Zestaw 49.
Fosfatydylocholina - metabolizm.
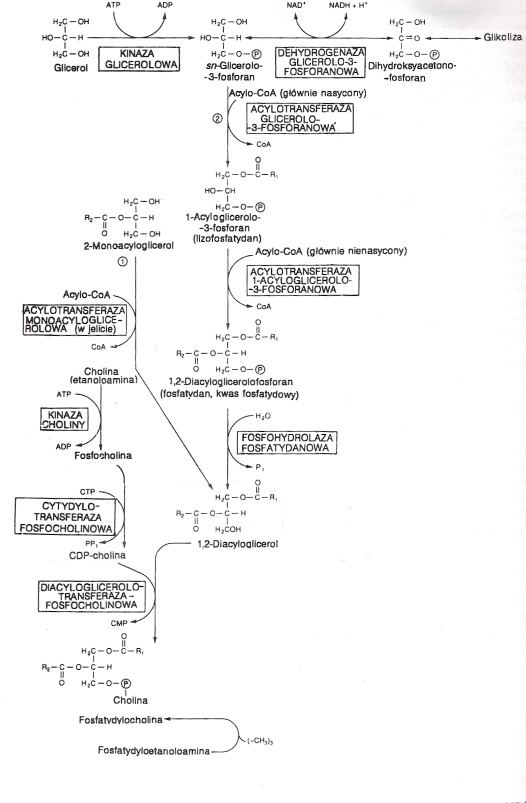
Fosfatydylocholina = lecytyna, fosfogliceryd zawierający cholinę. Są najczęściej występującymi składnikami błon lipidowych, stanowią dużą część zasobów choliny w organizmie (a ta odgrywa ważną rolę w przekaźnictwie nerwowym i jest zapasem grup metylowych). Dipalmitoilolecytyna jest składnikiem surfaktantu, zapobiegającego sklejaniu się płuc.
Lecytyny zazwyczaj mają resztę kwasu nasyconego w pozycji 1, a nienasyconego w poz. 2 glicerolu.
Biosynteza fosfatydylocholiny rozpoczyna się od 3-fosfoglicerolu, który może powstawać na dwóch drogach: albo poprzez bezpośrednią fosforylację glicerolu przy udziale kinazy glicerolowej i ATP albo, tam gdzie tego enzymu jest mało czy też ma małą aktywność, poprzez redukcję fosfodihydroksyacetonu pochodzącego z glikolizy przez enzym dehydrogenazę glicerolo-3-fosforanową i NADH + H+. Następnie acylotransferaza glicerolo-3-fosforanowa (powstaje lizofosfatydan) i acylotransferaza 1-acyloglicerolo-3-fosforanowa katalizują przeniesienie 2 cząsteczek acyloCoA na 3-fosfoglicerol, tworzy się fosfatydan. Fosfohydrolaza fosfatydanowa odczepia fosforan, dołącza wodę i tworzy się 1,2-diacyloglicerol. Ten z kolei łączy się ze zaktywowaną choliną (cholina jest fosforylowana do fosfocholiny przy udziale ATP, następnie reaguje z CTP, tworzy się cytydynodifosfocholina) za pomocą diacyloglicerolotransferazy fosfocholinowej. Tworzy się fosfatydylocholina.
Innym możliwym szlakiem biosyntezy, przebiegającym w wątrobie, jest metylacja fosfatydyloetanoloaminy przez S-adenozylometioninę.
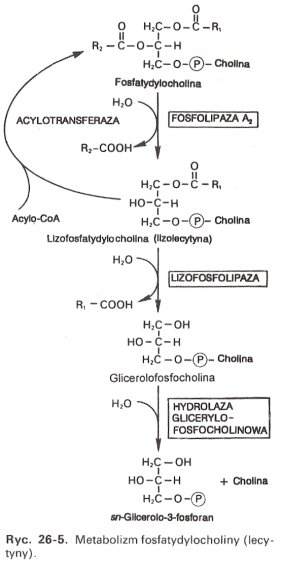
Degradacja do kwasów tłuszczowych, 3-fosfoglicerolu i choliny.
Wpierw następuje hydroliza lecytyny katalizowana przez fosfolipazę A2. Odczepia się kwas kwas tłuszczowy z pozycji 2, powstaje lizolecytyna. Lizolecytyna może slużyć albo do ponownej syntezy lecytyny (przyłącza się acylo-CoA w obecności acylotransferazy) albo jest dalej rozkładana. Następuje kolejna hydroliza, katalizowana przez lizofosfolipazę (fosfolipazę B), powstaje wolny kwas tłuszczowy i glicerolofosfocholina, ta dalej jest hydrolizowana, przez hydrolazę glicerolofosfocholinową, do 3-fosfoglicerolu i choliny.
Tworzenie lipoprotein osocza.
Wg schematu:
Acylotransferaza lecytyna:cholesterol
Lecytyna + Cholesterol --------------------------------> Lizolecytyna + Ester cholesterolowy Acylotransferaza lecytyna:cholesterol (LCAT), enzym syntetyzowany w wątrobie, występujący w osoczu, katalizuje przeniesienie reszty kw. tłuszczowego z poz. 2 lecytyny na cholesterol.
Tworzenie sfingomielin.
Ceramid + Fosfatydylocholina ------> Sfingomielina + Diacyloglicerol
Aminy biogenne.
Powstają w wyniku dekarboksylacji aminokwasów obojętnych i zasadowych.
katecholaminy (dopamina, noradrenalina, adrenalina)
Powstają z tyrozyny. W komórkach wątroby jest ona hydroksylowana przez monooksygenazę 3-tyrozynową. Powstaje 3,4-dihydroksyfenyloalanina (DOPA). Dekarboksylaza dopa, wymagająca fosforanu pirydoksalu, wytwarza dopaminę. Ta przemieszcza się czynnym transportem do pęcherzyków synaptycznych. Tam jest ponownie hydroksylowana przez β-oksydazę dopaminową (która potrzebuje miedzi i wit.C) do noradrenaliny (norepinefryny). Przekształcenie tej ostatniej w adrenalinę (epinefrynę) następuje w komórkach chromochłonnych rdzenia nadnerczy, gdyż tylko tam znajduje się enzym N-metylotransferaza fenyloetanoloaminowa, posługująca się S-adenozylometioniną w celu zmetylowania I-rzędowej gr. aminowej.
Inny szlak wiedzie od DOPA do syntezy melanin - barwników skóry i tęczówki
- dopamina jest odpowiedzialna za koordynację i napęd ruchowy (niedobór w ch. Parkinsona), procesy psychiczne i regulację uwalniania prolaktyny i gonadotropin.
- noradrenalina i adrenalina przyspieszają rytm serca, zwężają naczynia obwodowe, rozkurczają wieńcowe, rozkurczają oskrzela i źrenice, wzmagają przemianę glikogenu w glukozę. Noradrenalina jest metabolizowana przez katecholotlenometylotransferazę COMT do 3-metoksypochodnych oraz przez monoaminooksydazę MAO do aldehydu 3,4-dihydroksymigdałowego. Te metabolity są w końcowym efekcie utleniane do kwasów lub redukowane do alkoholi.
histamina
Pochodzi z histydyny. Dekarboksylacja jest katalizowana przez dwa różne enzymy:
dekarboksylazę L-aminokwasów aromatycznych, występującą w tkankach ssaków, jej substratami mogą być: dopa, 5-hydroksytryptofan, fenyloalanina, tyrozyna, tryptofan.
dekarboksylazę histydynową, występującą w większości tkanek, działa tylko na histydynę
Jest neuroprzekaźnikiem w OUN, a także wykazuje działanie prozapalne (poprzez kurczenie mieśni gładkich oskrzeli, jelit, macicy i dużych naczyń krwionośnych a rozszerzanie naczyń włosowatych i zwiększanie ich przepuszczalności, co powoduje obrzęk).
Jest metabolizowana przez N-metylotransferazę a następnie MAO.
serotonina
Pochodzi z tryptofanu, który najpierw jest hydroksylowany w pozycji 5. przez hydroksylazę tyrozynową wątroby do 5-hydroksytryptofanu. Ten związek jest dekarboksylowany do 5-hydroksytryptaminy (serotoniny) przez dekarboksylazę 5-hydroksytryptofanu.
Jest neuroprzekaźnikiem w OUN, zwęża naczynia krwionośne i kurczy mięśnie gładkie. Dostarcza również melatoniny (zachodzi równoczesna N-acetylacja grupy aminowej i O-metylacja grupy OH w poz.5), syntetyzowanej przez szyszynkę, odgrywającej rolę w utrzymaniu rytmów biologicznych, snu i czuwania. Wydzielanie melatoniny stymulowane jest przez ciemność i blokowane przez światło.
Serotonina metabolizowana jest przez niespecyficzną monoaminooksydazę MAO-A do aldehydu, który utlenia się do kwasu i jest wydalany z moczem.
GABA (kwas γ-aminomasłowy)
Powstaje z glutaminianu w reakcji katalizowanej przez dekarboksylazę L-glutaminianową, zależną od fosforanu pirydoksalu, w tkankach OUN, głównie substancji szarej. Może również powstawać z putrescyny poprzez deaminację lub N-acetylowane produkty pośrednie.
Neuroprzekaźnik w OUN, o działaniu hamującym.
Jest metabolizowany przez transaminazę GABA-T do semialdehydu bursztynowego.
inne aminy biogenne:
- kadaweryna powstaje z lizyny w procesach gnilnych białek, np. mięsa
- putrescyna powstaje z ornityny, j.w. należy do tzw. trupich jadów
- spermina i spermidyna powstają z ornityny, asoscjują z DNA i RNA, stymulują ich biosyntezę oraz razem z kadaweryną i putrescyną występują w plemnikach, broniąc ich DNA przed kwaśnym środowiskiem pochwy.
Metabolizm glukozy w warunkach tlenowych.
Glikoliza jest procesem zachodzącym w cytoplazmie wszystkich Prokaryota i Eucaryota. Substratem jest glukoza, produktami dwie cząsteczki pirogronianu, które w warunkach tlenowych przekształcane są do Acetylo-CoA wchodzącego do cyklu Krebsa lub służącego w syntezie kwasów tłuszczowych.
Fosforylacja glukozy katalizowana przez heksokinazę, która działa na anomery α i β i na inne heksozy (choć słabiej). W komórkach parenchymatycznych wątroby i w kom. wysepek trzustkowych tą reakcję katalizuje glukokinaza (wymagająca dużych stężeń glukozy co ma znaczenie po spożytym posiłku).
Dawcą fosforanu jest ATP, reagujący w formie kompleksu Mg2+-ATP. Reakcja jest praktycznie nieodwracalna, choć nie jest głównym miejscem regulacji procesu. Produkt - glukozo-6-fosforan wchodzi również do innych procesów: szlaku pentozofosforanowego, przemian glikogenu i in.
Izomeryzacja aldozowo-ketozowa G-6-P -> F-6-P
Enzym izomeraza fosfoheksozowa działa tylko na anomer α G-6-P.
Kolejna fosforylacja.
Reakcja w warunkach fizjologicznych nieodwracalna, główne miejsce regulacji procesu.
Rozszczepienie przez aldolazę na dwie fosfotriozy.
Aldolaza A w większości tkanek, aldolaza B w nerkach i wątrobie.
Izomeryzacja.
Do następnego etapu wchodzą więc dwie cząsteczki aldehydu 3-fosfoglicerynowego.
Utlenianie.
W obecności NAD+ i nieorganicznego fosforanu. Wśród produktów jest NADH, który w obecności tlenu w łańcuchu oddechowym prowadzi do powstania 3 cząsteczek ATP a w warunkach beztlenowych jest zużywany do syntezy mleczanu z pirogronianu.
Fosforylacja substratowa, przeniesienie fosforanu na ADP.
Powstają łącznie 2ATP, co rekompensuje 2ATP zużyte w reakcji 1 i 3.
Tą reakcję rozprzęga arsenian.
Przegrupowanie wewnątrzcząsteczkowe.
Przemieszczenie fosforanu z pozycji 3 na 2.
Dehydratacja.
Enolaza wymaga obecności Mg2+. Ten etap jest hamowany przez fluorki.
Kolejna fosforylacja substratowa, przeniesienie fosforanu na ADP.
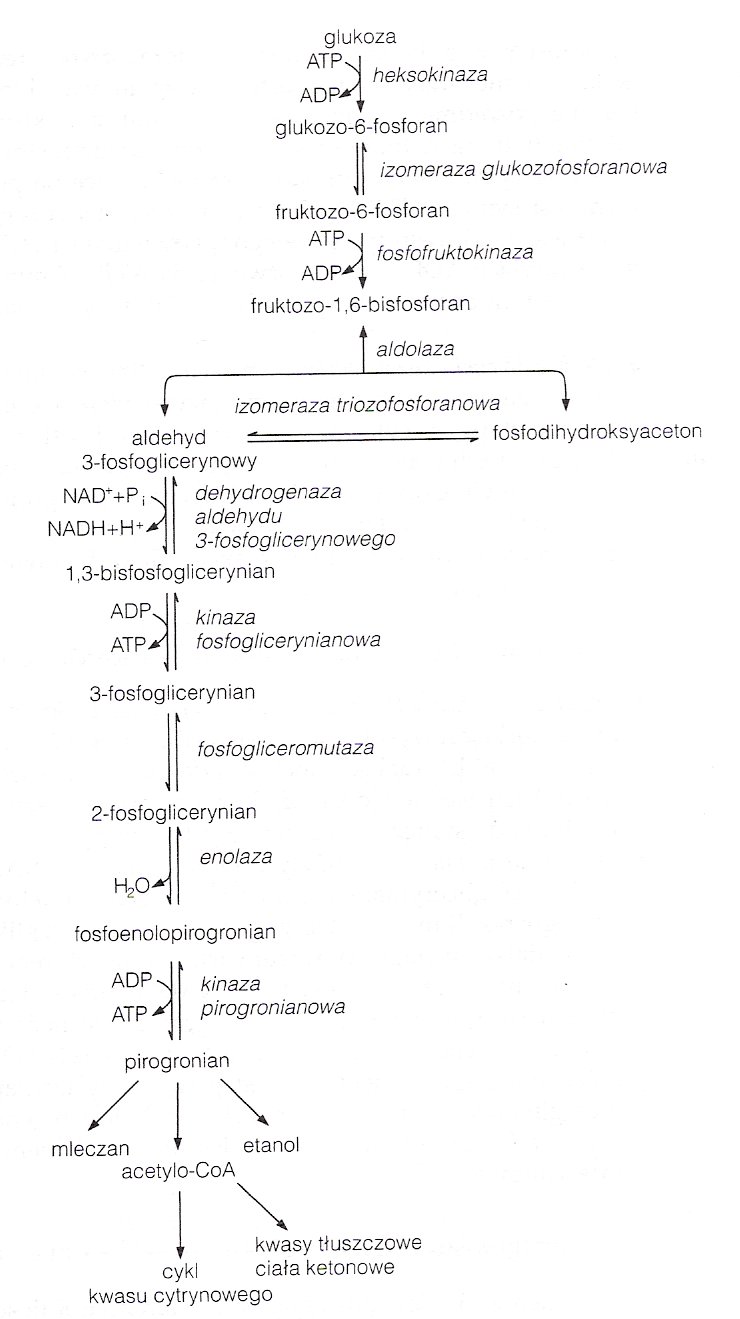
Reakcja praktycznie nieodwracalna. Produktem jest pirogronian i kolejne 2ATP. Ostatnie miejsce regulacji cyklu.
W warunkach tlenowych pirogronian wędruje do mitochondrium przez przenośnik pirogronianowy (symport z 1 protonem) i tam jest przekształcany do Acetylo-CoA w wyniku oksydacyjnej dekarboksylacji katalizowanej przez kompleks dehydrogenazy pirogronianowej (redukowany jest NAD+ do NADH i odszczepiany jest CO2). Pirogronian przy udziale dehydrogenazy pirogronianowej (pierwszego enzymu kompleksu) ulega dekarboksylacji do hydroksyetylowej pochodnej pierścienia tiazolowego difosfotiaminy. Ta reaguje z utlenionym lipoamidem (grupą prostetyczną drugiego enzymu - acetylotrasferazy dihydrolipoamidowej), oddając mu tą pochodną (tworzy się acetylolipoamid). Odtwarza się difosfotiamina. Następnie acetylolipoamid reaguje z koenzymem A, tworząc acetylo-CoA i zredukowany lipoamid. Trzeci enzym - dehydrogenaza dihydrolipoamidowa, którego koenzymem jest FAD - katalizuje utlenienie lipoamidu (redukuje sie FAD do FADH2). Acetylo-CoA wchodzi do cyklu Krebsa i tam ulega degradacji do CO2.
Powstałe w glikolizie równoważniki są przenoszone mostkami do mitochondrium (mostek glicerolo-3-fosforanowy i jabłczanowo-asparaginianowy) i tam włączane w łańcuch oddechowy.
1
Wyszukiwarka
Podobne podstrony:
Zestaw 54 Hanka Cywińska, materiały farmacja, Materiały 3 rok, Od Ani, biochemia, biochemia, opracow
Zestaw 51 Hanka Cywińska, materiały farmacja, Materiały 3 rok, Od Ani, biochemia, biochemia, opracow
Zestaw 50 Hanka Cywińska, materiały farmacja, Materiały 3 rok, Od Ani, biochemia, biochemia, opracow
Zestaw 52 Hanka Cywińska, materiały farmacja, Materiały 3 rok, Od Ani, biochemia, biochemia, opracow
Zestaw 53 Hanka Cywińska, materiały farmacja, Materiały 3 rok, Od Ani, biochemia, biochemia, opracow
ZESTAW 9 Monika Cywińska
ZESTAW 12 Monika Cywińska
ZESTAW 11 Monika Cywińska
ZESTAW 10 Monika Cywińska
ZESTAW 13 Monika Cywińska
ZESTAW 14 Monika Cywińska
zestaw nr 2
zestaw nr 3 (2)
Zestawy
zestaw 1 3
zestaw di 3 05
zestaw02rozw
zestaw7
więcej podobnych podstron