
Oporność drobnoustrojów na antybiotyki
prof. Marcin Świtała
Materiał pomocniczy do ćwiczeń z przedmiotu
farmakologia weterynaryjna (2010/11)
Katedra Biochemii, Farmakologii i Toksykologii
Uniwersytetu Przyrodniczego we Wrocławiu

Najprostszym typem oporności bakterii na
chemioterapeutyk jest oporność naturalna.
Oznacza ona całkowitą niewrażliwość danego
gatunku drobnoustroju na chemioterapeutyk.
Typowym przykładem oporności naturalnej jest
brak wrażliwość drobnoustojów beztlenowych na
działanie antybiotyków aminoglikozydowych.
W praktyce lekarskiej najczęściej o oporności drobno-
ustrojów patogennych mówi się wtedy, kiedy średnie
stężenia hamujące populację drobnoustrojów in vitro są
większe od stężeń możliwych do uzyskania in vivo.
Oznacza to, że stosując dawkę leczniczą antybiotyku
lekarz nie może uzyskać efektu terapeutycznego, gdyż
osiągane stężenie leku w tkankach jest
niewystarczające do zahamowania wzrostu populacji
patogennego drobnoustroju lub do jej zabicia.
W takim ujęciu mowa jest zawsze o tzw oporności nabytej,
która rozwinęła się w populacji dotąd wrażliwego patogenu
- w trakcie jego stosowania.
Ze względu na mechanizm powstania oporność
nabyta może występować w dwóch postaciach :
jako odporność nabyta p i e r w o t n a lub
jako oporność nabyta w t ó r n a

Oporność nabyta p i e r w o t n a powstaje w wyniku
spontanicznej mutacji szczepu należącego do gatunku
wrażliwego na chemioterapeutyk.
Może się ona pojawiać bez kontaktu z chemioterapeutykami.
Jest to tzw oporność chromosomalna gdyż jest zakodowana
genetycznie w chromosomach
Nie jest przekazywana na inne gatunki drobnoustrojów.
Częstość występowania mutantów w populacjach
bakteryjnych jest niewielka.
Oporność pierwotna występuje stosunkowo rzadko, co
sprawia, że ten typ oporności ma mniejsze znaczenie
Nie wygasa - w odróżnieniu od oporności wtórnej - po
zaprzestaniu stosowania chemioterapeutyku.

Oporność w t ó r n a rozwija się zawsze w warunkach
kontaktu drobnoustroju z chemioterapeutykiem.
Mechanizmy prowadzące do jej powstania mają 2 składowe:
• g e n e t y c z n ą (przeniesienie oporności) oraz
• b i o c h e m i c z n ą (sposób jej realizowania).
Jest ona nazywana opornością pozachromosomalą gdyż
mechanizm genetyczny leżący u jej podłoża jest związany z
genami znajdującymi się w cytoplazmie, poza chromoso-
mami, w strukturach zbudowanych z DNA zwanych
p 1 a z m i d a m i lub c z y n n i k a m i R (resistance factor)
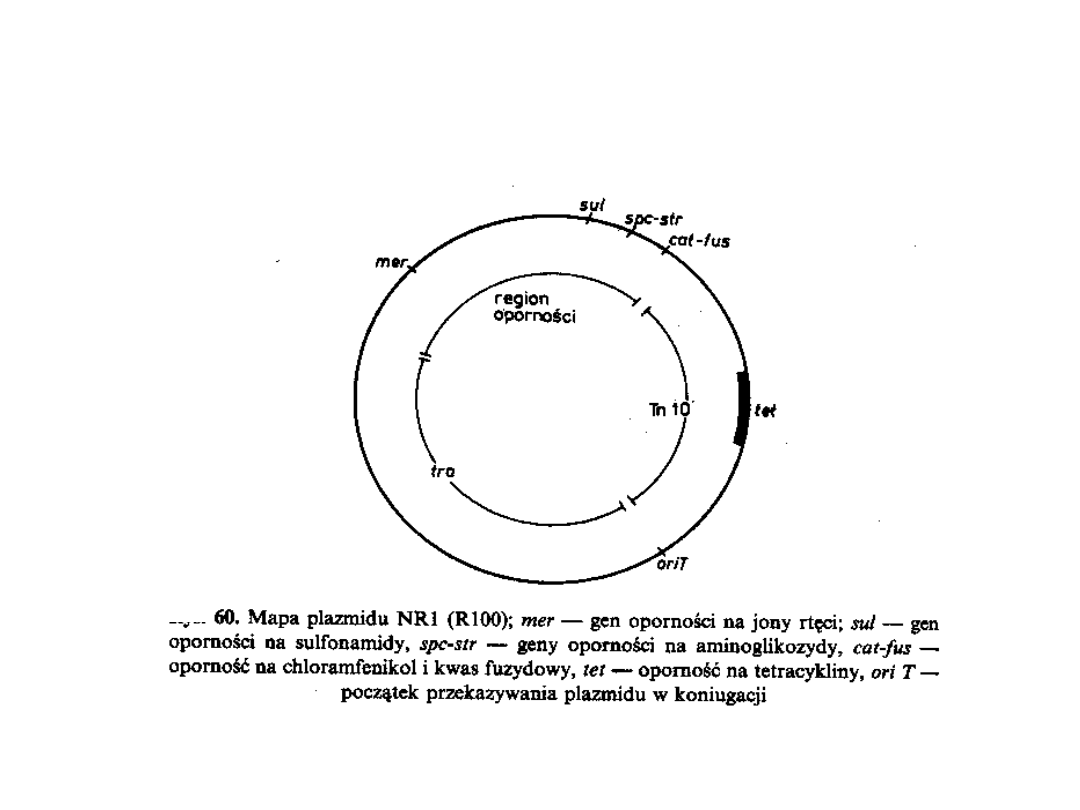
P l a z m i d y stanowią elementy dziedziczności
pozachromosonowej u większości bakterii, które mogą
przekazywać cechy genetyczne nie tylko w obrębie
danego gatunku bakterii, ale także pomiędzy
mikroorganizmami należącymi do różnych gatunków.
Przenoszenie lekooporności jest związane z
konkretnymi genami leżącymi w obrębie plazmidu lub
chromosomu.
Specjalną odmianą plazmidów są e p i z o m y, które
mogą wtórnie łączyć się z chromosomami,”wymusić”
replikację części genów zawartych w chromosomie i
przenieść zreplikowany materiał do innej komórki
Jeden plazmid może zawierać geny oporności
przeciwko kilku chemioterapeutykom.

Innymi czynnikami warunkującymi genetyczne
przenoszenie są - t r a n s p o z o m y.
Są to niewielkie sekwencje DNA, które mogą być
przenoszone z jednego plazmidu lub chromosomu
na inny.
Mogą one nieść geny oporności dla licznych
antybiotyków, nie wymagają specjalnego miejsca,
wbudowywania.
Transpozomy odgrywają istotną rolę w rozwoju i
szerzeniu się oporności bakterii w warunkach
szpitalnych.
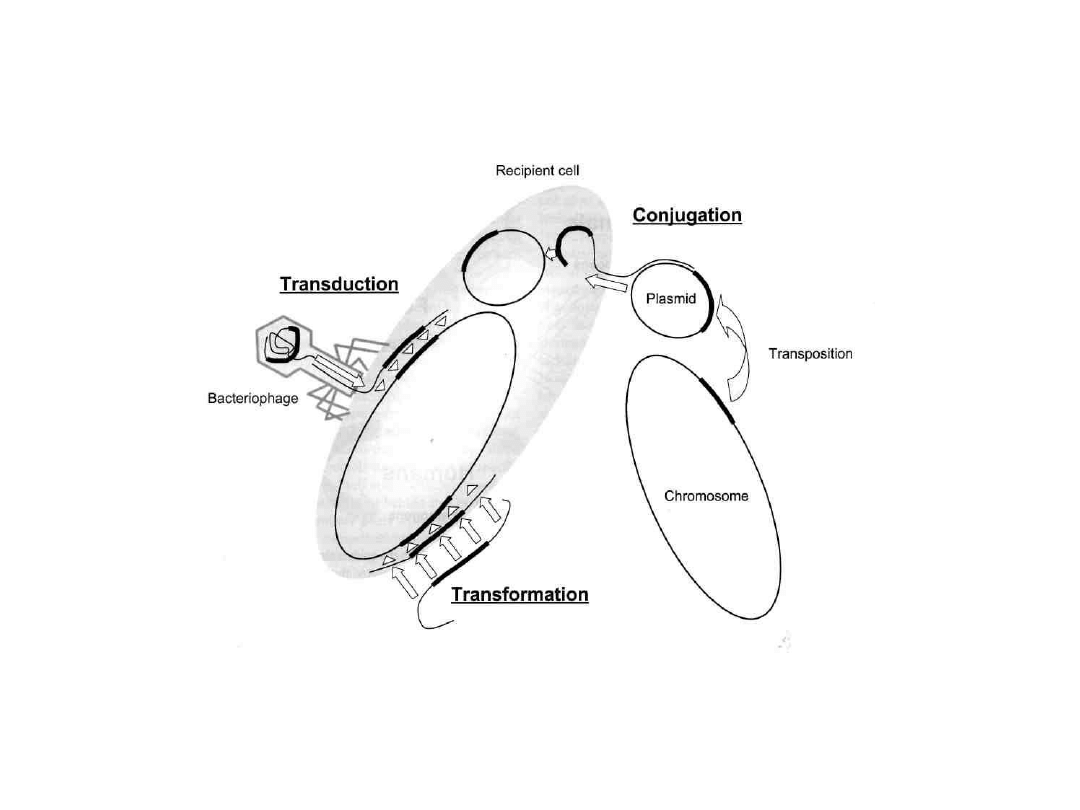
Przekazywanie plazmidów pomiędzy
komórkami bakteryjnymi może się odbyć
• drogą k o n i u g a c j i
• drogą t r a n s d u k c j i.
• drogą transformacji

W k o n i u g a c j i przekazanie plazmidów
zachodzi w czasie bezpośredniego połączenia dwóch
komórek (pilus) za pomocą nici białkowych.
Szczególnie niekorzystne jest przekazywanie w ten sposób
oporności z bakterii saprofitujących w przewodzie pokarmowym
na patogenne enterokoki.
plazmid
chromosom
pilus
dawca
biorca
Zjawisko to
występuje głównie
wśród bakterii
Gram-ujemnych.
KONIUGACJA
T r a n s d u k c j a
jest procesem przekazywania
plazmidów z komórki dawcy (donora) na komórkę
biorcy (akceptora) przez fagi ustrojowe
(bakteriofagi).
Proces jest swoisty gatunkowo.
Przykładem może być przenoszenie plazmidów
warunkujących wytwarzanie niektórych B-laktamaz w
obrębie szczepów gronkowcowych.
Mechanizmy biochemiczne , przez które najczęś-
ciej realizowane są procesy odpornościowe to:
1) zmniejszenie przenikania chemioterapeutyku przez
błonę komórkową;
2) unieczynnienie enzymatyczne chemioterapeutyku,
3) czynne “wypompowywanie" chemioterapeutyku z
komórki bakteryjnej.
4) zmiany w ilości lub konformacji receptora (lub/i jego
otoczenia) dla chemioterapeutyku;

W zależności od szybkości rozwoju oporności
wyróżnia się :
a) typ jednostopniowy oporności - gdy powstaje ona
po pierwszej ekspozycji na antybiotyk. Przyczyną jest
szybkie przekazywanie czynników oporności.
Jest to tzw. typ streptomycynowy gdyż rozwój opor-
ności na streptomycynę rozwija się bardzo szybko.
b) typ wielostopniowy oporności - gdy rozwija się
ona powoli, po wielokrotnej ekspozycji.
Przykładem może być rozwój oporności na
benzylopenicylinę (stąd nazwa typ penicylinowy).

Rozwój oporności na jeden chemioterapeutyk może
powodować oporność na inne chemioterapeutyki tej
samej grupy
Jest to tzw. o p o r n o ść k r z y ż o w a.
Oporność krzyżowa może być całkowita - gdy
dotyczy całej grupy lub częściowa.
Gdy rozwój oporności na chemioterapeutyk pociąga
wzrost oporności wśród chemioterapeutyków o
zbliżonym mechanizmie działania mówimy wówczas
o o p o r n o ś c i r ó w n o l e g ł e j.

Oporność
na antybiotyki beta-laktamowe
Najważniejszym mechanizmem oporności na
te leki jest wytwarzanie swoistych enzymów
tzw. beta-laktamaz, które wskutek hydrolizy
otwierają pierścień beta-laktamowy, całko-
wicie znosząc aktywność tych antybiotyków
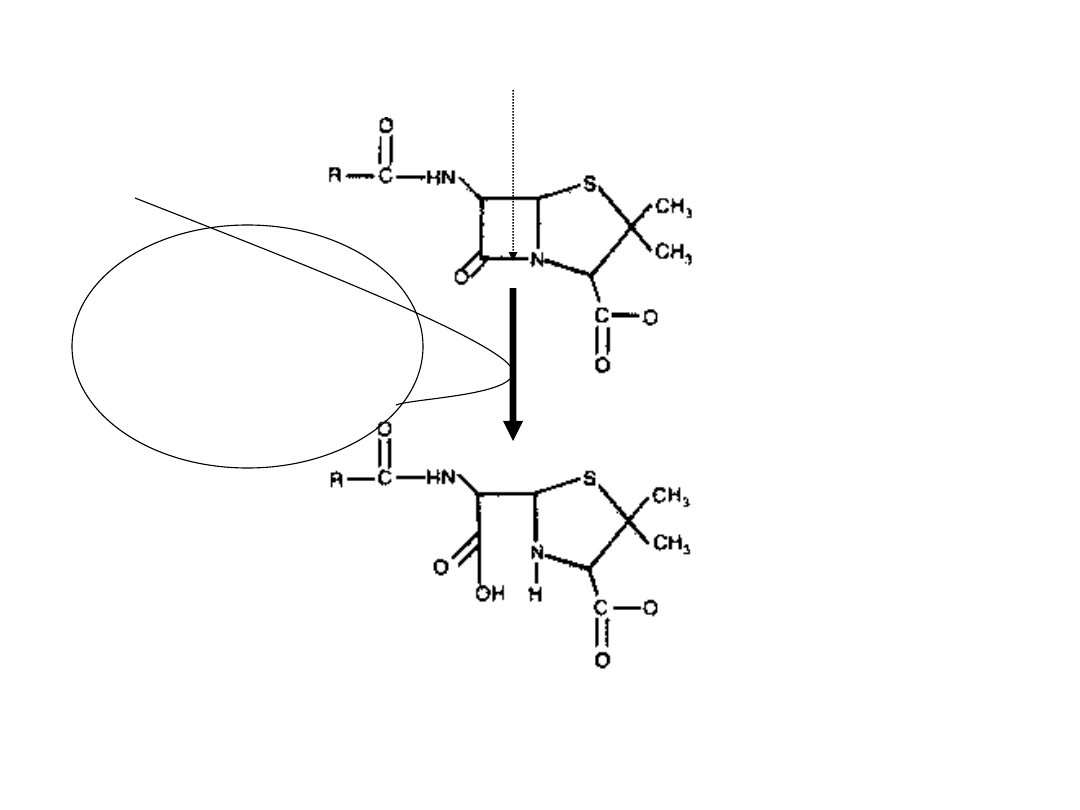
Beta-
laktamaza
wiązanie beta-laktamowe
kwas penicylinowy
penicylina
H
2
O

W świecie bakterii istnieje duża różnorodność
beta-laktamaz zróżnicowanych pod względem:
•budowy (z centrum serynowym lub z centrum
cynkowym),
•aktywności enzymatycznej (klasy enzymów TEM, OKSA,
PSE, TLE, SHV i inne) ,
•pochodzenia (grokowcowe, enterokokowe i in.)
•sposobu zakodowania (chromosomalne i plazmidowe)
•aktywnyności wobec różnych klas antybiotyków
(penicylinazy, cefalosporynazy)
•sposobu produkcji (konstytucjonalne lub indukowane)
•czynników inhibicyjnych
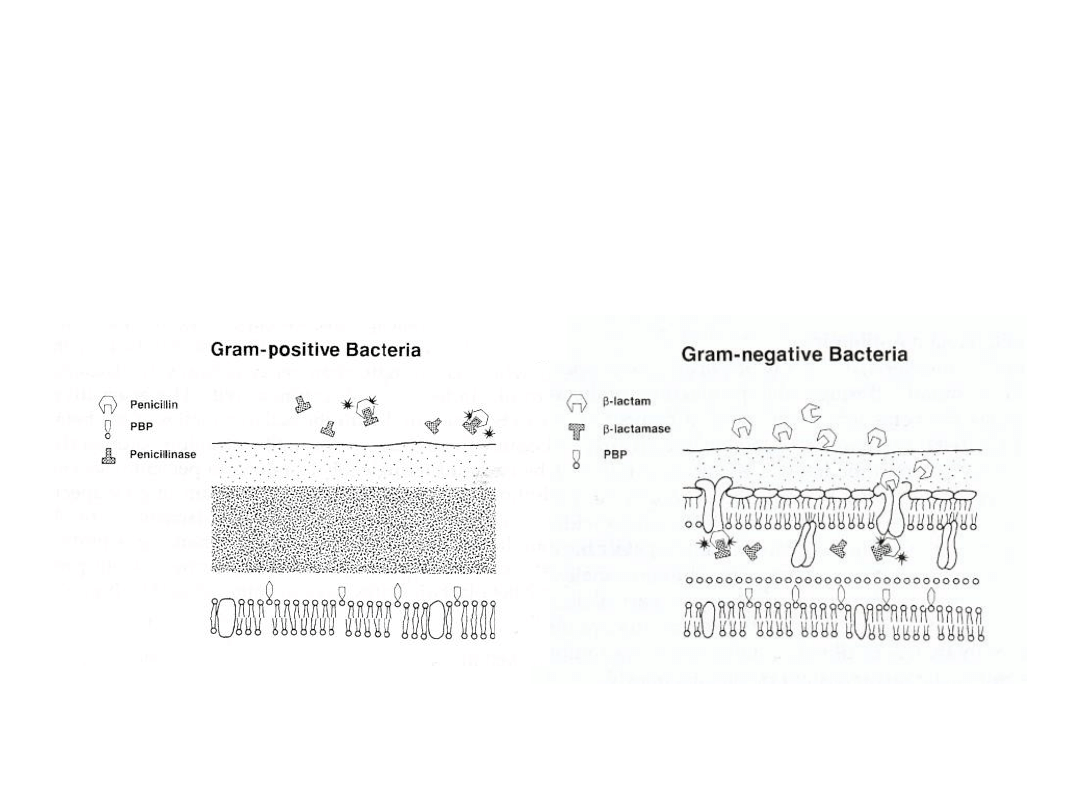
Beta-laktamazy produkowane przez bakterie Gram-
dodatnie są wydalone poza komórkę i działają w
bezpośrednim środowisku, natomiast produkowane przez
bakterie Gram ujemne w przestrzeni periplazmatycznej
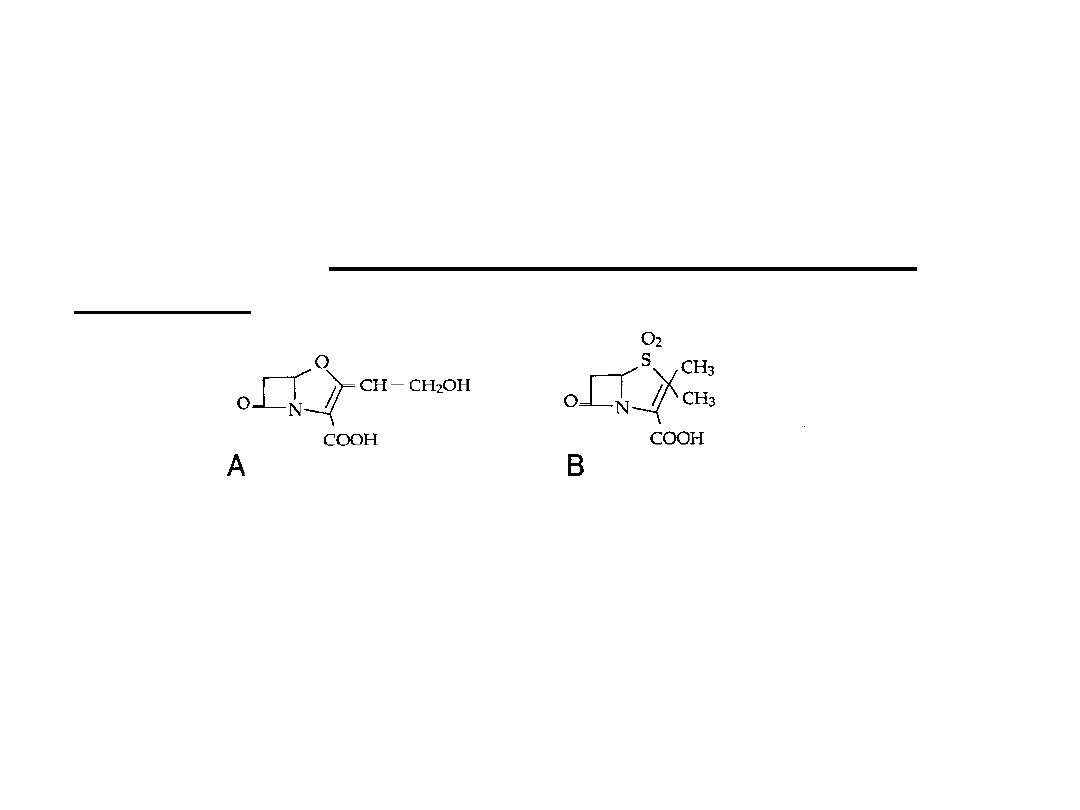
Obecnie w lecznictwie stosuje się na szeroką skalę inhibitory
beta-laktamaz, które podaje się łącznie z niektórymi
penicylinami lub cefalosporynami.
Należą do nich: kwas klawulanowy(A), sulbaktam (B),
tazobactam
W weterynarii znane są następujące kombinacje leków:
kwas klawulanowy+amoksycylina
sulbaktam+ampicylina
kwas klawulanowy+ tikarcylina tazobaktam+piperacylina

Oporność
na antybiotyki aminoglikozydowe
Oporność na te antybiotyki jest głównie wynikiem działania
związanych z plazmidami enzymów modyfikujących lek.
Enzymy te, do których należą acetylotransferazy(AAC),
fosfotransferazy (APH) i nukleotydylotransferazy(AAD),
modyfikują cząsteczkę antybiotyku w miejscu grup
aminowych lub hydroksylowych.
Do tych trzech grup należy wiele enzymów, które różnią się
znacznie
swoim
zakresem
działania
i
stopniem
unieczynniania różnych aminoglikozydów.
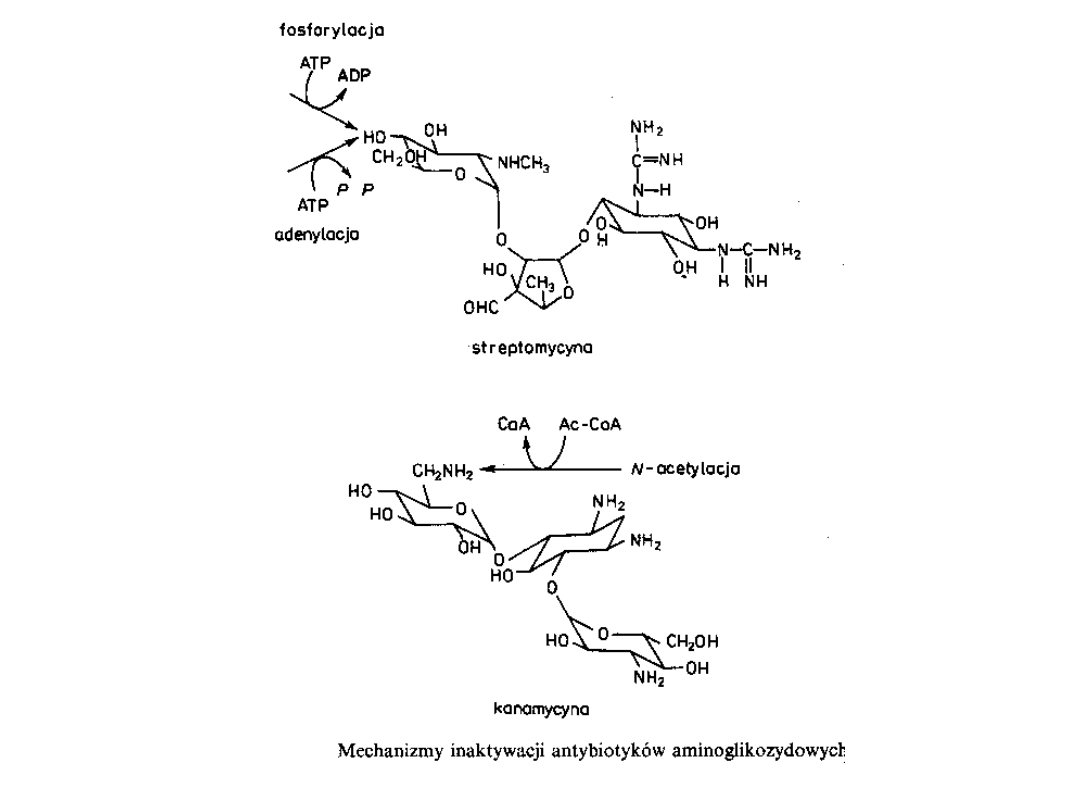

Enzymy modyfikujące aminoglikozydy i aminocyklitole

Oporność
na kolistynę
Oporność na kolistynę wśród bakterii
Gram-ujemnych powstaje rzadko.
Pojawienie się oporności na kolistynę jest
związane ze spadkiem przepuszczalności
błony komórkowej bakteryjnej dla leku.

Oporność
na fluorochinolony
Oporność na fluorochinoliny może powstawać
trzema drogami:
1. Spadek przepuszczalności błony zewnętrznej dla
leku w związku z modyfikacją por hydrofilowych
2. „Wypompowanie” leku z komórki przez
powstanie mechanizmu aktywnego transportu
3. Zmiana mutacyjna miejsca wiązania leku z
gyrazą DNA lub tomoizomerazą IV

Oporność
na tetracykliny
Oporność na tetracykliny związana jest z
następującymi mechanizmami
1. Wbudowaniem w błonę komórkową systemu
transportowego odpowiedzialnego za aktywne
„wypompowanie” leku z komórki
2. Zmiana mutacyjna miejsca wiązania leku z
rybosomem
Oporność ma charakter krzyżowy

Oporność
na kombinację sulfonamid-
trimetoprim
Oporność na trimetoprim jest zwykle związana z
kodowaną w plazmidzie syntezą nowej reduktazy
dihydrofolianu, która jest znacznie mniej wrażliwa na
trimetoprim niż forma występująca naturalnie.
Podobnie zależna od plazmidu oporność na
sulfonamidy często jest następstwem pojawienia się
nowej syntetazy dihydropteronianu, o zmniejszonym
znacznie powinowactwie do sulfonamidów, a nie do
kwasu p-aminobenzoesowego.

Oporność jest zwykle związana z wytwarza-
niem przez drobnoustrój acetylotransferaz,
które modyfikują i inaktywują leki z tej grupy.
W wyniku działania tego enzymu powstaje
mono- lub dioctan (chloramfenikol lub
tiamfenikol) lub tylko monooctan (florfenikol).
Oporność plazmidowa związana ze spadkiem
przepuszczalność komórki bakteryjnej dla leku
jest bardzo rzadka.
Oporność
na fenikole

Oporność plazmidowa na te antybiotyki jest
związana z modyfikowaniem miejsca wiązania
leku na rybosomie.
Jest duże prawdopodobieństwo powstania
oporności krzyżowej pomiędzy makrolidami a
linkosamidami i pleuromutylinami.
Oporność
na makrolidy i linkosamidy
Wyszukiwarka
Podobne podstrony:
biologia zakres materiau na egz Nieznany (2)
5 Wplyw dodatkow na recyklingu Nieznany
PISEMNY EGZAMIN TESTOWY NA STOP Nieznany
Aerobik Step aerobik na laweczk Nieznany (2)
Formowanie ladunkow na srodkach Nieznany
Leki wplywajace na mechanizmy k Nieznany
,fizyka 1 C, dzialania na wekto Nieznany (2)
2 Prognozowanie na podstawie s Nieznany (2)
GENETYKA DROBNOUSTROJOW NA EGZAMIN popr
7 Sterowanie logiczne na bazie Nieznany
FOLIE porz udek na strzelnicy i Nieznany
38 stron efektow zzo na obstet Nieznany (2)
330 przepisow na ciasta?likat Nieznany
Salatka z krewetkami NA RAUSZU Nieznany
330 przepisow na ciasta?bka z Nieznany
Wniosek o wyrazenie zgody na uz Nieznany
Wniosek o zezwolenie na wylaczn Nieznany
330 przepisow na ciasta?nanow Nieznany
więcej podobnych podstron