skanowanie0162

Cykl tyelowy komórki |Mj||fl|
Hornów. W tym stadium homnloglr/ne chromosomy (matczyne I ojeowskle) inuu ją Mly parami, następuje wymiana mit Inków chronnityd om/ synteza KNA I Prolaza I JohIi procesem ciągłym, lecz ze względu na zachodzące cha rat lei vui\ , zmiany w organizacji chromosomów podzltlono ją na 5 stadiów: I) leplnlcu, 'i hH goleń, 3) pnchyten, 4) diploten I 5) tllaklnczn (ryc, 8.2.4).
Leptoton (gr. leptos = cienki) rozpoczyna się kondensacją chromosomów. no chromatydy leżą ściśle obok siebie, tak że każdy chromosom wygląda (aldiy M złożony tylko z jednej chromatydy. Miejsca, wiążące chromatydy ściśle ze igjfl i^H krótkimi nie/repiikowanymi odcinkami DNA (ryc. 8.2.5). Pojawiają się zaczątki pn|ś; 1 czert syiuptemalnych, które w pełni rozwiną się dopiero w zygotenie. c In < nn.ii \ ,|y 1 utykają się telomerami z osłonką jądrową (ryc. 8.2.6). Ten mechanizm pomaga pi .. tlopotlobnie w uporządkowaniu chromosomów w dalszych stadiach prolazy I, i i.. -mosoiny wiążą się zazwyczaj z osłonką jądrową w jednym miejscu i tworzą h u mu 1 „bukietu". W czasie kondensacji nici DNA formują się chromomery, silnie sknndf^H suwane odcinki chromatyd, które rozdzielają odcinki słabiej skondensowane, l • i. M temu chromatydy przybierają wygląd nici, z nieregularnie nanizanymi koralll>ałH^| Koralikowy obraz chromatyd utrzymuje się aż do końca stadium diplotemi,
Zygoton (gr. zygon = łączyć) - rozpoczyna się proces łączenia się homologlc/nw^H chromosomów, czyli koniugacja. Choromosomy homologiczne, jeżeli si) oddalołjffl od siebie, zbliżają się i ustawiają w taki sposób, aby identyczne geny - a Hele Ipótlyl obok siebie. Proces łączenia się homologicznych chromosomów, czyli synapsls (git I napsts m połączenie), może brać początek w kilku miejscach, najczęściej syna puls rt)| poczyna się od lelomeru. Jeżeli jest on przyczepiony do osłonki jądrowej, to biegnie wzdłuż chromosomów, podobnie jak zasuwający się zamek blyskawlegj^H (ryc. 8.2.7). Połączenie synaptemalne powstaje pomiędzy homologicznymi chroifj^^H
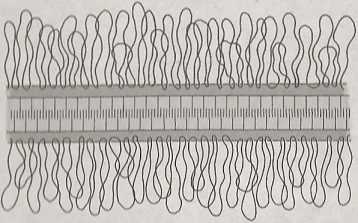
Ch
kompleks
połączeniowy
Ch
Ryc. tu ii Modol połączenia synaptemalnego. Połączenie synaptemalne powstaje pomiędzy homukąili * nyini chromosomami. Ma ono postać trójwarstwowogo pasma, w którym obecne są przebiegające |||| przecznic fllnmonty
soinaml. Kompleks synaplemalny ma posiać Irójwarslwowego pasma, w klóryul obeenu są przebiegające poprzecznie fllamenly (ryc, Połączenia synaplemalnp po opływie określonego czasu „rozwiązują się", I •zleje sly lak pod wpływem cn/y* mu lopot/omera/y II. lednyiu z elekaws/yi b /|a\v|n|», jakie /nchod/ą pode/,as zygo*
ngu I cm I proces 11 ni inl* i v | ii jl 1/ n JA, kl ńm Inicjuje knnlugin ję (dworni imoiuów,
& I<hUmm zanika, Część I )NA nle/repllkowanego w lazle 8 zoHtaje /replikowana dolin, w zygolenle I mml u fi/, w i,' • i/v; I >NA (0,1-0,2%). lii część PN A jt?nł matrycą dla p-KNA.
ISmliylon (gr, puchlin gruby) rozpoczyna się od momenlu zakończenia konluga-H|oninlogicznych chromosomów. Czas jego trwania określa się przeciętnie na kilka jui ilu tygodnia. W wyniku postępującej kondensacji chromalyny następuje skrócę-||ii i,lr | pogrubienie chromatyd, co sprawia, że chromosomy są dobrze widoczne, chromatydy zostają wyodrębnione, a połączone chromosomy homolo-jipnii, czyli biwalenty (podwójne), przybierają postać tetrad (gr. tetros = cztery), w tym sladium przebiega wymiana odcinków chromatyd, czyli proces określany Ja* ■ pri)HNlng-over (ang, wymiana na krzyż). Wymiana może zachodzić pomiędzy mpynlMmi czterema chromatydami w obrębie biwalentu. Przebieg crossing-over jest fiup||wy dzięki aktywności enzymu DNazy II, która rozcina nici DNA w miejscu, Bjtlńrym ma nastąpić wymianajej odcinków z sąsiednią chromatydą. W pachytenle Hi/i'biega również replikacja pozostałych odcinków DNA, tzw. P-DNA (DNA pachy-lęniiwy), które nie zostały zreplikowane w fazie S, i synteza białek histonowycll spe-MlfZnyeh dla mejozy. Synteza histonów wskazuje, że w profazie I musi następować lllikleoiiomowa reorganizacja chromatyny. Ponadto w pachytenie dojrzewnjąeyih filii ylów wielu zwierząt, np. płazów, obserwuje się replikację specjalnych odcinków | IMA, l/w, rDNA, który koduje informację dla RNA rybosomowego (rRNA), Wielu* Inni i iic zreplikowane odcinki rDNA przechodzą do cytoplazmy i funkcjonują lam ja jp |/,w, jąderka dodatkowe. Pod koniec pachytenu białka szkieletu osłonki jądrowe) Irtinlny (patrz rozdz. 4.2.1) ulegają reorganizacji, uwalniając związane z nimi lelo mn v, które uzyskują swobodę ruchu w całej wewnętrznej płaszczyźnie osłonki,
Diploten (gr. diplos = podwójny). W diplotenie zachodzi nadal kondensacja i lu o Bjglyny, n zatem obserwuje się dalsze skracanie i pogrubianie chromosomów. I lomo |ngii / nr chromosomy odpychają się od siebie, ale pozostają ze sobą zwlą/iiue miejscach skrzyżowania chromatyd (crossing-over). Miejsca te noszą nazwę i lilti/m, Większość połączeń synaptemalnych zanika. Pozostają jedynie bardzo małe pnląi y.enia, które można dopiero zobaczyć w obrazach mikroskopowo-eleklmmi* wyi li. I .iczba i pozycja chiazm w chromosomach jest różna dla prawie każdej komórki płciowej danego zwierzęcia, niemniej jednak na każdy chromosom przypada Hl/yntijmniej jedna chiazma. W diplotenie chromosomy odłączają się od osłonki ją di owe), jednym z najbardziej spektakularnych zjawisk jest przejściowa dekondenM-idu chromosomów, której następstwem są chromosomy szczoteczkowe (palrz pizd z, 8,2.4). Stadium diplotenu trwa najdłużej, bo rozciąga się w niektórych przypadkach na lata, o czym już wspomniano we wstępie do tego rozdziału.
Dlnklncza (złożenie gr. cliii *= poprzez, kinesis — ruch). Chromosomy .rozluźnione Iy dlplolenie powracają do pierwotnej skondensowanej formy i ponownie skracają |||, /unikają całkowicie pęlle w chromosomach szczoteczkowych, ustaje transkrypcja KNA 1 zanikają jąderka,1 limmosomy homologiczne oddalają się od siebie, W /a leżnoścl od liczby chla/in, pary chromosomów przybierają różne kształty, np, krzyża (psimi chiazma) lub wlelokioinych ósemek (liczne chłazmy) I są gotowe do pierwszego podziału mejołyc/oego
Wyszukiwarka
Podobne podstrony:
skanowanie0010 Cykl życiowy komórki i jej podział W puli i wio XIX stulecia definitywnie odrzucono h
skanowanie0152 Cykl łyclowy komórki 1
skanowanie0163 I Cykl żyelnwy tamArkl I
skanowanie0006 Ćwiczenie 6Wprowadzenie do zagadnień z genetyki. Cykl życiowy komórki. Podziały komór
skanowanie0011 !2MH Cykl lyclowy komórki I ]®j jtttfjtliłl / / / Hy< , 0,1,1. Schemat przebieg
56342 skanowanie0165 264 Cykl życiowy komórki i jej podzcf wej segregacji chromatyd. Dopiero w anafa
skanowanie0019 Schemat przepływu elektronów w łańcuchu oddechowym i związane z tym zjawiska energety
Skanowanie 10 01 12 58 (7) PAŁUBA że tym powiedzeniem mógłby wyzwolić w sobie sferę różnych apetytó
skanowanie0012 (62) J2 li-A 1P A A /fł 4- 1 Z i 1 2 tf 4 3 ■iliMi ~ fs® ^ 4- Wm. i WM» c> k,
skanowanie0027 (47) przez komórki prokambium, w których zanika wówczas aktywność genów markerowych t
skanowanie0028 (24) ; r * • * * • i ~s r y- i ’ * ^ * I Tl I *! : • .; • :.* ■ " . : ( ?vt 4 i
więcej podobnych podstron