136. Białka wiążące aktynę
Białka wiążące aktynę można podzielić na kilka typów pod kątem ich funkcji:
Białka wiążące G-aktynę i wpływające na polimeryzację: profilina ją promuje, tymozyna β4 hamuje (opisane wcześniej)
Białka tworzące zarodki nowych filamentów: forminy (opisane wcześniej)
białka rozcinające filamenty: np. żelsolina i kofilina. Służą degradacji niepotrzebnych filamentów, żeby udostępnić G-aktynę tam, gdzie jej trzeba. Łamią filamenty aktynowe wiążąc je w takiej konformacji, że puszczają wiązania między monomerami. Jednocześnie blokują + końce przed dodawaniem monomerów, co powoduje e takie filamenty są szybko degradowane. Ich aktywność jest hamowoana przez wiązanie z PIP2, kofilina jest dodatkowo sterowana przez fosforylację, a żelsolina aktywowana wzrostem stężenia jonów wapnia.
Białka blokujące jeden z końców filamentu (capping proteins): CapZ blokuje + koniec, tropomodulina w kompleksie z tropomiozyną - koniec. Blokują one zarówno dodawane jak i odpadanie polimerów, stabilizując filamenty. Takie ustabilizowane mikrofilamenty występują tam, gdzie aktyna ma tworzyć określone i niezmienne struktury np. pod błoną erytrocytu albo w sarkomerach mięśni. CapZ jet inhibowany przez wiązanie z PIP2
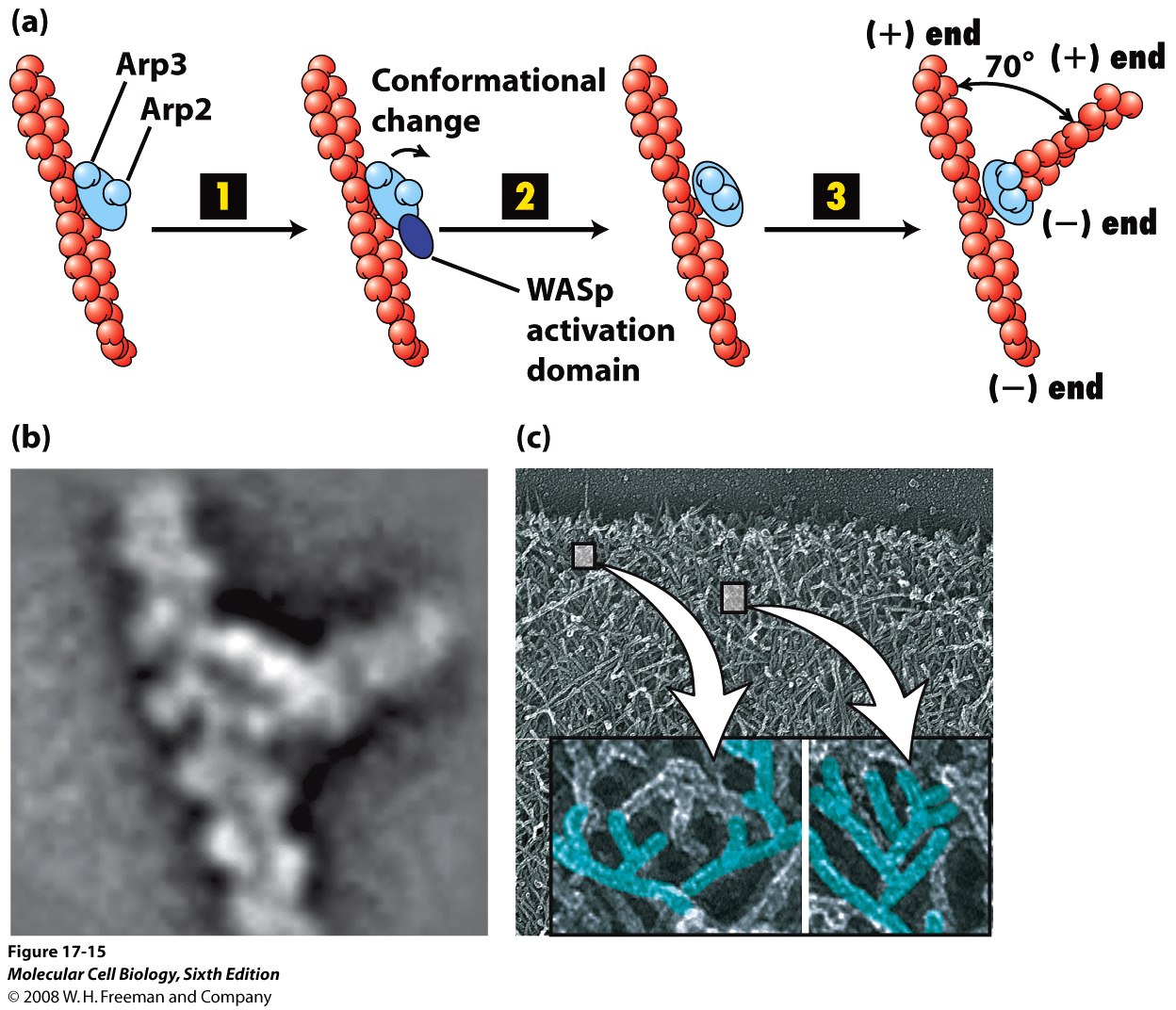
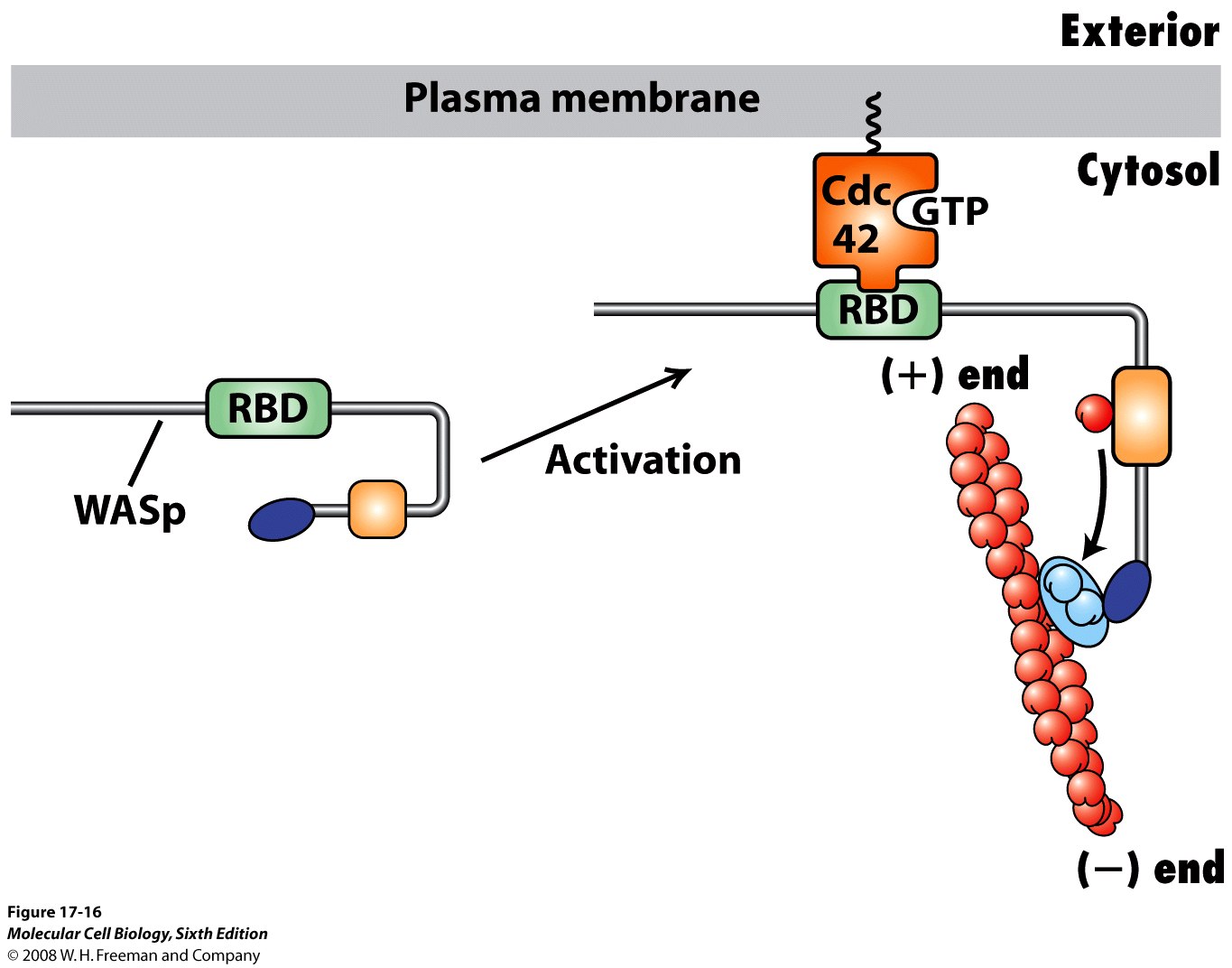
Arp 2/3 - kompleks, który wiąże się pod kątem 70 stopnie do boku filamentu aktynowego i powoduje powstanie nowego odgałęzienia. Jest to stymulowane przez białka WASp i kontrolowane przez Rho
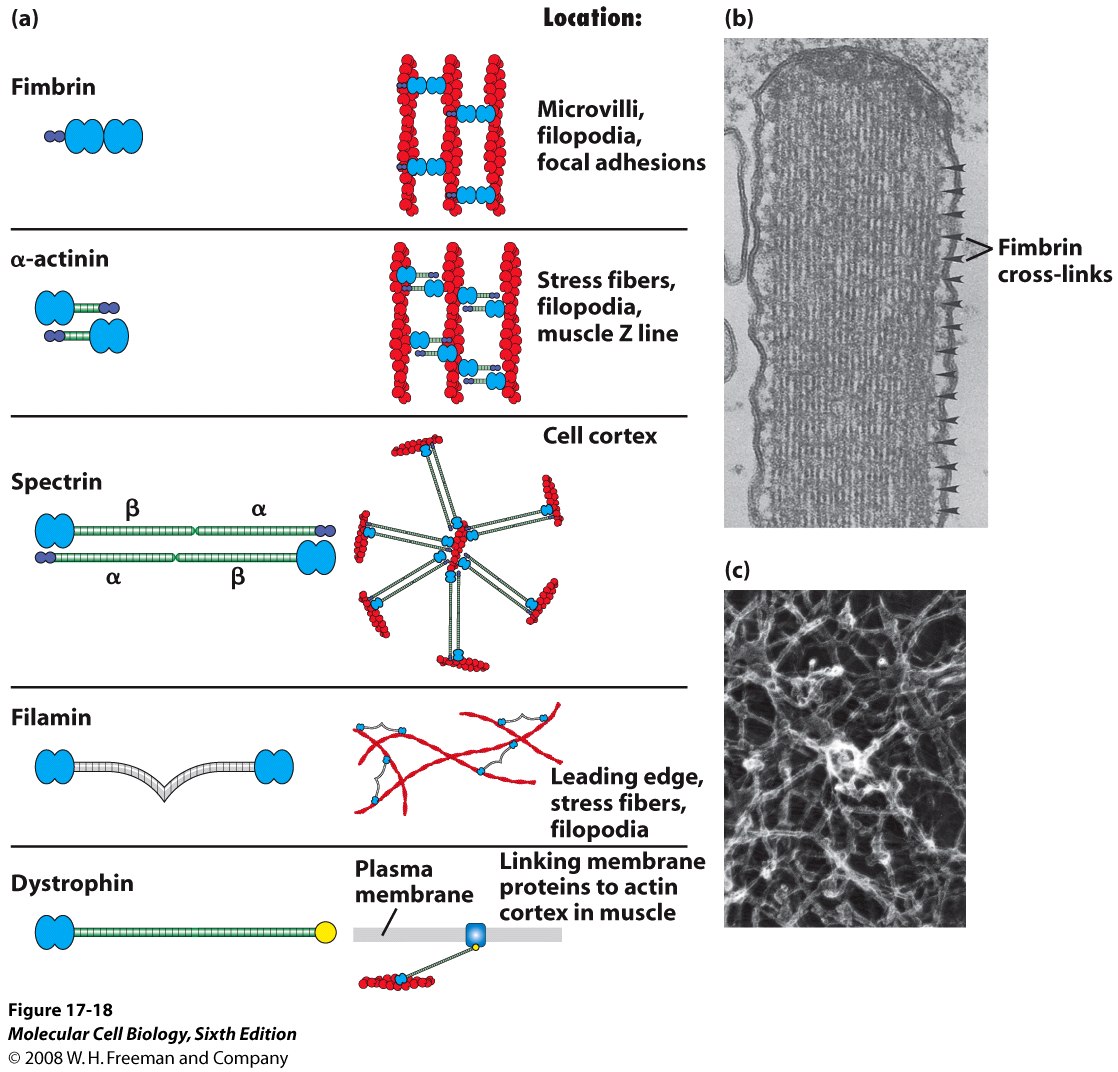
Jedno- lub wielopodjednostkowe białka łączące dwa lub więcej filamentów aktynowych (są w tabelce)
137. Tymozyna, profilina, kofilina
Wszystkie były opisane wcześniej, razem z obrazkami
138. Forminy i ARP2/3.
Forminy opisane wcześniej, z obrazkami. Do Arp należy jeszcze dodać, że rosnące odgałęzienia rosną plus końcem, Arp znaczy „actin-related protein” i wykazuje 50% homologii z aktyną. Aktywowane przez Rho (albo Rho-podobne białko) WASp powoduje zmianę konformacji Arp2/3 która daje nową gałąź.
Rozgałęzienia filamentów aktynowych pchają membranę i powodują zmianę kształtu oraz przesunięcie komórki.
139. Białka Rho.
Białka Rho to monomeryczne GTP-azy zakotwiczone w błonie resztami lipidowymi. W formie aktywnej, czyli związanej z GTP, aktywują one wiele czynników powodujących polimeryzację i rozgałęzianie mikrofilamentów. Białka Rho aktywują forminę, która jest odpowiedzialna za tworzenie się nowych filamenów, a także WASp, które aktywuje Arp2/3 powodując tworzenie rozgałęzień mikrofilamentów. Aktywność białka Rho jest kontrolowana przez błonowe receptory czynników wzrostu, cytokin itp. oraz przez inną GTP-azę - Rac.
(trochę to mało, ale nie wiem co tu dać)
140. Połączenia wiązek i włókien aktynowych z plazmolemmą.
Włókna aktynowe występują szczególnie obficie pod błoną komórkową. Aby zapewnić możliwość ruchu i zmiany kształtu komórce, aktyna musi być połączona z błoną. Takie połączenia służą też kotwiczeniu komórek w ECM, utrzymywaniu wypustek komórkowych (np. mikrokosmków) oraz biorą udział w procesie krzepnięcia krwi.
Jest kilka rodzajów takich połączeń:
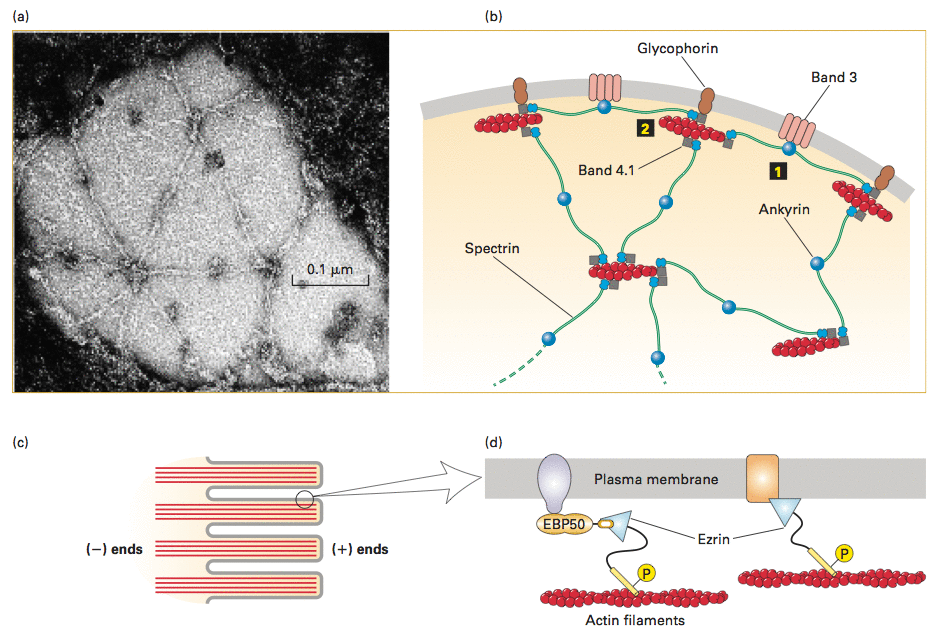
pod błoną ertytrocytów, a także niektórych innych komórek, aktyna tworzy krótkie 14-jednostkowe filamenty. Są one połączone przez ankirynę z BPA (białko przenoszące aniony, integralne białko błonowe) a przez spektrynę z innymi filamentami aktynowymi i z glikoforyną C, również transbłonowym białkiem. Każdy filament aktynowy jest połączony z sześcioma innymi przez spektrynę co daje heksagonalną sieć. Sieć ta zapewnia erytrocytom jednocześnie wytrzymałość mechaniczną i elastyczność.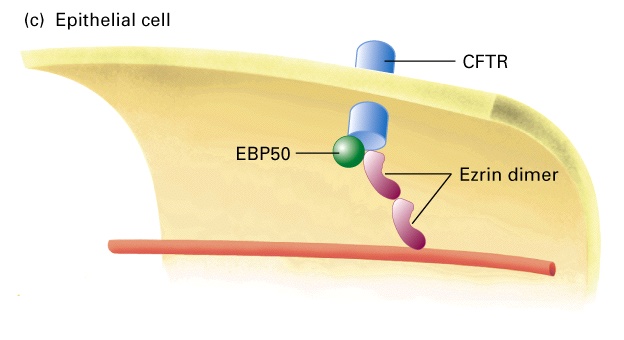
W komórkach nabłonka, np. w mikrokosmkach jelita, aktyna łączy się z dimerami ezryny, która przez EBP50 łączy się z białkami transbłonowymi - CFTR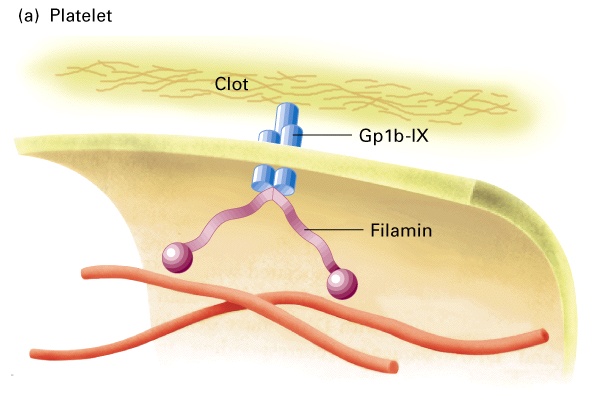
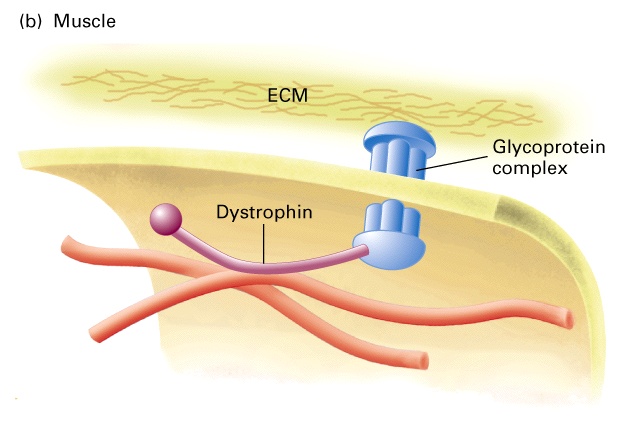
W płytkach krwi aktyna jest połączona przez filaminę do integralnych glikoprotein zwany Gp1b-IX, glikoproteiny te wiążą się ze skrzepemW komórkach mięśniowych włókna aktynowe łączą się przez dystrofinę z kompleksami błonowych glikoprotein, które kotwiczą komórkę w ECM.
141. Nuklepeotydy i alkaloidy wiążące się z aktyną.
Nukleotydy to ATP i ADP, ich rola była już opisana.
Alkaloidy mogące łączyć się z aktyną to np. falloidyna(^^) z muchomora sromotnikowego (Amanita phalloides). Stabilizuje ona F-aktynę do tego stopnia, że cała aktyna w komórce polimeryzuje i żadne procesy związane z ruchem nie są możliwe i komórka umiera.
Wyszukiwarka
Podobne podstrony:
136 -141, Materiały, Geologia, Geologia Historyczna
e str 136 141 fotograf
141 Future Perfect Continuous
Mahabharata Księga I (Adi Parva) str 73 136
135 136
140 141
139 141
136
141 145
136 Mr. Zoob - Mój jest ten kawałek podłogi, kwitki, kwitki - poziome
136, Prace z pedagogiki
136
136 137
Podstawy elektroniki str 101 141
135 136
K Wiech i J Kalmuk 126 136 allelopatia
jw pomal 2005
133 136
141 Przykłady pozycji opisów katalogowych IIIid 15704
więcej podobnych podstron