Opracowanie
wykładów
Biofizyka
MC_OMEN
BIOFIZYKA NARZADOW
1. Zasób energetyczny mięśnia
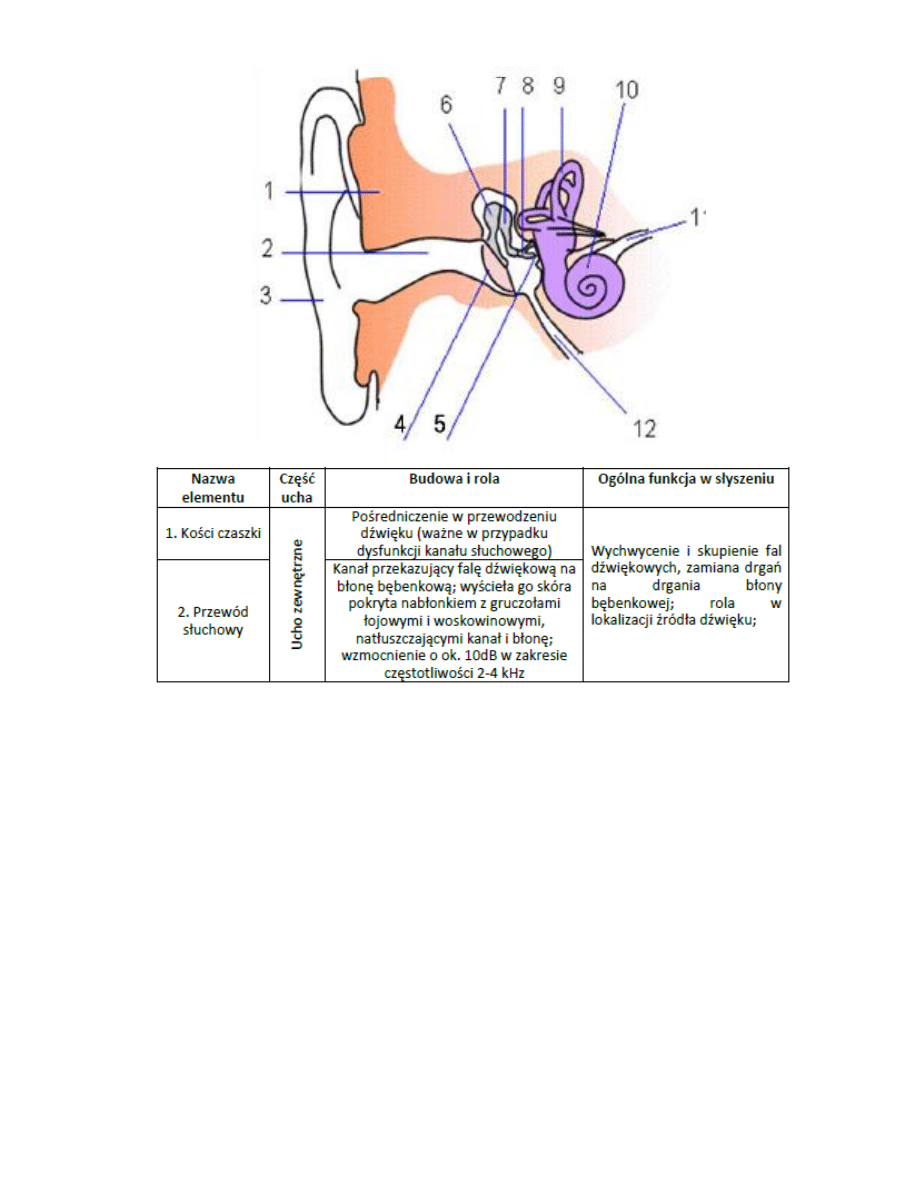
Najczęstszym źródłem energii dla mięśnia jest zmagazynowany w nim glikogen lub glukoza dostarczona
przez krew
2. Rodzaje tkanki mięśniowej
-
Mięśnie poprzecznie prążkowane: przytwierdzone są do kości za pomocą ścięgien – twardych i
elastycznych włókien tkanki łącznej. Mogą byd także przytwierdzone do skóry. Skurcze tych mięśni
są szybkie, krótkotrwałe i zależne od naszej woli (w przypadku zdrowego organizmu). Mięśnie
poprzecznie prążkowane charakteryzują się wysokim stopniem uporządkowania włókien
kurczliwych.
-
Tkanka mięśniowa gładka; obecna jest w ściankach przewodu pokarmowego, układu krwionośnego,
moczo-płciowego, limfatycznego, oddechowego. Skurcze mięsni gładkich są powolne, długotrwałe i
nie zależą od naszej woli. Komórki mięsni gładkich nie wykazują poprzecznego prążkowania, są
krótkie (u człowieka 0.02-0.05mm), o wrzecionowatym kształcie, ostro zakooczone. Zawierają jedno
jądro położone centralnie w komórce.
-
Tkanka mięśnia sercowego: występuje jedynie jako budulec serca. Wykonuje rytmiczne,
automatyczne, dośd szybkie i krótkotrwałe skurcze, które nie zależą od naszej woli. Komórki tej
tkanki są krótkie (u człowieka 0.05-0.12mm), o średnicy do 0.02mm. Zawierają jedno lub dwa jądra,
położone centralnie. Wykazują rozgałęzienia i silnie pofałdowane zakooczenia, a także delikatne
poprzeczne prążkowan
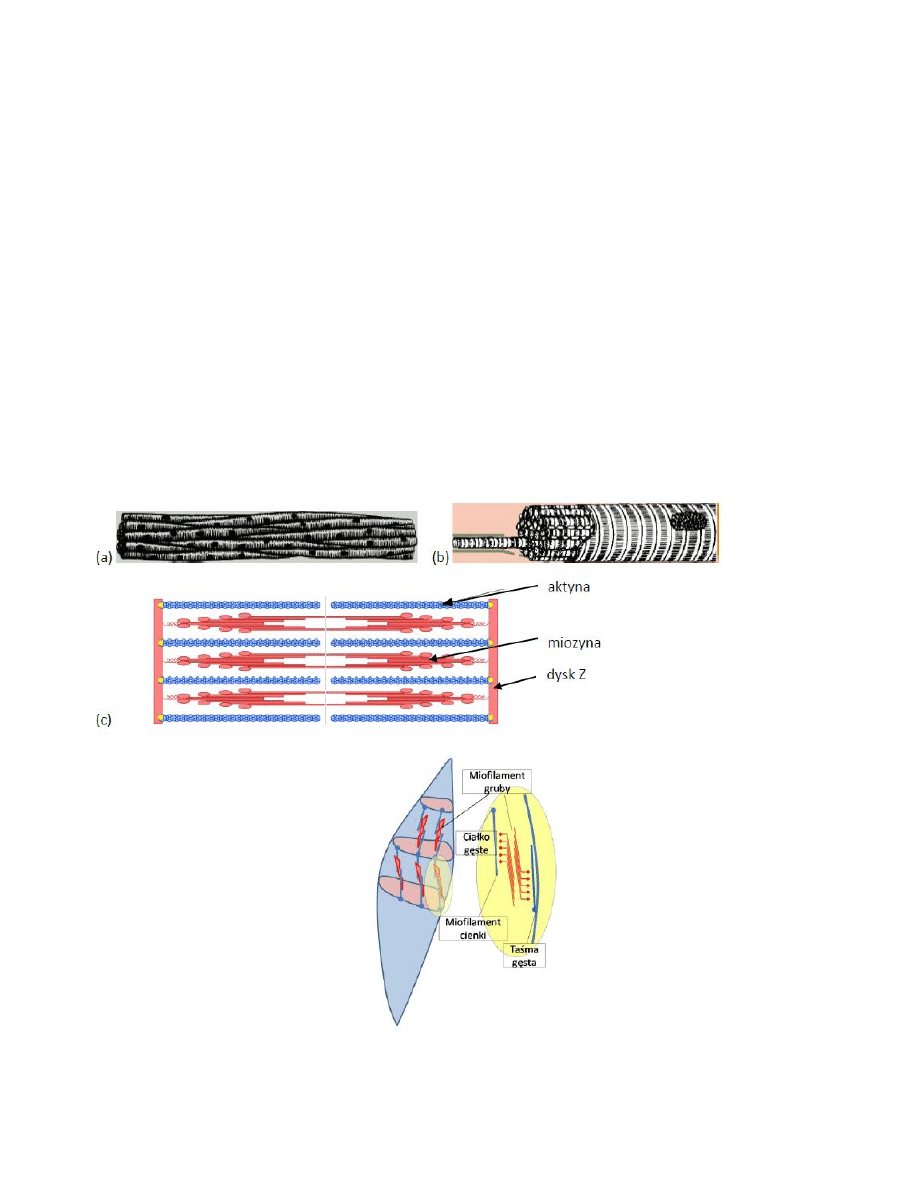
3. Budowa mięśnia poprzecznie prążkowanego:
4. Budowa komórki mięśnia gładkiego
5. Skurcze mięśnia :
Istotą skurczu jest zmiana długości mięśnia oraz/lub jego naprężenie. Zmiana długości lub napięcia mięśnia
wywiera siłę mechaniczną na miejsca przyczepu mięśnia lub wokół narządu otoczonego przez mięsieo
okrężny (np. jamy ustnej).
6. Podizał zmiany napięcia mięśnia i skruczy

-
Izotoniczne - gdy zmienia się długośd mięśnia przy stałym poziomie napięcia mięśniowego.
Wynikiem skurczu jest zmiana położenia części ciała względem siebie czyli w konsekwencji ruch.
-
Izometryczne – gdy wzrasta napięcie mięśnia przy jego stałej długości. Wynikiem nie jest ruch, ale
utrzymanie części ciała w stałym położeniu, np. odkręcanie mocno przykręconych śrub, stanie,
trzymanie ciężarów
-
Auksotoniczny – gdy następuje zarówno zmiana długości jak i napięcia mięśni (np. przy chodzeniu,
bieganiu).
7. Inny podział
-
Skurcz tężcowy – jeżeli impulsy docierają w czasie krótszym niż zdąży nastąpid rozkurcz mięśnia
-
Skurcz tężcowy niezupełny – jeżeli impulsy docierają do mięśnia w czasie dłuższym niż skurcz
-
Skurcz tężcowy zupełny (pojedyoczy) – wywołany przez pojedyoczy impuls nerwowy lub
elektryczny, po skurczu następuję rozkurcz, bardzo krótki skurcz.
8. Ślizgowa teoria skurczu

BIOFIZYKA ZMYSLU SLUCHU
1. Prędkośd rochodzenia sięfali dźwiękowej dla fali poprzecznej
E
c
2. Fali podłużnej
K
c
3. Impedancja akustyczna
Zdolnośd przewodzenia fali akustycznej w ośrodku
c
Z
w
0
4. Współczynnik odbicia fali
)
(
2
1
2
1
0
Z
Z
Z
Z
I
I
R
r
5. Natężenie fali
c
p
dtA
dW
I
kw
0
2
6. Budowa układu słuchowego
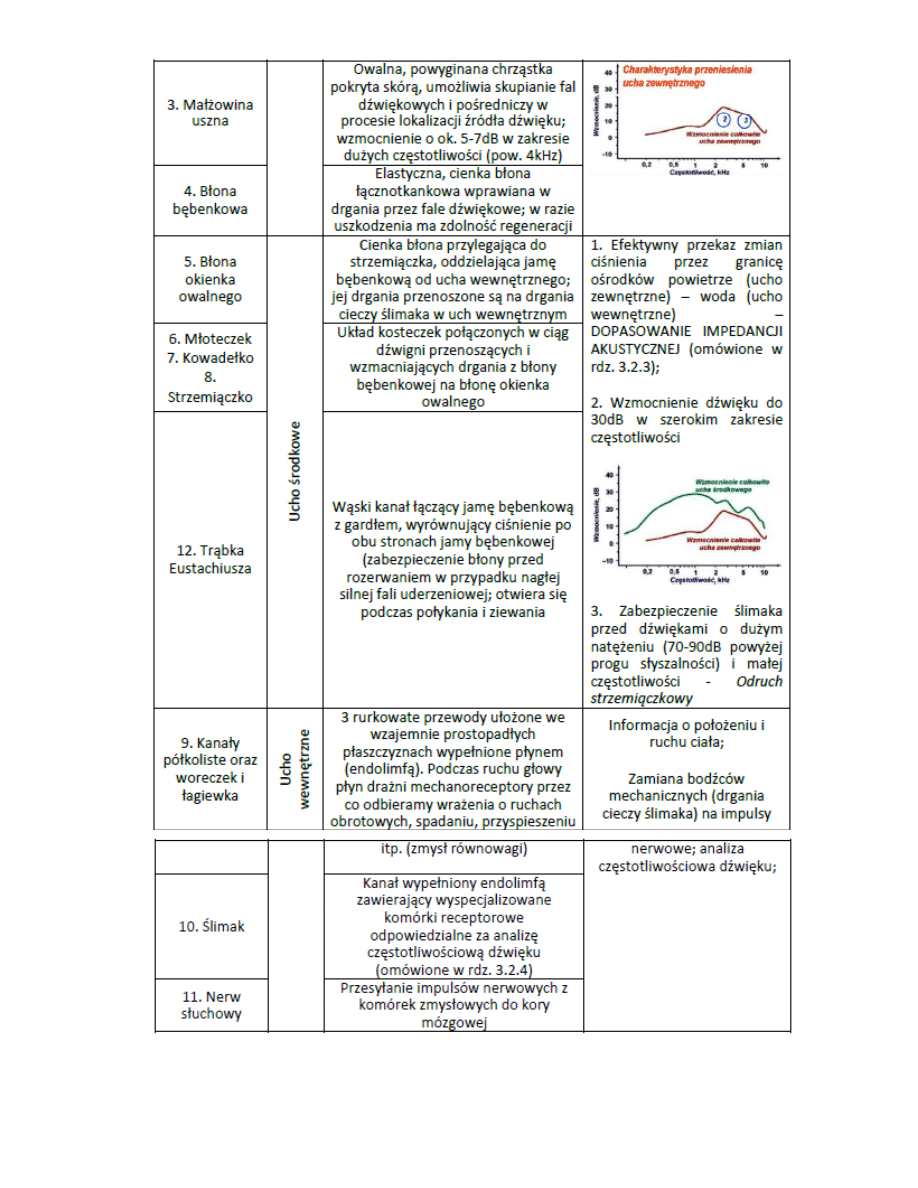
7. Mechanizm dopasowania impedancji w uchu środkowym
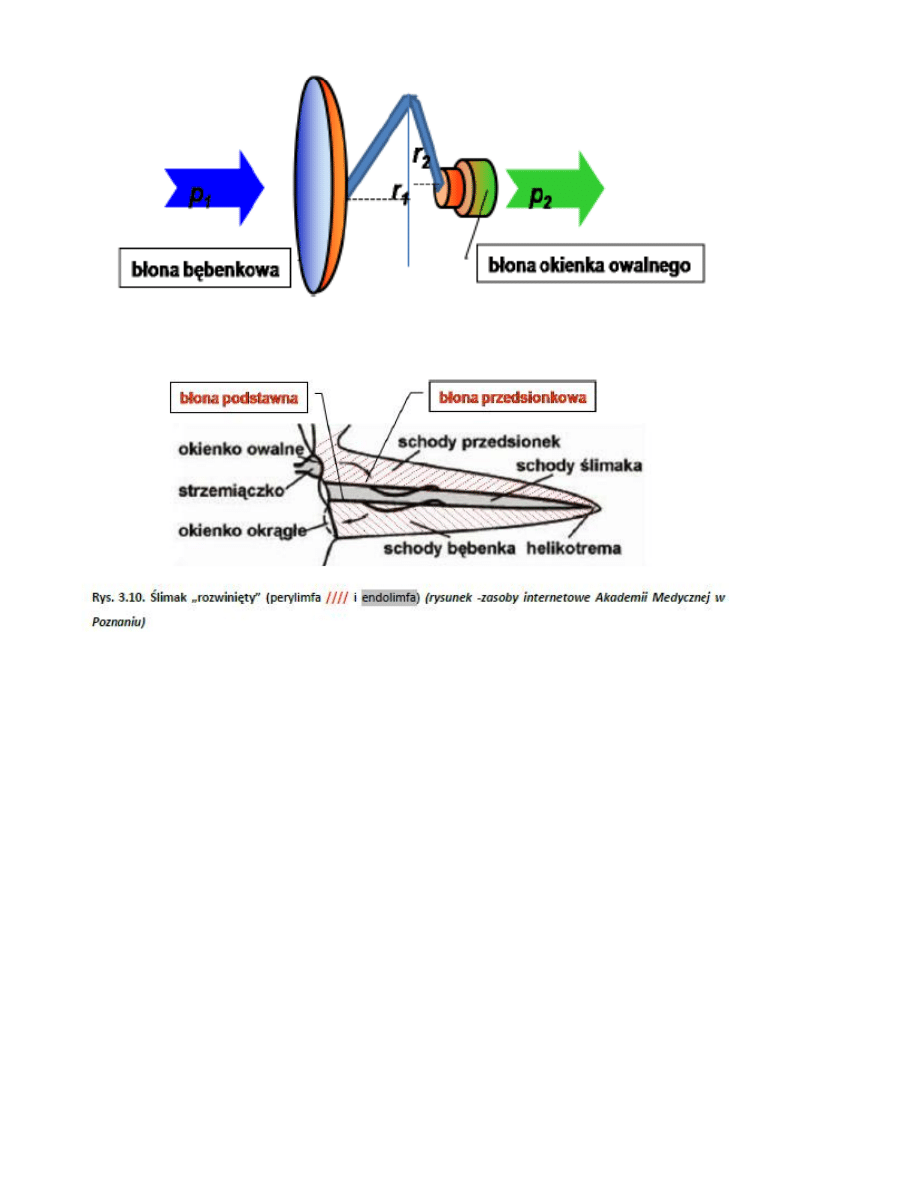
8. Budowa ślimaka (rozwinięty)
9. Mechanizm analizy dźwięku w ślimaku
-
ruch strzemiączka powoduje drgania błony okienka owalnego i wywołuje rozchodzące się
zaburzenie gęstości perylimfy wzdłuż ślimaka;
-
gdy fala ciśnienia dociera do helikotremy przechodzi dalej do obszaru schodów bębenka i
ostatecznie do okienka okrągłego, powodując jego odkształcenie
-
rozchodzenie się fali jest na tyle szybkie (prędkośd dźwięku w cieczy ok. 1500m/s, droga „tam i z
powrotem” ok. 7cm), że drgania błony okienka owalnego i okrągłego są w przeciwfazie.
-
fala rozchodząc się w perylimfie wzbudza drgania błony podstawnej, które są mocno zależne od
częstotliwości pobudzającego dźwięku
Elastycznośd błony jest mniejsza w niedużej odległości od okienka owalnego, a w pobliżu helikotre my błona
podstawna staje się szersza i mniej sztywna. Dźwięki o dużych częstotliwościach najsilniej wzbudzają drgania
błony w pobliżu podstawy błony, a tony niskie – w pobliżu helikotremy. Widzimy więc, w jaki sposób
dokonywana jest w ślimaku konwersja częstotliwośd - miejsce. Dzięki temu ślimak dokonuje analizy sygnału
pod względem częstotliwości, co nazywamy selektywnością częstotliwościową
10. Przekrój poprzeczny ślimaka
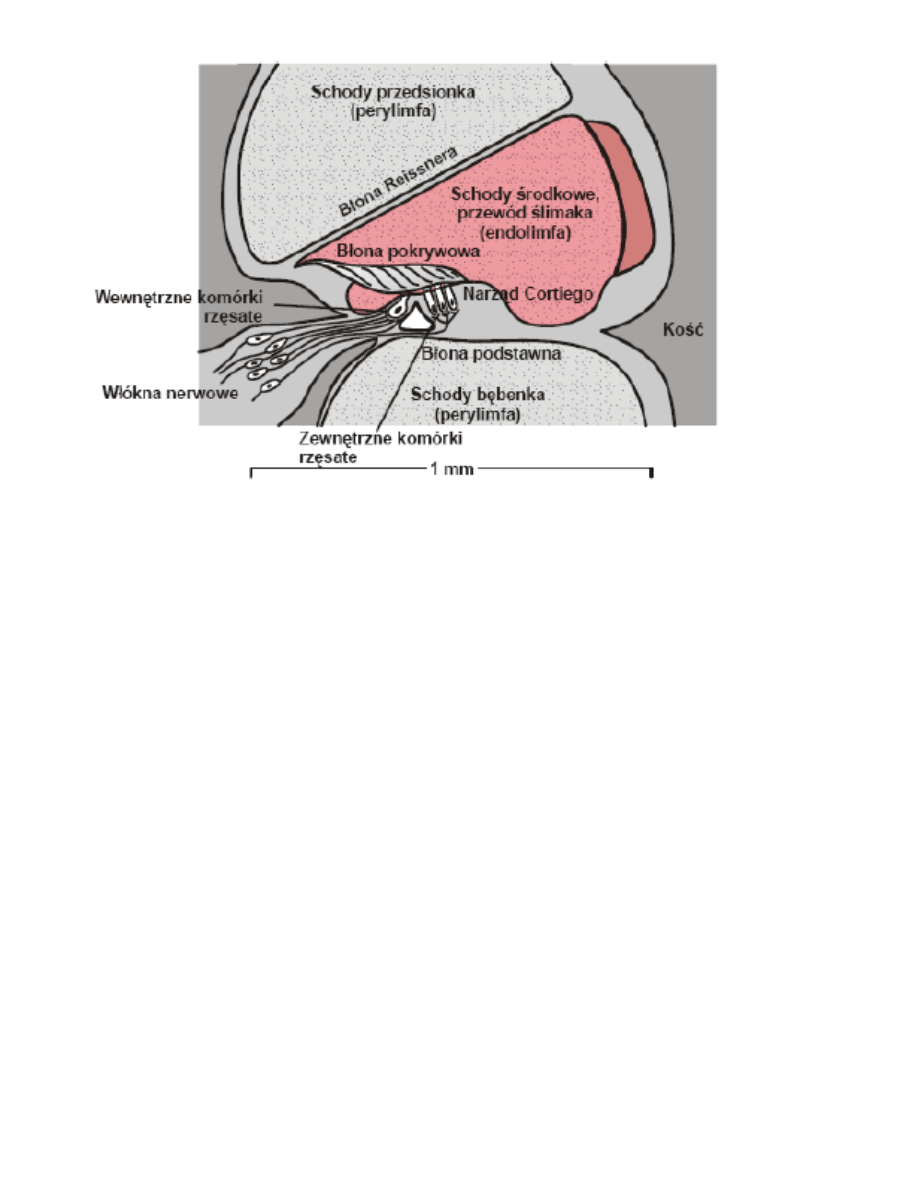
Komórki rzęsate wewnętrzne (ok. 3500 sztuk) wyposażone są w rzęski (ok. 40 rzęsek każd a), które w przypadku
braku wygięcia błony podstawnej prawdopodobnie nie mają kontaktu z błoną pokrywną. Gdy powstaje wygięcie
błony, na rzęski zaczynają działad siły ścinające, co powoduje otwieranie i zamykanie się kanałów jonowych i
przepływ jonów potasowych z endolimfy. W ten sposób generowana jest chwilowa zmiana potencjału komórek
rzęskowych, odbierana przez synapsy dołączonych do nich aferentnych komórek nerwowych i przenoszona do
wyższych partii układu nerwowego.
Grupa komórek rzęsatych zewnętrznych jest ponad 3-krotnie bardziej liczna jak również ok. 3 razy bardziej
„urzęstniona”. Połączona jest z 1800 neuronów eferentnych (przekazujących sygnały z mózgu). Komórki te
odpowiedzialne są za wzmocnienie wrażenia słuchowego dzięki zdolności ich kurczenia się pod wpływem zmiany
potencjału. Podobnie jak w przypadku komórek wewnętrznych, ruch błony podstawnej ku górze prowadzi do
otwarcia kanałów potasowych i napływu jonów K
+
z endolimfy. Wywołany tym napływem wzrost potencjału
powoduje skrócenie komórek rzęsatych zewnętrznych, a przez to większe zbl iżenie błony pokrywnej do komórek
rzęsatych wewnętrznych. Tym samym możliwe jest bardziej efektywne stymulowanie ich, a przez to wzmocnienie
wrażenia słuchowego. Mechanizm ten jest szczególnie ważny w przypadku dźwięków cichych.
11. Fale akustyczne
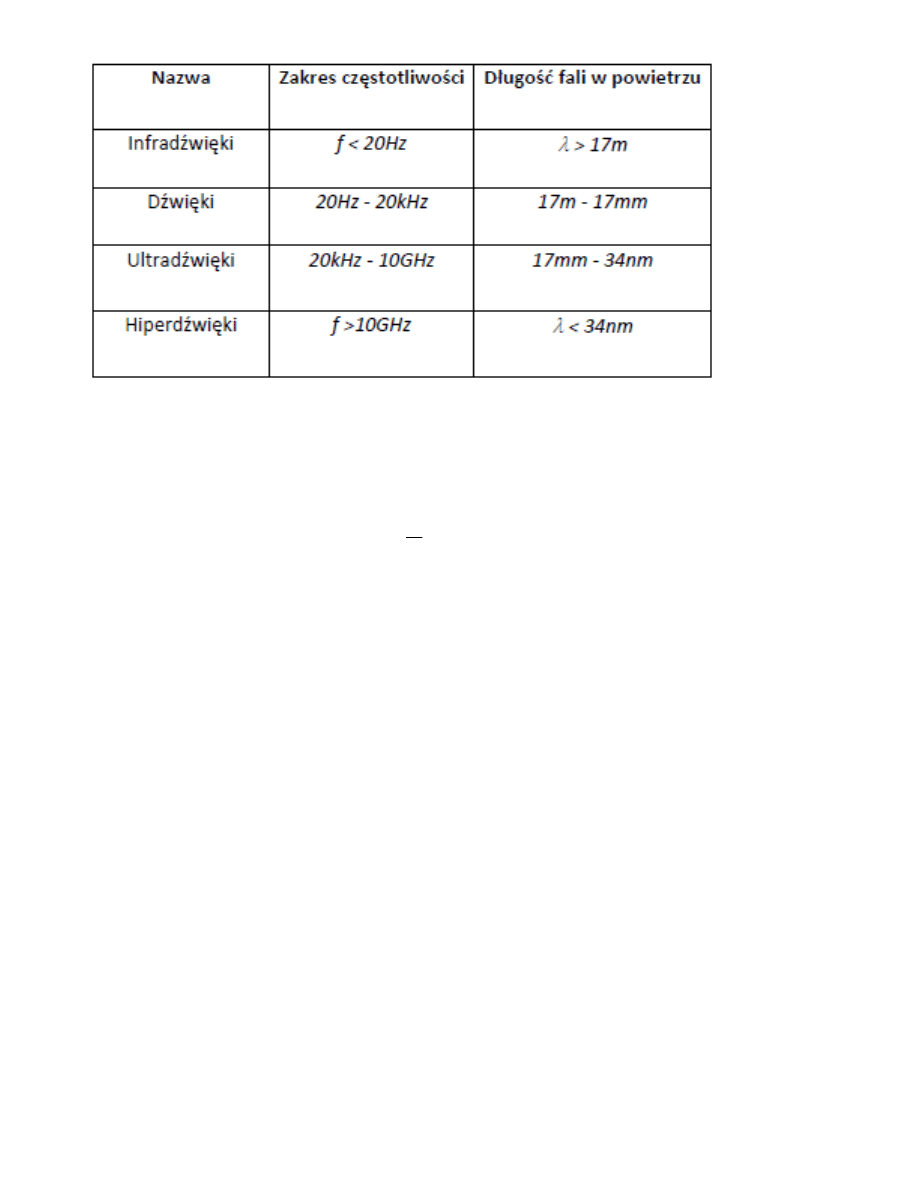
12. Dźwięk i wrażenie słuchowe
Z czysto fizycznego punktu widzenia, dźwięk opisywany jest przez tzw. parametry obiektywne (fizyczne):
częstotliwośd, natężenie i strukturę widmową. Wrażenie słuchowe jakie odbieramy wykorzystuje w opisie tzw.
cechy subiektywne (psychologiczne): wysokośd dźwięku, jego głośnośd i barwę. Obiektywne i subiektywne
cechy dźwięku odpowiadają sobie w znacznym stopniu.
.13 Poziom natężenia dźwięku (głośnośd)
0
1
log
10
I
I
L
- prawo Webera-Fechnera
13. Barwa dźwięku
Zdefiniowana jest jako atrybut wrażenia dźwiękowego umożliwiający rozróżnianie głosów różnych osób lub
tych samych nut granych na różnych instrumentach. Fizycznie wiąże się ze strukturą widmową
charakterystyczną dla danego źródła dźwięku.
14. Lokalizacja źródła dźwięku
Słyszenie dwuuszne (binauralne) sprawia, że sygnały docierające do nas ze źródła umieszczonego odosiowo (na
prawo lub na lewo względem osi pionowej ciała) różnią się zarówno momentem dotarcia do każdego z uszu
oraz natężeniem fali docierającej do nich.
15. Międzyuszna róźnica natężenia
Międzyuszna różnica natężenia zwana także międzyuszną różnicą poziomów wynika bezpośrednio z faktu, że
głowa i małżowiny uszne stanowią przeszkodę na drodze fali akustycznej i przez to dźwięk docierający z boku
odbierany jest przez ucho bliższe jako głośniejszy. W przypadku fal o długości porównywalnej lub większej od
rozmiarów głowy (częstotliwości poniżej 1500Hz), fala dźwiękowa ugina się na przeszkodzie i dociera także do
ucha dalszego różniąc się bardzo nieznacznie poziomem natężenia. Natomiast dla fal krótszych przeszkoda w
postaci głowy powoduje powstanie tzw. cienia akustycznego po stronie ucha dalszego i wówczas międzyuszna
różnica natężeo może wynosid nawet 30dB.
BIOFIZYKA ZMYSŁU OKA
1. Odbicie i załamanie światła
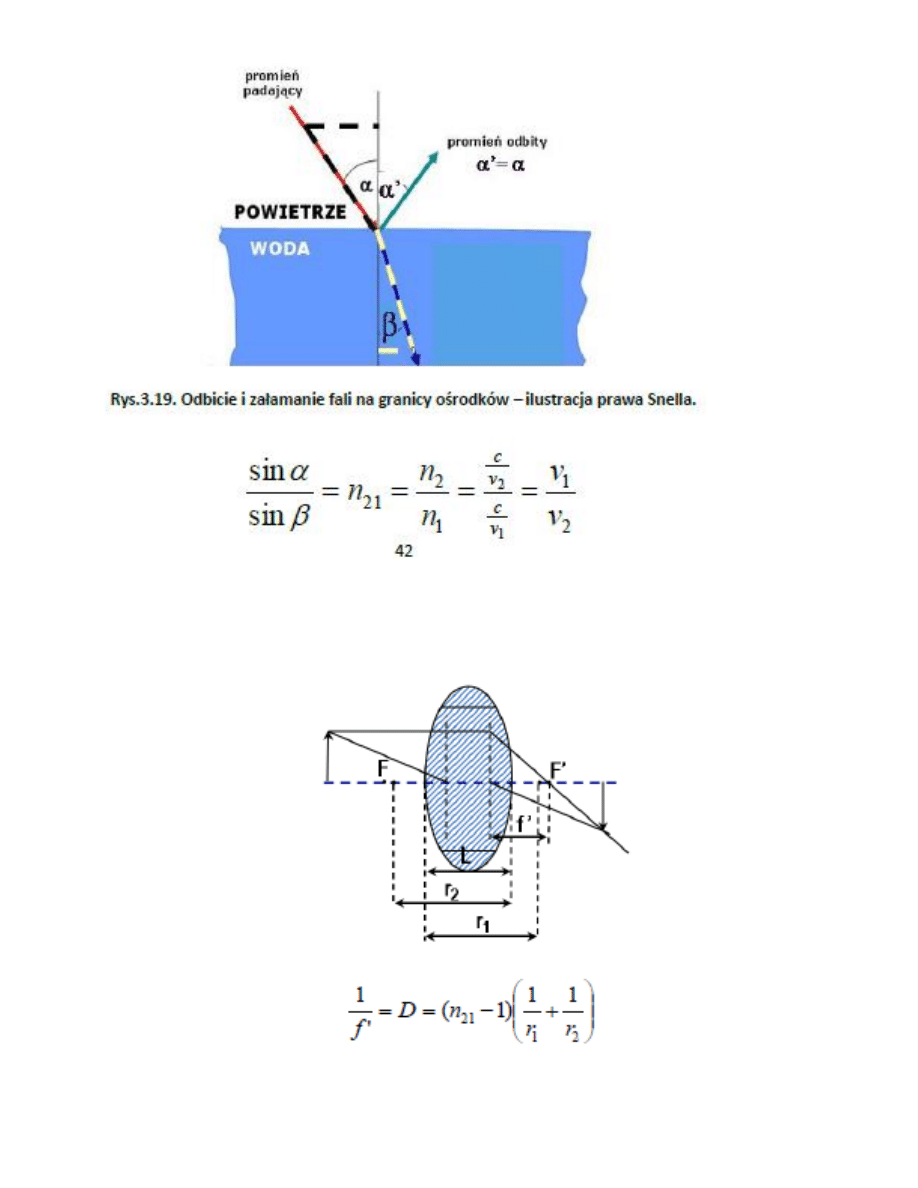
2. Soczekwa
to układ optyczny uformowany przez dwie powierzchnie sferyczne lub cylindryczne załamujące światło (Rys.
3.20) tworzące bryłę wypełnioną materiałem o innym współczynniku załamania niż otoczenie. W odróżnieniu
od powierzchni płaskiej, krzywizny soczewki umożliwiają skupienie lub rozproszenie wiązki za soczewką.
3. Zdolnośd skupiająca
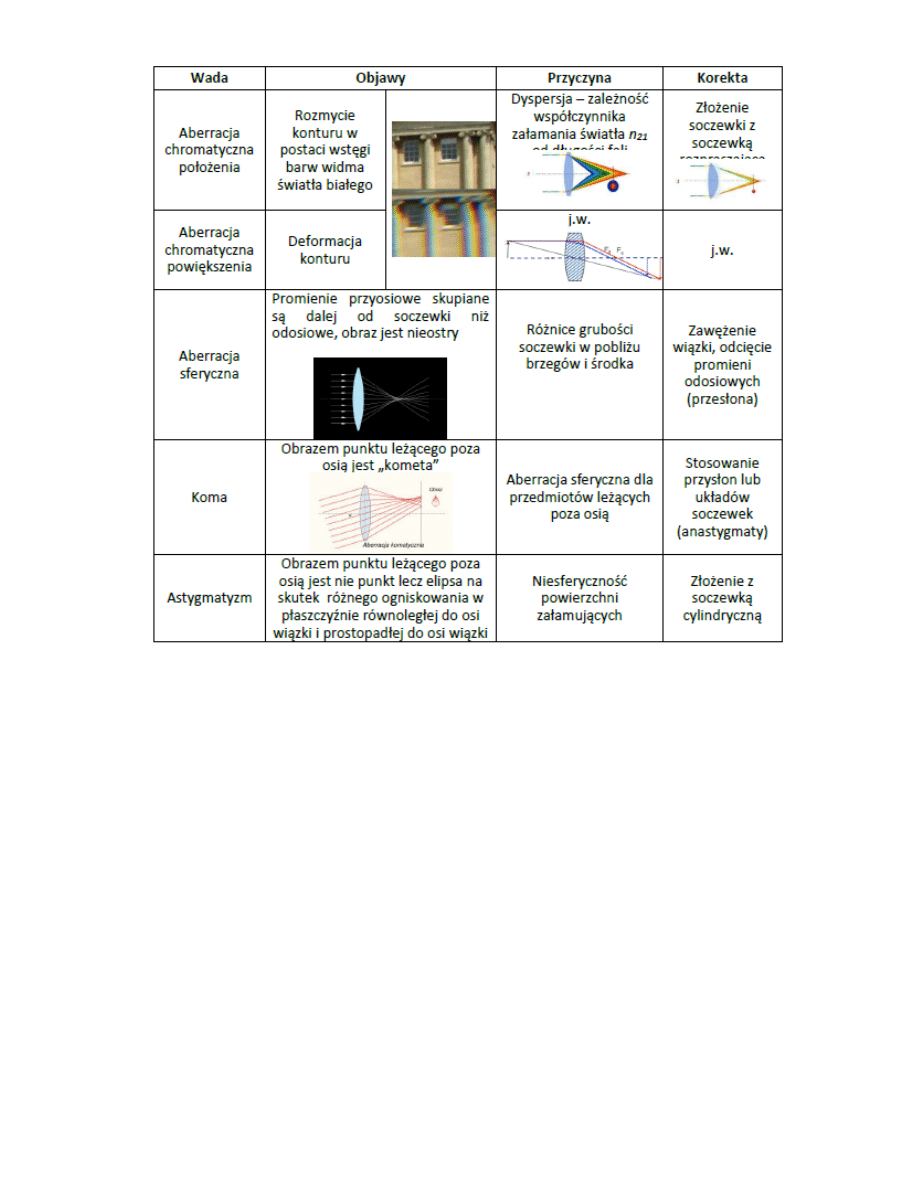
4. Wady odwozrowao soczewek optycznych
5. Budowa oka
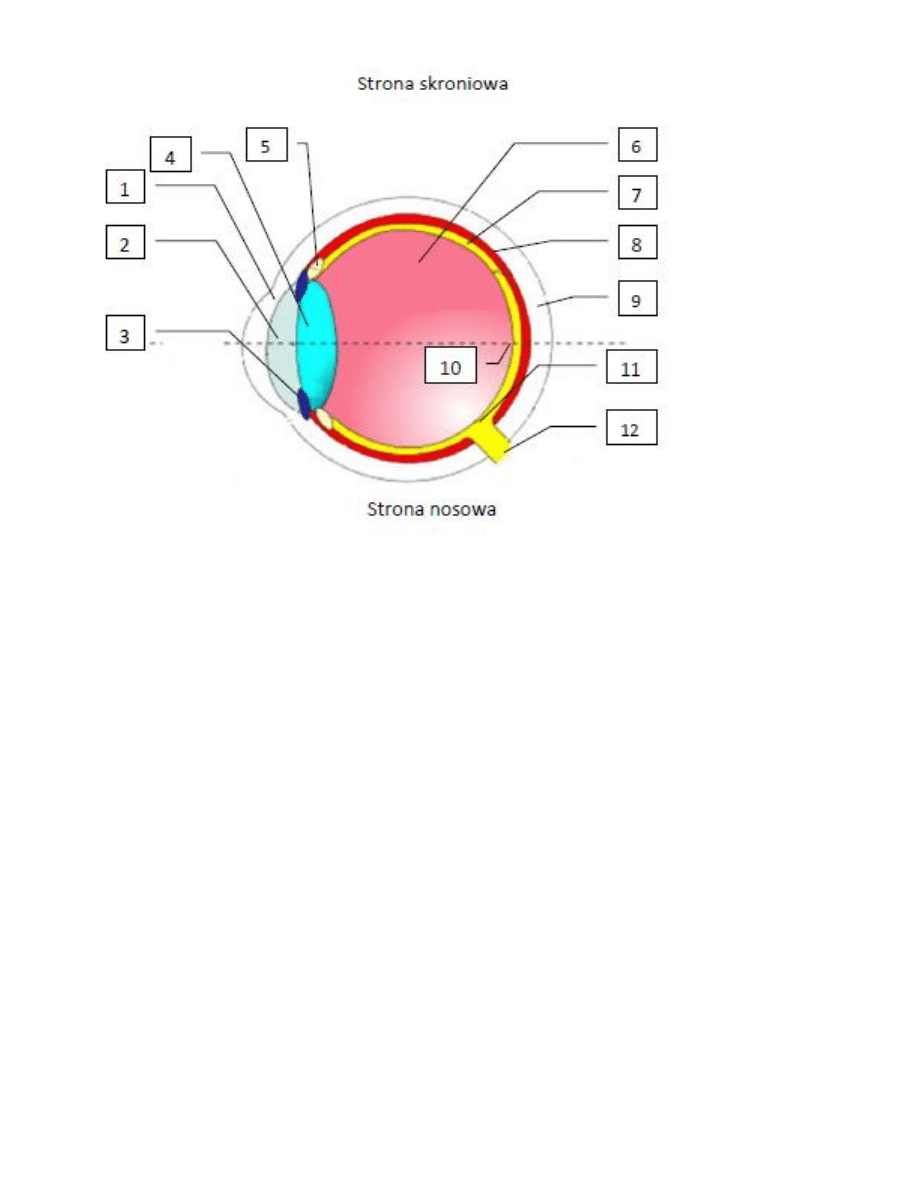
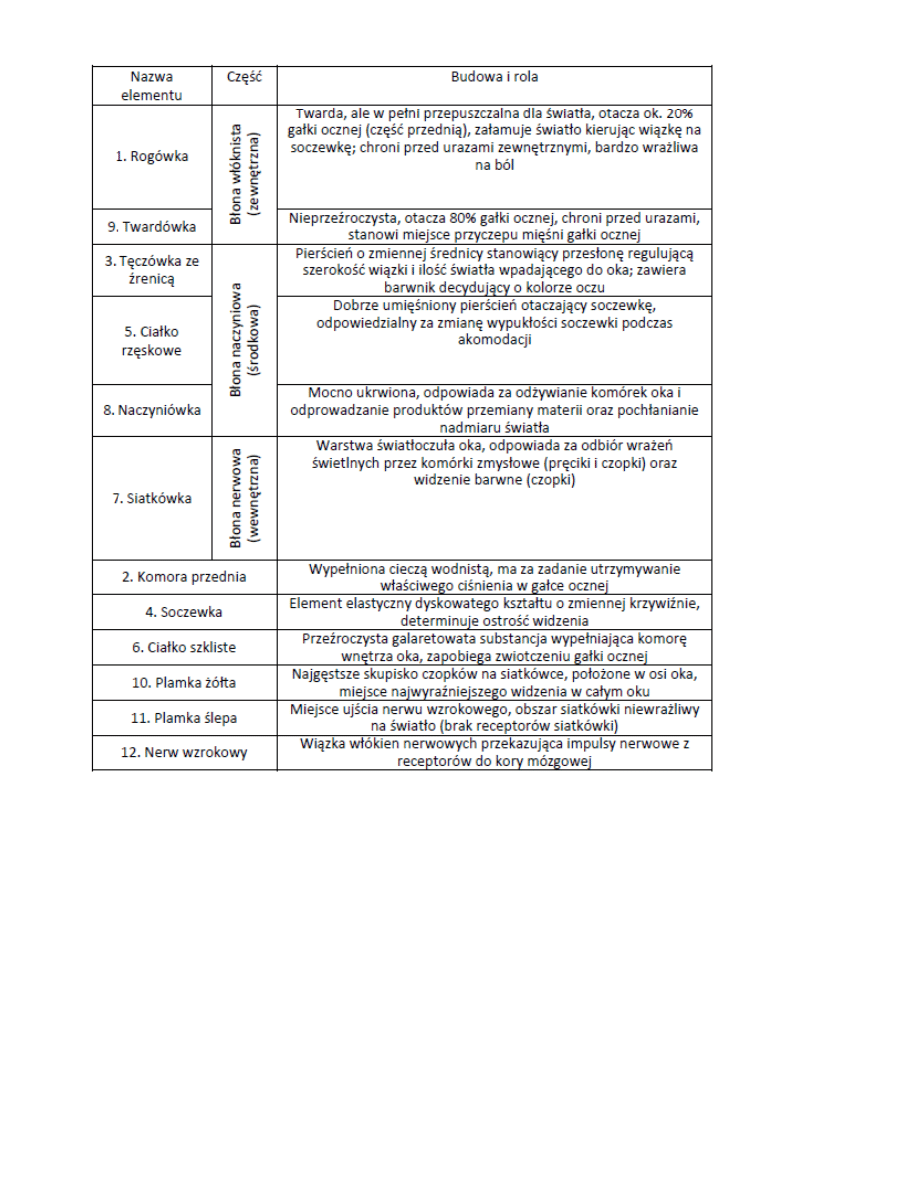
6. Opis budowy oka
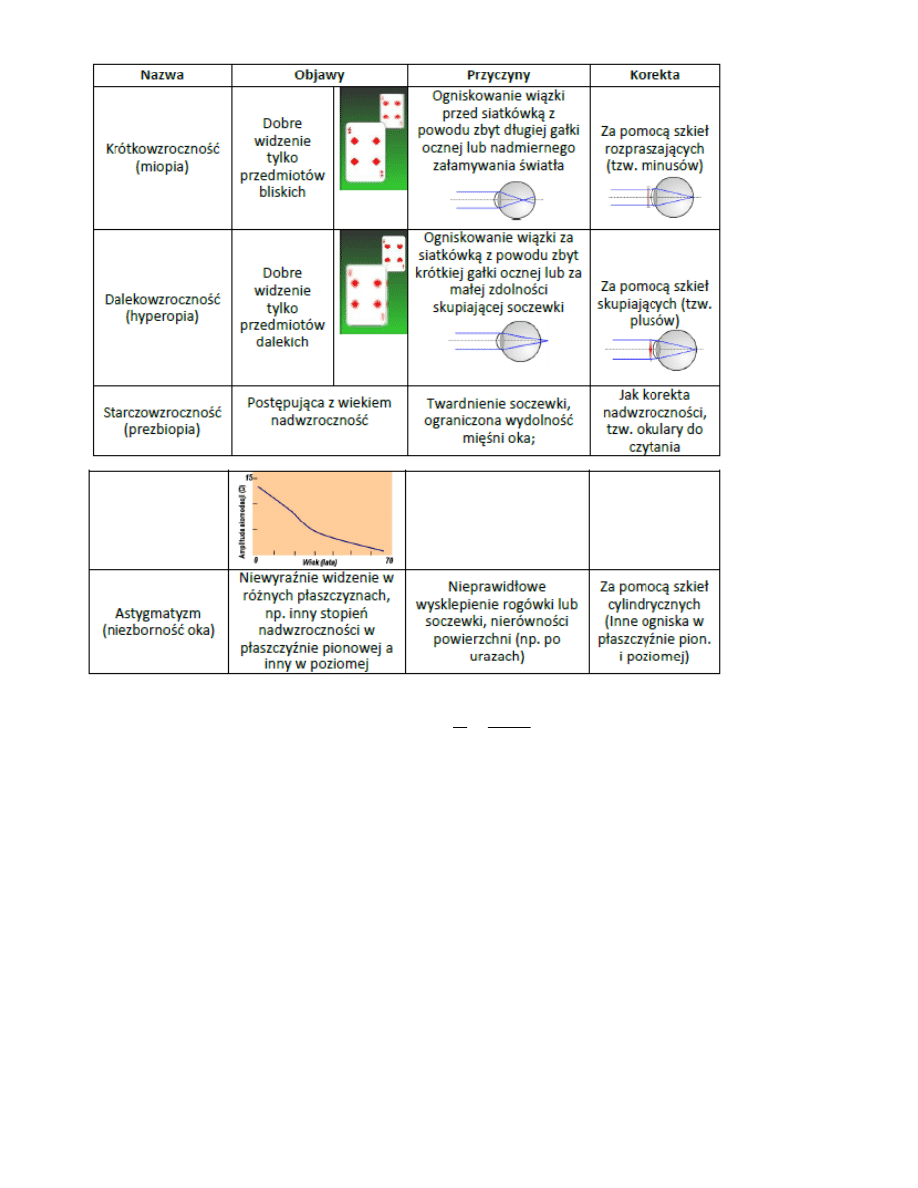
7. Wady wzroku
8. Kątowa rozdzielczośd oka
22
.
1
1
d
D
k
Jest to odwrotnośd najmnijszego kąta pod jakim widoczne są obrazy dwóch rozróżnialnych jeszcze
punktów
9. Czopki i pręciki
Siatkówka posiada wielowarstwową i niejednolitą strukturę komórkową, w której elementami światłoczułymi ją
dwa typy receptorów: czopki i pręciki. W obu typach komórek pod wpływem kwantów promieniow ania
inicjowane są reakcje chemiczne prowadzące do powstania impulsu nerwowego sygnalizującego wrażenie
wzrokowe. Jednak gęstośd fotoreceptorów w różnych częściach siatkówki nie jest jednakowa
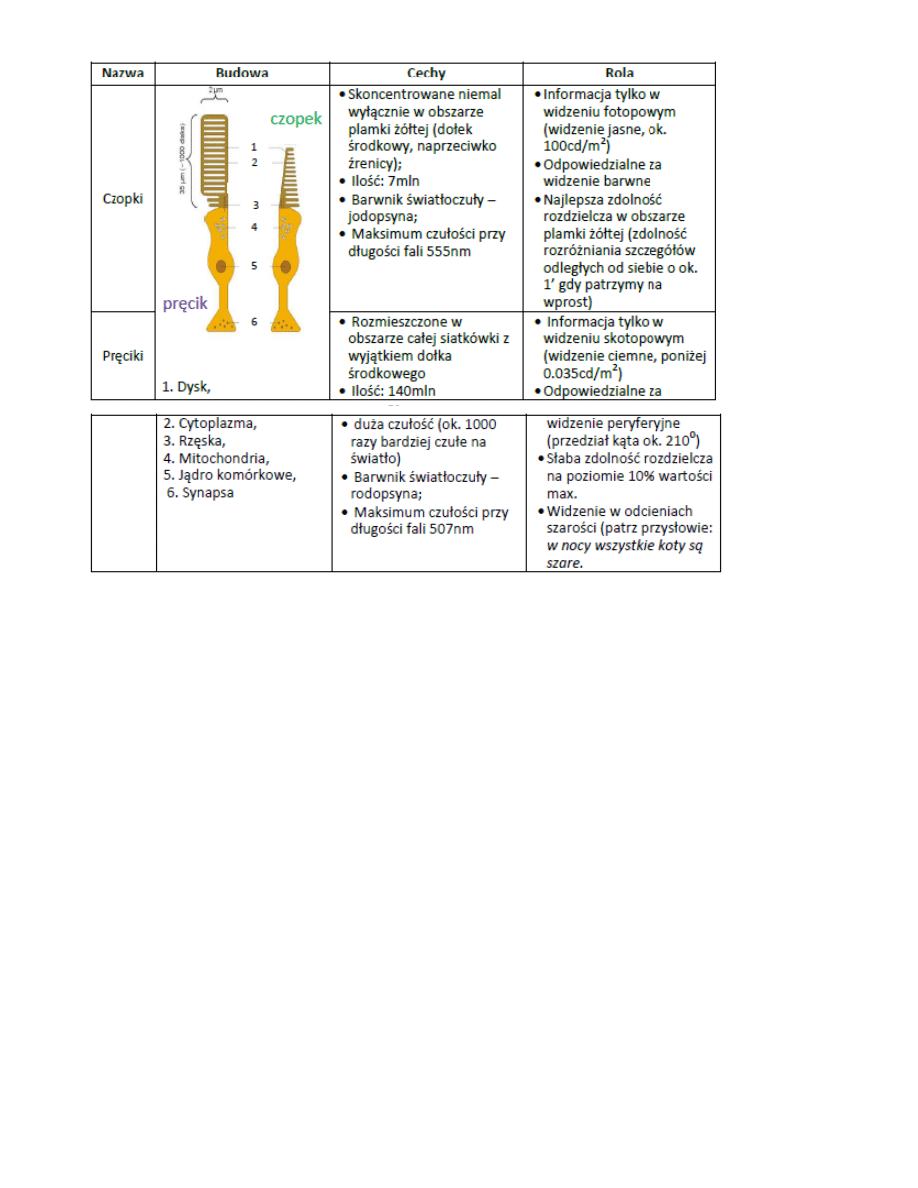
10. Adaptacja
Wspomniane już zjawisko adaptacji polega na dostosowaniu czułości oka do warunków oświetlenia, tak aby
możliwa była percepcja informacji wizualnej w możliwie szerokim przedziale luminancji. Mechanizm adaptacji
szerokości źrenicy zachodzi w ciągu kilku dziesiątych sekundy i zmieniając średnicę z 8mm do 2m m powoduje
16-krotne zmniejszenie strumienia światła padającego na siatkówkę. Drugim procesem jest adaptacja
fotochemiczna (bardziej powolna) polegająca na zmianie składu chemicznego pigmentów światłoczułych, a
następnie ich regeneracji, zachodzącej już w ciemności. Przy drastycznym spadku luminancji np. po zgaszeniu
światła w pokoju czyli z wartości ok. 100cd/m
2
do 0.001cd/m
2
(spadek o 5 rzędów wielkości) regeneracja
pigmentu zachodzi w pewnej skali czasowej. Jak pokazuje Rys.3.24, czopki przystosowują swó j próg czułości do
niższego poziomu (ok. 1cd/m
2
) w przedziale do 10 minut. Poniżej tego poziomu czopki przestają byd aktywne i
cała informacja zbierana jest przez pręciki, którym na całkowitą regenerację rodopsyny i odzyskanie
maksymalnej wrażliwości potrzeba przynajmniej 30 minut (do kilku godzin).
11. Rodopsyna
Rodopsyna – pigment światłoczuły obecny w pręcikach, pod wpływem światła rozkłada się na białko opsynę i
trans-retinal. W ciemności natomiast następuje ich ponowna synteza z udziałem witaminy A. Przy d użym
natężeniu oświetlenia procesy rozpadu przeważają nad procesami syntezy i ze wzrostem ilości światła pręciki
tracą swoją czułośd
Wyszukiwarka
Podobne podstrony:
Opracowanie wykładów biofyzka 1 MC OMEN
Opracowanie wykładów biofyzka 2 MC OMEN
Opracowanie pytań RÓŻNE MC OMEN
Opracowanie wykladow MC OMEN
Opracowanie wykladow MC OMEN
Opracowanie Sciaga MC OMEN
Opracowanie pytań MC OMEN 2
Opracowanie pytań MC KULA MC OMEN 2
Opracowanie pytań MC OMEN 3
Opracowanie projektu MC OMEN
Opracowanie pytań MC OMEN
Opracowanie pytań MC OMEN
Opracowanie do kolokwium part 2 MC OMEN
Opracowanie MC OMEN
Opracowanie Sciaga MC OMEN
Opracowanie pytań MC OMEN 2
więcej podobnych podstron