
FIZJOLOGIA
CZ OWIEKA
Ł
KOLOKWIUM NR 4
Fizjologia trawienia
DietetykaUP, Fizjologia człowieka, kolokwium nr 4 – Fizjologia trawienia
1

Skład śliny
*woda 99%
*związki nieorganiczne: KCl, NaCl, NaHCO3, NaHPO4, Ca(HCO3)2, Na, K, Ca, Mg
*związki organiczne:
mucyna – glikoproteina zwilżająca pokarm, ochraniająca błonę śluzową jamy ustnej
alfa-amylaza ślinowa (ptialina) – hydrolizuje wiązania 1-4 alfa, rozkłada skrobię na
maltozę i dekstryny
lipaza ślinowa (językowa) – rozkłada triglicerydy do kwasów tłuszczowych i 1,2-
diacylogliceroli
laktoferryna – wiązanie żelaza I działanie bakteriostatyczne
lizozym – enzym bakteriobójczy, niszczy błonę komórkową bakterii
immunoglobulina IgA
białka bogate w prolinę – ochraniają szkliwo zębów i wiążą toksyczne taniny
cholesterolu
kwas moczowy
mocznik
Funkcje śliny
*nawodnienie pokarmu
*ułatwianie połykania kęsów pokarmowych dzięki otaczaniu ich śluzem
*trawienie skrobi przy pomocy amylazy
*funkcja antybakteryjna
*bierze udział w odczuwaniu smaku
Enzymy śliny
*alfa-amylaza ślinowa (ptialina) – hydrolizuje wiązania 1-4 alfa, rozkłada skrobię na
maltozę i dekstryny
*lipaza ślinowa (językowa) – rozkłada triglicerydy do kwasów tłuszczowych i 1,2-
diacylogliceroli
Czynniki wzrostu w ślinie
*aktywne biologicznie peptydy, które stymuluja proliferację i różnicowanie komórek;
Różnią się one biologicznie od typowych hormonów, gdyż zarówno ich miejsce
syntezy, jak i miejsce działania nie jest ograniczone do danej tkanki.
*przykłady:
naskórkowy czynnik wzrostu EGF (utrzymywanie stanu zdrowia głównie jamy
ustnej, oddziałuje na gojenie uszkorzeń skóry i błony śluzowej przewodu
pokarmowego)
nerwowy czynnik wzrostu NGF
transformujący czynnik wzrostu TGF
czynniki wzrostu rodziny fibroblastów FGH
czynniki wzrostu rodziny insuliny IGF-1, IGF-2
Czynniki odpowiedzialne za odpowiedź immunologiczną
*sekrecyjna immunoglobulina A, która wspomaga proces aglutynacji i zapobiega
DietetykaUP, Fizjologia człowieka, kolokwium nr 4 – Fizjologia trawienia
2
adhezji, kolonizacji bakterii na tkankach miękkich i zębach oraz działa
synergistycznie z elementami obrony nieswoistej
Aktywność i czynniki zmieniające aktywność amylazy ślinowej
*zachowuje aktywność wyłącznie w obecności jonów chlorkowych Cl-
*optymalne pH – 6.0, niskie pH hamuje aktywnośc amylazy ślinowej
*optymalna temperatura – 37 st
*pH ok 4.0 hamuje właściwości amylazy ślinowej
Trzy fazy wydzielania soku żołądkowego
DietetykaUP, Fizjologia człowieka, kolokwium nr 4 – Fizjologia trawienia
3
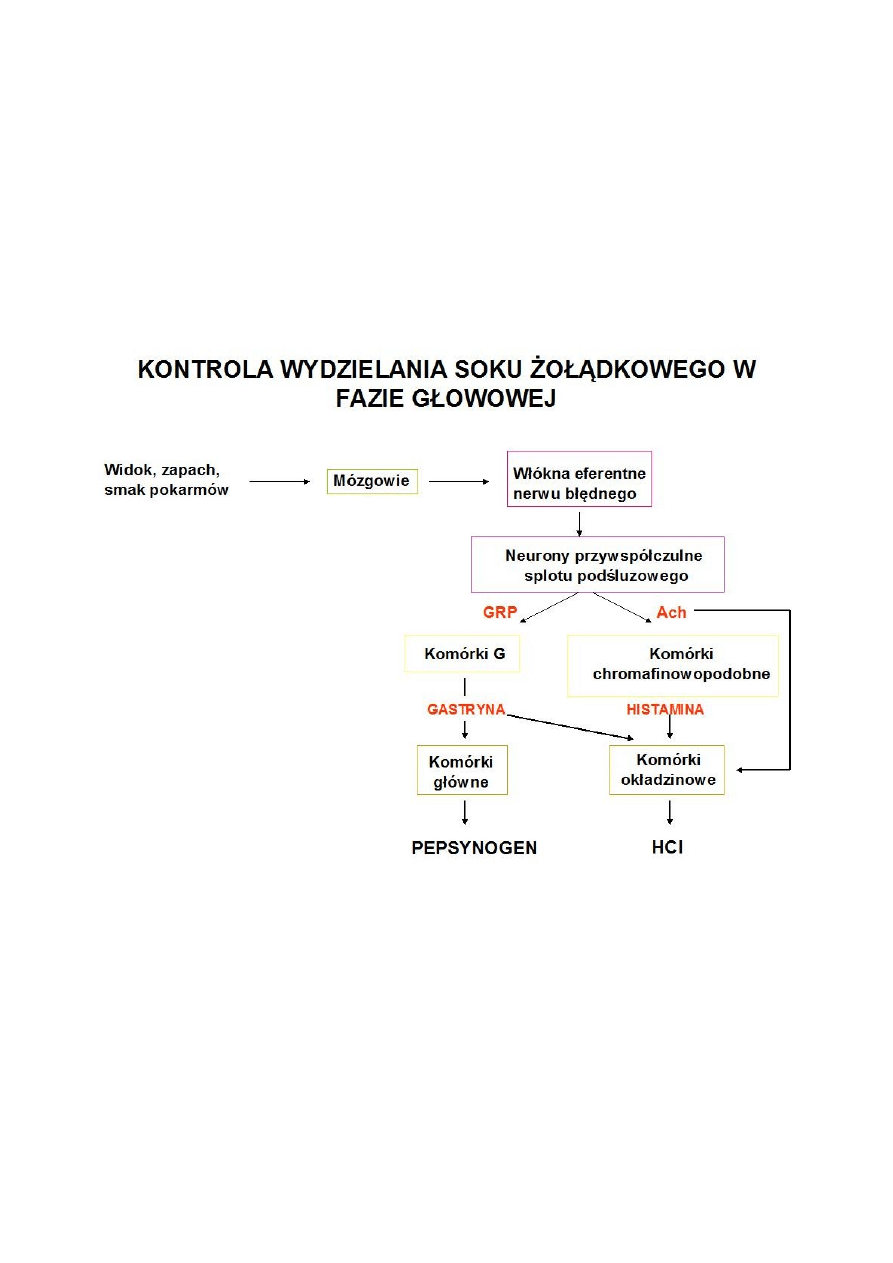
*Faza głowowa – jest związana z myśleniem o jedzeniu, widokiem, smakiem lub
zapachem. Zależna od unerwienia przywspółczulnego żołądka (n. błędny)
Eferentne włókna nerwu błędnego pobudzają wydzielanie HCL przez komórki
okładzinowe, gastryny przez komórki G oraz pepsynogenu przez kom. główne.
Prawie połowa soku żołądkowego jest wytwarzana w fazie głowowej i podczas
pobudzania n. błędnego
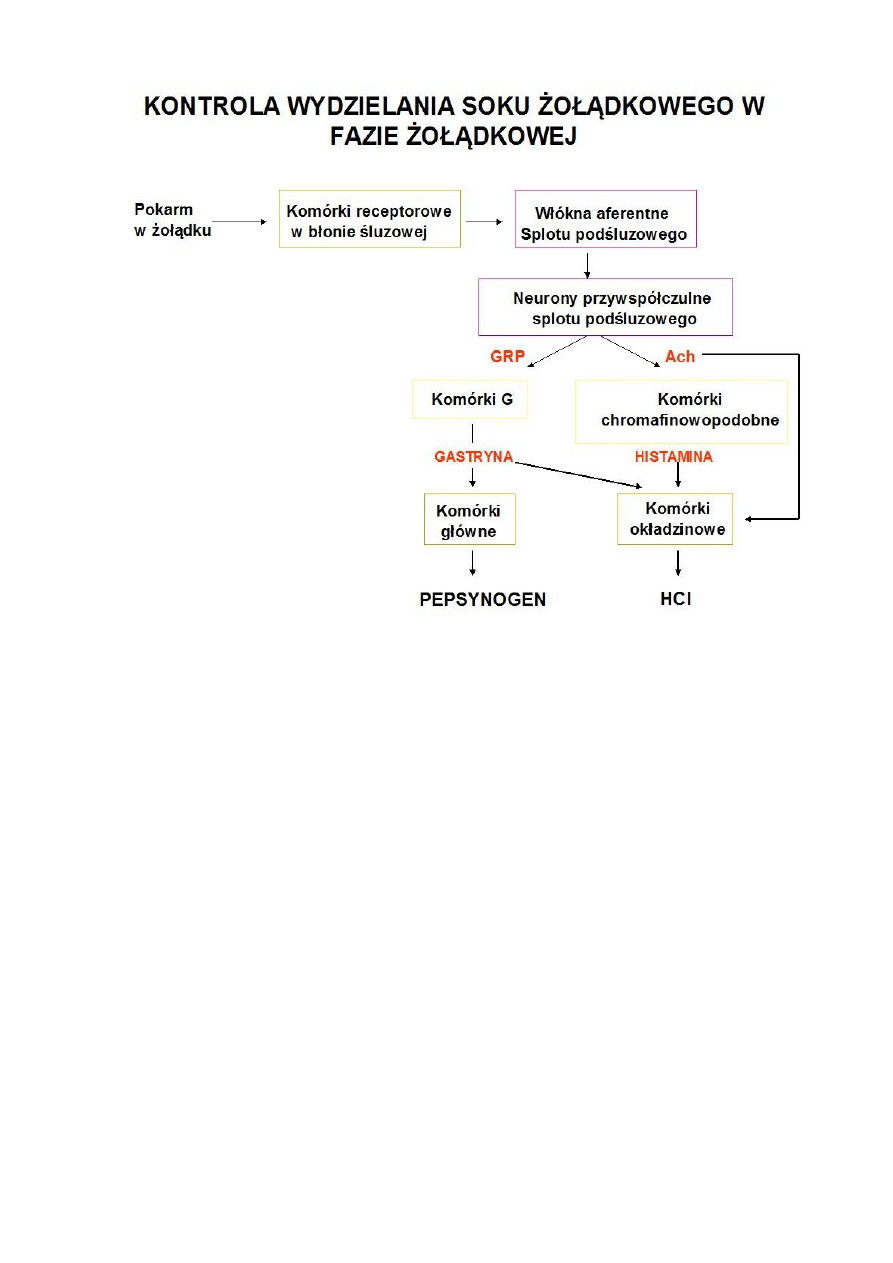
*Faza żołądkowa – rozpoczyna się w chwili dotarcia pokarmu do żołądka. Pokarm ten
buforuje częściowo kwas, podnosi pH i umożliwia działanie innych bodźców
wywołujących dalsze wydzielanie kwasu.
Rozciąganie trzonu żołądka prowadzi do wzrostu wydzielania HCL przez
pobudzenie odruchów miejscowych oraz wagonalnych.
Rozciągnięcie części odźwiernikowej pobudza wydzielanie gastryny za
pośrednictwem odruchów wagowagalnych miejscowych
Niskie pH wyzwala odruchy miejscowe co zwiększa wydzielanie pepsynogenu
Wydzielanie soku żołądkowego jest mniejsze w fazie żołądkowej, ale trwa dłużej.
*Faza jelitowa – zaczyna się w chwili opróżniania żołądka. Wydziela się w niej
niewiele soku żołądkowego; zachodzi tutaj zarówno pobudzanie jak i hamowanie
czynności żołądka; wytwarzana w dwunastnicy gastryna i CCK pobudzają
wydzielanie soku żołądkowego, natomiast sekretyna działa hamująco; za
pośrednictwem odruchu jelitowo-żołądkowego następuje hamowanie opróżniania
żołądka i wydzielania soku żołądkowego
DietetykaUP, Fizjologia człowieka, kolokwium nr 4 – Fizjologia trawienia
4

Skład soku żołądkowego
*kwas solny HCl – powstaje w kanalikach wewnątrz komórek okładzinowych
*pepsynogen – nieczynny enzym proteolityczny wytwarzany wkomórkach głównych
*śluz pokrywający ścianę żołądka
*kwas moczowy
*sód, chlor
*woda
Aktywność pepsyny i podpuszczki
*pepsyna – wytwarzana w komórkach głównych żołądka w postaci nieaktywnego
pepsynogenu;
aktywacja pepsynogenu do pepsyny polega na odłączeniu fragmentu białka w świetle
żołądka pod wpływem kontaktu z kwasem solnym;
pepsyna należy do endopeptydaz rozkładających wiązania peptydowe pomiędzy L-
aminokwasami aromatycznymi lub dwukarboksylowymi, a innymi aminokwasami;
największa aktywność pepsyny występuje w granicach pH od 1,6 – 3,2
*podpuszczka (chymozyna)
w obecności jonów wapnia ścina białka mleka (powoduje przechodzenie w formę
nierozpuszczalną) – kazeinę;
(przemienia rozpuszczalne białko mleka (kazeinogen) w nierozpuszczalny
parakazeinian wapnia (kazeinę))
Regulacja motoryki żołądka
*czynniki nerwowe:
nerw błędny X – przez przywspółczulne neurony śródścienne nasila skurcze i
przyspiesza perystaltykę żołądka
nerwy współczulne – działanie przeciwstawne – hamujące
*Włókna pobudzające o niższym progu pobudliwości działają na dalsze części żołądka,
natomiast hamujące, utworzone głównie przez włókna peptydergiczne,
purynergiczne i nitroergiczne wykazują wyższy próg pobudliwości i wywierają
wpływ hamujący głównie na motorykę początkowej części żołądka
*Na skurcze toniczne błony mięśniowej i perystaltykę żołądka mają wpływ czynniki
nerwowe i humoralne. Czynniki hamujące na drodze odruchowej i humoralnej
perystaltykę żołądka i zwalniające jego opróżnianie to:
;rozciągnięcie ścian dwunastnicy przez pokarm przechodzący z żołądka, czemu
towarzyszy zmniejszenie wartości pH w dwunastnicy
;zawartość kwasów tłuszczowych, częściowo strawionych białek i
węglowodanów w treści pokarmowej przechodzącej do dwunastnicy
;sekretyna i cholecystokinina powstające w dwunastnicy
;wysokie ciśnienie osmotyczne treści żołądkowej dostającej się do dwunastnicy
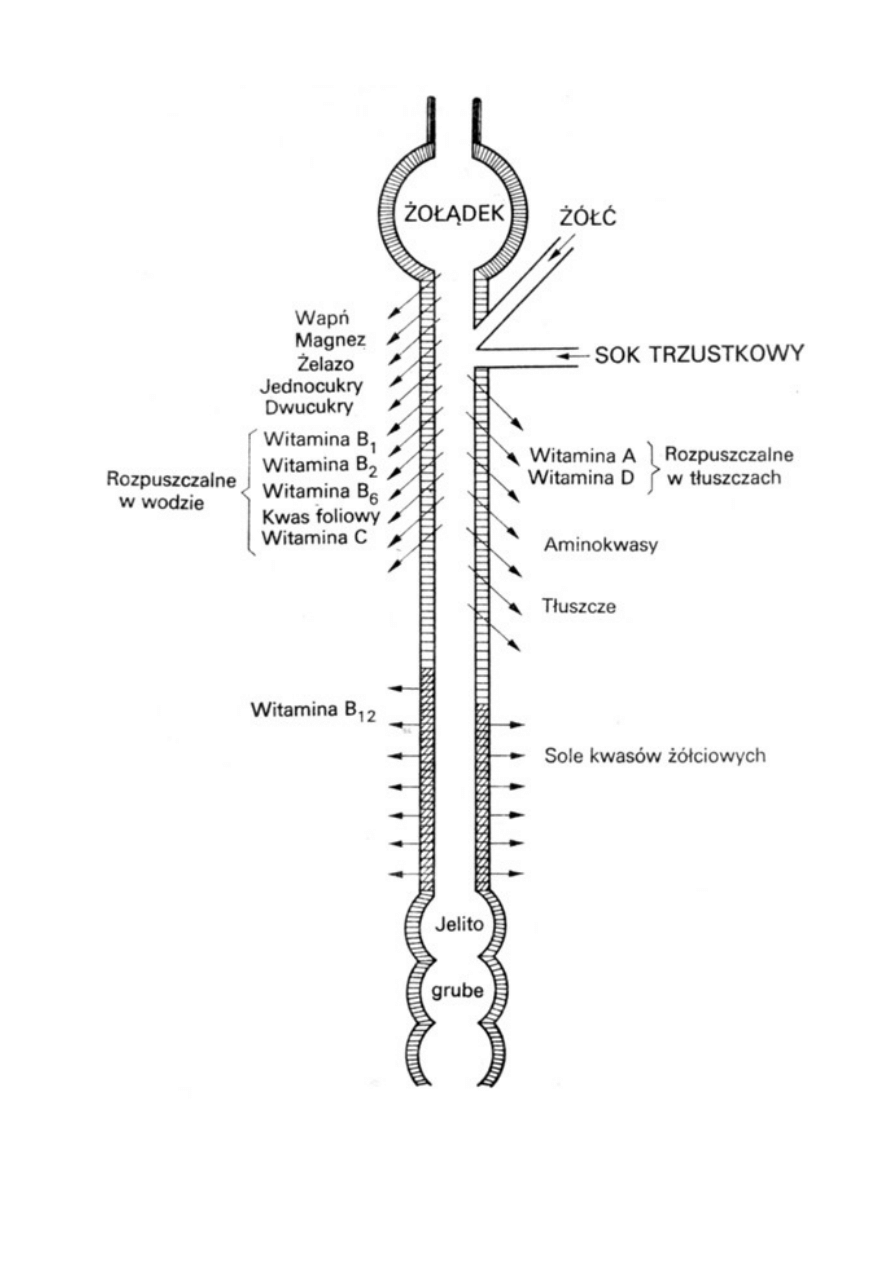
Wchłanianie witaminy B12
*Witamina B12 wchłania się w jelicie cienkim. Do prawidłowego jej wchłaniania
potrzebne jest połączenie z tak zwanym czynnikiem wewnętrznym, wydzielanym
w żołądku, oraz z jonami wapnia.
*Witamina B12 ulega związaniu z czynnikiem wewnętrznym, a następnie kompleks ten
jest przełączany przez kubilinę (apolipoproteinę o dużym powinowactwie,
DietetykaUP, Fizjologia człowieka, kolokwium nr 4 – Fizjologia trawienia
5

znajdującą się w receptorach w dystalnej części jelita cienkiego);
związanie z kubiliną zapoczątkowuje proces wchłaniania na drodze endocytozy;
następnie w erytrocytach jelita krętego witamina B12 jest przenoszona na białko
transportowe – transkobalaminę II, przenoszące tę witaminę w osoczu.
Sok trzustkowy
*odczyn zasadowy
*dobowe wydzialenie soku trzustkowego ok 1500 ml
*kationy: Na+, K+, Ca2+, Mg2+,
*aniony: HCO3-, Cl-, SO4, HPO4
*enzymy trawienne
Enzymy soku trzustkowego
*trypsyna - rozszczepia wiązania peptydowe aminokwasów zasadowych (argininy lub
lizyny) od końca karboksylowego
*chymotrypsyna – rozszczepia wiązania peptydowe aminokwasów aromatycznych od
końca karboksylowego
*karboksypeptydaza – rozszczepia wiązania peptydowe aminokwasów, które mają
pierścienie aromatyczne lub rozgałęzione boczne łancuchy elifatyczne, a także
aminokwasów, które mają zasadowe łancuchy boczne, od końca karboksylowego
*esterazy – rozszczepia estry karboksylowe; wymaga soli żółciowych
*nukleazy – rozkład DNA/RNA do nukleotydów – hydrolizuje wiązania estrowe
kwasów nukleinowych
*elastaza – rozszczepia wiązania peptydowe aminokwasów alifatycznych od końca
karboksylowego
*lipaza trzustkowa – rozkłada triglicerydy do monoglicerydów i kwasów tłuszczowych
*amylaza trzustkowa – hydrolizuje wiązania 1:4 alfa, powodując powstanie granicznych
alfa-dekstryn, maltotriozy i maltozy
Specyfikacja aktywacji i trawienia enzymów trzustki
*aktywacja enzymów trzustki:
^trypsynogen przechodzi w formę aktywną – trypsynę, w dwunastnicy pod
wpływem enzymu rąbka szczoteczkowego – enteropeptydazy.
^trypsyna katalizuje przejście chymotrypsynogenu do chymotrypsyny, proelastazy
do elastazy oraz prokarboksypeptydazy do karboksypeptydazy
DietetykaUP, Fizjologia człowieka, kolokwium nr 4 – Fizjologia trawienia
6

Skład żółci
*składniki nieorganiczne:
leucyna
fosfolipidy
kwas żółciowy
barwniki żółciowe
inne substancje nierozpuszczalne w wodzie (cholesterol, leki)
woda
kwasy tłuszczowe
tłuszcze i lipidy
Funkcje żółci
*obniża napięcie powierzchniowe wody, dzięki czemu zachodzi rozdrobnienie
tłuszczów i utrwalenie powstałej emulsji
*pomaga w absorbowaniu kwasów tłuszczowych, monoglicerydów, cholesterolu z
przewodu pokarmowego dzięki tworzeniu miceli
*aktywuje lipazę – enzym hydrolizuący tłuszcze
Rola żółci w trawieniu i wchłanianiu lipidów
*obniża napięcie powierzchniowe wody, dzięki czemu zachodzi rozdrobnienie
tłuszczów i utrwalenie powstałej emulsji
*pomaga w absorbowaniu kwasów tłuszczowych, monoglicerydów, cholesterolu z
przewodu pokarmowego dzięki tworzeniu miceli
*aktywuje lipazę – enzym hydrolizuący tłuszcze
Trawienie kontaktowe w jelicie cienkim
DietetykaUP, Fizjologia człowieka, kolokwium nr 4 – Fizjologia trawienia
7

*
trawienie przyścienne białek i polisacharydów opiera się na hydrolizie enzymatycznej
gdzie enzymy rąbka szczoteczkowego są przytwierdzona wiązaniami chemicznymi
do powierzchni błony enterocytu;
enzymy trawienia przyścinnego są wytwarzane w enterocytach i transportowane na
zewnętrzną powierzchnię błony komórkowej w wierzchołkowej części komórki,
gdzie ulegają zakotwiczeniu fragmentem łancucha;
*trawienie węglowodanów
enzymy rąbka szczoteczkowego trawią złożone węglowodany pokarmu takie jak:
sachacharoza, laktoza, maltoza, maltotrioza;
w rąbku szczoteczkowym obecne są enzymy: sacharaza, laktaza, maltaza, izomaltaza;
produktem trawienia maltozy i izomaltozy jest glukoza,
produktem trawienia sacharozy jest glukoza i fruktoza,
produktem trawienia laktozy jest glukoza i galaktoza.
*trawienie białek
peptydazy przytwierdzone są do błony komórkowej enterocytu;
ich zadaniem jest hydroliza peptydów powstałych w wyniku trawienia w świetle jelita
efektem trawienia przyściennego jest powstanie wolnych aminokwasów oraz
dipeptydów i tripeptydów
końcowe trawienie peptydów odbywa się dwuetapowo: w rąbku szczoteczkowym i
wewnątrz enterocytu
Wchłanianie cukrów, aminokwasów i lipidów
*wchłanianie lipidów
Proces wchłaniania produktów lipolitycznych składa się z następujących etapów:
1. wnikanie produktów do enterocytów,
2. resynteza triacylogliceroli i tworzenie chylomikronów w enterocytach
3. uwalnianie chylomikronów z enterocytów do naczyń chłonnych.
Produkty lipolityczne, uwolnione na powierzchni błony śluzowej jelita z
rozpadających się tu miceli żółciowych, dostają się na drodze dyfuzji przez
błonę komórkową enterocytu do jego wnętrza. Przy niższym pH kwasy
tłuszczowe ulegają protonowaniu, co zmniejsza ich micelarną rozpuszczalność
i ułatwia absorpcję przez enterocyty. Bezpośrednio po wchłonięciu zostają
połączone z glicerolem, tworząc triacyloglicerole, które po przyłączeniu
innych lipidów, takich jak cholesterol i jego estry z kwasami tłuszczowymi
oraz białka, zwłaszcza apoproteiny B, tworzą duże kompleksy, zwane
chylomikronami (są rozpuszczalne w wodzie) Są to największe ze znanych
lipoprotein przenoszących tłuszcze i zawierają około90% triacylogliceroli, 5—
9% fosfolipidów, 0,5— 1,0% białka, 0,7—1,5% cholesterolu i jego estrów oraz
l—7% wolnych kwasów tłuszczowych;
Po wytworzeniu chylomikrony uwalniane są z enterocytu przez błonę do przestrzeni
międzykomórkowej;
Chylomikrony z uwagi na duże wymiary nie mogą przenikać przez nabłonek naczyń
włosowatych kosmków i nie mogą tą drogą ulec wchłonięciu do krwi;
przedostają się więc do naczyń limfatycznych, charakteryzującymi się
szerokimi porami;
Wchłanianie lipidów odbywa się głównie w dwunastnicy i początkowym odcinku
jelita czczego;
Produkty trawienia lipidów ulegają wchłanianiu przez błonę wierzchołkową
enterocytu na zasadzie transportu ułatwionego oraz prostej dyfuzji;
*wchłanianie węglowodanów
Wchłanianie monosacharydów (glukoza, galaktoza, fruktoza) zachodzi szybko już
DietetykaUP, Fizjologia człowieka, kolokwium nr 4 – Fizjologia trawienia
8

w dwunastnicy i w górnym odcinku jelita czczego, kończąc się w zasadzie
w obrębie pierwszych 50 cm jelita czczego. Specjalne transportery
enterocytów łączą się z powstającymi w wyniku trawienia
monosacharydami i transportują je na zasadzie dyfuzji ułatwionej przez
warstwę lipidową błony komórkowej. Glukoza i galaktoza wchłaniają się
czynnie przy udziale transporterów. Proces ich wchłaniania wymaga
obecności jonów Na+, które zwiększają powinowactwo glukozy i galaktozy
do wspólnego transportera. Po wchłonięciu do enterocytu glukoza opuszcza
go za pośrednictwem transportera zgodnie ze swoim gradientem stężeń do
płynu zewnątrzkomórkowego i krwi. Proces wchłaniania glukozy zachodzi
nawet przeciwko gradientowi stężeń i trwa aż do zupełnego jej wchłonięcia
z treści jelitowej.
Wchłanianie fruktozy zachodzi około dwukrotnie wolniej niż glukozy i odbywa
się przy udziale transportera (transport ułatwiony).
LUB:
Transport cukrów jest uzależniony od stężenia jonów Na+ w świetle jelita; duże
stężenie Na+ na powierzchni komórek błony śluzowej jelita ułatwia napływ
glukozy do enterocytów, podczas gdy małe stężenie jonów Na+ utrudnia ten
proces; jest to spowodowane tym, że glukoza i jony Na+ korzystają z tego
samego kontransportera – symportu, czyli transportera glukozy zależnego od
sodu (SGLT); stężenie jonów Na+ w enterocytach pozostaje małe. Jony Na+
przechodzą ze światła jelita do komórek zgodnie z gradientem stężeń; glukoza
przechodzi łącznie z jonami Na+, a nastapnie uwalniana jest w komórce; jony
Na+ transportowane są do bocznych przestrzeni międzykomórkowych, a
glukoza przy udziale białka transportującego GLUT-2 do przestrzeni
międzykomórkowej, a potem do naczyń włosowatych; transport glukozy jest
przykładem transportu wtórnie aktywnego – energia potrzebna do transportu
glukozy jest dostarczana pośednio, poprzez aktywny transport jonów Na+ poza
komórkę; W ten sposób gradient stężeń jonów Na+ w poprzek błony zwróconej
do światła jelita pozostaje zachowany, co z kolei ułatwia wchłanianie glukozy;
Natomiast transport fruktozy ze światła jelita do enterocytów odbywa się na drodze
dyfuzji ułatwionej przy udziale białka transportującego GLUT-5, a następnie
do przestrzeni międzykomórkowej przy udziale GLUT-2; część cząsteczek
fruktozy jest w enterocytach przekształcana w cząsteczki glukozy
*wchłanianie białek
Produkty hydrolizy białka wchłaniają się w jelitach, częściowo w postaci wolnych
aminokwasów, a częściowo jako składowe małych peptydów, głównie
dipeptydów i tripeptydów. Te ostatnie ulegają ostatecznie hydrolizie do
aminokwasów w samych enterocytach. Wchłanianie aminokwasów
zachodzi niemal tak szybko, jak ich uwalnianie z peptydów pod wpływem
proteaz. Ciekawe, że transport wolnych aminokwasów z jelita do krwi
odbywa się wolniej niż transport dipeptydów i tripeptydów. Transportery
dla wolnych aminokwasów są inne niż te dla oligopeptydów. Wchłanianie
aminokwasów i oligopeptydów do enterocytów zachodzi na drodze
czynnego transportu zachodzącego przeciwko gradientowi chemicznemu i
prowadzącego do nagromadzenia tych aminokwasów w enterocytach, skąd
już na drodze dyfuzji ułatwionej zgodnie z gradientem stężeń przedostają
się one do płynu zewnątrzkomórkowego i dalej do krwi. Wchłanianie
zachodzi sprawnie pod warunkiem, że aminokwasy są L-izomerami. D-
DietetykaUP, Fizjologia człowieka, kolokwium nr 4 – Fizjologia trawienia
9

izomery wchłaniają się bardzo powoli. Wchłanianie aminokwasów zachodzi
na zasadzie podobnej do wchłaniania monosacharydów. Dotyczy to
zwłaszcza korzystania z transporterów wspólnych z jonami Na+.
Elektrogeniczna pompa sodowo-potasowa w bocznej ścianie enterocytów
zapewnia usuwanie Na
+
z ich wnętrza i stanowi jednocześnie siłę napędową
czynnego transportu aminokwasów i oligopeptydów w przeciwległej,
zwróconej do światła jelita, błonie komórkowej. Źródłem energii
podtrzymującym aktywność tej pompy jest ATP, który zapewniając
utrzymanie odpowiednich gradientów Na
+
i K
+
wpływa na wchłanianie
aminokwasów.
W przypadku di- i tripeptydów transport odbywa się przy udziale jonów H+
zamiast jonów Na+
DietetykaUP, Fizjologia człowieka, kolokwium nr 4 – Fizjologia trawienia
10
DietetykaUP, Fizjologia człowieka, kolokwium nr 4 – Fizjologia trawienia
11

Rola gastryny, sekretyny, CCK w regulacji trawienia
*gastryna – pobudza wydzielanie soku żołądkowego (głównie),
pobudza gruczoły jelita cienkiego do wydzielania soku jelitowego,
komórki zewnątrzwydzielnicze trzustki do wydzielania soku trzustkowego,
komórki wątroby do wydzielania żółci;
nasila motorykę żołądka oraz jelit, skurcze ścian pęcherzyka żółciowego
*CCK (cholecystokinina) – pobudza wydzielanie soku trzustkowego (głównie),
soku żołądkowego, jelitowego, żółci;
wzmaga perystaltykę jelit;
hamuje perystaltykę żółądka,
pobudza uwalnianie insuliny z wysp trzustkowych (efekt inkretynowy)
*sekretyna – pobudzenie wydzielania soku trzustkowego ,
hamowanie wydzielania kwasu solnego i gastryny w żołądku,
opóźnienie oprózniania żołądka,
pobudzenie wydzielania gruczołów dwunastniczych,
pobudzenie sekrecji żółci;
Rola leptyny i greliny w regulacji pobierania pokarmu
*
Leptyna to hormon produkowany przez komórki tłuszczowe. Jest przenoszona przez krew
do mózgu, gdzie działa na neurony w przysadce mózgowej, które hamują apetyt i
potrzebę jedzenia. Leptyna wstrzymuje więc chęć pobierania pokarmu, gdy zapasy
tłuszczu są duże, a wzmaga apetyt, gdy są małe. Po związaniu leptyny z receptorami
w podwzgórzu, neurony przestają wytwarzać neurotransmiter - neuropeptyd Y, który
jest stymulatorem apetytu.
Spożyte i wchłonięte w przewodzie pokarmowym składniki pokarmowe
pobudzają adipocyty do wydzielania leptyny bezpośrednio i pośrednio poprzez
insulinę z wysp trzustkowych. Leptyna na zasadzie ujemnego sprzężenia
zwrotnego poprzez ośrodki pokarmowe w podwzgórzu hamuje przyjmowanie
pokarmów i tym samym hamuje magazynowanie energii w tkance tłuszczowej
*Grelina stymuluje pobieranie pokarmów i poprzez aktywację neuronów NPY
(neuropeptydowych) w podwzgórzu zwiększa bilans energetyczny
DietetykaUP, Fizjologia człowieka, kolokwium nr 4 – Fizjologia trawienia
12
Wyszukiwarka
Podobne podstrony:
Fizjologia Krew zagadnienia III koło
FIZJOLOGIA zagadnienia IV zal 3
FIZJOLOGIA, zagadnienia IV zal 2
FIZJOLOGIA, zagadnienia IV zal
wykaz zagadnien do iv kolokwium 2011 2012, materiały farmacja, Materiały 3 rok, Farmakognozja, Do ko
fizjologia roslin IV kolo, Szkoła Rolnictwo studia, Szkoła, Materiały studia, fizjo roslin
FIZJOLOGIA zagadnienia IV zal 2
Fizjologia krążenia zagadnienia (II kolokwium)
Opracowane zagadnienia na koło z podstaw turystyki, Notatki na koła
IV koło
fizjologia trawienia 1
ZAGADNIENIE NA KOŁO RF 12
Zagadnienia na kolo z patologii społ
Postępowanie administracyjne - zagadnienia, IV
ZAGADNIENIA na koło
więcej podobnych podstron