Krzysztof H. Włodarski
KREW I SZPIK
Krew stanowi płynną tkankę, należącą do szerokiej grupy tkanki łącznej. Tak jak każdy rodzaj tkanki łącznej, krew składa się z elementów komórkowych oraz substancji międzykomórkowej, zwanej surowicą, w której zawieszone są komórki, zwane krwinkami.
Krew jest wytworem kilku tkanek i narządów: tkanki hematopoetycznej (krwiotwórczej), narządów limfopoetycznych (węzłów chłonnych, śledziony, grasicy, migdałków, grudek limfatycznych rozsianych w jelitach i płucach), które dostarczają do krwi elementy komórkowe, oraz wątroby i narządów limfatycznych, które dostarczają elementy surowicy: białka albuminy (m. in. wiążące wodę), globuliny (większość z nich stanowią przeciwciała) oraz fibrynogen, będący odpowiednikiem włókien w substancji międzykomórkowej tkanki łącznej, który w procesie krzepnięcia krwi przekształca się w nierozpuszczalne nici włóknika (fibryny).
Krew jest łatwo dostępna tkanką, którą można pobierać od pacjentów do analizy, gdyż badanie cytologiczne i chemiczne krwi wnosi bardzo wiele podstawowych informacji o stanie pacjenta. Wynika to z faktu, że o składzie i chemii krwi decyduje stan narządów, dostarczających do krwi komórki i elementy surowicy i znajdujące się w niej produkty metabolizmu i wydzielania.
Krew spełnia w organizmie dwie podstawowe funkcje: 1) rozprowadza po całym organizmie tlen, składniki odżywcze (aminokwasy, glukozę i lipidy) i hormony oraz przenosi zużyte produkty przemiany materii do nerek i płuc, gdzie są usuwane; 2) utrzymuje homeostazę (tj. stałe warunki środowiska) ustroju.
KOMÓRKI KRWI OBWODOWEJ
Elementami morfotycznymi krwi obwodowej są: erytrocyty (czerwone ciałka krwi), trombocyty (płytki krwi) - elementy pozbawione jąder komórkowych, a więc nie będącymi prawdziwymi komórkami, oraz komórki jądrzaste, zwane leukocytami lub białymi ciałkami krwi. Wśród leukocytów wyróżniamy: 1) granulocyty, tj. komórki zawierające w cytoplazmie ziarnistości o różnym powinowactwie (chłonności) do barwników - a) obojętnochłonne, b) kwasochłonne, c) zasadochłonne, i odpowiednio wyróżniamy granulocyty obojętnochłonne (neutrofile), granulocyty kwasochłonne (eozynofile) i granulocyty zasadochłonne (bazofile) oraz 2) komórki pozbawione ziarnistości: monocyty i limfocyty.
Erytrocyty sa najliczniejszymi elementami morfotycznymi krwi: 4.5 - 5.5 x 106/µL, dla mężczyzn wartości są wyższe niż dla kobiet. Dwuwklęsły kształt tych komórek zapewnia najlepszy stosunek powierzchni do objętości, optymalny dla wydajnej wymiany gazowej. Średnica erytrocytu wynosi 7-8 µm, a więc odpowiada przekrojowi najmniejszych naczyń włosowatych. Erytrocyty stanowią około 45% całkowitej objętości krwi. Stosunek objętości upakowanych na drodze wirowania erytrocytów do objętości pełnej krwi nazywa się hematokrytem. U mężczyzn wynosi on 47%, u kobiet - 42%, ale może zmieniać się w dość dużych granicach w stanach chorobowych.
Zasadniczą funkcją erytrocytów (w nomenklaturze laboratoryjnej RBC - ang. red blood cells) jest transport gazów pomiędzy płucami a tkankami. Zarówno tlen, jak i dwutlenek węgla przenoszone są w erytrocytach za pomocą hemoglobiny (Hb), której obecność nadaje erytrocytom charakterystyczny kolor.
Powinowactwo hemoglobiny do tlenu zmniejsza się wraz ze wzrostem kwasowości. Dzięki temu krew, znajdująca się w tych miejscach, gdzie zachodzi intensywna przemiana materii (= metabolizm) i gdzie produkowane są kwaśne metabolity, zmniejsza swe powinowactwo do tlenu, który zostaje uwolniony do tych aktywnch metabolicznie miejsc.
Powinowactwo hemoglobiny do tlenu jest niższe przy niskim ciśnieniu parcjalnym tlenu (w tkankach) a wysokie przy wysokim ciśnieniu parcjalnym tlenu (w płucach). Ciśnienie parcjalne dwutlenku węgla także wpływa na powinowactwo hemoglobiny do tlenu: im niższe ciśnienie parcjalne dwutlenku węgla, tym większe powinowactwo hemoglobiny do tlenu. Ta reguła ma zasadnicze znaczenie fizjologiczne, gdyż pozwala na wydajne łączenie tlenu w płucach (gdzie panuje wysokie ciśnienie parcjalne tlenu i niskie ciśnienie parcjalne dwutlenku węgla) i uwalnianie tlenu w tkankach, gdzie ciśnienie parcjalne tlenu jest niższe, niż dwutlenku węgla.
Erytrocyty osobników dorosłych nie zawierają jądra komórkowego, ale u płodów i u noworodków posiadają jądra, które jednak nie są zdolne do ekspresji genów, gdyż mają bardzo zbitą chromatynę. Erytrocyty nie posiadają organelli komórkowych - aparatu Golgiego, siateczki śródplazmatycznej, mitochondriów i rybosomów, aczkolwiek niekiedy młode postacie erytrocytów zawierają agregaty rybosomów, które łącząc się z barwnikami zasadowymi tworzą obraz siateczki. Stąd nazwa takich komórek - retikulocyty (rete = sieć). W pewnych stanach chorobowych (anemia hemolityczna) liczba retikulocytów znacznie wzrasta.
Czas życia erytrocytów wynosi ok. 4 miesięcy, po tym czasie zużyte erytrocyty zostają zniszczone przez komórki żerne (makrofagi) w śledzionie, wątrobie i w szpiku, a z uwolnionej hemoglobiny zostaje uwolnione żelazo, wykorzystywane do produkcji hemoglobiny w nowych erytrocytach.
Płytki krwi (trombocyty) stanowią składniki morfotyczne krwi, które można ujawnić stosując specjalne metody barwienia. Są to bardzo małe elementy krwi o średnicy 2-4 µm, a w 1 µL krwi znajduje się ich ok. 200 - 400 000. Płytki krwi stanowią fragmenty cytoplazmy olbrzymich komórek, występujących w szpiku i w śledzionie, zwanych megakariocytami. Odszczepianie trombocytów od cytoplazmy megakariocytów zachodzi w świetle szerokich naczyń włosowatych (zatokowych), do których megakariocyty wciskają swe wypustki
cytoplazmatyczne.
Płytki krwi odpowiedzialne są za powstawanie skrzepu - koagulacji krwi. W miejscu uszkodzenia naczynia krwionośnego tworzą one agregaty i uwalniają substancje, aktywujące przemianę rozpuszczalnego białka fibrynogenu we włókienkowatą postać włóknika (fibryny). W sieci tego włóknika znajdują się nie tylko płytki krwi, ale także leukocyty. Razem tworzą one czop o galaretowatej konsystencji, uszczelniający miejsce zranienia.
Leukocyty (krwinki białe)
Krwinki jądrzaste (leukocyty) są to komórki zawierające jądro oraz ziarnistości w cytoplazmie (granulocyty) lub też pozbawione są ziarnistości cytoplazmatycznych (agranulocyty). Pośród granulocytów wyróżniamy granulocyty obojętnochłonne (neutrofile), granulocyty kwasochłonne (eozynofile) i granulocyty zasadochłonne (bazofile), a pośród agranulocytów wyróżnia się limfocyty oraz monocyty.
Całkowita zawartość leukocytów we krwi obwodowej wynosi 7000-8000/µL, lecz wartość ta nie jest stała i zmienia się w różnych stanach patologicznych.
Analiza morfologiczna rozmazów krwi, określająca procentową zawartość różnych leukocytów nazywa się wzorem leukocytarnym lub składem procentowym krwi i stanowi podstawowy test laboratoryjny. Tzw. badanie morfologiczne krwi (= „morfologia krwi”) składa się z oceny zawartości erytrocytów i leukocytów w 1 ul. (= 1 mm3) krwi oraz określenia składu procentowego leukocytów.
Tabela
Skład odsetkowy krwinek białych we krwi obwodowej
|
% |
Wartości bezwzględne (x 1000) w 1 μL |
Granulocyty: - obojętnochłonne - kwasochłonne - zasadochłonne |
45 - 65 1 - 5 0 - 1 |
3,1 - 5,5 0,2 - 0,4 0,0 - 0,1 |
Limfocyty |
28 - 42 |
2,1 - 3,3 |
Monocyty |
4,5 - 8,5 |
0,3 - 0,7 |
Razem |
100 |
4,5 - 13,0 |
Granulocyty obojętnochłonne (neutrofile)
Jest to najliczniejsza (60-70%) grupa krwinek białych. Komórki te posiadają jądro segmentowane, a liczba segmentów jądra (tzw. płatów) jest funkcją wieku komórki. Młode postacie neutrofilów (ok. 3% w krwi obwodowej) posiadają jądro w kształcie wygiętej pałeczki (tzw. jądro pałeczkowate), natomiast jądra starszych granulocytów obojętnochłonnych podzielone są na 2-7 płatów. Tak więc zwiększony odsetek komórek o jądrach pałeczkowatych wskazuje na uwalnianie do krwi postaci młodych, niedojrzałych (tzw. przesunięcie wzoru leukocytarnego w lewo), co ma miejsce np. w ostrych procesach zapalnych. Obniżona produkcja granulocytów obojętnochłonnych, np. w przypadku uszkodzenia szpiku, manifestuje się zwiększeniem odsetka komórek o jądrach wielopłatowych (tzw. przesunięcie wzoru w prawo).
Cytoplazma granulocytów obojętnochłonnych posiada liczne, drobne ziarnistości, w rutynowej ocenie rozmazu niedostrzegalne, tak że granulocyty obojętnochłonne identyfikuje się na podstawie morfologii jąder komórkowych (jądra podzielone na płaty).
Neutrofile, po kilku godzinach przebywania w naczyniach krwionośnych przechodzą przez ściany drobnych naczyń włosowatych do otaczających je tkanek. W ostrych stanach zapalnych migracja granulocytów do miejsca zakażonego jest bardzo pokaźna. Granulocyty obojętnochłonne stanowią główny rodzaj komórek w tego typu reakcji zapalnej.
Granulocyty obojętnochłonne odgrywają kluczową rolę w obronie organizmu przed zakażeniem. Pożerają one (fagocytują) bakterie, wykorzystując enzymy obecne w ziarnistościach cytoplazmatycznych do trawienia wychwyconych bakterii. Granulocyty obojętnochłonne biorą także udział w usuwaniu (= fagocytowaniu) z tkanek martwych komórek.
Granulocyty kwasochłonne (eozynofile)
Te komórki są nieco większe od granulocytów obojętnochłonnych, ich cytoplazma obładowana jest dużymi, wyraźnymi ziarnistościami, barwiącymi się eozyną (stąd nazwa „eozynofile”). Ziarnistości te zawierają wiele substancji, w tym enzym rozkładający histaminę - związek uwalniany w przebiegu reakcji immunologicznych oraz specyficzne białko o działaniu toksycznym dla pasożytów. Komórki te odgrywają więc istotną rolę w zwalczaniu pasożytów i w chorobach pasożytniczych liczba eozynofilów we krwi wzrasta ponad normę.
Jądra eozynofilów są bardzo charakterystyczne - składają się z dwóch płatów, połączonych wąskim przesmykiem chromatyny (tzw. „jądro okularowe”).
Granulocyty zasadochłonne (bazofile)
Granulocyty zasadochłonne (bazofile) są najrzadziej spotykaną we krwi krwinką, często w rozmazie krwi nie spotyka się jej. Komórka ta posiada bardzo liczne, duże, intensywnie barwiące się barwnikami zasadowymi ziarnistości cytoplazmatyczne, całkowicie zasłaniające pojedyncze jądro.
Ziarnistości bazofilów zawierają liczne substancje o silnym działaniu biologicznym, m.in. histaminę. Uwalnianie tych ziarnistości (= degranulacja komórki) w przebiegu reakcji alergicznych powoduje przedostanie się tych substancji do tkanek, w następstwie czego dochodzi do rozmaitych reakcji miejscowych, np. rozszerzenia naczyń krwionośnych, ściągnięcia (= chemotaksja) innych komórek (granulocytów kwasochłonnych, obojętnochłonnych, płytek krwi) w miejsce reakcji.
Liczba tych komórek wzrasta w stanach uczuleniowych (alergiach).
Limfocyty
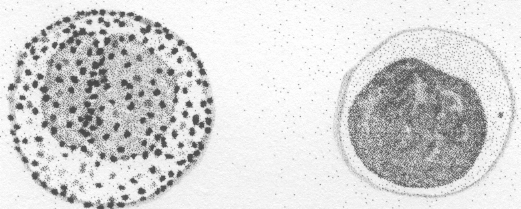
Są to komórki stanowiące ok. 30-35% składu leukocytów. Ich średnica wynosi ok. 8-12 µm, a więc są to najmniejsze komórki jądrzaste krwi. Posiadają okrągłe jądro o bardzo zbitej chromatynie, intensywnie barwiące się, oraz zawierają bardzo wąski rąbek cytoplazmy, niekiedy nawet nie widoczny w mikroskopie. Jednak w tej niewielkiej ilości cytoplazmy występują wszystkie organelle komórkowe.
Krążące we krwi limfocyty należą do trzech kategorii: limfocytów B (15-30%), limfocytów T (55-75%) oraz limfocytów NK (0-5%). W rutynowych badaniach rozmazów krwi tych rodzajów limfocytów nie rozróżnia się. Jednak limfocyty NK zawierają pojedyncze ziarnistości podobne do ziarnistości w bazofilach. Limfocyty można zidentyfikować stosując jedynie specjalne metody immunologiczne, ujawniające pewne cechy antygenowe, występujące na powierzchni tych komórek.
Limfocyty biorą udział w reakcjach obronnych (immunologicznych) ustroju, przy czym poszczególne klasy limfocytów spełniają odrębne funkcje.
Limfocyty B odpowiedzialne są za wytwarzanie krążących przeciwciał (tzw. „przeciwciała humoralne”) ale dopiero po przejściu w stan pobudzenia przez kontakt z odpowiednimi antygenami.
Limfocyty T odpowiedzialne są za niszczenie komórek zakażonych wirusami lub wprowadzonych do ustroju komórek obcych (np. przeszczepów tkankowych). Limfocyty T zaangażowane więc są w odpowiedź immunologiczną typu komórkowego.
Limfocyty NK są komórkami nie posiadającymi cech antygenowych (= markerów) ani dla limfocytów B, ani dla limfocytów T, dlatego też występują pod nazwą „komórki żadne” (ang. null cells) . Limfocyty K zaangażowane są w odpowiedź przeciwnowotworową ustroju.
Monocyty, stanowiące od 5 do 8% populacji leukocytów, są komórkami o dość znacznej heterogenności. Duże monocyty, o średnicy 30-40 µm, posiadają duże, nerkowatego kształtu jądro o luźnej strukturze chromatyny i z wyraźnym jąderkiem. Małe monocyty, o średnicy 20-30 µm, posiadają jądra owalne lub okrągłe, a jąderka są niewidoczne. Taką postać monocytu trudno jest odróżnić od dużego limfocytu. Mimo, że monocyty zalicza się do agranulocytów, w ich cytoplazmie spotyka się pojedyncze ziarnistości. Cytoplazma tych komórek zawiera też wakuole oraz wiele lizosomów - organelli komórkowych związanych z trawieniem wewnątrzkomórkowym.
Spadek zawartości monocytów we krwi nazywa się monocytopenią, wzrost zawartości - monocytozą.
Ich główną funkcją jest fagocytowanie (pożeranie) bakterii, wirusów, kompleksów antygen-przeciwciało, uszkodzonych komórek i starych erytrocytów. Monocyty, po opuszczeniu krążenia, zasiedlają różne tkanki i przekształcają się w różne formy makrofagów -tkankowych komórek o wybitnej zdolności fagocytarnej, a także stają się prekursorami komórek degradujących tkankę kostną - osteoklastów.
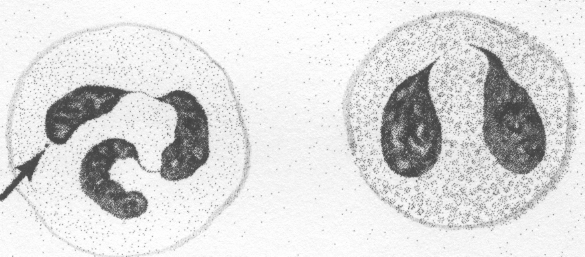
GRANULOCYT GRANULOCYT MONOCYT
OBOJĘTNOCHŁONNY KWASOCHŁONNY
GRANULOCYT LIMFOCYT
ZASADOCHŁONNY
Ryc. 1
Schematyczne obrazy leukocytów.
SZPIK
Szpik stanowi tkankę hemopoetyczną, to jest miejsce powstawania i dojrzewania wszystkich rodzajów krwinek, tak jądrzastych, jak i pozbawionych jąder.
Ogólnie biorąc, szpik składa się z dwóch przedziałów tkankowych: 1) zrębu, do którego zalicza się szerokie naczynia włosowate (sinusoidy), komórki siateczki (odmiana fibroblastów) i wytworzone przez nie włókna siateczkowate, makrofagi (=komórki żerne), komórki tłuszczowe oraz komórki wyścielające jamę szpikową (komórki śródkostnej); 2) przedziału hemopoetycznego (krwinkotwórczego), ulokowanego w rusztowaniu przedziału zrębowego.
Komórki przedziału hemopoetycznego stanowią dominujący element komórkowy szpiku, aczkolwiek w tzw. szpiku żółtym, hematopoetycznie nieczynnym, dominującym elementem są zrębowe komórki tłuszczowe. Szpik żółty może jednak w pewnych sytuacjach przekształcić się w krwiotwórczy (hematopoetyczny) szpik czerwony.
Wszystkie elementy krążące we krwi biorą swój początek z tzw. wielopotencjalnej (pluripotencjalnej) komórki macierzystej, nazwanej CFU (z ang. Colony Forming Unit) - komórka wytwarzająca w śledzionie napromieniowanych zwierząt kolonie komórek krwiotwórczych. Z komórki tej, drogą licznych podziałów i stopniowego różnicowania się powstają wszystkie linie komórkowe krwi. Proces ten nazywa się hematocytopoezą.
Na hematocytopoezę składają się:
Erytropoeza - powstawanie erytrocytów z dużych komórek prekursorowych, zwanych erytroblastami. Erytroblasty w miarę postępujących licznych podziałów komórkowych syntetyzują coraz więcej hemoglobiny, przez co barwliwość ich cytoplazmy zmienia się od zasadochłonnej (brak hemoglobiny), poprzez wielobarwliwą (pojawia się hemoglobina), aż po kwasochłonną. (Hemoglobina, przypominam, wykazuje silne powinowactwo do kwaśnego barwnika - eozyny). Tak więc w miarę dojrzewania komórek szeregu erytropoetycznego cytoplazma ich przypomina coraz bardziej cytoplazmę erytrocytu, a ich jądra komórkowe --> stają [Author:WP] się coraz bardziej zbite i komórki tracą zdolność proliferacji. W końcowym etapie różnicowania z komórki zwanej erytroblastem kwasochłonnym (= ortochromatycznym) usunięte zostaje jądro, a powstały w ten sposób bezjądrzasty erytrocyt przedostaje się do światła naczyń zatokowych szpiku. Uwalniane jądra erytroblastów ortochromatycznych zostają sfagocytowane przez makrofagi.
Granulopoeza - wytwarzanie wszystkich rodzajów granulocytów. Proces ten rozpoczyna się od komórki, zwanej mieloblastem. Jest to komórka, nie posiadająca jeszcze ziarnistości. Ziarnistości nieswoiste pojawiają się w cytoplazmie kolejnej formy - promielocycie. Dopiero jego następcy - mielocyty wykształcają ziarnistości specyficzne (kwaso-, zasado- i obojętnochłonne), a jądra tych komórek ulegają kondensacji i przybierają nieregularną formę. Z mielocytów powstają metamielocyty, zawierające już wykształcone ziarnistości swoiste, ale proces kondensacji jąder nie jest w nich jeszcze zakończony. To staje się dopiero na etapie dojrzałych granulocytów, które przechodzą do naczyń krwionośnych, pokonując tzw. „barierę szpikową” - mechanizmy molekularnego rozpoznawania, czy komórka jest już w pełni dojrzałą, czy jeszcze wymaga przebywania w mikrośrodowisku szpiku. Czas potrzebny do wykształcenia się dojrzałego granulocytu ocenia się na ok. dwa tygodnie.
Limfocytopoeza. Limfocyty, obecne we krwi, pochodzą nie tylko ze szpiku, ale również z narządów limfopoetycznych. Komórką progenitorową jest limfoblast, a jego potomnymi komórkami są limfocyty wszystkich trzech rodzajów. Limfocyty T powstają w szpiku, ale nabywają swą cechę limfocytów T (tzw. kompetencję) po przejściu przez środowisko zrębu grasicy. Limfocyty B swą kompetencję nabywają w środowisku szpiku.
Monocytopoeza. Monocyty rozwijają się ze wspólnej komórki macierzystej dla mielopoezy - CFU, w której wydrębnia się gałąź, dającą początek linii monocytarnej.
Trombocytopoeza (wytwarzanie płytek krwi). Płytki krwi powstają przez odłączenie się fragmentu cytoplazmy z wielojądrzastych, dużych (o średnicy ponad 100 μm) komórek, zwanych megakariocytami. Te zaś powstają z megakarioblastów, a te wywodzą się ze wspólnej komórki macierzystej - CFU. Odłączanie się płytek od cytoplazmy megakariocytu następuje w świetle naczynia zatokowego, do światła którego wnikają wypustki cytoplazmatyczne tej olbrzymiej komórki. Czas życia płytek wynosi 7-10 dni.
Procesy namnażania i różnicowania komórek hemopoetycznych regulowane są przez liczne czynniki wzostu, tak produkowane przez komórki zrębu szpiku, jak też wytwarzane poza szpikiem. Do prawidłowej erytropoezy potrzebny jest udział erytropoetyny - hormonu produkowanego w nerkach, a do trombocytopoezy - trombopoetyny.
Udział czynników wzrostu w regulacji hemopoezy odbywa się na różnych poziomach różnicowania się komórek i najczęściej do przebiegu prawidłowej hemopoezy potrzebny jest jednoczesny udział kilku czynników wzrostu. Jest to jednak tak złożony proces, że nie można go lapidarnie opisać w kilku zdaniach. Podkreślić jednak należy, że do prawidłowego przebiegu hemopoezy niezbędne jest mikrośrodowisko (tzw. „nisza ekologiczna”), wytworzone przez komórki zrębowe szpiku. Zrąb ten należy więc traktować nie jako „rusztowanie” dla komórek hemopoetycznych, lecz jako regulatora i „opiekuna” różnicujących się komórek krwiotwórczych.
Różnicowanie komórek macierzystych (CFU) szpiku:
CFU
MEGAKARIOCYTOBLAST LIMFOBLAST MIELOBLAST PROERYTROBLAST
LIMFOCYT LIMFOCYT LIMFOCYT
MEGAKARIOCYT B T K
MIELOCYT ERYTROBLAST
ZASADOCHŁONNY
PŁYTKI
EOZYNOFIL NEUTROFIL BAZOFIL ERYTROBLAST
POLICHROMATOFILNY
ERYTROBLAST
KWASOCHŁONNY
RETIKULOCYT
ERYTROCYT
SZPIK JAKO ŻRÓDŁO KOMÓREK KOŚCIOTWÓRCZYCH
Szpik erytropoetyczny posiada zdolność wytwarzania tkanki kostnej. Przeszczepiony w miejsce złamania kości przyspiesza proces wytwarzania tzw. blizny kostnej, a przeszczepiony w miejsca heterotopowe, np. do nerki, wywołuje osteogenezę.
Komórkami szpiku, odpowiedzialnymi za produkcję tkanki kostnej są komórki zrębowe szpiku, nie zaś komórki hemopoetyczne.
Komórki zrębowe szpiku mogą także różnicować się w komórki tłuszczowe i w komórki chrzęstne.
Komórki zrębowe szpiku można namnażać w hodowli in vitro i wykorzystywać dla celów odtwarzania tkanki kostnej.
Wyszukiwarka
Podobne podstrony:
grasica i nadnercza, Szkoła, studia, histologia, do nauki
układ płciowy żeński, Szkoła, studia, histologia, do nauki
histo test stoma, Studia, Histologia
progam zajęć, studia, histologia
Ukł. dokrewny, STUDIA, histologia
histologia-tkanka nabłonkowa, Szkoła, studia, histologia
Krew, Studia - ratownictwo medyczne, 3 rok, Zawansowane procedury ratunkowe
AnM 2ROK, studia, histologia
układ moczowy, Szkoła, studia, histologia, do nauki
Wątroba, studia, histologia
HISTO GIEŁDA opisowa, Szkoła, studia, histologia, do nauki
Krew, Prywatne, Studia, Fizjologia
Krew, Pomoce naukowe na studia powiązane z medycyną
07. Układ oddechowy, Studia - materiały, Histologia, Wykłady - histologia
Krew i mięśnie, Lekarski I rok ŚUM, histologia, histologia
więcej podobnych podstron