BK4

3.5. POTENCJAŁ WODY
Z przedstawionych rozważań wynika, że o kierunku i szybkości przepływu wody przez błonę półprzepuszczalną nie decyduje tylko gradient stężenia wody lub tylko gradient ciśnienia hydrostatycznego, ale suma tych dwóch typów oddziaływań. Obserwacja ta doprowadziła do powstania koncepcji tzw. całkowitej siły napędowej (ang. total driving force) w ruchu wody, którą jest gradient energii swobodnej wody lub inaczej gradient potencjału chemicznego wody.
Potencjał chemiczny wody, l/yw (psi), jest to ilość energii swobodnej (inaczej zdolność do wykonania pracy użytecznej lub aktywność wody) wnoszonej do układu przez każdy mol wody. Nie można go zmierzyć bezpośrednio, ale można porównać potencjały chemiczne wody w różnych systemach, przyjmując jako punkt odniesienia potencjał chemiczny czystej wody (/i°w) przy ciśnieniu atmosferycznym i w takiej samej temperaturze, jak układ badany. Wobec tego potencjał wody jest różnicą między potencjałem chemicznym wody w układzie (pj a potencjałem chemicznym czystej wody (/r°w), który przy ciśnieniu atmosferycznym ma wartość 0:
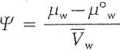
gdzie: Vw - parcjalna molalna objętość wody, tzn. objętość 1 mol wody (18 cm3). Podzielenie przez ten wyraz obu składników równania umożliwia określenie potencjału wody w jednostkach energii na jednostkę objętości, co jest równoważne jednostkom siły przypadającym na jednostkę powierzchni, czyli jednostkom ciśnienia:
energia • mol-1 wody energia
objętość • mol-1 wody objętość
Potencjał wody można zatem wyrażać w następujących jednostkach układu SI1: J-dm”3, gdzie 1 J = 1 N • 1 m, albo Pa (1 Pa — 1 N-m~2) i jego wielokrotności: kPa (103 Pa) lub MPa (106 Pa). W starszych podręcznikach można napotkać inne jednostki F, nie należące do obowiązującego dziś układu SI: erg cm-3 (1 erg = 10_7J), dyn-cm-2 (1 dyn = 10~5N), bar (1 bar = 105Pa = 0,987 atm), atmosfera
(1 atm = 101 325 Pa).
Ponieważ ciśnienie pary wodnej zależy od jej potencjału chemicznego, potencjał wody w układzie można wyrażać w parametrach tego ciśnienia:
RT\n 4
e
F = —=-
Vw
gdzie: R - stała gazowa, T - temperatura absolutna, e - ciśnienie pary wodnej w układzie, e° - ciśnienie pary czystej wody w tej samej temperaturze. Pomiary ciśnienia pary można wykorzystać do oznaczania potencjału wody w układzie. Stosunek procentowy e
obu tych ciśnień: — -100 - to wilgotność
względna (RPI - ang. relative humidity), miara często stosowana do określania stosunków wodnych w atmosferze.
Do oznaczenia potencjału wody w tkankach roślin wykorzystuje się wiele metod, np.:
• Bada się zmiany świeżej masy tkanek, inkubowa-nych w roztworach różnych substancji o znanym potencjale wody (nie zdysocjowanych i nie wnikających do protoplastu, np. glikolu polietylenowego). W roztworze o potencjale wody równoważnym potencjałowi tkania nie zajdzie przepływ wody i nie ulegnie zmianie masa tkanki. Metoda ta ma wiele ograniczeń, gdyż w czasie inkubacji tkanki w roztworach masa komórek może ulegać zmianom w wyniku procesów wzrostowych lub oddechowych
• Techniki psychrometryczne polegają na rejestrowaniu spadku temperatury tkanki spowodowanego parowaniem wody i na oznaczeniu momentu, w którym prężność pary wodnej w komorze pomiarowej osiągnie równowagę z potencjałem wody tkanki. Zmiany temperatury próby rejestruje się za pomocą termopary powiązanej z odpowiednio wyskalowanym miernikiem prądu elektrycznego.
• Zastosowanie komory (bomby) ciśnieniowej (wg Scholandera i wsp. 1965), którego podstawą jest założenie, że potencjał wody w ksylemie dobrze odzwierciedla potencjał wody całego organu. Metoda polega na równoważeniu ujemnego ciśnienia hydrostatycznego, występującego w ksylemie większości roślin (patrz rozdz. 3.5.4), znanym ciśnieniem ze zbiornika ciśnieniowego. Jeśli znane jest ciśnienie osmotyczne soku ksylemu (wyznaczone np. krioskopowo, na podstawie oznaczenia punktu zamarzania soku), to łatwo wyliczyć potencjał wody ksylemu, a więc i potencjał wody w badanym organie.
3.6. STOSUNKI WODNE W KOMÓRCE ROŚLINNEJ
Wartości potencjału wody w komórkach roślinnych są na ogół mniejsze od 0, a więc ujemne, np. w stosunkowo dobrze uwodnionych komórkach wahają się w granicach od — 0,1 do —1,0 MPa. O wartości potencjału wody w komórce decydują trzy siły: l) ciśnienie hydrostatyczne, (Fp), wyższe od atmosferycznego, zwiększa energię swobodną (aktywność) wody, a więc podwyższa potencjał wody w komórce, 2) siły osmotyczne zmniejszają energię swobodną wody, wobec czego potencjał osmotyczny (Fn) ma wartość ujemną (potencjałowi osmotycznemu odpowiada liczbowo ciśnienie osmotyczne, ale ma ono znak +: w roztworze o Fn równym —0,3 MPa ciśnienie osmotyczne wynosi +0,3 MPa), 3) siły matrycowe (oddziaływania elektrostatyczne, wiążące wodę z makromolekularni obdarzonymi ładunkiem, siły pęcznienia, oddziaływania międzycząsteczkowe na granicy faz). Siły te, określane symbolem Fx (czasem Fm), zmniejszają energię swobodną wody, a więc FT ma waitość ujemną.
Zależność potencjału wody w komórce od tych trzech typów oddziaływań określa równanie:
Fw = FP+(-)Fn + (-)Fi
gdzie: Fp - ciśnienie turgorowe ściany komórkowej, Fn - oddziaływania osmotyczne związków małocząsteczkowych, głównie wakuolamych, Fi - napięcie powierzchniowe w mikrokapilarach ściany komórkowej oraz przyciąganie wody przez grupy polanie (hydroksylowe, karboksylowe i aminowe) cząsteczek wchodzących w skład struktur komórkowych (ścian komórkowych, białkowych koloidów cytoplaz-my, fosfolipidów).
Wartości poszczególnych parametrów decydujących o Fw w komórce nie są stałe: zależą od etapu rozwojowego komórki, jej stanu fizjologicznego i czynników środowiskowych. Ciśnienie turgorowe zależy między innymi od właściwości mechanicznych ściany komórkowej, które ulegają zmianom w czasie wzrostu i różnicowania komórek lub pod wpływem niektórych bodźców środowiskowych. W naczyniach ksylemu może powstawać ujemne ciśnienie hydrostatyczne, w wyniku intensywnej transpiracji (patrz rozdz. 3.9.3), a więc Fp w ksylemie może mieć wartość ujemną. Hydroliza makromo-lelcul do związków małocząsteczkowych os-motycznie czynnych (np. skrobi do cukrów prostych) lub odwrotność tego procesu - polimeryzacja małych cząsteczek do związków wielkocząsteczkowych - mogą stanowić mechanizm regulacji ciśnienia osmotycznego komórki.
J - dżul, N - niuton, Pa - paskal.
Wyszukiwarka
Podobne podstrony:
BK4 3.5. POTENCJAŁ WODY Z przedstawionych rozważań wynika, że o kierunku i szybkości przepływu
scan0007 (35) Z przedstawionych rozważań wynika, że w ogólnym przypadku własności zastosowanego w uk
scan0007 (35) Z przedstawionych rozważań wynika, że w ogólnym przypadku własności zastosowanego w uk
skanuj0044 (48) WSPÓŁCZESNA TURYSTYKA KULTUROWA 47 Z przedstawionej tabeli wynika, że przedmiotem za
img194 194 Z powyższych rozważać wynika, że najbardziej istotną częścią widma sygnału ASK, bo wynika
16.3. Programowanie mikroprocesorów Z dotychczasowych rozważań wynika, że
foto (14) Z przedstawionych rozkładów wynika, że im większy wskaźnik tolerancji k tym większe obciąż
Z tak przedstawionych definicji wynika, że organizacja formalna ma następujące cechy: -
82127 skanuj0295 (2) Stąd po przekształceniach 3 / 2Mobl • ą • cos/^ (11.74) Z dotychczasowych rozwa
foto (14) Z przedstawionych rozkładów wynika, że im większy wskaźnik tolerancji k tym większe obciąż
67 (18) Z przedstawionej zależności wynika, że płetwy mogą wytwarzać siłę nośną wy łącznie na płynąc
Z przedstawionych danych wynika, że osoby prywatne i firmy widzą szansę we wprowadzeniu precyzyjnych
14) Co to jest cofka ? Jest to spiętrzenie wody przed mostem wynikające ze zmniejs
CCF20081203�028 Stąd po przekształceniach 2Mobi -ą-cosP X-z- kgj (11.74) Z dotychczasowych rozważań
7 (418) Z przedstawionego materiału wynika, że ojcowie pragną wychowywać swe dzieci w posłuszeństwie
73 Synteza układów kompensacji.. Rys. 7 Z przeprowadzonych rozważań wynika, że składowę
Z przedstawionego zapisu wynika, że elastyczność funkcji mierzymy stosunkiem względnej (procentowej)
więcej podobnych podstron