faza ciemna C4-przebieg 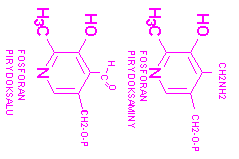
Fotosynteza C4 to proces fotosyntezy, wiązania dwutlenku węgla u roślin określanych nazwą rośliny C4. Proces wiązania CO2 przebiega w komórkach mezofilowych, gdzie dwutlenek węgla przyłączany jest do fosfoenolopirogronianu. W reakcji tej powstaje związek czterowęglowy - szczawiooctan. Jest on w zależności od gatunku rośliny przekształcany do asparaginianu lub jabłczanu i w tej postaci przenoszony do komórek pochew okołowiązkowych. Tam zachodzi reakcja dekarboksylacji i wydzielenie CO2, która jest włączany do cyklu Calvina-Bensona. Brak cyklu Calviana-Bensona w komórkach mezofilowych związany jest z brakiem enzymu, przyłączającego CO2 do cząsteczki rybulozo-1,5-bisfosforanu (RuBP,) określanego nazwą rubisco (karboksylaza oksygenaza rybulozo-1,5-bisfosforanu). Enzym ten może katalizować także reakcję przyłączenia do RuBP tlenu. Proces ten nosi nazwę fotooddychania i obniża on wydajność fotosyntezy roślin C3. Tlen i dwutlenek węgla konkurują o centrum aktywne enzymu rubisco. Dzięki zwiększonemu stężenie CO2 w komórkach pochew okołowiązkowych proces fotooddychania jest zahamowany,a tym samym wydajność fotosyntezy roślin C4 jest wyższa niż roślin C3. Rośliny Proces fotosyntezy u roślin C4 przebiega wydajniej, jednak nakład energetyczny na związanie jednej cząsteczki CO2 jest większy niż u roślin C3. Rośliny C4 podzielono na trzy podtypy: podtyp NADP-ME, podtyp NAD-ME i podtyp PEP-CK. Podstawą wydzielenia trzech podtypów jest enzym odpowiedzialny za przeprowadzenie reakcji dekarboksylacji w komórkach pochew okołowiązkowych. Jest to odpowiednio: enzym jabłczanowy zależny od NADP, enzym jabłczanowy zależny od NAD i karboksykinaza fosfoenolopirogronianu. Do roślin C4 należą gatunki z wielu rodzin np.: kukurydza, trzcina cukrowa, proso zwyczajne, sorgo, proso olbrzymie. Są to rośliny pochodzące z klimatu zwrotnikowego. Proces fotosyntezy u roślin C4 przebiega wydajniej, jednak nakład energetyczny na związanie jednej cząsteczki CO2 jest większy niż u roślin C3.
2. fosforan pirydoksolu- wzór, przykład reakcji z udziałem koenzymu
witamina B6 ma trzy formy: pirydoksynę, pirydoksamine, pirydoksal. Koenzymem B6 jest fosforan pirydoksaminy i pirydoksalu, które współdziałają z enzymami w przemianach aminokwasów i uczestniczą w: RACEMIZACJI OPTYCZNIE CZYNNYCH AMINOKWASÓW, TRANSMINACJI POMIĘDZY AMINOKWASEM I 2-OKSOKWASEM, DEKARBOKSYLACJI AMINOKWASÓW, ELIMINACJI TYPU 3,4 (ODWODNIENIE DERYNY DO PIROGRONIANU) SYNTEZA TRYPTOFANU Z SERYNY I INDOLU, FOSFOROLIZA SACHARYDÓW ZŁOŻONYCH reakcje zachodzą dzięki aktywności grupy aldehydowej 5-fosforanu pirydoksalu. Łatwo zachodzą w obrębie aminokwasów, które z tą grupą tworzą zasadę Schiffa o bardzo nietrwałym układzie elektronów. Sprzyja temu grupa OH, które chelatuje jony miedzi i glinu. Odmienna funkcję wykazuje on we współdziałaniu z fosforylazą glikogenową, do działania której jest niezbędny jako kofaktor. Funkcją fosforanu pirydoksalu jest reakcja transaminacji, kóra może przebiegac bez udziału białka enzymowego, a jedynie w obecności koenzymu i dwuwartościowego jonu metalu.
3. mechanizm działania koenzymów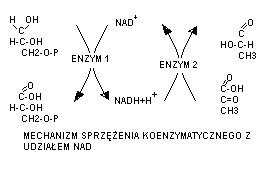
Dzialanie koenzymow polega na ich powiązaniu stechiometrycznym z substratem za pośrednictwem określonej jego grupy oraz z bialkiem enzymowym. W reakcji przeniesienia protonów i elektronów z jednego związku na inny może uczestniczyc jeden lub dwa enzymy; w drugim przypadku nastepuje tzw. Sprzężenie koenzymatyczne. Przykładem takiego sprzeżenia jest powiazanie z udzialem dinukleotydu nikotynoamidoadeninowego.
Enzym 1 (dehydrogenaza fosforanu gliceraldehydu, katalizuje przeniesienie atomow wodoru z wodzianu fosforanu triozy na NAD+, przy czym tworzy się 3-fosfoglicerynian i zredukowany NAD+. Reakcja jest sprzężona z druga w której regeneruje się NAD+ przez przekazanie atomów wodoru na pirogronian z udziałem innego enzymu- dehydrogenazy mleczajowej, w wyniku czego tworzy się mleczan. W tym przypadku koenzym jest lluxno powiązany z bialkiem enzymu i może współdziałać z róznymi apoenzymami. W innych reakcjach koenzym jest silniej powiązany z apoenzymem i wtedy jest on pośrednikiem w przekazaniu określonej grupy atomów z jednego substratu na drugi przy udziale tego samego enzymu.
Reakcja utlenienia β-D-glukopiranozy do laktonu kwasu glukonowego, katalizowana przez oksydazę glukozową, współdziałającą z FAD. Atomy wodoru odłączone z glukozy są tu przekazane za pośrednictwem FAD, współdziałającego z białkiem enzymu, na tlen cząsteczkowy, w wyniku czego tworzy się nadtlenek wodoru H2O2.
4. prebieg i znaczenie cyklu kwasu cytrynowego
cykl Krebsa, cykl kwasów trójkarboksylowych) zachodzi w macierzy mitochondrialnej. Polega na utlenieniu cząsteczki acetylokoenzymu A do dwóch cząsteczek dwutlenku węgla. Wytwarzana energia jest magazynowana w wysokoenergetycznych wiązaniach cząsteczek ATP, natomiast uwolnione atomy wodoru są przenoszone przez koenzymy NAD i FAD na enzymy łańcucha oddechowego. Cykl kwasu cytrynowego jest pośrednim etapem katabolizmu wielu związków chemicznych utlenianych w komórce.

Cykl rozpoczyna się od kondesacji szczawiooctanu z acetylo-CoA , w wyniku czego powstaje cytrynian, a następnie w drodze izomeryzacji jego powstaje izocytrynian. W następnej kolejności następuje dekarboksylacja izocytrynianu do a- ketoglutaranu, a ten po przez oksydacyjną dekarboksylację daje bursztynylo-CoA. Wiązanie tioestrowe w bursztynylo-CoA zostaje rozerwane przez fosforan, powstaje bursztynian i 1 cząsteczka GTP. W wyniku utleniania bursztynianu powstaje fumaran, który po uwodnieniu daje jabłaczan. Cykl kończy regeneracja szczawiooctanu, powstałego w wyniku utleniania jabłaczanu. Bilans energetyczny cyklu kwasu cytrynowego stanowi 10 wysokoenergetycznych wiązań fosforanowych powstałych z całkowitego utleniania każdego fragmentu dwuwęglowego do CO2 i H2O
5. Omów na dowolnym przykładzie proces syntezy kwasu tłuszczowego
Biosynteza kwasów tłuszczowych odbywa się w cytoplazmie komórek tłuszczowych. Do procesu potrzebny jest acetylokoenzym A. W biosyntezie kwasów tłuszczowych wyróżnić można kilka etapów:
1. acetylokoenzym A ulega karboksylacji do malonylokoenzymu A (malonylo-CoA).
2. Synteza kwasów tłuszczowych na kompleksie enzymatycznym - syntetazie kwasów tłuszczowych. W skład syntetazy wchodzi ACP, ktrory przenosi acyle. ACP zawiera z kolei panteteinę Tworzony kwas tłuszczowy jest związany kowalencyjnie z enzymem. reszta acetylowa jest przeniesiona na ACP. Następnie dochodzi do połączenia reszty malonylowej z resztą acetylową, powstaje 4-węglowa cząsteczka acetoacetylo-S-ACP. Wydzielony wówczas jest CO2 i HS-ACP. CO2 jest uwolniony w wyniku działania syntazy 3-oksoacylo-ACP.
3. Etap redukcji odbywa się przy udziale NADPH i reduktazy 3-oksoacylo-ACP. Dochodzi do redukcji grupy -okso. Powstaje reszta beta-hydroksyacylowa, która poddana jest dehydratacji przy udziale dehydratazy 3-hydroksy-ACP. Powstaje reszta alfa, beta-dehydro-acylowa, która zostaje poddana redukcji przy udziale NADPH i reduktazy enoilo-ACP. Powstaje 4-węglowy rodnik butyrylowy. W następnym obrocie reakcji powstaje kolejna jego cząsteczka, przy czym reszta acylowa z wcześniej wytworzonego rodnika jest przeniesiona na tę następna, wydzielony jest wówczas dwutlenek węgla i powstaje reszta beta-ketoacylowa. Ta znów podlega kondensacji z kolejną. W ten sposób tworzony kwas ulega wydłużaniu do odpowiedniej masy.
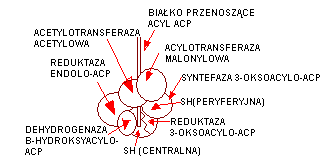
4. Uwalnianie gotowego łańcucha kwasu tłuszczowego odbywa się przy pomocy deacylazy. Odłącza ona kwas od HS-ACP,
CH3CO ~ S-CoA + 7HOOC-CH2CO~S-CoA + 14 NADPH + 14 H+ CH3(CH2)14CO~S-CoA + 7 CO2 + 7 HS-CoA + 14 NADP+ + 6H2O
Powstałe kwasy tłuszczowe są gromadzone w komórkach w postaci estrów glicerolu.
MODEL SYNTEZY KW. TŁUSZCZOWYCH
6. proces dekarboksylacji aminokwasow, rola otrzymanych produktów
Reakcje dekarboksylacji aminokwasów polegają na rozerwaniu wiązania między grupą karboksylową -COOH i resztą cząsteczki aminokwasu, w wyniku czego wydziela się CO2 i powstaje odpowiednia amina. Reakcję katalizują dekarboksylazy aminokwasowe. Dekarboksylacja aminokwasowa jest źródłem amin biogennych - substancji o dużej aktywności fizjologicznej, np. histamina (po dekarboksylacji histydyny), tyramina (po dekarboksylacji tyroksyny), tryptamina (po dekarboksylacji tryptofanu), serotonina = 5-hydroksytryptamina (po dekarboksylacji 5-hydroksytryptofanu). W Wyniku dekarboksylacji niektórych aminokwasów tworzą się ważne części składowe koenzymów, np. 2-propanolamina (składnik koenzymu B12), cysteamina (składnik koenzymu A). Z kwasu glutaminowego powstaje kwas gamma-aminomasłowy.
W wyniku dekarboksylacji aminokwasów obojętnych i zasadowych powstają aminy biogenne, które z wyjątkiem histaminy zwężają naczynia krwionośne i podwyższają ciśnienie krwi. Produkt dekarboksylacji histydyny - histamina rozszerza naczynia krwionośne, powodując spadek ciśnienia krwi oraz wywołując objawy alergiczne. Histamina jest stosowana do wydzielania soku żołądkowego w próbach czynnościowych. Natomiast produktem dekarboksylacji 5-hydroksytryptofanu jest serotonina. Serotonina zwęża naczynia krwionośne, powodując zwiększenie ciśnienia krwi, wywołując skurcz mięśni gładkich. Działa pobudzająco na ośrodkowy układ nerwowy.
5-hydroksytryptofan Serotonina
Rys. 16 Dekarboksylacja aminokwasów
Aminy biogenne można podzielić na aminy alifatyczne (monoaminy i poliaminy), katecholowe (fenolowe, o pierścieniach aromatycznych) i heterocykliczne. Aminy katecholowe (tyramina, dyzamina, adrenalina), imidazolowe (histamina) i indolowe (tryptamina, serotonina) powodują zmiany ciśnienia krwi. Aminy te są też neuroprzekaźnikami, odgrywają ważną role w przenoszeniu impulsów nerwowych przez zakończenia nerwowe (noradrenalina, a jako inhibitor dopamina) oraz poprzez synapsy komórek nerwowych (serotonina). Nadmiar serotoniny w krwi powoduje stany halucynacyjne, a niedobór depresje.
7.rola tranclacjii w biosyntezie bialek Translacja - to w biologii proces syntezy łańcucha polipeptydowego białek na matrycy mRNA. Proces ten katalizowany jest przez rybosomy. Translacja polega na interpretacji informacji zawartej w kolejności ułożenia nukleotydów mRNA, zgodnie z zasadami kodu genetycznego, na kolejność ułożenia reszt aminokwasowych w białku. Translacja odbywa się w kierunku od 5' do 3' mRNA, a syntetyzowane białko powstaje od końca aminowego do karboksylowego. Proces składa się z trzech etapów: inicjacji, elongacji i terminacji. Zakończenie odbywa się gdy w mRNA zostanie napotkany nonsensowny (stop) kodon nie odpowiadający żadnemu tRNA.
Etap translacji (odczytania, przetłumaczenia)- zachodzi w rybosomach.- biorą w nim udział cząsteczki tRNA. - aminokwas łącząc się z cząsteczką tRNA ulega uaktywnieniu:
tRNA + aminokwas = aminoacylo tRNA
- cząsteczki mRNA umiejscawiają się między dwiema podjednostkami rybosomu.- cząsteczki tRNA dostają się do rybosomu i jeśli maja antykodon komplementarny do kodonu w mRNA to zostawiają tam transportowany aminokwas.- między aminokwasami powstają wiązania peptydowi, rybosom przesuwa się wzdłuż nici mRNA, a łańcuch peptydowy wydłuża się.- translacja zaczyna się kodonem startowym- metionina (AUG), a kończy kodonem nonsensownym (UAA, UAG, ULG).- translacja wymaga: udziału rybosomów, mRNA, dostatecznej ilości aminokwasów, energii, tRNA. translacja jest zatem kluczowym procesem warunkujacym zycie organizmow
8. Omow znaczenie w przemianach metabolicznycha: jonu amonowego
Cykl ornitynowy, zwany też cyklem mocznikowym lub mocznikowym cyklem Krebsa to cykl przemian biochemicznych (reakcji enzymatycznych) trzech aminokwasów: ornityny, cytruliny i argininy. Do cyklu wprowadzany jest karbamoilofosforan powstały z amoniaku i dwutlenku węgla. Cykl przebiega w mitochondriach i cytoplazmie komórek wątroby (hepatocytów) i wymaga dostarczenia energii w postaci ATP, a jego produktem końcowym jest mocznik.
Reakcje zużywania aminokwasów rozpoczynają się od odłączenia grupy aminowej w procesie deaminacji. Produktami tych reakcji są łańcuchy węglowe (tzw. ketokwasy) oraz jon amonowy. W komórkach człowieka łańcuchy węglowe po przemianach mogą być włączone w glikolizę lub do cyklu Krebsa. Jon amonowy jest silnie toksyczny. W organizmie człowieka jest więc szybko przekształcany w substancję mniej toksyczną i dobrze rozpuszczalną w wodzie - mocznik. W ciągu doby człowiek wydala średnio kilkanaście gramów mocznika w postaci stężonego moczu.
b)Glukoneogeneza, zespół przemian biochemicznych zachodzących w wątrobie, prowadzący do wytworzenia glukozy lub glikogenu z substancji niecukrowych: aminokwasów, kwasu mlekowego, pirogronowego i in. Glukoneogeneza pobudzana jest i kontrolowana przez glikokortykoidy. Głównym substratem jest pirogronian. Glukoneogeneza ma miejsce głównie w komórkach wątroby, częściowo również w nerkach. W wyniku glukoneogenezy wydzielają się duże ilości energii.
Jest to proces syntezy glukozy z mleczanu i związków nie cukrowych. W warunkach tlenowych, pod wpływem dehydrogenazy mleczanowej, mleczan przechodzi w pirogronian. Właściwy proces glukoneogenezy rozpoczyna się pirogronianu i jest w dużej mierze odwróceniem procesu glikolizy. Jednak nie wszystkie etapy glikolizy można w prosty sposób odwrócić. Do takich etapów należy przejście:
- glukozy w glukozo-6-fosforan,
- fruktozo-6-fosforanu we fruktozo1,6-dwufosforan,
- fosfoenolopirogronianu w pirogronian,
to enzymatyczny proces tworzenia przez organizm glukozy z metabolitów nie będących węglowodanami. Większość czynników wpływających na aktywność szlaku glukoneogenezy to substancje powodujące zmniejszenie czynności wykorzystywanych w nim enzymów, jednak zarówno acetylo-CoA jak i cytrynian działają na nie aktywująco. Glukoneogeneza rozpoczyna się od wytworzenia α-ketoglutaranu przez karboksylację pirogronianu kosztem jedej cząsteczki ATP. Reakcja jest katalizowana przez odpowiednią karboksylazę. α-ketoglutaran jest następnie dekarboksylowany i równocześnie fosforylowany do fosfoenolopirogronianu przez odpowiednią karboksykinazę. W czasie tej reakcji jedna cząsteczka GTP jest hydrolizowana do GDP. Obie reakcje zachodzą w mitochondriach.