CCF20080601�003

Rys. 16-10. Tlenek azotu (NO) uruchamia rozkurcz mięśnia gładkiego w ścianie naczynia krwionośnego. (A) Na rysunku przedstawiono połączenie nerwu z naczyniem krwionośnym. (B) Sekwencja zdarzeń prowadząca do rozszerzenia się naczynia krwionośnego.
Acetylocholina uwolniona z zakończeń nerwów w ścianie naczynia krwionośnego uruchamia syntezę i uwalnianie NO przez komórki śródbłonka wyściełające to naczynie. NO dyfunduje z komórek śródbłonka do przylegających komórek mięśnia gładkiego, powodując rozkurcz (relaksację) komórek mięśnia. Należy podkreślić, że gazowy NO jest wysoce toksyczny przy wdychaniu i nie należy go mylić z podtlenkiem azotu (N2O) znanym jako gaz rozweselający
(B)
pobudzone zakończenie nerwu
(A)
komórki mięśnia gładkiego błona podstawna

komórka śródbłonka
arginma
■V
ł
ł
-acetylocholina
tlenek
azotu
NO związany z białkiem docelowym
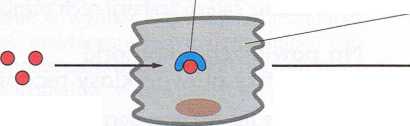
SZYBKA DYFUZJA NO PRZEZ BŁONY
SZYBKA RELAKSACJA KOMÓRKI MIĘŚNIA GŁADKIEGO
komórka mięśnia gładkiego
komórka
śródbłonka
klicznego AMP, znacznie częściej używanej wewnątrzkomórkowej czą-eczki sygnałowej, której działanie omówimy później.
iektóre hormony przenikają przez błonę komórkową wiążą się z receptorami wewnątrzkomórkowymi
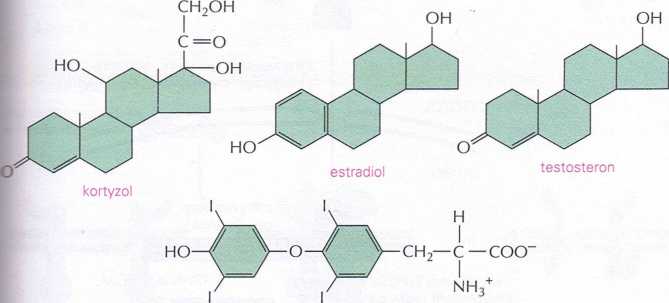
Gazy takie jak NO to nie jedyne cząsteczki sygnałowe, które mogą przenikać przez błony komórkowe. Hydrofobowe cząsteczki sygnałowe takie jak hormony steroidowe (w tym kortyzol, estradiol i testosteron) oraz hormony tarczycy takie jak tyroksyna (rys. 16-11) przechodzą przez błonę komórkową morki docelowej. Jednakże nie aktywują one enzymu wewnątrzkomórko-ego, ale wiążą się z białkami receptorowymi znajdującymi się w cytozolu łub jądrze. Tymi receptorami są białka zdolne do regulacji transkrypcji genów, przy czym w komórce nie poddanej stymulacji zwykle obecne są one w formie nieaktywnej. Po związaniu hormonu receptor ulega dużej zmianie konformacyjnej, co prowadzi do jego aktywacji, dzięki której może on inicjować lub hamować transkrypcję wybranego zespołu genów (rys. 16-12). Dla każdego typu hormonu istnieje odrębne białko receptorowe, a każdy
tyroksyna
Rys. 16-11. Niektóre małe hydrofobowe hormony wiążą się z receptorami wewnątrzkomórkowymi, które pełnią funkcję białek regulujących geny. Chociaż cząsteczki sygnałowe tego typu różnią się budową chemiczną i funkcją, wszystkie działają za pośrednictwem wewnątrzkomórkowych białek receptorowych. Receptory te nie są identyczne, ale są ewolucyjnie spokrewnione i należą do nadrodziny receptorów hormonów steroidowych białek regulujących geny. Miejsca powstawania i funkcje tych hormonów podano w tab. 16-1 (s. 537)
Ogólne zasady sygnalizacji komórkowej
541
Wyszukiwarka
Podobne podstrony:
CCF20080601�005 Rys. 16-13. Większość cząsteczek sygnałowych wiąże się z receptorem białkowym na pow
CCF20081203�069 Rys. 13.10. Kształty i wymiary kola pasowego rowkowego: a) z jednym rowkiem, b) z ki
DSC03242 Rozluźnienie powodują: Tlenek azotu (NO), prostacyklina (PGI), prostaglandyna K (PCiK) Skur
Obraz (6) 2 Substancje regulujące szerokość naczyń krwionośnychNaczyniorozszerzaiace Tlenek azotu (N
img027 3 Rys. 2-16 O 10 kN -1,11 yw=3-78 or3-270° <y.lx.^Qf1 = 240o (xi=2, y-i=-i) (x3=-3,y3=-2)
DSC02781 TLENEK AZOTU (NO) Funkcje fizjologiczne NO w OUN: • udział w prokafnictwi
DSC02399 Rozluźnienie powodują: Tlenek azotu (NO), prostaeyklina (POI), prostaglandyna E (P(»E) Skur
DSC02400 Rozluźnienie powodują: Tlenek azotu (NO), prostacyklina (PCil), prostsiglandnu E (PGE) Skur
DSC02401 Rozluźnienie powodują: Tlenek azotu (NO), prostacy klina (PGI), prostaglamlyna E (PGE) Skur
Jakie jest działanie tlenku azotu (NO) na komórki mięśniowe? Tlenek azotu (NO) w komórkach mięśniowy
Rys. 16.3 Do tego samego wyniku można dojść, jeżeli skorzysta się ze wzoru na ugięcie f rozpatrywane
DSC03544 252525255B1024x768 252525255D 16 ROZDZIAŁ I godzin. Kompresy no bólu reumatyczne, mięśniow
skany patofizjo�9 2.16. Zawał (infarcłus) Zawał jest to martwica tkanki spowodowana niedrożnością na
więcej podobnych podstron