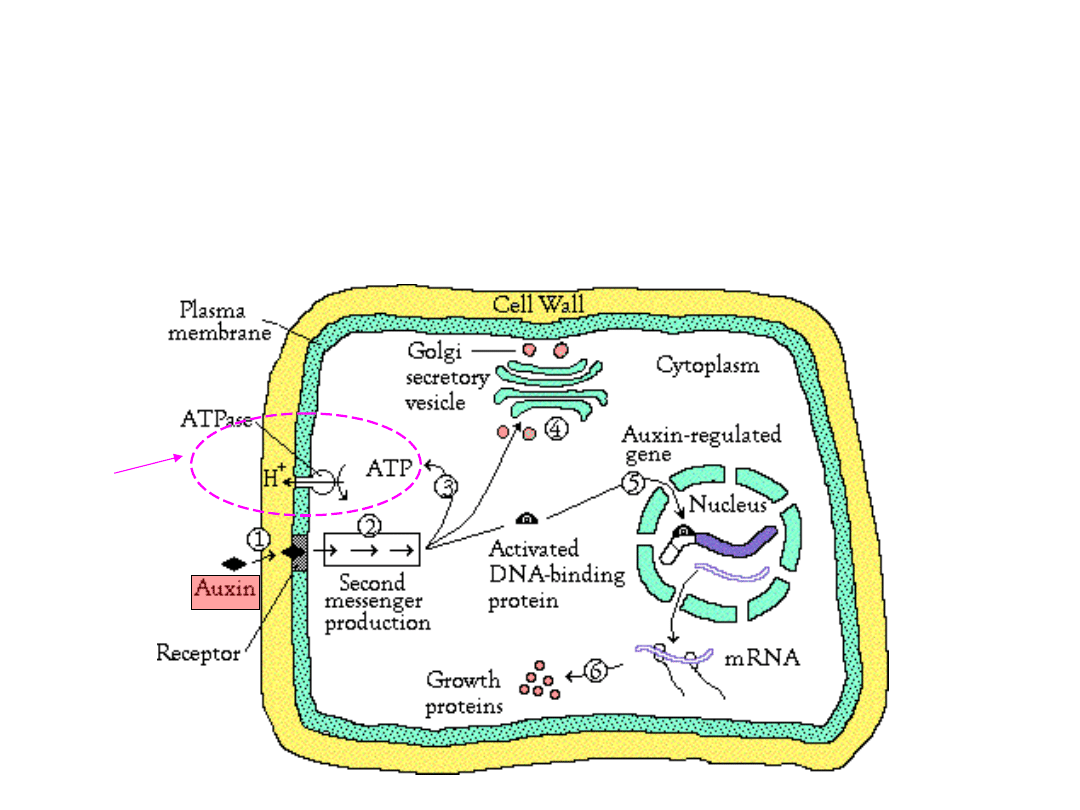
Szlak transdukcji sygnału
indukowanego przez auksynę
W szlaku transdukcji sygnału biorą udział różne wtórne
przekażniki sygnału (np. jony wapnia, IP3))
Szybka
odpowiedż
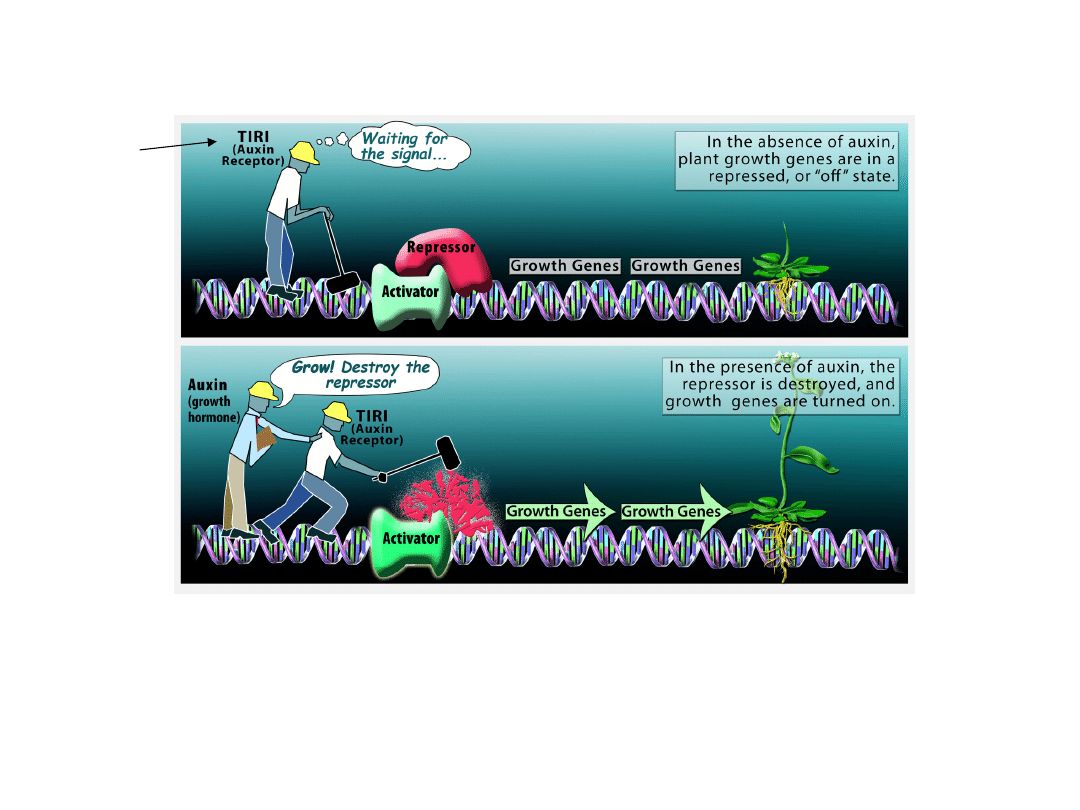
Molekularny mechanizm sygnału
auksynowego
• (a) Ekspresja genów odpowiedzi na auksynę jest zablokowana przez
represor
• (b) Auksyna łączy się z receptorem TIR1
Następuje destrukcja represora
Uruchomiona jest ekspresja genów związanych z odpowiedzią
fizjologiczną rośliny na ten hormon ------- WZROST
TIRI
recepto
r

Gibereliny- hormony
„wysokości”
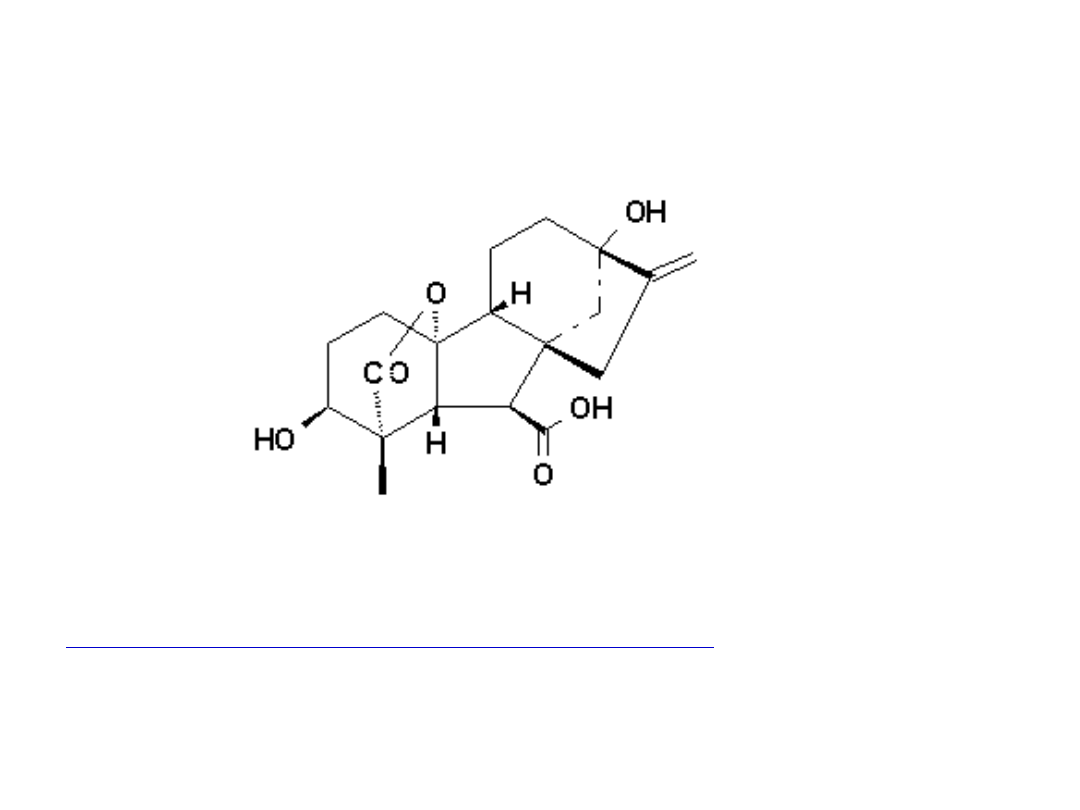
Struktura chemiczna giberelin
Znanych jest obecnie ponad 120 giberelin.

Występowanie i formy
giberelin
Występują u wszystkich roślin
• jako wolne,
aktywne
związki
• w kompleksach typu glikozydów i estrów
glikozydowych
• związane z białkiem
Organy obficie produkujące gibereliny to:
• części wierzchołkowe korzeni i najmłodsze liście, ale
nie merystem,
• węzły u traw i pędów dwuliściennych,
• pręciki,
• rozwijające się nasiona.

Gibereliny- efekty fizjologiczne
działania
• Stymulacja wzrostu pędu przez stymulację podziałów
komórkowych i wzrost elongacyjny komórek (
wraz z
auksynami
).
• Stymulacja kwitnienia u roślin krótkiego dnia
• Przerywanie spoczynku nasion, bulw, pąków
• Stymulacja kiełkowania nasion
• Indukcja tworzenia owoców partenokarpicznych (
wraz z
auksynami
).
• Opóźnianie procesu starzenia liści i owoców cytrusowych
.
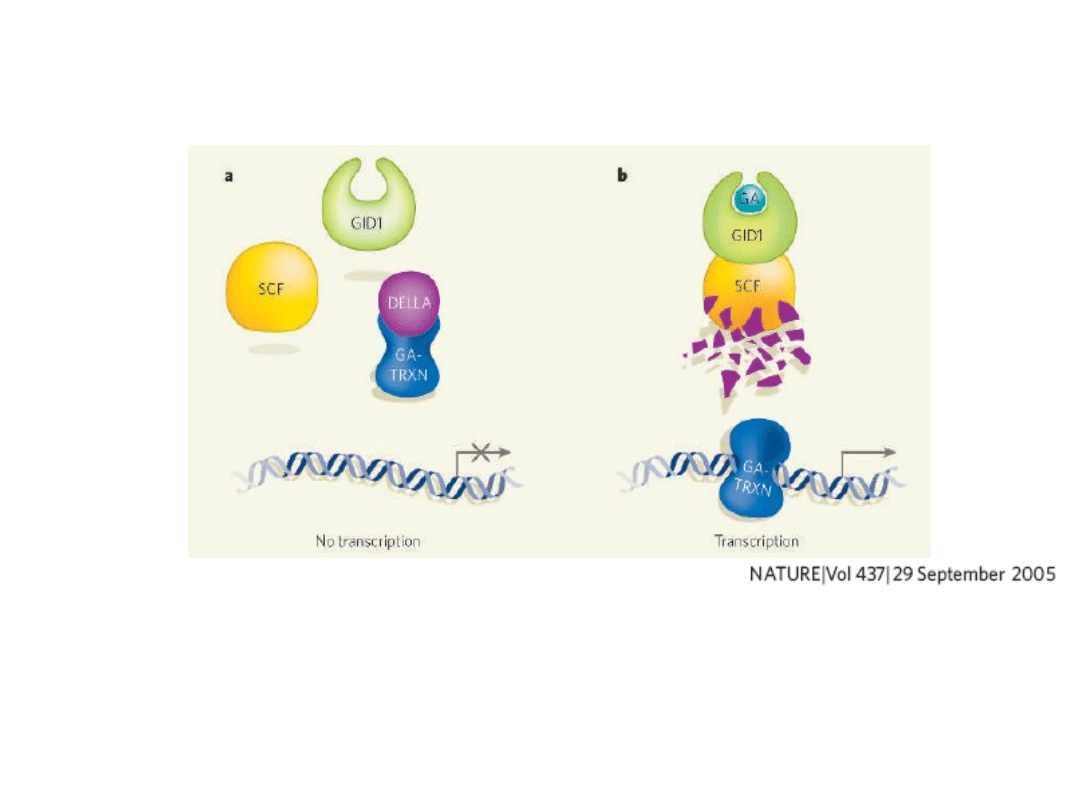
Molekularny mechanizm sygnału
giberelinowego
Obecność negatywnych regulatorów
DELLA
uniemożliwia ekspresje genów regulowanych przez GA,
które kontrolują
kiełkowanie, wydłużenie łodygi,
kwitnienie ...... inne procesy.

Praktyczne zastosowanie giberelin
(GA
3
)
• Winogrona beznasienne o dużych rozmiarach
• GA
4
+ GA
7
+cytokininy stosowane dla polepszenia kształtu
jabłek (bez wpływu na plon i smak)
• Przedłużenie trwałości owoców cytrusowych na drzewach
• Zwiększenie plonu trzciny cukrowej
• Przyspieszenie hydrolizy skrobi w produkcji piwa
• Zastosowanie inhibitorów syntezy giberelin pozwala otrzymać
rośliny o krępym pokroju (lilie, chryzantemy, poinsecje) i
zapobieganiu wylegania (zboża)

Cytokininy –
regulatory podziałów
komórkowych
Występowanie cytokinin
CK
syntezowane są
głównie w merystemie
wierzchołkowym korzeni
, skąd są
transportowane przez ksylem do pędu.
CK syntetyzowane są również w młodych
organach rośliny tj. rozwijających się
zarodkach, liściach, owocach.

Aktywność biologiczna cytokinin
• Stymulują
podziały komórkowe (w kulturach in vitro)
• Stymulują fotosyntezę, syntezę chlorofilu
• Biorą udział w regulacji morfogenezy (przełamują
dominację wierzchołkową, powodują wybijanie pędów
bocznych)
• Aktywują transport asymilatów i rozwój liści
• Przyspieszają rozwój owoców
• Zwiększają odporność na patogeny
• Hamują
proces starzenia (liści)

Cytokininy
Stymulatory
procesów anabolicznych
we wczesnych fazach ontogenezy
oraz
Inhibitory
procesów katabolicznych
(degradacji) podczas końcowych faz
ontogenezy roślin.

Nadekspresja genów kodujących białka związane z
biosyntezą CK (ipt) (
wzrost zawartości CK
) hamują
starzenie liści tytoniu
CK
:
-
indukują proces fotosyntezy
-stymulują aktywność Rubisco
-stymulują syntezę chlorofilu
-stymulują rozwój wiązek
przewodzących
kontrola
rośliny zmodyfikowane

Praktyczne zastosowanie cytokinin
• W kulturach tkankowych in vitro (BA, BAP, KIN) pobudzają
podziały komórkowe, stymulują wyrastanie pędów
bocznych
• Kształtowanie pokroju rośliny, rozkrzewianie roślin,
zniesienie dominacji wierzchołkowej pędu głównego,
wybijanie pędów bocznych, hamują wydłużanie pędów
• Odmładzanie eksplantatow (in vitro), sadzonek roślin
sadowniczych
• Spowalnianie procesów starzenia roślin

Brasinosteroidy (BR) –
najaktywniejsze stymulatory
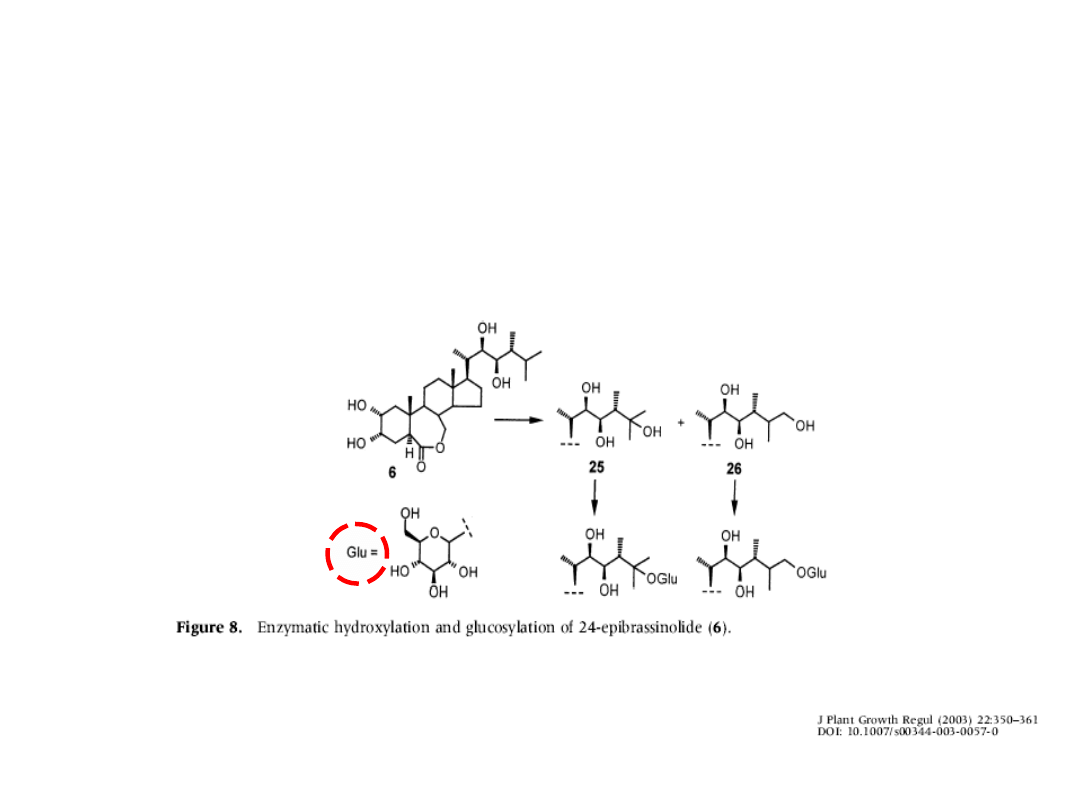
Metabolizm i transport BR
• Brasinolid może być przekształcany do nieaktywnych
koniugatów z glukozą np. 23-o-beta-D-gluko-piranozylo-
brasinolu

Produkcja i występowanie BR
• Brasinosteroidy są syntetyzowane w niemal
wszystkich organach rośliny
• Występują w bardzo małych ilościach
(np. w 1 kg nasion
Arabidopsis znajduje się 220 ng epibrasinolidu i 360 ng
kastasteronu)
• Wykazują aktywność fizjologiczną w stężeniach ok.
1000 razy mniejszych niż inne fitohormony
• BR są transportowane przez ksylem lub floem

Aktywność fizjologiczna BR
•Stymulują wzrost koleoptyli i pędów (najaktywniejsze
ze wszystkich hormonów roślinnych)
•Stymulują aktywność ATP-azy (wydzielanie H
+
do
apoplatu), co prowadzi do rozluźniania struktury
ściany komórkowej
•Stymulują podziały komórkowe
•Indukują proces różnicowania komórek mezofilu w
elementy ksylemu, pobudzają transport związków
pokarmowych
•Zwiększają tolerancję na stresy
•Przyspieszają starzenie liści
•Stymulują syntezę etylenu

BR zwiększają tolerancję na stresy
(wysokiej temperatury)
BR zwiększają odporność roślin (rzepak) na stresy
poprzez:
- stymulację syntezy białek HSP (70, 90,101),
- aktywację systemu antyoksydacyjnego oraz
- ograniczenie degradacji barwników
fotosyntetycznych
EBR -
epibrasinolid

Praktyczne znaczenie BR
• W kulturach tkankowych in vitro zastępują działanie
cytokinin
• Opryski środkami zawierającymi BR (BIOBRAS-16)
zwiększają plonowanie roślin i poprawiają wartości
odżywcze zbiorów (np. zwiększają zawartość cukru w
burakach cukrowych i owocach – w arbuzach, winogronach)
• Stosowane jako ukorzeniacze sadzonek.

HORMONY
ROŚLINNE
INHIBITORY
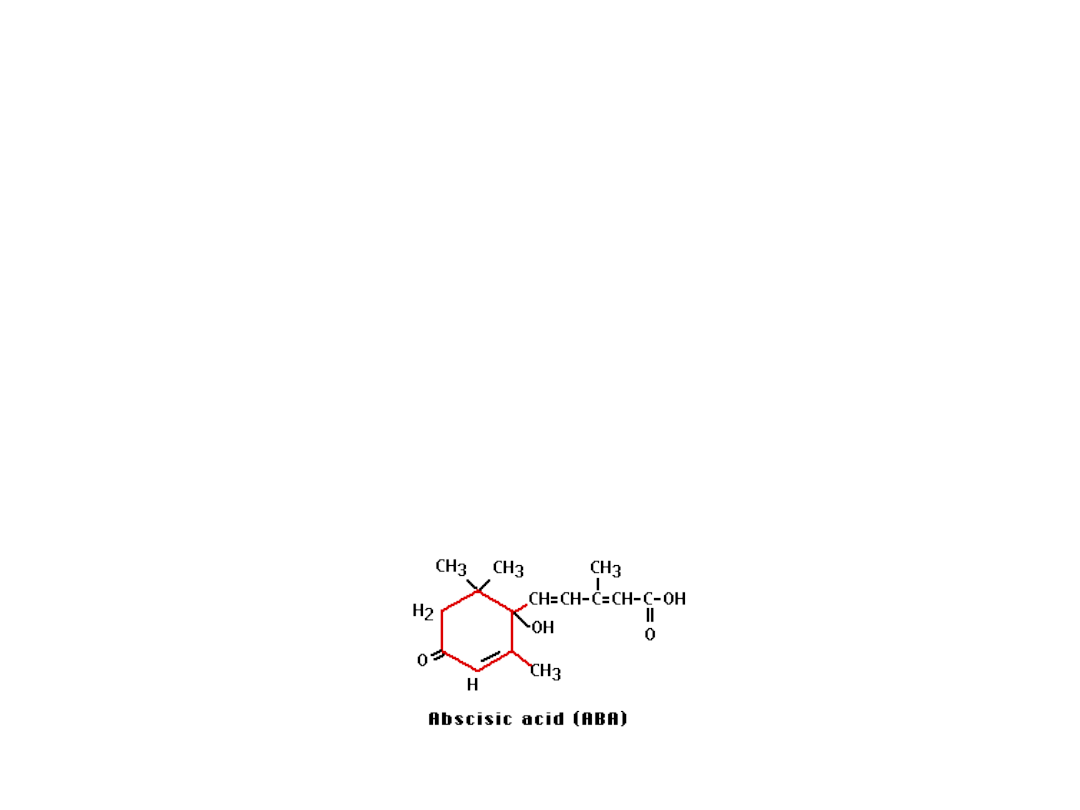
Kwas abscysynowy
–
hormon spoczynku
i
odporności na stresy

Produkcja i transport ABA
• ABA – jest produkowany przez wszystkie organy
rośliny:
liście, nasiona, korzenie i owoce
• jest transportowany przez ksylem i floem (głównie) w
formie niezdysocjowanej
• w formie zdysocjowanej (ABA
-
) łatwo
transportowany przez błony.

Aktywność biologiczna ABA
• hamuje
wzrost pędów (koleoptyli)
• hamuje kiełkowanie nasion
• indukuje stan spoczynku nasion, pąków
• stymuluje starzenie organów roślinnych
• indukuje zrzucanie liści, kwiatów i owoców (współdziałanie z
etylenem
w tworzeniu warstwy odcinającej)
w warunkach stresu
• stymuluje
zamykanie aparatów szparkowych
• hamuje fotosyntezę, syntezę chlorofilu, transport jonów przez
błony
• stymuluje ekspresję genów kodujących białka związane z
aklimatyzacją roślin
• zwiększa odporność roślin na stresy (susza, zalanie, zranienie)

Mechanizm działania ABA
Najlepiej poznane efekty ABA:
-
hamowanie kiełkowanie nasion
- udział w reakcji na stresy
(susza) –
zamykanie
aparatów szparkowych

Spoczynek i kiełkowanie nasion jest
regulowane przez hormony roślinne
Aktywacja zarodka
GA,
ETYLEN
ABA
Spoczynek nasion
– przewaga inhibitorów (
ABA
) nad stymulatorami
(
GA, etylen
)
Kiełkowanie nasion
– przewaga stymulatorów (
GA, etylen
) nad
inhibitora (
ABA
)

Porastanie zbóż
Zbyt niskie stężenie ABA
(brak spoczynku)
powoduje przedwczesne
kiełkowanie nasion na
roślinie matecznej, co
powoduje duże straty
plonu.
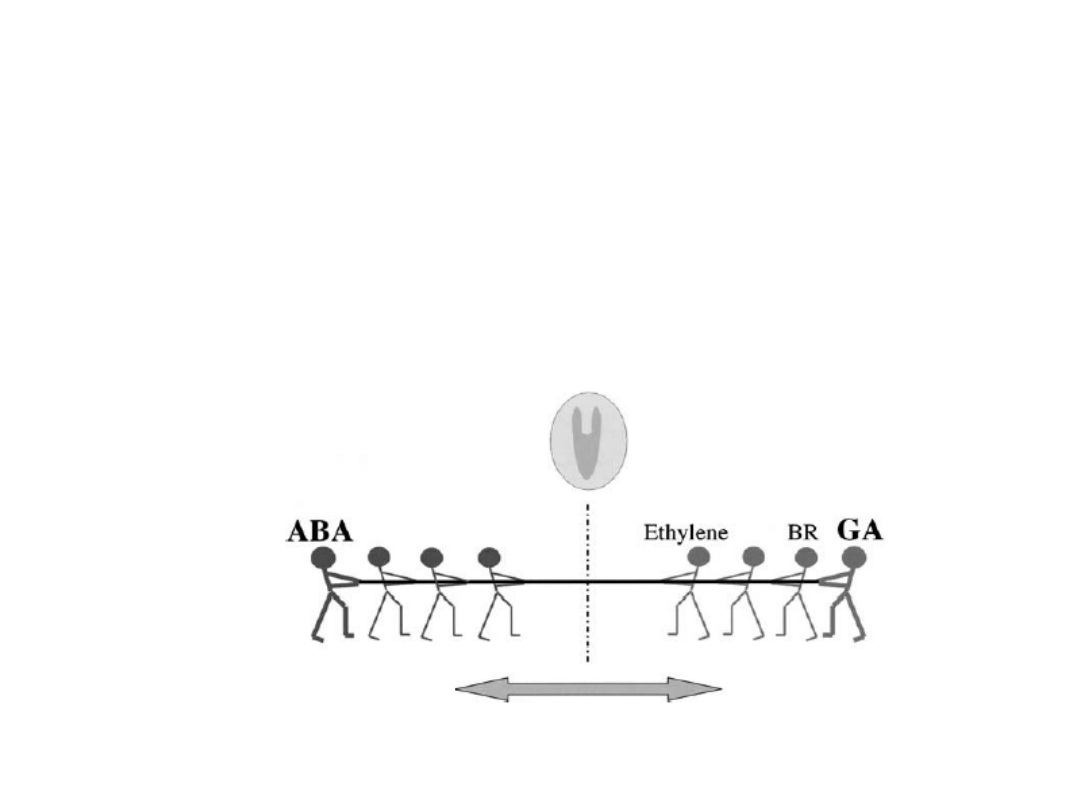
ABA
hormon o działaniu antagonistycznym do auksyn,
giberelin, cytokinin, etylenu, brasinosteroidów w
regulacji wielu procesów fizjologicznych m. in. spoczynku
nasion.
spoczynek
brak spoczynku
Etylen –
hormon dojrzewania,
starzenia
i reakcji na stres
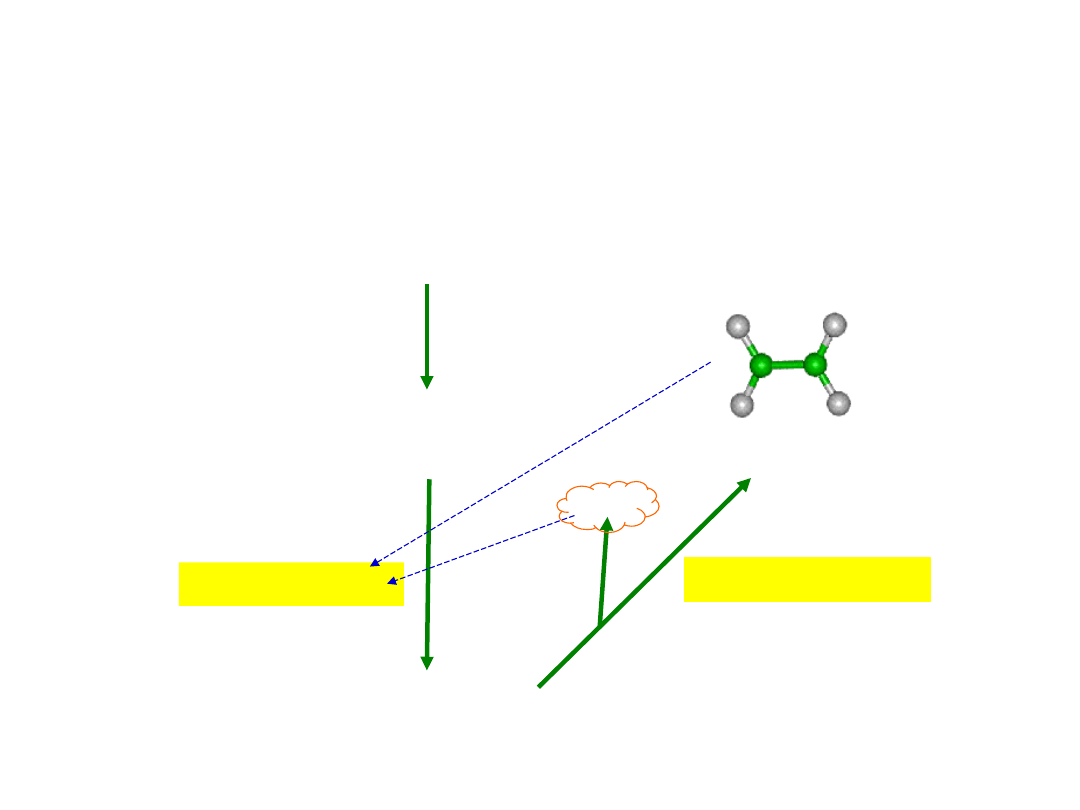
Synteza etylenu
(kwas 1-amino-cyklopropano-1-
karboksylowy)
syntaza ACC
Metionina
SAM
syntaza
SAM
Etylen C
2
H
4
ACC
Autostymulacja
syntezy etylenu przez
etylen lub/i HCN
(produkt uboczny
syntezy etylenu)
HC
N
oksydaza ACC

Produkcja etylenu
• Etylen jest produkowany przez
wszystkie części rośliny.
• Intensywność produkcji zależy od stanu fizjologicznego i typu
tkanki.
• Najbardziej aktywna produkcja etylenu:
- w tkankach merystematycznych
- dojrzewających owocach
- starzejących tkankach
- w tkankach poddanych działaniu warunków stresowych
(susza, chłód, zalanie, atak patogenu,
zranienie)
• Stymulatory syntezy etylenu: auksyny i cytokininy
(stymulują
ACS, ACO)

Aktywność fizjologiczna etylenu
• reguluje wzrost i rozwój siewek
• indukuje epinastie (ruchy roślin)
• przyspiesza dojrzewanie owoców
• stymuluje kiełkowanie i wzrost młodych siewek
• przyspiesza starzenie
• przyspiesza opadanie organów (liści, owoców)
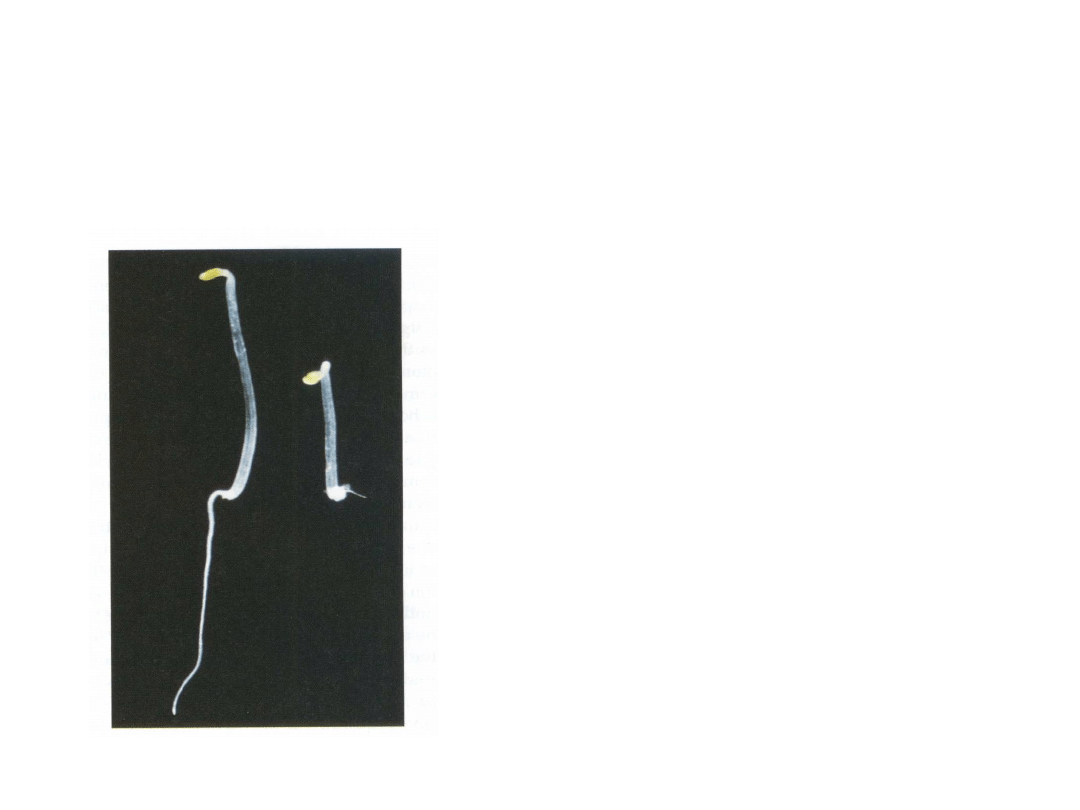
Etylen reguluje wzrost i rozwój siewek
• hamuje wzrost elongacyjny, indukuje wzrost na grubość
siewek rosnących w ciemności.
Kontrola
+ etylen
Efekty działania etylenu („potrójne”):
• skrócenie i pogrubienie hypokotyla
• zahamowanie wzrostu korzeni
• zwiększenie „kolanka”
podwierzchołkowego
pędu
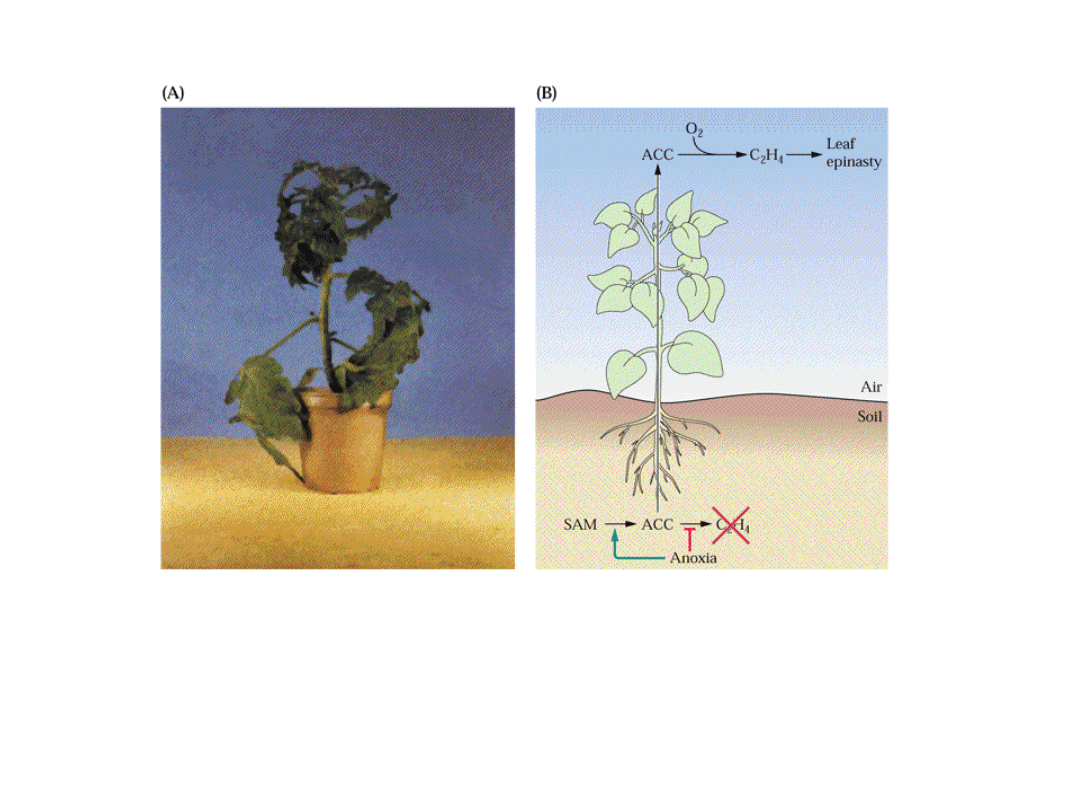
Etylen indukuje epinastie liści
Podczas anoksji lub zalania ACC syntetyzowane w korzeniach jest
transportowany przez ksylem do pędów i tam przekształcany do
etylenu (w glebie jest deficyt tlenu w skutek zalania, chłodu)
Etylen powoduje niesymetryczną dystrybucję auksyn w pędzie i
nierównomierny wzrost obu stron ogonka liściowego (efekt
„zwiędłych” liści).
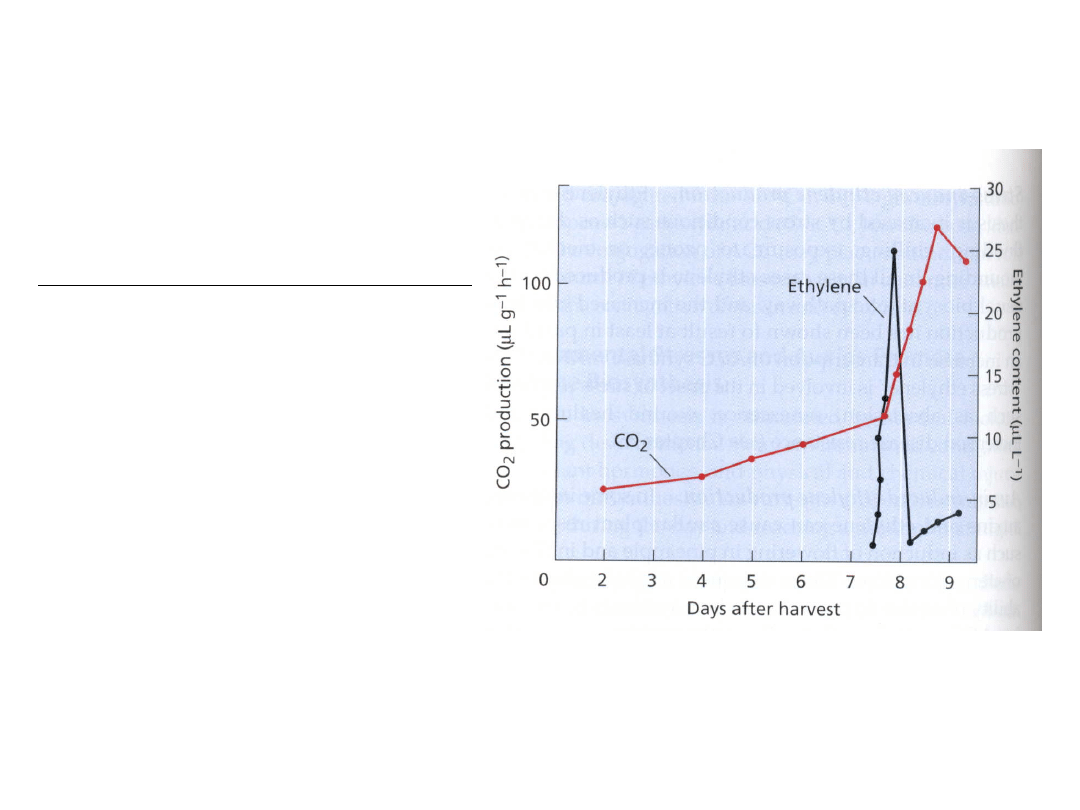
Etylen reguluje dojrzewanie owoców
klimakterycznych
O W O C E
klimakteryczne
nieklimakteryczne
jabłka
cytrusy
banany
winogrona
figi
ananasy
mango
truskawki
pomidory arbuzy
śliwki
brzoskwinie
Zmiany intensywności oddychania i
produkcji etylenu podczas dojrzewania
bananów
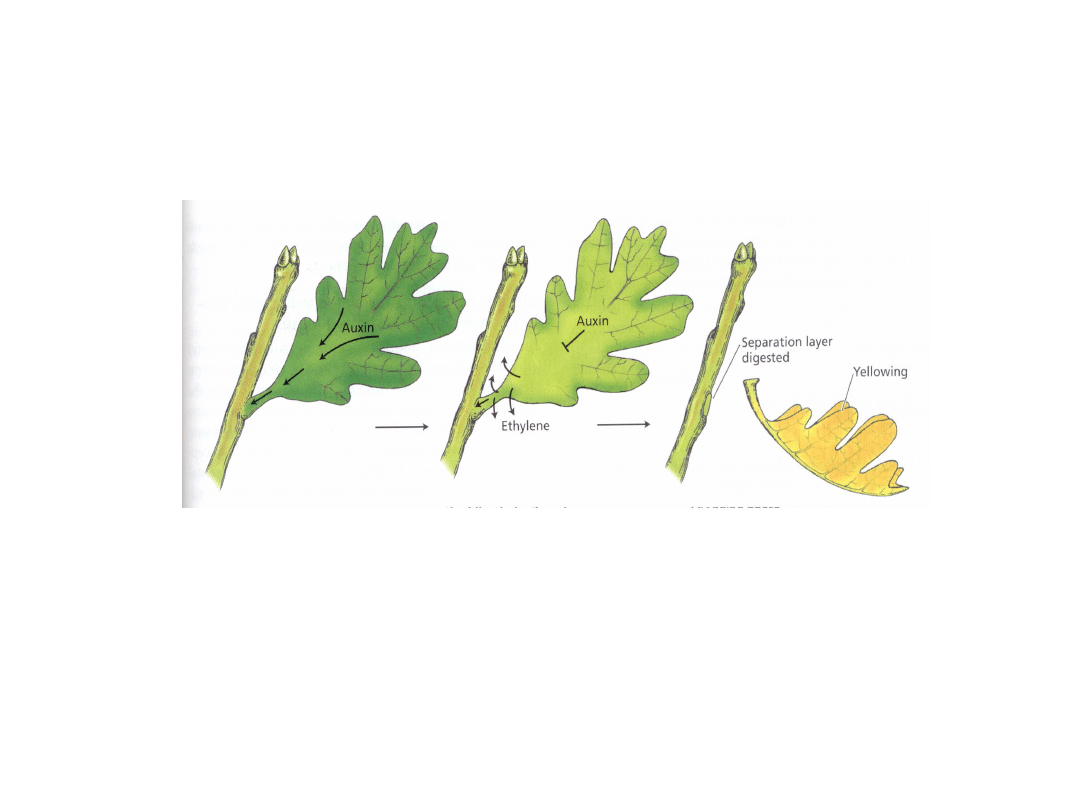
Etylen indukuje powstawanie warstwy
odcinającej
Wysokie stężenie
auksyn hamuje
syntezę etylenu
Spadek stężenia
auksyny
stymuluje
syntezę etylenu
Etylen stymuluje
degradację ścian
komórkowych przy
udziale enzymów
hydrolitycznych (celulazy,
poligalakturonazy)

Praktyczne znaczenie
etylenu
W praktyce stosowanych jest wiele
preparatów, które emitują etylen
(etefon, etrel) lub zawierają IAA i ACC w
celu:
- przyspieszania dojrzewanie owoców
-
synchronizacji zakwitanie
niektórych
roślin
(ananasów)
-
modelowania pokroju roślin
(skracanie,
promocja wzrostu na grubość)
-
ukorzeniana
odciętych fragmentów
pędów
Preparaty blokujące syntezę etylenu
opróżniają dojrzewanie owoców,
opadanie kwiatów, przedłużają trwałość
kwiatów ciętych.

Kwas jasmonowy (JA) –
hormon obronny roślin
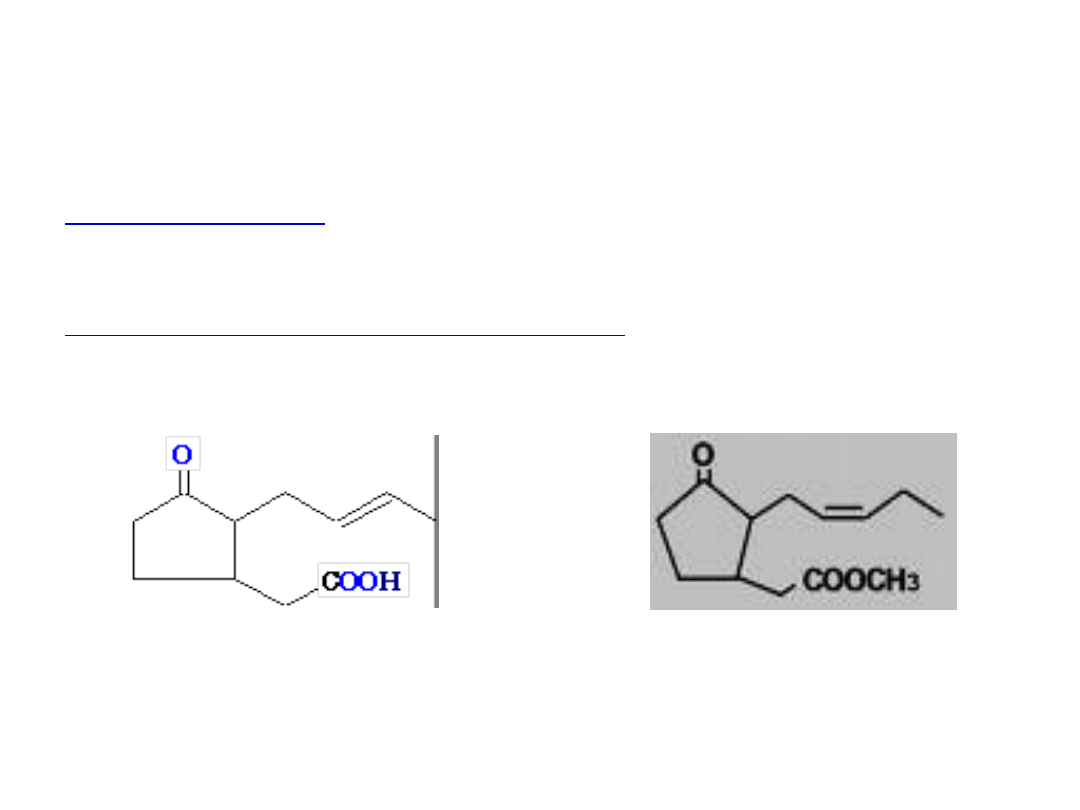
Budowa chemiczna JA
Kwas jasmonowy (JA) jest prostym, 12-węglowym związkiem,
podstawę budowy stanowi cyklopentenowy pierścień
Ester metylowy kwasu jasmonowego – JA-Me – jest lotnym
związkiem o podobnej aktywności biologicznej
JA
JA-Me
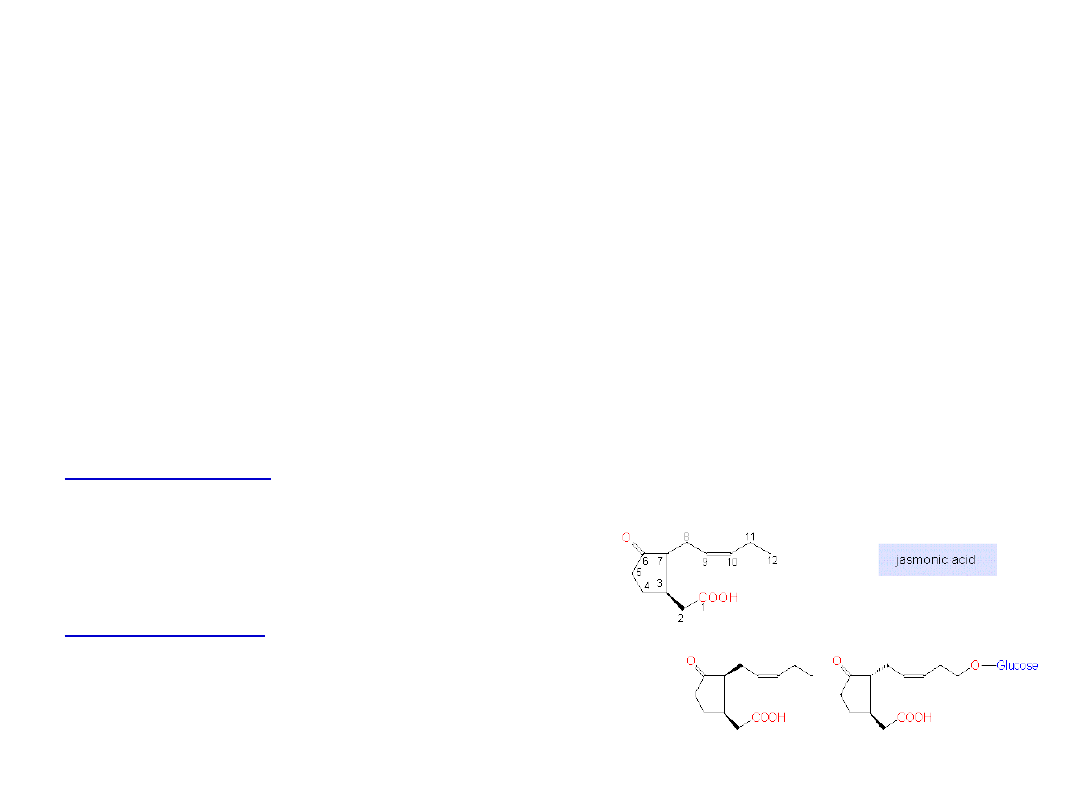
Występowanie JA
Występowanie JA i JA-Me wykazano niemal we wszystkich
grupach systematycznych roślin wyższych, paprociach,
mchach, grzybach i glonach
Występują w bardzo małych ilościach - od 0,1 do 5 mg / kg
ś.m. tkanki.
Koniugaty JA:
• z aminokwasami m. in. z leucyną, waliną, tyrozyną
• z cukrami m. in. z glukozą
Inhibitory JA:
aspiryna, salicylan, ibuprofen,

Aktywność biologiczna JA
Działa
podobnie do ABA
w trakcie ontogenezy rośliny
• Hamowanie wzrostu korzeni
• Hamowanie fotosyntezy i aktywności niektórych
enzymów chloroplastowych
* * *
• Stymuluje
procesy starzenia
• Stymuluje opadanie liści
• Stymuluje dojrzewanie owoców
• JA indukuje reakcje obronne przeciwko atakowi
patogenów, owadów, promieniowaniu UV, działaniu
ozonu, zranieniu
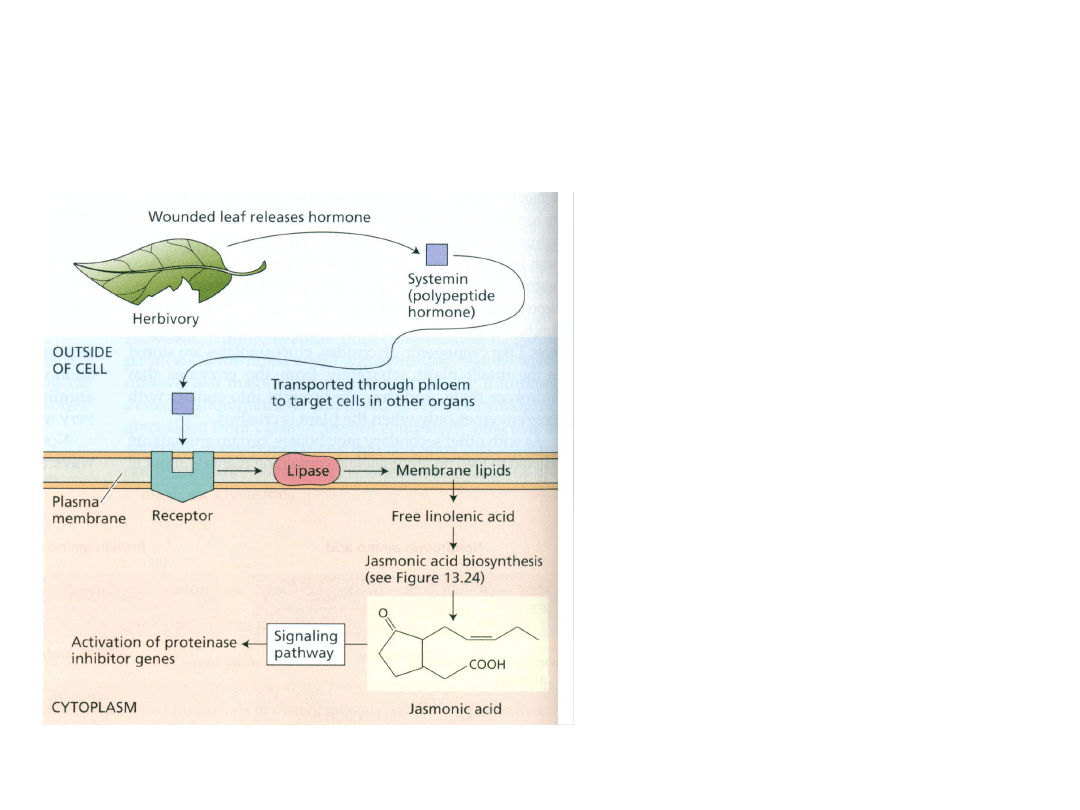
Udział JA w reakcji na stresy biotyczne
(atak patogenów)
Uszkodzenie rośliny przez owady
(zranienie) uwalnia systeminę
(18-aminokwasowy polipeptyd),
która łączy się z receptorem, co
aktywuje lipazę, uwalniającą z błon
kwas linolenowy – prekursor JA

Współdziałanie hormonów
cross-talk

Współdziałanie hormonów
• Plejotropowy charakter działania
hormonów polega na udziale poszczególnych
hormonów w regulacji wielu procesów
fizjologicznych.
• Decydująca jest odpowiednia równowaga
hormonalna
Współdziałanie hormonów –
synergistyczne lub antagonistyczne
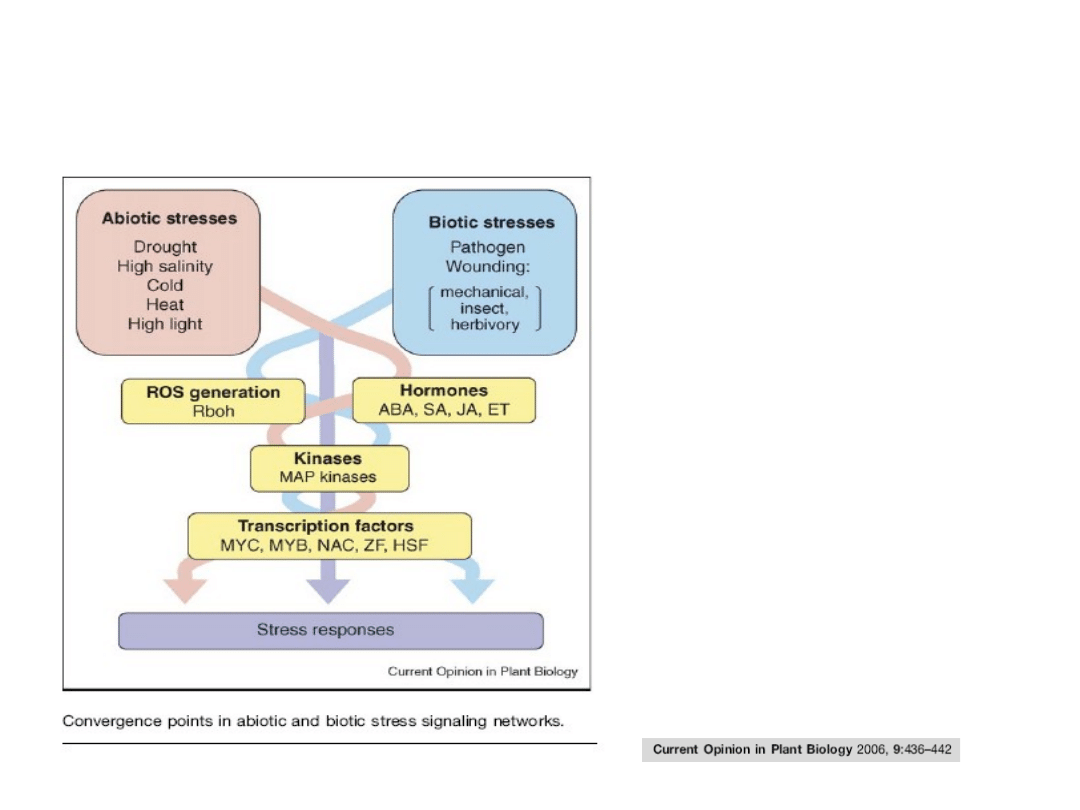
Współdziałanie szlaków transdukcji sygnałów („sieć”)
biorących udział w reakcji roślin na stresy biotyczne i
abiotyczne
Wspólne elementy szlaków
indukowanych przez różne
hormony:
• ROS,
• kinazy MAP,
• niektóre czynniki
transkrypcyjne.

Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
- Slide 38
- Slide 39
- Slide 40
- Slide 41
- Slide 42
- Slide 43
- Slide 44
- Slide 45
- Slide 46
- Slide 47
- Slide 48
- Slide 49
- Slide 50
- Slide 51
- Slide 52
- Slide 53
- Slide 54
- Slide 55
- Slide 56
- Slide 57
Wyszukiwarka
Podobne podstrony:
5 Hormony jako cz sygnalna skrócony
biol hormony
SPRAWDZ. BIOL - GIM IIBUDOWA I ROLA SKÓRY ORAZ UKŁADU HORMONALNEGO 2005r, sprawdziany, gim2
SPRAWDZ. BIOL - GIM II UKŁAD NERWOWY i HORMONALNY 2005r, sprawdziany, gim2
Biol kom cz 1
TEMAT 9 hormony a odporność
HORMONY NADNERCZY
W 8 Hormony 2010 2011
w4 skrócony
w3 skrócony
78 Hormony wysp trzustki
Biol Mol wyklad 9
Hormony kory nadnerczy 4
więcej podobnych podstron