Medycyna Wet. 2006, 62 (3)
311
Praca oryginalna
Original paper
Etiopatogeneza powstawania torbielowatego roz-
rostu gruczo³ów b³ony luzowej i zapalenia b³ony lu-
zowej macicy u suk nie jest do koñca wyjaniona. Z³o-
¿ona regulacja hormonalna funkcji jajników z równo-
czesnym bezporednim oddzia³ywaniem tej gonady na
czynnoæ i strukturê macicy nastrêcza du¿e trudnoci
w okreleniu poszczególnych czynników odpowie-
dzialnych za wyst¹pienie zmian chorobowych w ma-
cicy. Dotychczas wykonano liczne badania maj¹ce na
celu wyjanienie typu zaburzeñ hormonalnych, których
efektem jest wyst¹pienie zmian w macicy suk w po-
staci rozrostu torbielowatego gruczo³ów b³ony luzo-
wej oraz ropnego zapalenia b³ony luzowej macicy
(hyperplasia cystica (pyometra complex)), lecz etiolo-
gia nadal nie jest do koñca poznana. Tak¿e próby eks-
perymentalnego wywo³ania zmian typowych dla tego
zespo³u chorobowego przez podanie egzogennych es-
trogenów i progesteronu nie wyjani³y do koñca etio-
patogenezy tego procesu. W zwi¹zku z tym mo¿na
przypuszczaæ, i¿ etiopatogeneza tak czêsto wystêpu-
j¹cych zmian chorobowych w macicy suk jest bardziej
z³o¿ona.
Rozwój zmian chorobowych w macicy bez w¹tpie-
nia stanowi efekt zaburzeñ funkcji jajników. Produ-
kowane w jajnikach hormony reguluj¹ funkcjê komó-
rek macicy za porednictwem obecnych w j¹drach tych
komórek swoistych receptorów dla estrogenów (ER)
i progesteronu (PR). Ta zale¿noæ funkcjonalna znaj-
duje swoje odbicie w fizjologicznych zmianach struk-
tury macicy zachodz¹cych w cyklu, jak i w stanach
patologicznych. Mo¿na wiêc przypuszczaæ, ¿e pra-
wid³owe funkcjonowanie jajników ma zasadnicze
znaczenie dla zachowania homeostazy komórkowej
w macicy.
Celem badañ by³o okrelenie zmian w strukturze
jajników u suk z chorobami macicy oraz okrelenie
charakteru zaburzeñ hormonalnych w oparciu o bada-
nie ekspresji receptorów dla estrogenów (ER), proge-
steronu (PR) oraz alfa-inhibiny. Spodziewane wyniki
badañ mog¹ stanowiæ przyczynek do poznania etiopa-
togenezy zespo³u torbielowatego rozrostu b³ony lu-
zowej macicy i ropomacicza u suk.
Materia³ i metody
Materia³ do badañ pochodzi³ od 61 suk, w wieku od 2,5
do 7 lat, które nale¿a³y do ró¿nych ras i zosta³y poddane
chirurgicznemu zabiegowi usuniêcia jajników i macicy
(owariohisterektomii) z powodu klinicznie stwierdzonych
chorób macicy. Kontrolê stanowi³a grupa 5 suk, u których
równie¿ wykonano owariohisterektomiê i makroskopowo
nie stwierdzono obecnoci zmian chorobowych. Zwierzêta
podzielono na trzy grupy:
A (n = 45) o wysokim stopniu zaawansowania choro-
by; B (n = 16) w mniejszym stopniu nasilenia zmian cho-
robowych; oraz C (n = 5) zdrowych suk, które stanowi³y
grupê kontroln¹.
Zmiany strukturalne w jajnikach suk
z chorobami macicy
MARIA KATKIEWICZ, ZDZIS£AW BORYCZKO
Katedra Nauk Klinicznych Wydzia³u Medycyny Weterynaryjnej SGGW, ul. Nowoursynowska 159c, 02-776 Warszawa
Katkiewicz M., Boryczko Z.
Structural ovarian changes in bitches with uterus diseases
Summary
The aim of this study was to investigate the ovaries structure in bitches with HC/pyometra complex
disease. The description of the hormonal disturbances responsible for the etiopathogenesis of this disease
syndrome were based on the expression of the nuclear ER and PR receptors and the cytoplasm alfa-inhibin
expression. The microscopic examination of ovaries of bitches with uterus diseases revealed the presence of
the simultaneous changes in the ovarian intertitium. This was manifested by the appearance of the so called
secondary interstitial cells. The main features of these cells microscopic structure were the yellowish, lipid
loaded cytoplasm. Their nuclei expressed PR receptors or ER receptors, and all of them expressed faint
cytoplasm alfa-inhibin receptors. The number of the so called secondary interstitial cells increased with the
degree of uterus pathological lesions. The hypothetic mechanism of the endocrine disturbances as a cause of
ovarian changes as well as their supposed relation to etiopathogenesis of endometrial cystic hyperplasia and
pyometra complex in bitches were discussed.
Keywords: bitches, endometritis

Medycyna Wet. 2006, 62 (3)
312
Wycinki rogów macicy zawsze pobierano w po³owie d³u-
goci rogu, oba jajniki utrwalano w 10% formalinie bufo-
rowanej fosforanami, zatapiano w parafinie. Skrawki mi-
krotomowe barwiono nastêpuj¹cymi metodami: przegl¹do-
w¹ hematoksylin¹ i eozyn¹ oraz metod¹ histochemiczn¹
AB/PAS w celu uwidocznienia glikozaminoglikanów
(GAG).
Reakcje immunocytochemiczne wykonano na skrawkach
parafinowych wybranych jajników, naklejonych na szkie³-
ka pokryte Silanem, w celu wykrycia ekspresji receptorów:
progesteronu (PR) przeciwcia³a monoklonalne firmy
Novocastra NCL-PGR (clone 1 A6), estrogenów (ER)
przeciwcia³a monoklonalne firmy Novocastra NCL-ER-
-LH2 (clone CC4-5), inhibiny alfa przeciwcia³a mono-
klonalne firmy Serotec Inhibin alfa subunits R1 MCA 951 S.
Wyniki badañ i omówienie
W badaniu makroskopowym macicy u 59 suk gru-
py A i B stwierdzono typowe zmiany dla ropomaci-
cza, to jest: powiêkszenie narz¹du, przekrwienie i zgru-
bienie ciany macicy, a w wietle wysiêk ropny lub
posokowaty. W 2 przypadkach ciana macicy by³a cien-
ka, a w wietle znajdowa³ siê p³yn luzowo-surowiczy.
W grupie C, suk kontrolnych nie stwierdzono widocz-
nych makroskopowo zmian w budowie macicy.
Badanie mikroskopowe macicy. Grupa A. W wy-
cinkach rogu macicy 45 suk tej grupy stwierdzano
typowe zmiany strukturalne dla zespo³u rozrostu tor-
bielowatego/ropomacicza, a mianowicie: proliferacjê
gruczo³ów endometrium z towarzysz¹cymi zmianami
torbielowatymi, proliferacjê nab³onka powierzchow-
nego endometrium, transformacjê nab³onków w fazê
sekrecji, co manifestowa³o siê wystêpowaniem jasnej
cytoplazmy w komórkach nab³onka gruczo³owego
i powierzchownego. W przypadkach, kiedy obserwo-
wano wysoki stopieñ nasilenia rozrostu komórek na-
b³onka endometrium, przybiera³y one wygl¹d komó-
rek jasnych tak okrelanych z uwagi na budowê
cytoplazmy. W zrêbie endometrium stwierdzano ognis-
kowe, komórkowe nacieki zapalne, które g³ównie by³y
z³o¿one z komórek monoj¹drowych, natomiast na po-
wierzchni nab³onka powierzchownego i w wietle gru-
czo³ów dodatkowo obecne by³y bardzo liczne gra-
nulocyty obojêtnoch³onne. W sporadycznych przypad-
kach stwierdzano zapalenie g³êbszych warstw ciany
macicy metritis, a w jednym przypadku wystêpowa-
³o adenomyosis. Zmiany mikroskopowe w macicy suk
tej grupy sumarycznie przedstawiono w tab. 1. W bar-
wieniu histochemicznym AB/PAS wykazano, ¿e wy-
dzielina zawarta w torbielach gruczo³owych zawiera
ró¿ne typy GAG. Przewaga silnie AB+ wydzieliny
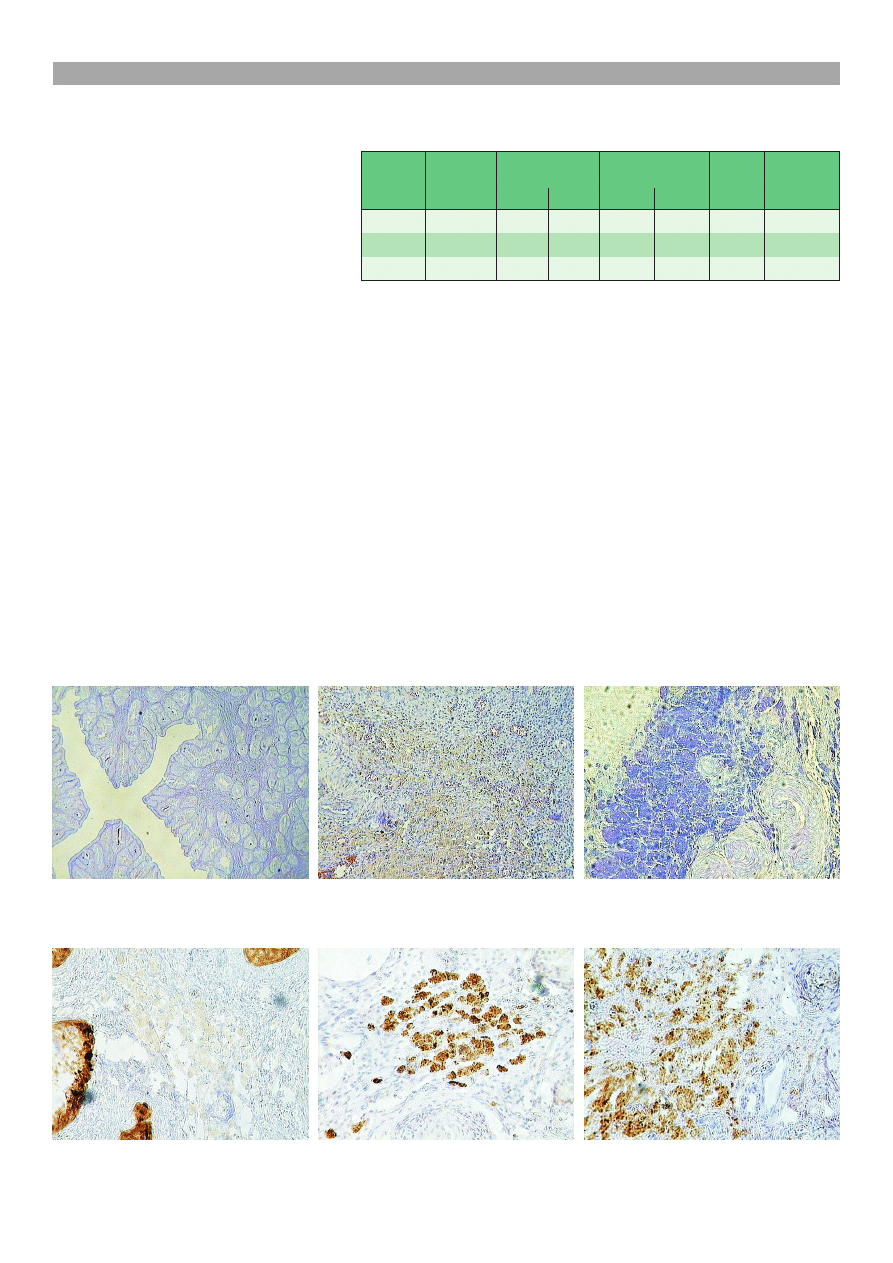
(ryc. 1) dowodzi nadmiaru wydzielanych kwanych
GAG, natomiast wydzielina PAS+ wiadczy o prze-
wadze wydzielania obojêtnych GAG (ryc. 2).
Grupa B. W tej grupie chorych suk (n = 16) zmiany
mikroskopowe w macicy ogranicza³y siê do wystêpo-
wania tylko niektórych z opisanych zmian histopato-
logicznych typowych dla zespo³u rozrostu torbielowa-
tego endometrium i ropomacicza (tab. 1). Z wyj¹tkiem
dwóch suk, u których stwierdzono w³óknienie ciany
macicy (fibrosis endometrium) i luzomacicze (mu-
cometra) we wszystkich pozosta-
³ych przypadkach obserwowano
proliferacjê gruczo³ów endome-
trium oraz powiêkszenie siê ich
wiat³a. Komórki nab³onkowe en-
dometrium ulega³y transformacji
w fazê wydzielnicz¹. W barwieniu
AB/PAS uzyskano podobne rezul-
taty, jak u suk grupy A. U suki
z du¿ego stopnia zanikiem endome-
trium stwierdzono obfity naciek za-
palny w zrêbie endometrium, z³o¿o-
ny g³ównie z komórek monoj¹dro-
wych. Jednak¿e, mimo wyranego
zaniku endometrium, komórki na-
Tab. 1. Typy zmian histologicznych stwierdzonych w macicy suk chorych (n=61) w porównaniu z prawid³ow¹ struktur¹
mikroskopow¹ macicy suk grupy kontrolnej (n=5)
a
p
u
r
G
a
j
c
a
r
e
fi
l
o
r
P
w
ó
³
o
z
c
u
r
g
y
n
o
³
b
j
e
w
o
z
u
l
e
l
e
i
b
r
o
T
w
ó
³
o
z
c
u
r
g
y
n
o
³
b
j
e
w
o
z
u
l
a
j
c
a
m
r
o
f
s
n
a
r
T
w
ó
k
n
o
³
b
a
n
ê
z
a
f
w
¹
n
j
y
c
e
r
k
e
s
w
ó
k
n
o
³
b
a
n
a
j
c
a
r
e
fi
l
o
r
P
j
e
n
l
a
n
o
j
c
k
n
u
f
i
c
ê
z
c
w
y
n
l
a
p
a
z
k
e
i
c
a
N
y
n
o
¿
o
³
z
k
e
r
ó
m
o
k
z
h
c
y
w
o
r
d
¹
j
o
n
o
m
y
n
p
o
R
k
e
i
c
a
n
y
n
l
a
p
a
z
s
it
ir
t
e
M
e
n
n
I
o
g
e
³
a
m
a
i
n
p
o
t
s
o
g
e
¿
u
d
a
i
n
p
o
t
s
)
5
4
=
n
(
A
5
4
=
n
5
4
=
n
5
4
=
n
0
5
4
=
n
0
5
4
=
n
3
=
n
s
i
s
o
y
m
o
n
e
d
A
1
=
n
)
6
1
=
n
(
B
5
1
=
n
5
1
=
n
9
=
n
9
=
n
0
6
=
n
3
=
n
0
s
i
s
o
r
b
if
1
=
n
a
rt
e
m
o
c
u
m
1
=
n
m
u
ir
t
e
m
o
d
n
e
a
i
h
p
o
rt
a
1
=
n
)
5
=
n
(
C
0
0
0
0
0
0
0
0
0
Ryc. 1. Macica suki z grupy A. W torbie-
lach gruczo³owych przewaga wydzieliny
AB+, kwanych GAG. Reakcja AB/PAS
× 10
Ryc. 2. Macica suki grupy A. W torbie-
lach gruczo³owych wydzielina o przewa-
dze PAS+, obojêtnych GAG. Ab/PAS
× 10
Medycyna Wet. 2006, 62 (3)
313
b³onka powierzchownego wykazywa³y
tak¿e cechy transformacji w fazê sekre-
cyjn¹. Ogólnie mówi¹c zmiany choro-
bowe w obrazie mikroskopowym by³y
u suk tej grupy znacznie mniej zaawanso-
wane w porównaniu do suk grupy A, lecz
typ zmian by³ analogiczny.
Grupa C. W badaniu mikroskopowym
wycinków rogów macicy suk tej grupy
(n = 5) stwierdzano prawid³ow¹ budowê
endometrium, charakterystyczn¹ dla fazy
lutealnej cyklu p³ciowego. Aktywnoæ wydzielnicza
komórek macicy w fazie lutealnej jest niska i charak-
teryzuje siê sk¹p¹ iloci¹ wydzieliny w wietle gru-
czo³ów, które mia³y cechy GAG kwanych, AB+ lub
mieszanej wydzieliny AB/PAS+ (ryc. 3).
W badaniu makroskopowym jajników w 11 przy-
padkach stwierdzano obecnoæ ma³ych torbieli jaj-
nikowych, których charakter zosta³ dok³adnie okrelo-
ny w badaniu mikroskopowym i przedstawiony w ta-
beli 2, podobnie jak obecnoæ cia³ek ¿ó³tych w jajni-
kach.
Badanie mikroskopowe jajników suk. Grupa A.
Zmiany mikroskopowe w jajnikach suk tej grupy
przedstawiono w tab. 2. Dominuj¹c¹ cech¹ stwierdzon¹
w jajnikach suk tej grupy by³a obecnoæ w zrêbie pól
komórek cechuj¹cych siê blado¿ó³t¹, ob³adowan¹ li-
pidami cytoplazm¹ i okr¹g³ymi j¹drami (ryc. 4). Ko-
mórki te g³ównie by³y zlokalizowane w czêci rdzen-
nej jajnika, wokó³ du¿ych naczyñ krwiononych, lecz
w przypadkach du¿ej liczebnoci nacieka³y czêæ
korow¹ jajnika. W tych przypadkach obserwowano
du¿e pola zrêbu jajnika utworzone przez ten typ ko-
mórek. Niekiedy komórki te tworzy³y w¹skie pasma
widoczne w os³once pêcherzykowej torbieli jajniko-
wych lub wokó³ cia³ka ¿ó³tego. Komórki te okrelono
mianem komórek wtórnych zrêbu.
W barwieniu AB/PAS komórki wtórne zrêbu bar-
wi³y siê na kolor ciemnogranatowy (ryc. 5). Komórki
te wykazywa³y cieñ ekspresji alfa inhibiny (ryc. 6),
a tak¿e stwierdzano w nich ekspresjê j¹drowego re-
ceptora progesteronu PR (ryc. 7) lub receptora dla es-
trogenów ER (ryc. 8). W jajnikach suk tej grupy stwier-
dzano tak¿e obecnoæ, mniej licznych du¿ych, a czê-
ciej mikroskopowej wielkoci torbieli pêcherzykowych
oraz jednego lub kilku cia³ek ¿ó³tych. W czterech przy-
padkach stwierdzono obecnoæ pêcherzyków wieloja-
jowych. Na szczególne podkrelenie zas³uguje tak¿e
fakt wystêpowania rozrostu komórek nab³onka jajni-
Tab. 2. Charakter zmian w strukturze mikroskopowej jajników suk z choro-
bami macicy (n=61) oraz suk grupy kontrolnej (n=5)
Objanienia: ++++/+++++ bardzo liczne; +++ liczne
a
p
u
r
G
i
k
r
ó
m
o
K
e
n
r
ó
t
w
u
b
ê
r
z
e
l
e
i
b
r
o
T
e
w
o
k
y
z
r
e
h
c
ê
p
e
l
e
i
b
r
o
T
e
w
o
k
n
o
³
b
a
n
o
k
³
a
i
C
e
t³
ó
¿
i
k
y
z
r
e
h
c
ê
P
e
w
o
j
a
j
o
l
e
i
w
e
³
a
m
e
¿
u
d
e
³
a
m
e
¿
u
d
)
5
4
=
n
(
A
+
+
+
+
+
/
+
+
+
+
3
=
n
6
=
n
0
4
=
n
5
=
n
3
4
=
n
5
=
n
)
6
1
=
n
(
B
+
+
+
9
=
n
9
=
n
2
1
=
n
)
5
=
n
(
C
e
z
c
n
y
d
e
j
o
p
2
=
n
5
=
n
Ryc. 6. Jajnik suki z grupy A. Ekspresja
receptora alfa-inhibiny w cytoplazmie
wtórnych komórek zrêbu. IPOX × 40
Ryc. 7. Jajnik suki z grupy A. Ekspresja
receptora progesteronu (PR) w j¹drach
wtórnych komórek zrêbu. IPOX × 40
Ryc. 8. Jajnik suki z grupy A. Ekspresja
receptora estrogenów (ER) w j¹drach
wtórnych komórek zrêbu. IPOX × 40
Ryc. 3. Macica suki grupy C, kontrolnej.
Widoczna sk¹pa sekrecja zarówno kwa-
nych, jak i obojêtnych GAG. AB/PAS × 10
Ryc. 4. Wtórne komórki zrêbu w jajniku
suki z grupy A, cechuj¹ce siê blado¿ó³-
tym zabarwieniem cytoplazmy. HE × 10
Ryc. 5. Jajnik suki z grupy A. Reakcja
AB/PAS w wtórnych komórkach zrêbu.
AB/PAS × 20

Medycyna Wet. 2006, 62 (3)
314
kowego (ryc. 9). Stopieñ nasilenia tego rozrostu by³
osobniczo zró¿nicowany od licznych ma³ych torbie-
li zlokalizowanych pod powierzchni¹ jajnika do du-
¿ych torbieli nab³onkowych (ryc. 10).
Grupa B. Struktura jajników tej grupy suk chorych
w porównaniu do suk grupy A znamiennie ró¿ni³a siê
znacznie mniejsz¹ liczebnoci¹ wtórnych komórek
zrêbu (tab. 2). Tak¿e nie u wszystkich suk stwierdza-
no obecnoæ torbieli pêcherzykowych i cia³ek ¿ó³tych.
Ekspresja badanych receptorów zachowywa³a siê po-
dobnie w wtórnych komórkach zrêbu jak to opisano
w jajnikach suk grupy A.
Grupa C. W jajnikach suk zdrowych stwierdzano
obecnoæ prawid³owo zbudowanych pêcherzyków jaj-
nikowych i cia³ka ¿ó³te. W zrêbie jajnika nie wyka-
zano obecnoci du¿ych skupisk wtórnych komórek
zrêbu.
W ostatnim okresie ukaza³a siê publikacja dotycz¹-
ca patogenezy chorób macicy u suk (1), w której auto-
rzy przedstawili opiniê, i¿ rozrost torbielowaty gru-
czo³ów endometrium macicy i ropomacicze mog¹ sta-
nowiæ dwie odrêbne jednostki chorobowe o wspólnej
etiopatogenezie, i bêd¹ce na ró¿nym etapie rozwoju
procesu chorobowego. Opiniê tê autorzy poparli wy-
nikami badañ klinicznych i patomorfologicznych
macicy suk. Równoczenie wykazali oni, ¿e mo¿na
czêsto pope³niaæ b³êdy w rozpoznawaniu choroby,
jeli podstaw¹ diagnozy jest wy³¹cznie kliniczna oce-
na stanu narz¹du rozrodczego suki. W cytowanej pra-
cy autorzy w bardzo uporz¹dkowany i nowoczesny
sposób przedstawili obraz zmian mikroskopowych
w endometrium chorych suk od zaniku ciany macicy
do rozwoju typowego zespo³u rozrostu torbielowate-
go/ropomacicza. Brak jednak w tym opisie dok³adnych
danych dotycz¹cych typu zapalenia i momentu poja-
wiania siê nacieku zapalnego w endometrium i jamie
macicy. Tak¿e nie uwzglêdniono zachowania siê ko-
mórek nab³onków gruczo³owego i powierzchownego,
które, podobnie jak ma to miejsce w cyklu, tak i w
stanach patologicznych ulegaj¹ zmianom odzwiercied-
laj¹cym fazê dzia³ania hormonów p³ciowych. Wiado-
mo bowiem, ¿e zmiany w wygl¹dzie komórek nab³on-
ka endometrium pozostaj¹ w cis³ym zwi¹zku ze zmia-
n¹ ich funkcji i charakterem stymulacji hormonalnej.
Ilustracj¹ tych zmian w budowie en-
dometrium s¹ od dawna znane zmia-
ny zale¿ne od fazy cyklu.
Wyniki niniejszych badañ jedno-
znacznie wskazuj¹, ¿e w jajnikach
suk z klinicznie rozpoznawanymi
chorobami macicy, niezale¿nie od
stopnia nasilenia choroby, pojawia
siê w zrêbie populacja komórek, któ-
rych struktura ró¿ni siê od struktury
prawid³owych komórek zrêbu jajni-
ka. Komórki te za Ericssonem (2)
okrelono mianem wtórnych komó-
rek zrêbu. Autor ten opisa³ wtórne
komórki zrêbu w jajnikach kobiet, wyra¿aj¹c pogl¹d,
¿e pochodz¹ one z transformowanych komórek zrêbu
jajnika. Na podstawie uzyskanych w niniejszej pracy
wyników badañ mo¿na przypuszczaæ, i¿ komórki wtór-
ne zrêbu mog¹ pochodziæ tak¿e z innych struktur jaj-
nika, takich jak cia³ko ¿ó³te lub komórki pêcherzykowe.
Cech¹ charakterystyczn¹ budowy wtórnych komó-
rek zrêbu jest obecnoæ du¿ej zawartoci lipidów
w cytoplazmie, co mo¿e stanowiæ wyraz spichrzenia
lipochromów, jak i sugeruje wystêpowanie zaburzeñ
w metabolizmie lipidów w komórkach, które posiada-
j¹ zdolnoæ do steroidogenezy. Cytoplazmatyczne
spichrzenie lipochromów nadawa³o tym komórkom
barwê intensywnie ¿ó³t¹, wyró¿niaj¹c¹ je w prepara-
tach barwionych hematoksylin¹ i eozyn¹. W skraw-
kach jajników, w których wykonano reakcjê histoche-
miczn¹ PAS/AB, cytoplazma tych komórek w miarê
nasilania siê procesu spichrzenia barwi³a siê intensyw-
nie w reakcji PAS/AB. Obecnoæ analogicznych zmian
w strukturze jajników po raz pierwszy opisano u 14-
-miesiêcznych myszy (3), które powsta³y w wyniku
zaburzeñ hormonalnych spowodowanych podaniem
oseskom mysim diethylstilbestrolu w pierwszych
3 dniach ¿ycia. Komórki te, podobnie jak stwierdzono
to w jajnikach chorych suk, uk³ada³y siê w pola lub
sznury, a ich cytoplazma by³a tak¿e ob³adowana lipi-
dami. U starszych myszy obserwowano ponadto pro-
liferacjê nab³onka jajnikowego, któr¹ to zmianê stwier-
dzano tak¿e w jajnikach chorych suk. Stopieñ prolife-
racji komórek nab³onka jajnikowego by³ u chorych suk
w znacznym stopniu zró¿nicowany, czego najlepszym
dowodem by³o powstawanie ma³ych lub bardzo du-
¿ych torbieli nab³onkowych. Ten du¿y stopieñ zró¿ni-
cowania osobniczego nie zezwala na uznanie tej zmia-
ny za cechê charakterystyczn¹ dla badanego zespo³u
chorobowego u suk, w przeciwieñstwie do obecnoci
w zrêbie jajników wtórnych komórek zrêbu. Stwier-
dzenie rozrostu nab³onka jajnikowego, zarówno u cho-
rych suk, jak i myszy z eksperymentalnie wywo³any-
mi zaburzeniami hormonalnymi, nasuwa pewne w¹t-
pliwoci odnonie do interpretacji wystêpowania w jaj-
nikach suk tzw. struktur podnab³onkowych, co stano-
wi powszechnie przyjête okrelenie dla ma³ych tor-
bieli nab³onkowych w jajniku suk (6), jako cechy cha-
Ryc. 10. Jajnik suki z grupy A. Torbiele
nab³onkowe. HE × 10
Ryc. 9. Jajnik suki z grupy A. Prolifera-
cja nab³onka pokrywaj¹cego jajnik. HE
× 20

Medycyna Wet. 2006, 62 (3)
315
rakterystycznej dla tego gatunku. Autorzy niniejszej
pracy nie podzielaj¹ tej opinii i uwa¿aj¹, i¿ prolifera-
cja nab³onka jajnikowego u suk stanowi odbicie wy-
stêpuj¹cych zaburzeñ hormonalnych.
Wyniki przeprowadzonych obserwacji jajników suk
z chorobami macicy oraz rezultaty badañ immunocy-
tochemicznych ekspresji receptorów PR, ER i alfa-
-inhibiny nasuwaj¹ przypuszczenie, ¿e komórki wtór-
ne zrêbu to transformowane w wyniku choroby ko-
mórki lutealne i komórki os³onki wewnêtrznej oraz
komórki pêcherzykowe. Dowodem na takie pochodze-
nie wtórnych komórek zrêbu mo¿e byæ obserwowana
w tych komórkach ekspresja receptorów j¹drowych
odpowiednio PR lub ER. Jakkolwiek obecna w cyto-
plazmie wtórnych komórek zrêbu ekspresja receptora
alfa-inhibiny by³a o bardzo s³abym nasileniu, to jed-
nak wskazuje na to, i¿ komórki te cechuje zdolnoæ
steroidogenezy. To pozwala wnioskowaæ, i¿ groma-
dzenie lipochromów w cytoplazmie tych komórek sta-
nowi wyraz zaburzenia procesu steroidogenezy. W wa-
runkach fizjologicznych lipidy s¹ zu¿ywane do synte-
zy odpowiednich hormonów jajnikowych.
Przedstawione wyniki s¹ rezultatem badañ przepro-
wadzonych na materiale klinicznym, a wiêc bardzo
zró¿nicowanym pod wzglêdem osobniczym. Fakt ten
z jednej strony stanowi³ o wyst¹pieniu pewnej trud-
noci w uchwyceniu dominuj¹cych i charakterystycz-
nych zmian patomorfologicznych w jajnikach suk,
u których choroby macicy by³y w ró¿nej fazie i stop-
niu zaawansowania, lecz z drugiej strony, zró¿nico-
wany obraz zmian chorobowych pozwoli³ na przele-
dzenie ich narastania oraz wykazanie obecnoci kore-
lacji miêdzy stopniem nasilenia zmian patologicznych
w macicy a liczebnoci¹ komórek wtórnych zrêbu.
Wspó³zale¿noæ liczby wtórnych komórek zrêbu
obecnych w jajnikach suk narastaj¹ca wraz z rozwo-
jem zmian chorobowymi w macicy dowodzi, i¿ oba
typy zmian s¹ w bli¿ej nieznany sposób zwi¹zane
z patogenez¹ rozrostu torbielowatego b³ony luzowej
i ropomacicza. Jakkolwiek wniosek ten ma charakter
hipotetyczny, to argumentem przemawiaj¹cym za rol¹
wtórnych komórek zrêbu jajnika w patogenezie zmian
chorobowych obserwowanych w endometrium jest
fakt, i¿ podobnie jak to opisano w jajnikach myszy
(3, 5) pozostaj¹ one w zrêbie w postaci nieprawid³o-
wych, patologicznych komórek zrêbu, jakkolwiek win-
ny ulec apoptozie. Obecnoæ receptorów PR i ER mo¿e
ponadto sugerowaæ, i¿ komórki te zachowa³y zdolnoæ
syntezy i sekrecji hormonów jajnikowych. Równoczes-
na obecnoæ ladowej ekspresji alfa-inhibiny hor-
monu pe³ni¹cego rolê autokrynowego regulatora ste-
roidogenezy mo¿e wskazywaæ, i¿ komórki wtórne
zrêbu jajnika wymknê³y siê z fizjologicznych pro-
cesów regulacji steroidogenezy.
Niektórzy autorzy (4) opisuj¹ grupy komórek zrêbu
o podobnych cechach morfologicznych jako cia³ko
¿ó³te resztkowe. Nie mo¿na zgodziæ siê z tak¹ inter-
pretacj¹, bowiem w procesie luteolizy komórki luteal-
ne ulegaj¹ procesowi fizjologicznej apoptozy, które-
mu to procesowi nie towarzyszy proces spichrzenia
cytoplazmatycznych lipochromów. Zachowanie siê
komórek wtórnych zrêbu mo¿e wskazywaæ na utratê
wra¿liwoci na sygna³y prowadz¹ce do indukcji apop-
tozy lub te¿ na obecnoæ zmian, które nast¹pi³y w pro-
cesach metabolicznych uszkodzonej komórki i sta³y
siê przyczyn¹ uszkodzenia ³añcucha metabolicznego
apoptozy. To ostatnie zjawisko cechuje komórki, któ-
re ulegaj¹ metaplazji nowotworowej. Wystêpowanie
w jajniku grup komórek, które z patologicznego punktu
widzenia mo¿na okreliæ jako komórki stanowi¹ce
wyraz zaburzenia homeostazy komórkowej lub dys-
plastycznej, jak i obserwowana proliferacja nab³onka
jajnikowego mog¹ stanowiæ interesuj¹ce informacje
do wykorzystania w patologii porównawczej, a w
szczególnoci z problematyk¹ zwi¹zan¹ z badaniami
mechanizmów metaplazji nowotworowej w jajniku.
Nawi¹zuj¹c do opisanych u myszy zmian mikrosko-
powych w strukturze jajników po eksperymentalnym
podaniu diethylstilbestrolu podobnych do obserwowa-
nych u suk z chorobami macicy mo¿na wniosko-
waæ, ¿e pojawienie siê w zrêbie jajników chorych suk
licznych wtórnych komórek zrêbu stanowi podobnie
wyraz patologicznej stymulacji spowodowanej przez
estrogeny. Wiadomo tak¿e, ¿e w indukcji procesów
zwi¹zanych z rozrostem edometrium u suk, która za-
pocz¹tkowuje chorobê macicy rozwijaj¹c¹ siê z up³y-
wem czasu w zespó³ zwany rozrostem torbielowatym
b³ony luzowej macicy i ropomaciczem musi mieæ
miejsce patologiczna stymulacja wywo³ana przez es-
trogeny. A za tym obecnoæ wtórnych komórek zrêbu
w jajnikach chorych suk stanowi w jajniku wyraz za-
burzeñ w procesie follikulogenezy/oogenezy, jak i lu-
teolizy cia³ka ¿ó³tego.
Przedstawione zmiany w strukturze mikroskopowej
jajników chorych suk z zespo³em rozrostu torbielo-
watego endometrium i ropomaciczem stanowi¹ dowód
na to, i¿ byæ mo¿e przyczyn¹ wyst¹pienia choroby
macicy s¹ pierwotne, bli¿ej niesprecyzowane zaburze-
nia hormonalne o charakterze endokrynowym lub pa-
rakrynowym, którym towarzyszy pojawienie siê w zrê-
bie jajników nietypowych komórek, okrelonych mia-
nem wtórnych komórek zrêbu.
Pimiennictwo
1.De Boschere H. De., Ducatelle R., Vermeirsch H., Van Den Broeck W.,
Coryn M.: Cystic endometrial hyperplasia pyometra complex in the bitch:
should the two entities be disconnected. Theriogenology 2001, 55, 1509-1519.
2.Erickson F. G., Magfoffin D. A., Dyer A. C., Hofeditz C.: The ovarian androgen
producing cells: a review of structure/function relationships. Endocr. Rev. 1985,
6, 371-399.
3.Mori T.: Age related changes in ovarian responsiveness to gonadotropins
in normal and neonatally estrogenized mice. J. exp. Zool. 1979, 207, 451-458.
4.Schoon H. A., Schoon D., Nolte I.: Untersuchungen zur Pathogenese des En-
dometritis-Pyometra-Komplexes der Hundin. J. Vet. Med. A 1992, 39, 43-56.
5.Tenenbaum A., Forsberg J. G.: Structural and functional changes in ovaries
from adult mice treated with diethystilbestrol in the neonatal period. J. Reprod.
Fert. 1985, 73, 465-477.
6.Jones T. C., Hunt R. D., King N. W.: Veterinary Pathology, Genital System,
Lippincott Williams&Wilkins, Baltimore 1997, s. 1158.
Adres autora: prof. dr hab. Maria Katkiewicz, ul. Ebro 45, 01-490 War-
szawa
Wyszukiwarka
Podobne podstrony:
Zmiany struktur demograficznych i społecznych w Polsce w Europie i na świecie
59 Zmiany strukturalne w dwuwarstwie po zwiazaniu z bialkami lub peptydami
Handel zagraniczny - bilans płatniczy (27 stron) , Tendencje rozwoju i zmiany struktury polskiego ha
ZMIANY W STRUKTURZE SPOŁECZNEJ I MODELU ŻYCIA JEDNOSTKI OD SPOŁECZNOŚCI OPARTYCH NA ŁOWIECTWIE
4 Zmiany w strukturze etnicznej ziem polskich w czasie i po II wojnie światowej Kopiax
Zmiany struktury i własności powierzchni materiałów
Zmiany struktury w 322 osa
Zmiany łagodne jajnika (2)
Zmiany struktury skóry towarzyszące procesowi starzenia
Derma-zmiany skórne 1, Podstawowe wykwity chorobowe.
VII. GOSPODARKA A ZMIANY STRUKTURY SPOECZNEJ, GOSPODARKA A ZMIANY STRUKTURY SPOŁECZNEJ
Zmiany struktur demograficznych i społecznych w Polsce w Europie i na świecie
Starzenie się – zmiany zachodzące w ludzkim organizmie Choroby wieku podeszłego 2
Zmiany struktury tkanki roślinnej wywołane odwadnianiem osmotycznym
Barwiński, Marek Wpływ granic oraz uwarunkowań politycznych na zmiany struktury narodowościowej i w
Zmiany strukturalne DNA
więcej podobnych podstron