12. Budowa chromosomów eukariotycznych
Chromosomy eukariotyczne mają postać liniową, a ich DNA jest ściśle połączony z dużą ilością specyficznych białek, które pełnią rolę funkcjonalną i strukturalną. Znajdują się one w jądrze komórkowym. Zawartość DNA w chromosomach eukariotycznych jest dużo większa niż w prokariotycznych i dlatego w każdym chromosomie eukariotycznym znajduje się wiele miejsc początku replikacji.
Chromosomy eukariontów można zwykle obserwować jedynie w czasie podziału komórki, gdy w wyniku replikacji utworzyły się już dwie identyczne struktury, zwane chromatydami (chromosomy siostrzane). Można je wtedy obserwować w mikroskopie świetlnym.
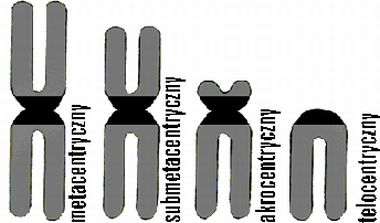
Chromosomy klasyfikuje się na podstawie morfologii, w zależności od położenia centromeru (przewężenia pierwotnego):
▪ chromosomy metacentryczne - centromer położony środkowo, dzieli chromosom na dwie połowy (ramiona). Ramiona p i q są sobie równe.
▪ chromosomy submetacentryczne - centromer znajduje się w pobliżu środka i dzieli chromosom na ramię krótsze p oraz dłuższe q.
▪ chromosomy akrocentryczne - centromer położony bliżej końca jednego z ramion (bardzo krótkie ramię p).
▪ chromosomy telocentryczne - brak krótkich ramion p. Nie występują u ludzi.
Molekularna struktura chromosomów:
Chromatyna (materiał budujący chromosomy) - kompleks białek i DNA (kompleks nukleoproteinowy):
▫ 30% białka histonowe (zasadowe)
▫ 30% białka wiążące DNA (kwaśne białka niehistonowe)
▫ 30% DNA
▫ 10% RNA - występuje przejściowo i jest eksportowany do cytoplazmy
Struktura chromatyny służy upakowaniu i organizacji chromosomowego DNA i jest zdolna do zmiany poziomów skondensowania w zależności od etapów cyklu komórkowego.
Euchromatyna - luźniejsza struktura, najbardziej rozproszony region chromosomu interfazowego, aktywna część chromatyny, biorąca udział w transkrypcji genów.
Heterochromatyna - struktura skondensowana, zawiera wiele powtórzeń DNA, pozbawiona aktywności genowej:
- konstytutywna - bezwzględnie pozbawiona aktywności genowej.
- fakultatywna - fragmenty chromatyny występujące zarówno w postaci heterochromatynowej, jak i euchromatynowej; wyłączona aktywność genów, np. ciałko Baara.
Histony to grupa białek o masie mniejszej niż 23 kDa (wyjątek stanowi histon H1 o masie 23 kDa). Charakteryzują się dużą zawartościa aminokwasów zasadowych, głównie lizyny i argininy. W fizjologicznym pH mają ładunek dodatni, co ułatwia ich ścisłe powiązanie z kwaśnym polianionowym DNA. Wyróżniamy 5 typów histonów: H1, H2A, H2B, H3 i H4, które oprócz H1 wystepują u wszystkich gatunków. Histon H1 może mieć kilka różnych odmian. W pozostałych histonach sekwencja aminokwasów jest wysoce konserwatywna (nie zmieniła się w czasie ewolucji).
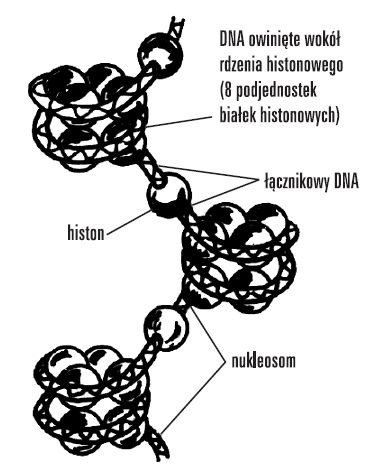
Podstawową jednostką struktury chromosomu jest nukleosom, złożony z białkowego oktameru, zawierającego po 2 histony rdzeniowe każdego typu (H2A, H2B, H3, H4) oraz owijającego go lewoskrętnie 1,8 razy odcinka DNA o dł. 146 pz (co razem stanowi rdzeń nukleosomu) i histonu H1. Nawinięcie DNA na białkowy oktamer wyjaśnia ujemną superhelikalność eukariotycznego DNA.
Pojedyncza cząsteczka H1 stabilizuje DNA w miejscu, w którym wchodzi on do nukleosomu i opuszcza go, oraz organizuje DNA pomiędzy nukleosomami.
Rdzeń nukleosomu i histon H1 nazywany jest chromatosomem.
Powtarzająca się jednostka nukleosomowa ma ok. 200 pz (przeciętna długośc łącznikowego DNA między rdzeniami nukleosomów wynosi ok. 55 pz).
Solenoid (włókno 30 nm) - struktura wyższego rzędu zorganizowania chromatyny, zbudowana z lewoskrętnej helisy nukleosomów, w której na 1 obrót przypada około 6 nukleosomów. W tej formie występuje większość chromatyny.
W większej skali, chromosomowy DNA jest zorganizowany w pętle dochodzące do 100 kpz, występujące w formie włókien 30 nm, utrzymywane przez białkowe rusztowanie matriks jądrowej.
Najwyższy stopień kondensacji osiągają chromosomy podczas metafazy podziału mitotycznego. Organizacja tych struktur wymaga połączenia włókien chromatynowych z chromosomowym rusztowaniem. Jest ono zbudowane głównie z niehistonowego białka jądrowego (topoizomerazy II). Specyficzne rejony DNA o dł. kilkuset pz, bogate w adeninę i tyminę to rejony łącznikowe z rusztowaniem (SAR - scaffold attachment regions), bo wiążą DNA z rusztowaniem chromosomowym. Odcinki DNA od jednego SAR do drugiego układają się w pętle różnej długości. Pętle chromosomowe tworzy chromatyna w formie włókien 30 nm.
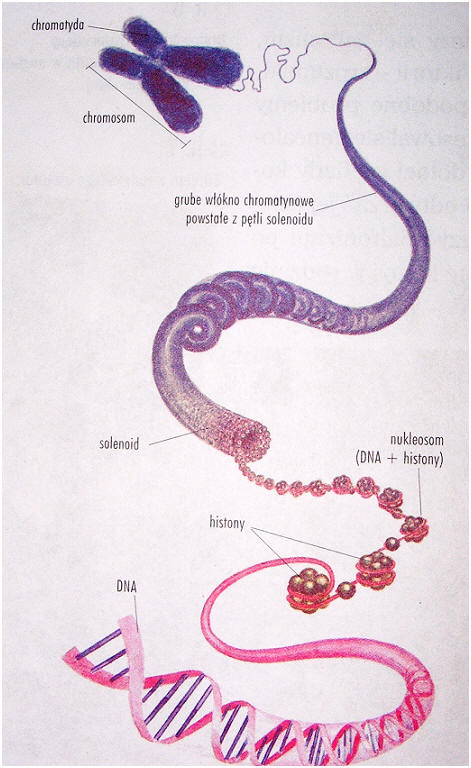
Upakowanie DNA w chromosomach eukariotycznych:
٠ nukleosom (złożony z podwójnego dysku, utworzonego z 8 cząsteczek, po dwie każdego z histonów H2A, H2B, H3 i H4. Wokół takiego oktameru owinięty jest odcinek DNA o dł. 146 pz). Każdy nukleosom jest powiązany z następnym za pomocą DNA łącznikowego. Z zewnętrzną stroną rdzenia nukleosomu łączy się pojedyncza cząsteczka histonu H1 odpowiedzialna za dalsze fałdowanie się nukleosomów w solenoid i kolejne, bardziej kompleksowe struktury. W tworzeniu rusztowania chromosomowego i regulacji genów biorą udział kwaśne białka.
٠ nić nukleosomowa.
٠ spirala utworzona z nici nukleosomowej.
٠ spirala tworzy pętle przytwierdzone do białkowego szkieletu w chromosomie.
٠ ściśle upakowane pętle tworzą ramiona chromosomu.
Elementy budowy chromosomów eukariotycznych:
elementy niezbędne:
◦ origin replikacji.
◦ centromer - miejsce, w którym w czasie podziału komórki dochodzi do przyłączenia włókien wrzeciona kariokinetycznego. Zbudowany jest z wysoce powtarzalnego (repetytywnego), satelitarnego DNA. Białka związane z centromerem tworzą kinetochor (wielowarstwową strukturę łączącą się z włóknami wrzeciona podziałowego).
◦ telomery - wyspecjalizowane sekwencje DNA, tworzące końce liniowej cząsteczki chromosomu. Zbudowane z krótkich, powtarzających się sekwencji DNA (TTAGGG u człowieka), które są syntetyzowane przez telomerazę w sposób niezależny od zwykłej replikacji DNA. Liczba tych powtórzeń w komórkach somatycznych maleje z wiekiem (molekularny marker procesu starzenia), lecz w komórkach rozrodczych i nowotworowych jest przez telomerazę utrzymywana na stałym poziomie. Telomery zapobiegają rekombinacji pomiędzy końcami chromosomów. Wśród eukariontów istnieje
mała zmienność sekwencji telomerowych.
Telomeraza - rybonukleoproteina, enzym utrzymujący na właściwym poziomie długość telomerów. Zawiera RNA komplementarny do powtarzających się sekwencji DNA w telomerach (RNA służy jako matryca do wydłużania telomerów). Nieobecna w komórkach somatycznych; pojawia się w przypadku ich transformacji w komórki nowotworowe, u których długość telomerów nie ulega skracaniu.
element dodatkowy (u niektórych chromosomów):
◦ organizator jąderka (NOR) - zlokalizowany w przewężeniu wtórnym. Zawiera tandemowo 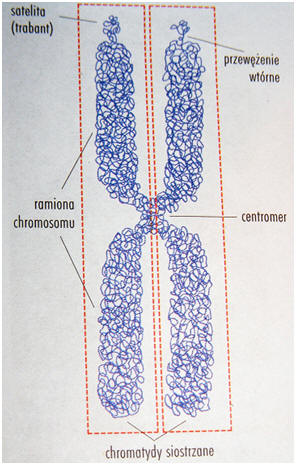
powtarzające się geny rRNA: 5,8S, 18S, 28S. Geny 5S rRNA u większości gatunków są rozproszone po
całym genomie. Jeśli poza NOR wystaje pewien odcinek chromosomu, to nazywany jest on satelitą lub trabantem. U człowieka liczba NOR-ów wynosi 10, są one obecne na wszystkich krótkich ramionach chromosomów akrocentrycznych, z wyjątkiem chromosomu Y. Każdy NOR składa się z ok. 80-100 powtórzeń. W czsie interfazy NOR-y ulegają dekondensacji, a wokół nich tworzy się jąderko. NOR-y z
różnych chromosomów mogą się łączyć ze sobą w jedno jąderko. Gdy komórka wchodzi w stadium metafazy w mitozie chromosomy mogą wyglądać jakby były wciąż połączone krótkimi ramionami. Jest to tzw. połączenie satelitarne.
Informacje dodatkowe:
Nadwrażliwość na trawienie deoksyrybonukleazą I (DNazą I) charakteryzuje aktywne regiony chromatyny, w których włókna 30 nm są zmienione przez przyłączenie do DNA specyficznych białek, lub regiony aktualnie podlegające transkrypcji.
Sekwencje 5'-CG-3' (CpG) w DNA ssaków zwykle mają zmetylowane cytozyny, chociaz w pobliżu promotorów często transkrybowanych genów występują „wyspy” niemetylowanych CpG, które tworzą regiony szczególnie wrażliwe na trawienie DNazą.
Wyszukiwarka
Podobne podstrony:
Biologia część I, Budowa komórki Eukariotycznej i funkcje jej organelli
6. Technika PCR i jej wykorzystanie-ok, Biologia II, Biologia molekularna
Biologia część I Budowa komórki Eukariotycznej i funkcje jej organelli
3. Przyklady regulacji ekspresji genow u Eukaryota-ok, Biologia II, Biologia molekularna
Budowa chemiczna organizmow, dydaktyka, biologia-praktyki, II gimnazjum
10. ZJAWISKO APOPTOZY I JEGO ZNACZENIE-ok, Biologia II, Biologia molekularna
11. Charakterystyka mitochondrialnego DNA-ok, Biologia II, Biologia molekularna
Budowa chromosomu, BiologiaMedyczna
4. Bakteriofagi charakterystyka i cykle rozwojowe-ok, Biologia II, Biologia molekularna
Biologia część I, Budowa komórki Eukariotycznej i funkcje jej organelli
10 Biologia molekularna 5 12 2011
biologia molekularna 12 pytania
tematy cwiczen - ii rok biologii, Biol UMCS, IV semestr, Biologia molekularna, Egzamin
II.DWUDZIESTOLECIE MIĘDZYWOJENNE W EUROPIE, 12.Budowa socjalizmu w ZSRR, Marek Biesiada
II.DWUDZIESTOLECIE MIĘDZYWOJENNE W EUROPIE, 12.Budowa socjalizmu w ZSRR, Marek Biesiada
Budowa komórki eukariotycznej część II
zagadnienia na egzamin- hydro os ekzzp nbiol, II rok, Biochemia, Biologia molekularna, Biotechnologi
ĆWICZENIE II biol mol, far, III rok IV sem, biologia molekularna, II
Budowa komórki, Ratownicto Medyczne, Biologia
więcej podobnych podstron