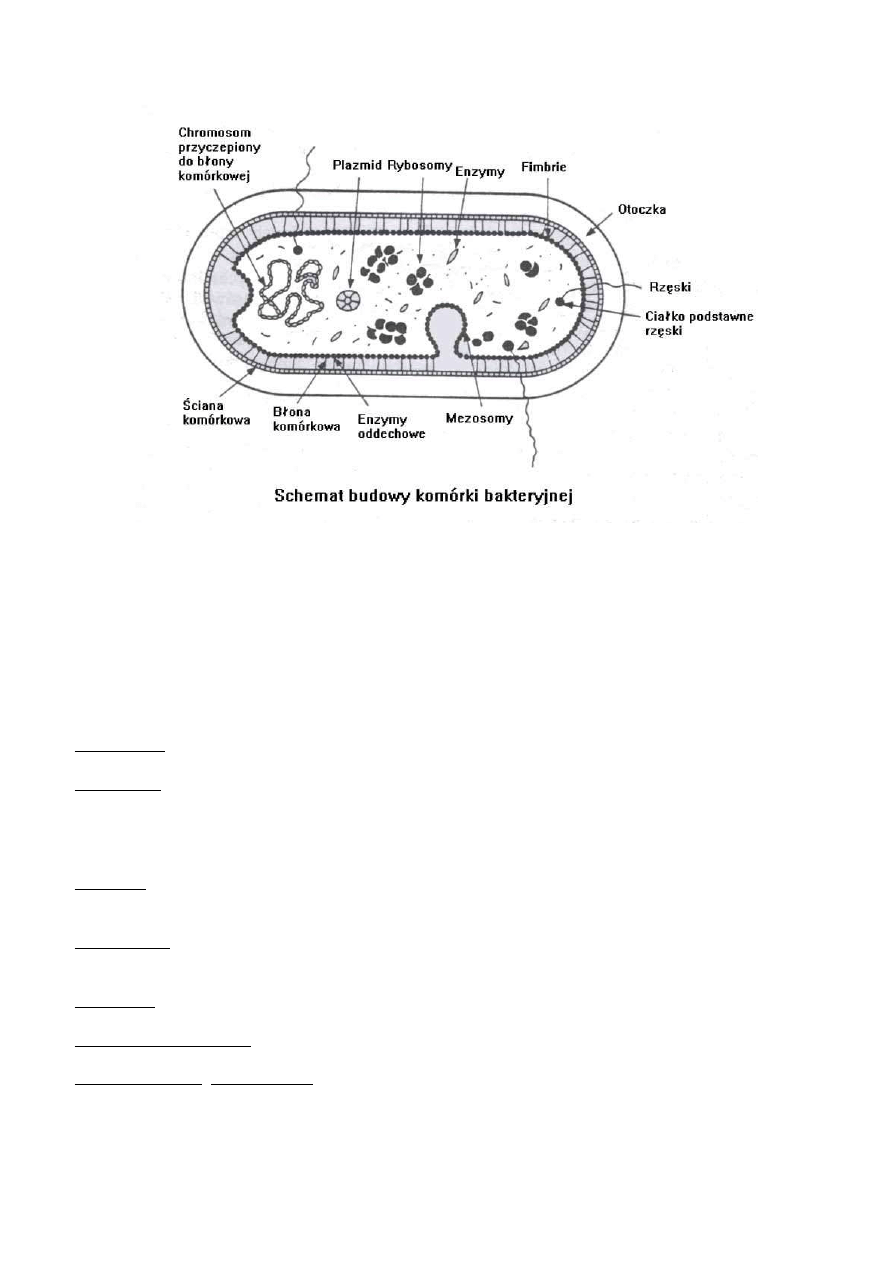
Budowa komórki bakteryjnej
cytoplazma
nukleoid (DNA)
rybosomy
błona cytoplazmatyczna (błona wewnętrzna)
ściana komórkowa
dodatki powierzchniowe: otoczka, rzęski, fimbrie, przetrwalniki (endospory), wtręty
cytoplazmatyczne (ziarnistości), inne plazmidy, transpozony
Cytoplazma - (protoplazma) - skład : woda, nieorganiczne jony, metabolity niskocząsteczkowe,
wysokocząsteczkowe polimery kwasów nukleinowych, białka, rybosomy, substancje zapasowe.
Nukleoid - w komórkach bakterii brak jądra komórkowego i jąderek. W genomie (materiał
genetyczny) w postaci DNA nie występują chromosomy, nie osłania go błona. Nukleoid (DNA)
nazywamy chromosomem bakteryjnym inaczejmateriałem chromosomowym. Jest w nim zawarta
informacja genetyczna dotycząca funkcji życiowych komórki (chromosomalne czynniki
dziedziczenia).
Plazmidy - koliste cząsteczki DNA (ulegają w cytoplazmie autonomicznej replikacji) determinują
cechy zwiększające możliwość przeżycia komórek bakteryjnych w określonych warunkach
środowiska.
Transpozony - fragmenty DNA podatne na przemieszczenia (transeokację) z jednego locus w inne. W
stanie nie zmienionym zostaje wbudowany w nowe miejsce w DNA. Transpozony zawierać mogą
różne geny także determinujące oporność na chemioterapeutyki.
Rybosomy - w komórce może być ponad 10.000 rybosomów. Mogą łączyć się ze sobą - tworzyć
agregaty (polirybosomy) w rybosomach zachodzi synteza białek.
Błona cytoplazmatyczna - otacza cytoplazmę wszystkich bakterii, przez błonę zachodzi komunikacja
komórek ze środowiskiem zewnętrznym.
Funkcje błony cytoplazmatycznej:
transport elektronów
synteza i transport prekursorów
wydzielanie zewnątrzkomórkowych i periplazmatycznych enzymów i toksyn.
regulacja segregacji chromosomalnego i plazmidowego DNA
wytwarzanie układów transportowych
przenoszenie receptorów i innych białek do układu chemotaktycznego
Budowa błony: 70% białka, 30% fosfolipidy, niewielka ilość węglowodanów
Mezosomy - to wpuklenia błony cytoplazmatycznej do wnętrza komórki.
Funkcje : mezosomy ścienne - miejsce przyczepienia nukleoidu do błony cytoplazmatycznej.
Znajdują się w nich enzymy potrzebne do syntezy składników ściany komórkowej.
znajdują się tu enzymy oddechowe (cytochromy)
zachodzą tu procesy oksydoredukcyjne
Ściana komórkowa - występuje tu peptydoglikan (mureina); kwasy tejchojowe
peptydoglikan ma kwas muraminowy, u niektórych bakterii jest zamiast niego kwas
talosaminouronowy, wtedy peptydoglikan nazywa się pseudomureiną.
Peptydoglikanu brak u : Tenericutes (mykoplazmy), Mendosicutes, Halobacterium (sololubne).
Cząsteczka peptydoglikanu zbudowana jest z długich łańcuchów polisacharydowych połączonych
w sieci poprzez mostki peptydowe.
Ze względu na różnice w budowie ściany komórkowej (ma to związek z barwieniem
metodzie Grama), bakterie podzielone są na : Gram-dodatnie i Gram-ujemne.
U bakterii Gram-dodatnich - grubość ściany - (20 - 80 nm)
skład ściany - (60 - 100% z peptydoglikanu tworzącego trójwymiarową sieć).
U wszystkich Gram-dodatnich występują polimery N-acetyloglukozoaminy i kwasu N-
acetylomuraminowego, mogą różnić się długością mostków peptydowych i składem aminokwasowym
Peptydoglikan - u bakterii Gram-dodatnich może być hydrolizowany przez lizozym (obecny w
płynach ustrojowych: łzy, ślina) także jest w białku jaja kurzego. Hydrolizuje on (lizozym)
glikozydowe wiązania między kwasem N-acetylomuraminowym, a N-acetyloglikozo-aminą.
Zachodzi wtedy liza komórki bakteryjnej. Mureina tworzy siatkę trójwymiarową.
Kwasy tejchojowe - to polimery w długiej cząsteczce. W jej skład wchodzi rybitol tzn. alkohol
z 3-węglowym cukrem. Polimery są połączone ze sobą fosfodwuestrowymi wiązaniami.
Kwasy tejchojowe (2 postacie): ribitolowy, glicerolowy
Kwas tejchojowy - rybitolowy jest zwykle związany z grupą 6-hydroksylową kwasu-N-
acetylomuraminowego (nazywany jest inaczej kwasem tejchojowym ściany komórkowej)
Kwas tejchojowy - glicerolowy - zawsze połączony z glikolipidami błony (nazywamy inaczej
lipotejchojowy).
Rola kwasów tejchojowych nie została dokładnie poznana. Przypuszczalnie biorą udział w
regulacji przechodzenia jonów przez warstwę peptydoglikanu (maja bowiem ładunek ujemny).
W ścianie mogą także być kwasy mykolowe (wolne kwasy tłuszczowe) np.
Corynebacterium,Mycobacterium. Różnica w ich strukturze (kw. mykolowych) jest kryterium do
identyfikacji i klasyfikacji.
W ścianie są także białka związane kowalencyjne lub niekowalencyjne nie tworzące struktury
błonowej.
Występują ponadto mutanty Bacillus subtilis i S. aureus nie mające kwasów tejchojowych, są one
zdolne do życia jednak występuje u nich zaburzenie procesu podziału komórek. Podobną
sytuację można obserwować w przypadku Streptococcus pneumoniae. U tego gatunku zamiast
choliny (innegoskładnika kwasów tejchojowych) występuje etanoloamina. W związku z tym w
podziale komórek nie tworzą się dwoinki ale długie łańcuchy.
Ściana bakterii Gram- ujemnych - ma bardziej złożoną budowę, skład : peptydoglikan (mureina)
błona zewnętrzna : fosfolipidy, białka, glikilipid (lipopolisacharyd) (LPS)
Warstwa mureiny jest cieńsza w porównaniu z Gram-dodatnimi drobnoustrojami (5 - 10% masy
ściany komórkowej) tworzy dwuwymiarową siatkę.
Jest bardziej elastyczna (jednak na tyle mocna aby utrzymać kształt komórki, jak również ochronić
ją przed osmatyczną lizą). W mureinie brak mostków wiążących krzyżowo, dlatego mostki
peptydowe łączące łańcuchy glikanu wiążą bezpośrednio grupę karboksylową D-alaniny jednego
łańcucha z wolną grupą aminową kwasu dwuaminopimelinowego łańcucha przyległego.
W odróżnieniu od gram_dodatnich utowaobnych jest 50% tetrapeptydów zakończonych wolną
D-alaniną.
Mureina z błoną cytoplazmatyczną jest połączona poprzez wiązania jonowe, natomiast z
błoną zewnętrzną przez lipoproteinę (lipoproteina mureinowa). Funkcja lipoproteiny - to utrzymanie
na miejscu innych białek błony zewnętrznej. Część białkowa kowalencyjnie jest związana z
mureiną, a lipidowa znajduje się w błonie zewnętrznej przyczepiona przy pomocy hydrofobowych
wiązań między lipoproteina, a fosfolipidami błony zewnętrznej.
Błona zewnętrzna - skład : fosfolipidy, lipopolisaeharyd (LPS), białka.
Fosfolipidy - dwie warstwy z których części hydrofobowe znajdują się naprzeciw siebie.
Błona zewnętrzna nie przepuszcza składników hydrofobowych jest ponadto oporna na działanie
detergentów dlatego też wiele bakterii Gram-ujemnych rośnie w ichTobecności. Zwiększenie
przepuszczalności błony powodują związki chelatujące np. EDTA (staje się przepuszczalna dla

antybiotyków i detergentów). Dla cząstek hydrofilowych błona zewnętrzna odznacza się duże
przepuszczalnością.
Białka główne - (70% ogółu białek w błonie zewnętrznej) powiązane są z LPS, ze sobą jak
i z lipoproteina mureinowa. W związku z powyższym tworzy się stabilna struktura.
Lipopolisacharydy (LPS - to długie heteropolimery zbudowane z trzech strukturalnie
odmiennych regionów które są powiązane ze sobą wiązaniami kowalencyjnymi. Są nimi : lipid
A (I), oligocukier rdzeniowy (II) i wielocukier o swoistości antygenowej 0, jest on antygenem
somatycznym (III).
Oligosacharyd rdzeniowy - podobny jest u wszystkich bakterii Gram-ujemnych w związku z
tym określony grupowo swoisty antygen (antygen wspólny) wykazany u Escherichia coli 014.
LPS bakterii Gram-ujemnych wykazuje właściwości toksyczne w stosunku do ssaków. Z uwagi
na znajdujący się w LPS toksyczny czynnik (wielocukier i/lub lipid A) będący integralną częścią
ściany komórkowej nazwano go endotoksyną inaczej toksyną lipopolisacharydową.
Antygen somatyczny - jest to najbardziej zewnętrzna część LPS będąca polimerem powtarzających
się jednostek oligocukrów utworzonych przez 2 lub 8 reszt jednocukrowych.
Lipid A - zbudowany jest z dwucukru : D-glikozaminylowego. Lipid A hamuje dyfuzję
związków hydrofobowych np. kwasów żółciowych, niektórych antybiotyków i detergentów przez
błonę zewnętrzną.
Endotoksyna - wywołuje wiele objawów chorobotwórczych m.inn.: wstrząs endotoksyczny
(wstrząs septyczny), miejscowe reakcje skórne, gorączka (pirogenność), leukocytoza, obniżenie
ciśnienia krwi indukcja nieswoistej odporności na zakażenie, wiele innych.
Znikome tj. nanogramowe ilości endotoksyny będą w wyjałowionych roztworach do wlewów
dożylnych mogą być powodem u pacjentów efektu pirogennego. Konieczne jest dlatego
badanie płynów infuzyjnych pod względem obecności endotoksyny.
Do zewnętrznej powierzchni ściany komórkowej mogą przylegać wydzielone przez bakterie
substancje o charakterze polimerów. W przypadku gdy substancje te tworzą grubą warstwę
otaczającą pojedynczą komórkę lub parę komórek nazywa się ją otoczką. Otoczki są zbudowane
z polisacharydów. Polisacharydy mają niskie powinowactwo do barwników i dlatego nie
widać ich w metodach stosowanych rutynowo. Można wykazać ich obecność w metodzie tzw.
negatywnej. Na ciemnym tle „halo" widać je niezabarwione. Można także wykazać ich obecność w
metodach serologicznych np. -odczyny : pęcznienia otoczek, aglutynacji, immunofluorescencyjnej, a
także aglutynacji lateksowej.
Funkcja otoczek - to ochrona komórki przed wysychaniem. Są z reguły dobrym antygenem,
indukują wytwarzanie swoistych przeciwciał biorących udział w niszczeniu bakterii. Bakterie
wytwarzające otoczki rosną na podłożach z zawartością np. cukru, surowicy, w atmosferze CO
2
w
postaci śluzowatych kolonii typu S, M. Ich odmiany bezotoczkowe tworzą kolonie R-szorstkie. Na
powierzchni komórki mogą znajdować się inne substancje, które nie są warstwą komórkową.
Nazywane są mikrootoczką, glikokaliksem, śluzem lub egzopolisacharydem.
Rzęski - narząd ruchu bakterii, są to długie, w środku puste, spiralnymi filamentami (nici). Ich
długość jest kilka razy większa niż długość komórki, a średnica w granicach 12-20 nm. Rozmiary te
są mniejsze od zdolności rozdzielczej, mikroskopu świetlnego dlatego nie mogą być widoczne w
rutynowym barwieniu preparatów, jedynie przy użyciu odpowiedniej bejcy, które doprowadza do
powiększania średnicy rzęsek. W mikroskopie świetlnym można obserwować ruch drobnoustrojów
w tzw. kropli wiszącej. W mikroskopie elektronowym są łatwo dostrzegalne. Wzrost urzęsionych
drobnoustrojów można obserwować na podłożach stałych w postaci „mgiełki - pełzanie", jak
również wzrost w podłożu z dodatkiem TTC (chlorek trifenylotetrazoliowy) - wzrost poza linią
wkłucia w całej objętości podłoża. Ich wytwarzanie jest cechą gatunkową zależną także od
warunków środowiska np. Listeria monocytogenes, Yersinia enterocolitica i Yersinia
pseudotuberculoris nie wytwarzają w temperaturze 37°C, a są obecne w temperaturze niższych (22°C
- 30°C). Ważną cechą taksonomiczną bakterii jest ich rozmieszczenie i liczba. Charakterystyczna
dla rodzaju jest ich lokalizacja. Umiejscowienie rzęsek : dookoła komórki (wkołorzęse = peritricha)
na jednym biegunie (czuorzęse = lophotricha) na obu biegunach po 1 rzęsce (dwurzęse = ditricha)
jedna rzęska na biegunie (jednorzęse = monotricha).
Rzęski wychodzą z cytoplazmy komórki. Rzęska przymocowana jest za pomocą ciałka podstawnego
(w postaci haczyka, pierścienia albo płytki). Różni się ono budową u bakterii Gram-ujemnych (2
pary pierścieni), i Gram-dodatnich (1 para pierścieni). Pierścienie te pełnia rolę tulejki, przez którą
przechodzą nici rzęski (filamenty). Rzęski zbudowane są z białka tzw. flageliny (białko rzęskowe),
która jest immunogenem (antygenem H).
Bakterie za pomocą rzęsek poruszają się dzięki ich ruchowi obrotowemu przypominającemu ruch
śruby okrętowej. Występuje u bakterii zjawisko chemotaksji tzn. poruszanie się w kierunku
substancji odżywczych lub uciekanie od substancji szkodliwych. Ruch komórki zależy od kierunku,

w którym obraca i rzęska np. jeżeli odwrotny do kierunku wskazówek zegara - komórka porusza się
po linii prostej, jeśli zgodnie to „koziołkuje" w miejscu. Jeśli płynie do substancji odżywczej ruchu
po linii prostej są dłuższe, fazy koziołkowania są krótsze, jeśli oddala się od substancji wabiącej i
zbliża się do szkodliwej wzmagają się fazy koziołkowania, aż do momentu ustalenia odpowiedniego
kierunku.
Fimbrie - (pile) - nitkowate dodatki jedynie widoczne w mikroskopie elektronowym. Są proste i
krótsze od rzęsek. Występują u bakterii Gram-ujemnych u Gram-dodatnich np. Streptococcus
spp., Corynetobacterium spp. Zbudowane są z białka składającego się z podjednostek zwanych
pillinami. Jako białko są swoistym immunogenem. Komórka może nie posiadać fimbrii jest wtedy
nieufimbriowana, jeśli wytwarza je jest ufimbriowana. Typy fimbri : zwykłe i płciowe.
Zwykłe - występują u bakterii Gram-ujemnych z rodziny Enterobacteriaceae, rodzaju Pseudomonas i
Haemophilus oraz u Neisseria gonorrhoae. Są wyznacznikiem chorobotwórczości bakterii. Ich
wytwarzanie uwarunkowane jest najczęściej przez geny w chromosomie, rzadko przez plazmidowe.
Płciowe - ich wytwarzanie zależne od genów w plazmidach (F, R, col). Fimbrie te maja kanał,
przez który może przechodzić transpozonowy chromosomalny, plazmidowy lub bakteriofagowy
DNA.
Przetrwalniki - (endospory) - wytwarzane przez rodzaj Bacillus spp. i Clostridium spp. np. B.
subtilis, B. anthracis, C. botulinum, C. perfingens, S. tetanii. Rola przetrwalników - przetrwanie w
niekorzystnych warunkach życia (brak składników odżywczych, wody). Dzięki nim mogą bakterie te
przeżywać setki lat (anabioza). Proces tworzenia przetrwalnika - proces sporulacji w warunkach
doświadczalnych początek jego następuje po skończeniu logarytmicznej fazy wzrostu, na początku
fazy stacjonarnej wtedy kiedy brakuje źródeł węgla i azotu, a gromadzą się metabolity. Proces
(sporulacja), przebiega w wielu etapach. Następuje zagęszczenie cytoplazmy wokół jednego
nukleoidu, tworzy się błona cytoplazmatyczna i warstwa peptydoglikanu tzn. osłon które otaczają
tworzący się przetrwalnik. W nim zatrzymany zostaje proces metaboliczny tzw. „pauza
metaboliczna". Przetrwalnik jest oporny na działanie szkodliwych dla niego czynników
zewnętrznych na brak wody, podwyższona temperatura, promienie UV, brak składników odżywczych,
zmiany pH. Z uwagi na obecność kwasu dipikolinowego nie niszczy go po 1 godzinne gotowanie w
temp. 100°C. Proces kiełkowania tzw. przejście w formę komórki wegetatywnej nazywany jest
germinacją. Leki infuzyjne, produkty spożywcze np. konserwy nieprawidłowo wyjałowione (tz.
w temperaturze mniejszej 121°C) mogą zawierać przetrwalniki. Po wykiełkowaniu w konserwie
powodują one wystąpienie nie tzw. bombażu, a lek nie może być podawany pacjentowi. Lek można
jałowic także w procesie filtracji. Wymiary ednospor mogą być mniejsze od poprzecznego wymiaru
komórki macierzystej lub większe np. Clostridium spp. Umiejscowienie przetrwalników w
komórce jest: centralne, subcentralne lub biegunowe, a komórki mają kształt buławy lub wrzeciona.
Cechą taksonomiczną jest umiejscowienie przetrwalnika a także jego wielkość. Od chorego mogą
być izolowane między innymi z ropy, plwociny, kału, rany. Przetrwalniki są widoczne w
mikroskopie świetlnym po wybarwieniu metodą np. Schaeffer-Fultona. Wybarwiają się na kolor
podgrzanego barwnika.
Ziarna cytoplazmy - (ciałka wtrętowe) - mają funkcje zapasowe są źródłem energii, mogą być
otoczone cienką błoną. Ziarnistości mogą być zbudowane z polimeru kwasu p-hydroksymasłowego
albo zawierają nieorganiczne fosforany. Ziarnistości wolutynowe występują u Corynebacterium
diphtheriae (jakc ciałka Emsta-Babesa). Oglądane są w mikroskopie świetlnym po wybarwieniu
metodą Neissera. Ziarnistości wybarwione są na kolor ciemnobrązowy,' komórka
wegetatywna na kolor żółty. Corynebacterium spp. - pałeczki Gram-dodatnie w preparacie
układają się w postaci liter chińskich, I,Y,X,L Występują w wodzie, glebie, na skórze i błonach
śluzowych. Corynebacterium diphtheriae izolowanych jest z gardła, pępka, rany.
Prątki - Mycobacterium spp. - należy 26 gatunków określonych jako wolnorosnące i 28 jako szybko
rosnące. Są bakteriami wydłużonymi, pręcikowatymi, prostymi lekko zagiętymi. Są Gram-dodatnie,
kwasooporne (w ścianie komórkowej są duże ilości lipidów tzw. wosków). Rosną na podłożach w
warunkach tlenowych, tworzą bezbarwne lub barwne kolonie. Niektóre są saprofityczne (Myc.
smegatis) jest w ziemi w wodzie, w wydzielinach łojotokowych skóry, w moczu, w mastce. Za
postać gruźlicy płucnej odpowiedzialny jest Myc. tuberculosis (najczęściej u człowieka), Myc bovis
i Myc bovis BCG (gruźlica odzwierzęca u ludzi). Myc africanum (gruźlica płucna w Afryce), Myc
microti występuje u zwierząt.
Podział prątków niegruźliczych - Fotochromogenne (gr I) powolny wzrost od 2-10 tygodni, barwnik
stymulowany przez światło (M. kansasii, M. simiae). - Skotochromogenne (gr II) powoduje wzrost,
barwnik w obecności lub bez obecności światła (M. scrofulaceum, M. xenopi). - Niefotochronogenne
(gr III) - powolny wzrost (M. avium, M. intracellulare). - Szybkorosnące (gr IV) - wzrost w ciągu 7
dni (M. fortuitum, M. smegmatis). Materiał do badań - plwocina, popłuczyny żołądkowe, płyny
wysiękowe, płyn mózgowo-rdzeniowy,wymaz krtani, mocz. Metoda barwienia - met. Ziehl -
Neelsena (prątki zabarwione czerwono, elementy morfotyczne na niebieskie)
Antybiotyki p-laktamowe hamują tworzenie peptydowych, krzyżowych mostków w mureinie u
bakterii Gram-dodatnich. Podobne właściwości (hamowanie syntezy mureiny) wykazują :
wankomycyna,
fosfomycyna, bacytracyna, ykloseryna. Także działanie liozymu powoduje rozerwanie połączeń
glukozoamina, a kwas acetylomuraminowy. Komórki takie - bez mureiny u bakterii Gram-
dodatnie
nazywano protoplastami.
Sferoplasty - wytwarzane podobnie komórki bez mureiny u bakterii Gram-ujemnych.
Protoplasty i steroplasty są: wrażliwe na działanie ciśnienia osmotycznego środowiska.
In vitro żyją w środowisku o wyższym ciśnieniu osmotycznym np. sole magnezu, albumina
bydlęca,
żelatyna, 20% sacharoza.
Przeniesione do podłoża, które nie zawiera czynnika hamującego syntezę mureiny zmieniają
swój
kształt do wyjściowego (normalnych komórek).
Postacie -Ł-bakterii są to sferoplasty i protoplasty hodowane in vitro i in vivo. Na podłożach
stałych tworzą kolonie wrastające w podłoża o kształcie zbliżonym do „sadzonego jaja"
Badania mikroskopowe - Elementy komórki obserwowane są przy użyciu mikroskopów:
kontrastowo-fazowy, fluorescencyjny, ultrafioletowy, świetlny zwykły, ultramikroskop, elektronowy,
skaningowy
Mikroskop fazowo-kontrastowy - oglądanie drobnoustrojów umieszczonych w kropli wody, jak
również odróżnienie bez barwienia struktur komórkowych. Wykazywana jest także obecność
drobnoustrojów w niebarwionych fragmentach tkanek. Optyka w tym mikroskopie uwzględnia
różnice w gęstości materiałów biologicznych i ich współczynników załamania światła.
Mikroskop luminescencyjny ( fluoroscencyjny) - Jako oświetlenie preparatu zastosowane są
promienie
UV będące niewidzialne dla ludzkiego oka. Otrzymane zjawisko świecenia preparatu pod wpływem
tego
promieniowania nazwano fluoresćencją.
Mikroskop świetlny - zwykły - zasadą działania tego mikroskopu jest wzmocnienie dwóch
soczewek tworzących zespół. Jedna z nich jest obiektywem, druga to okular. Otrzymane
powiększenie jest iloczynem powiększeń : obiektywu i okularu. Obecnie najsilniejsze
stosowanie soczewki będące obiektywami powiększają do 100x, natomiast okulary do 25x.
Zastosowanie tego mikroskopu : w laboratoriach mikrobiologicznych i cytologicznych oglądane są w
nim najczęściej drobnoustroje zabite i barwione.
W mikroskopie świetlnym obserwowane są: wielkość, kształt, ułożenie, sposób barwienia. Ponadto
przy
zastosowaniu specjalnych technik barwienia, obecność rzęsek, otoczek, przetrwalników, ziaren
wolutynowych.
Mikroskop ultrafioletowy - tu także wykorzystano promienie UV. Przechodzi ono przez szkło
kwarcowe będące częścią składową systemu optycznego. Zwiększona jest tu zdolność rozdzielcza
mikroskopu do
0,1 mikrona. Oglądane obrazy rejestrowane są na kliszy fotograficznej.
Mikroskop elektronowy - wykorzystywany jest tu strumień elektronów przechodzących przez
pole magnetyczne (elektromagnesy) Obrazy mogą być oglądane na ekranie fluoryzującym lub
rejestrowane na kliszy fotograficznej. Obraz oglądany jest niby cień obserwowanego przedmiotu.
Mikroskop ten umożliwia powiększenie obrazu do 1 miliona razy. Najczęściej stosowane
powiększenia to od 30.000 do 50.000 razy. Oglądane są wysuszone , niezabarwione preparaty
poddawane zabiegom nazwanym cieniowania tj. impregnacja solami metali ciężkich np. solami złota.
Mikroskop elektronowy - (skaningowy) w tym przypadku strumień elektronów nie przenika
przez preparat. Oglądana jest przestrzenna budowa drobnoustrojów.
Ultramikroskop - obserwacja preparatów w ciemnym polu widzenia. Jest on zmodyfikowaną
postacią
mikroskopu świetlnego zaopatrzoną w kondensor. Zadaniem kondensora jest zatrzymanie centralnych
promieni świetlnych padających na preparat, który oświetlany jest promieniami bocznymi
padającymi pod pewnym kątem. Obserwowane są komórki na ciemnym tle. Mogą być oglądane
mniejsze cząsteczki aniżeli w mikroskopie świetlnym. Oglądane preparaty są świeże i niebarwione.
Techniki przygotowania preparatów - z pobranych próbek wykonuje się rozmazy, preparaty świeże,

preparaty utrwalone, barwione, niebarwione.Z materiałów płynnych (krew, mocz, płyn
mózgowordzeniowy) można wykonać preparat w postaci tzw. kropki wiszącej (głównie do oglądania
w ciemnym polu widzenia).
Wyszukiwarka
Podobne podstrony:
Filtry akwarium budowa id 1710 Nieznany
ProgSieciowe 15 Budowa id 39746 Nieznany
Jak dawniej budowano id 223573 Nieznany
budowa i dzialanie FDD id 94136 Nieznany (2)
4 Budowa ciala stalego id 3714 Nieznany
Budowa osrodka sportowo1 id 943 Nieznany
Budowa materii id 94290 Nieznany (2)
budowa wyrazow i zdan id 94443 Nieznany (2)
BCh Budowa zwiazkow id 81937 Nieznany (2)
BUDOWA DREWNA id 94109 Nieznany (2)
Budowa Drog1 id 94112 Nieznany
Budowanie systemu 11 id 94500 Nieznany (2)
budowa malej sieci id 94283 Nieznany (2)
Budowa krtani id 94261 Nieznany (2)
Budowa Lampy Elektronowej id 94 Nieznany (2)
Budowa i sklad atm id 94182 Nieznany (2)
Budowa bakterii id 94078 Nieznany (2)
budowa komputera id 94246 Nieznany
więcej podobnych podstron