Medycyna Wet. 2007, 63 (3)
270
Artyku³ przegl¹dowy
Review
Wirus influenzy wywo³uj¹cy ptasi¹ grypê jest te¿
czynnikiem etiologicznym grypy cz³owieka, a powo-
duj¹cy g¹bczast¹ encefalopatiê byd³a (BSE) prion rów-
nie¿ wariantu choroby Creutzfeldta-Jakoba (vCJD).
Zoonotyczny charakter tych patogenów stanowi uza-
sadnienie do przedstawienia w³aciwej oceny stopnia
zagro¿enia zdrowia cz³owieka ze strony rezerwuaru
zwierzêcego, zw³aszcza wobec ró¿nych, upowszech-
nianych na ten temat pogl¹dów. Kolejnym wskazaniem
wyboru tematu jest aktualnoæ wymienionych chorób
w skali globalnej, w tym w Polsce. Dodaæ nale¿y, i¿
u zwierz¹t s¹ one przyczyn¹ olbrzymich strat, tak bez-
porednich, jak te¿ porednich. Równoczenie szereg
niejasnoci dotycz¹cych patogenezy, profilaktyki
i zwalczania, wymaga dalszych badañ.
Grypa ptasia jako zoonoza
W wieku XX wyst¹pi³y u ludzi, obok corocznych
sezonowych zachorowañ, trzy pandemie grypy odzwie-
rzêcej (13, 18). Pierwsza, w latach 1918-1919, wywo³a-
na przez podtyp H1N1 wirusa influenzy typu A, okre-
lana jako hiszpanka, spowodowa³a w skali wiatowej
mieræ oko³o 50 mln osób. Druga z lat 1957-1958, któ-
rej czynnikiem etiologicznym by³ podtyp H2N2, sta³a
siê przyczyn¹ zejæ miertelnych oko³o 1 mln ludzi,
a chorobê nazwano gryp¹ azjatyck¹. W wyniku trze-
ciej pandemii, która wyst¹pi³a w latach 1968-1969,
zmar³o oko³o 700 tys. ludzi. Wywo³a³ j¹ podtyp H3N2.
Okrelona zosta³a jako grypa Hong Kong. Wymienio-
ne podtypy wirusa influenzy wywodzi³y siê od ptaków,
nabywaj¹c w wyniku dokonuj¹cej siê zmiennoci, któ-
ra jest omówiona w dalszym tekcie, w³aciwoci cho-
robotwórczych dla cz³owieka.
Obecna pandemia grypy ptasiej rozpoczê³a siê
w po³udniowo-wschodniej Azji w grudniu 2003 r.
Z uwagi na wysokiego stopnia chorobotwórczoæ dla
ptaków domowych okrela siê j¹ jako wysoce pato-
genn¹ grypê ptasi¹ (highly pathogenic avian influen-
za), w przeciwieñstwie do niskopatogennej grypy pta-
siej, wywo³anej przez podtypy o mniejszej chorobo-
twórczoci dla drobiu (9). Wywo³ana przez podtyp
H5N1 u ptaków dzikich i domowych, zw³aszcza u kur,
Znaczenie ptasiej grypy i g¹bczastej encefalopatii
byd³a w wystêpowaniu zachorowañ u ludzi
MARIAN TRUSZCZYÑSKI, EL¯BIETA SAMOREK-SALAMONOWICZ, MIROS£AW P. POLAK
Pañstwowy Instytut Weterynaryjny Pañstwowy Instytut Badawczy, Al. Partyzantów 57, 24-100 Pu³awy
Truszczyñski M., Samorek-Salamonowicz E., Polak M. P.
Significance of avian influenza and bovine spongiform encephalopathy in human diseases
Summary
There is a wide range of differing opinions relating to assessing the risk of zoonotic pathogens which cause
avian influenza (AI) and bovine spongiform encephalopathy (BSE), and, in the case of humans, variants of
Creutzfeldt-Jakob disease (vCJD) and influenza. The aim of the study was to analyze the most essential
publications in this field. It compared the number of cases of disease and death in animals and humans and
presented the variety of mechanisms within influenza viruses as well as the consequences of their adaptations
in relation to both birds and humans. The study concluded that subtype H5N1 of the influenza virus was the
cause of the pandemic influenza in domestic and wild birds which started in 2003, is still continuing, and has
so far caused the death of over 200 million domestic birds and an undefined number of wild birds, but that it
is not a danger to humans to such a degree as subtypes H1N1, H2N2 and H3N2. These subtypes caused the
death of about 50 million, 1 million, and over 700 000 humans during respective outbreaks in the 20
th
century.
The present situation, related to the H5N1 strain has, on the other hand, only caused 129 deaths so far with the
most recent cases of mortality occurring in Asia, Turkey and Egypt. As far as BSE is concerned, cases of
transmitting the prion which causes vCJD from cattle to man have been much fewer, with the highest
numbers in the past 20 years occurring in the United Kingdom about 160 human fatalities, France 15 cases
and a few other countries with smaller numbers of fatalities. No cases of vCJD have been encountered to date
in some countries where BSE has been diagnosed such as Switzerland, Germany and Poland. Even in the U.K.
however, where in 2000 the number of fatalities of vCJD were 28, this number decreased in 2005 to 5 cases,
indicating the disappearance of this zoonosis.
Keywords: Avian influenza, bovine spongiform encephalopathy, human health
Medycyna Wet. 2007, 63 (3)
271
wysoce patogenna influenza, najpierw wyst¹pi³a i roz-
przestrzeni³a siê na obszarze Azji Po³udniowo-
-Wschodniej, a nastêpnie przenios³a siê do Chin,
Indii, Mongolii, wschodniej czêci Federacji Rosyj-
skiej, Kazachstanu, Azerbejd¿anu, Gruzji, Turcji,
Rosji europejskiej, Ukrainy, Rumunii, Bu³garii, Chor-
wacji, Wêgier, Niemiec, Republiki Czech, S³owacji,
Polski, Austrii, Francji, W³och, Zjednoczonego
Królestwa, Szwajcarii, Grecji, Boni i Hercegowiny,
Serbii i Czarnogóry, Izraela, Egiptu, Nigru, Nigerii,
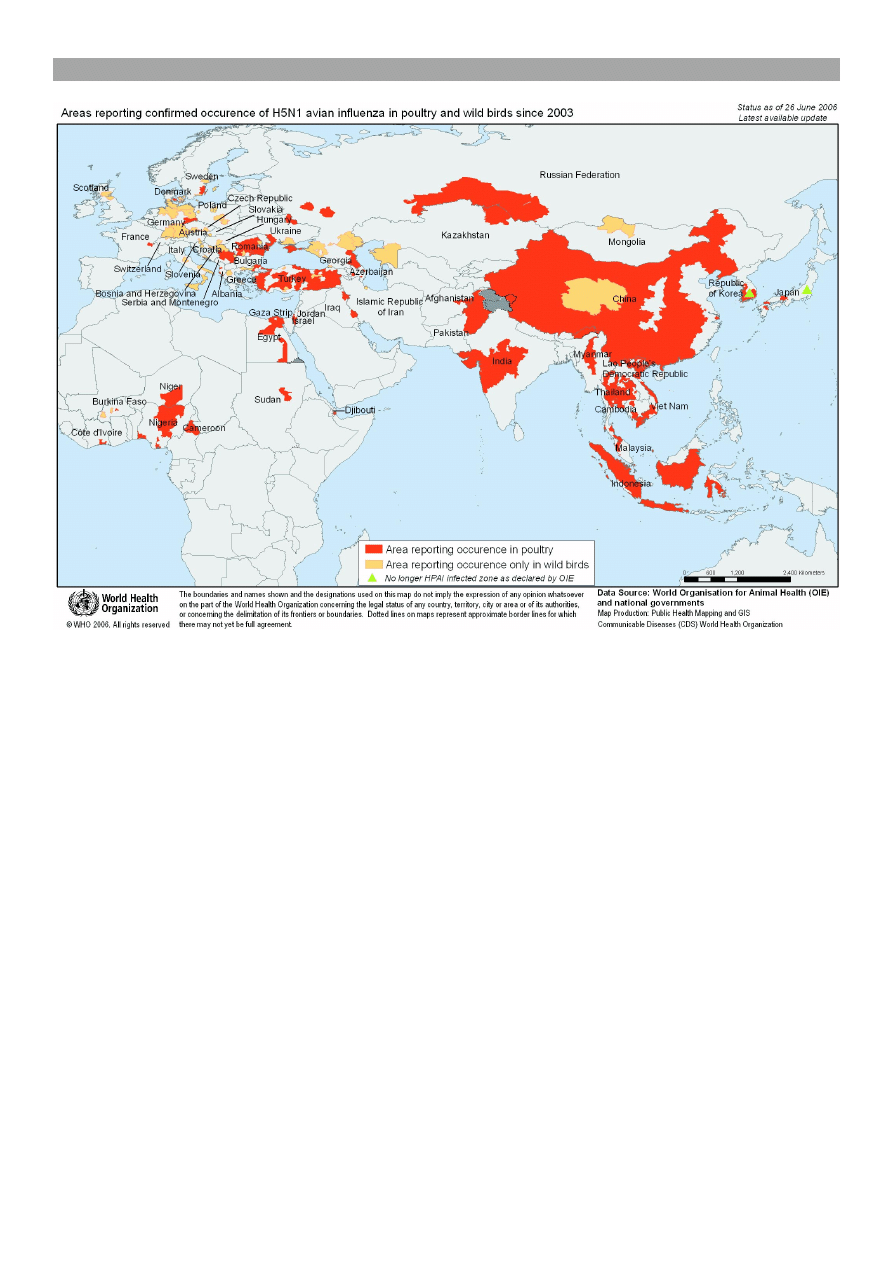
Kamerunu (31). Dane te przestawia ryc. 1. Nowe przy-
padki zachorowañ podawane s¹ na bie¿¹co drog¹ in-
ternetow¹. Znajduj¹ siê te¿ w sprawozdaniu o sytuacji
epizootiologicznej na wiecie w latach 2005-2006 (34).
Dotychczas pad³o lub zabito z koniecznoci ponad 200
mln sztuk drobiu domowego. Do tego nale¿y dodaæ
pad³e ptaki nieudomowione, których liczba nie jest do-
k³adnie okrelona. Ocenia siê (34), ¿e nigdy przedtem
¿adna choroba zakana zwierz¹t, w tym ptaków, nie
rozprzestrzenia³a siê tak szybko, na tak olbrzymi ob-
szar obejmuj¹cy ponad 50 pañstw, jak obecna ptasia
grypa, wywo³ana przez podtyp H5N1. W Azji, Turcji,
Afryce i w znacznie mniejszym zasiêgu w niektórych
krajach europejskich wyst¹pi³a u ptaków dzikich i u
domowych, zw³aszcza kur i indyków. Natomiast w po-
zosta³ych krajach europejskich, w tym w Polsce, wy-
³¹cznie u ptaków dzikich (ryc. 1).
Wywo³uj¹cy pandemie grypy u ptaków wirus H5N1
jest, jak dotychczas, w znacznie mniejszym stopniu
gronym dla cz³owieka ni¿ dla ptaków. Spowodowa³
zachorowania ponad 207 osób, z zejciem miertel-
nym ponad 129. W porównaniu do skutków wymie-
nionych pandemii grypy ptasiej, a w³aciwie odptasiej
grypy cz³owieka wieku XX, jest to liczba bardzo nis-
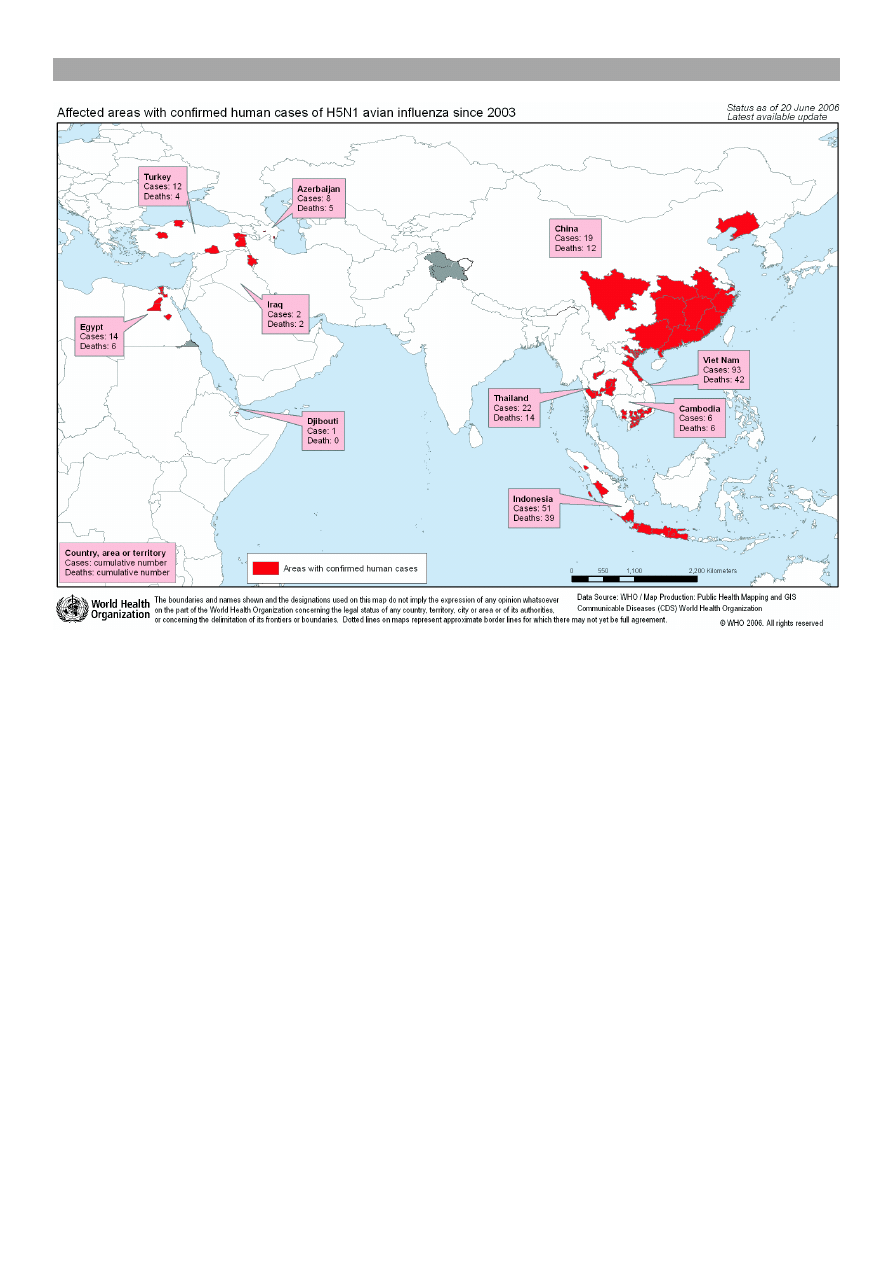
ka. Jak przedstawia ryc. 2, najwiêcej przypadków
miertelnych zanotowano w Wietnamie (42 osoby),
nastêpnie w Indonezji (38), Tajlandii (14), Chinach
(12). Po kilka osób zmar³o w Kambod¿y, Iraku, Azer-
bejd¿anie, Turcji i Egipcie (32). W Europie nie stwier-
dzono ani jednego przypadku zejcia miertelnego
u cz³owieka, wywo³anego przez podtyp H5N1 wirusa
influenzy (32). G³ówn¹ przyczyn¹ ró¿nicy zakanoci
dla ludzi miêdzy podtypami, które wywo³a³y pande-
mie grypy w wieku XX, a podtypem H5N1 jest jego
dotychczasowa niemo¿noæ przenoszenia siê z cz³o-
wieka na cz³owieka, a wy³¹cznie z zaka¿onego nim
ptaka na cz³owieka (15, 20, 23, 25).
Istnieje 16 typów hemaglutyniny (HA1-HA16) i 9
typów neuraminidazy (NA1-NA9), a poszczególne
podtypy mog¹ posiadaæ wszystkie mo¿liwe kombina-
cje typów HA i NA (26). Hemaglutynina jest to g³ów-
ne bia³ko powierzchniowe odpowiadaj¹ce za wi¹za-
nie siê z receptorami komórkowymi zaka¿onego gos-
podarza. Aminokwasy w pozycji 226 i 228 okrelaj¹
Ryc. 1. Obszary wystêpowania w latach 2003-2006 pandemii grypy ptasiej u drobiu i ptaków dzikich, wywo³anej przez pod-
typ H5N1 (31)
Medycyna Wet. 2007, 63 (3)
272
specyficznoæ receptorow¹. Hemaglutyniny z Leu 226
i Ser 228, charakterystyczne dla szczepów ludzkich,
rozpoznaj¹ struktury SA-a-2,6-Gal receptorów ludz-
kich, natomiast HA z Gln-226 i Gly-228 s¹ typowe
dla szczepów ptasich i rozpoznaj¹ SA-a-2,3-Gal. Ta
ró¿nica pozwala na zrozumienie, dlaczego replikacja
ptasich szczepów grypy u cz³owieka jest ograniczona
i dlaczego transmisja wirusa H5N1 od cz³owieka do
cz³owieka jest trudna (15). Wykazano bowiem, ¿e
obecnie kr¹¿¹ce szczepy ptasiej grypy, które wywo³a-
³y u ludzi ciê¿kie, miertelne zapalenie p³uc, zaatako-
wa³y i replikowa³y siê w komórkach dolnych partii dróg
oddechowych, których receptory maj¹ struktury mem-
branowe SA-a-2,3-Gal. Jest to zgodne z obrazem kli-
nicznym pacjentów, gdy¿ u nich obserwowano ciê¿k¹
infekcjê dolnych dróg oddechowych, czyli pêcherzy-
ków p³ucnych i uchodz¹cych do nich oskrzelików,
z szybk¹ progresj¹ do objêcia stanem zapalnym ca³e-
go mi¹¿szu p³ucnego. Ponadto stwierdzono, ¿e wirus
H5N1 wci¹¿ mutuje, gdy¿ aminokwasy zlokalizowa-
ne blisko miejsca wi¹zania receptorów ulegaj¹ zmia-
nom. Zmiany tego rodzaju wykryto w szczepie izolo-
wanym od zmar³ego Wietnamczyka po zaka¿eniu
wirusem H5N1 (23). Wykazano szereg podobieñstw
tego szczepu ze zrekonstruowanym szczepem wirusa
H1N1, który wywo³a³ hiszpankê (15, 23). Zauwa-
¿ono te¿ zmiany w wirusowej polimerazie PB2. W pta-
sich szczepach influenzy w pozycji 627 znajduje siê
zazwyczaj kwas glutaminowy, podczas gdy w ludz-
kich szczepach jest lizyna. Jednak¿e szczepy H5N1
izolowane od pacjentów zmar³ych po zaka¿eniu wiru-
sem H5N1 posiada³y lizynê w pozycji 627. Równie¿
lizyna w tej pozycji znajduje siê w szczepach izolo-
wanych w drugiej po³owie 2005 r. od dzikich ptaków
wodnych z jeziora Qinghai w Chinach (15).
Nie mo¿na wykluczyæ, ¿e w wyniku zmiennoci
podtyp H5N1 uzyska zdolnoæ ³¹czenia siê z recepto-
rami SA-a-2,6-Gal, które s¹ zlokalizowane w górnych
drogach oddechowych cz³owieka. Wtedy zaistnia³aby
mo¿liwoæ transmisji drog¹ kropelkow¹ infekcji z cz³o-
wieka na cz³owieka, co mog³oby groziæ rozwojem
pandemii analogicznych do tych, które wyst¹pi³y w XX
wieku (23).
Generalnie, wirusy grypy cechuj¹ siê du¿¹ tenden-
cj¹ do zmiennoci (26). Znane s¹ dwa rodzaje zmien-
noci: reasortacja genowa, nazywana skokiem anty-
genowym (antigenic shift) oraz przesuniêcie antyge-
nowe (antigenic drift) (11, 19, 24, 26).
Reasortacja czyli skok antygenowy mo¿e wyst¹piæ,
gdy komórka gospodarza zostanie jednoczenie zaka-
¿ona przez dwa ró¿ni¹ce siê od siebie szczepy wirusa
grypy. Dochodzi wówczas, miêdzy innymi, do wymia-
ny poszczególnych segmentów RNA, co jest przyczy-
n¹ powa¿nych zmian antygenowych na powierzchni
Ryc. 2. Obszary wystêpowania zachorowañ u ludzi, wywo³anych przez podtyp H5N1 (32)

Medycyna Wet. 2007, 63 (3)
273
wiriona. Skok antygenowy mo¿e dotyczyæ ka¿dego
z omiu segmentów genomu wirusa. Potencjalnie mo¿e
wyst¹piæ 256 ró¿nych genetycznie szczepów potom-
nych. Wymiana segmentów genomu jest równie¿ mo¿-
liwa pomiêdzy szczepami wirusa pochodz¹cymi od
ró¿nych gatunków zwierz¹t (19). Równoczesne zaka-
¿enie komórki szczepem ludzkim i ptasim mo¿e do-
prowadziæ do powstania nowych podtypów, ró¿nych
od ju¿ istniej¹cych. Powstaj¹cy wirus posiada czêæ
segmentów pochodz¹cych z genomu od jednego szcze-
pu, a czêæ od drugiego szczepu i mo¿e staæ siê podty-
pem daj¹cym pocz¹tek epidemii, a nawet pandemii (11,
19). Przyk³adem tego s¹ podtypy H2N2 oraz H3N2.
Wirus H2N2 uzyska³ na drodze reasortacji trzy geny
PB1, HA i NA od wirusa wystêpuj¹cego u dzikich
kaczek, natomiast wirus H3N2 naby³ poprzez rearota-
cjê dwa geny PB1 i HA równie¿ od wirusa wystêpuj¹-
cego u kaczek (21, 23).
Zmiennoæ antygenowa wirusów grypy mo¿e odby-
waæ siê równie¿ na drodze przesuniêcia antygenowe-
go (antigenic drift). Dotyczy ono drobnych zmian
w segmentach koduj¹cych antygeny powierzchniowe.
Polimeraza RNA zale¿na od RNA jest enzymem do-
konuj¹cym licznych pomy³ek, polegaj¹cych na wsta-
wianiu b³êdnych zasad podczas syntezy potomnych
³añcuchów RNA. Polimerazy RNA nie maj¹ tzw. w³as-
noci korektorskich i nie potrafi¹ tych zasad usun¹æ.
W wyniku tego powstaj¹ mutacje punktowe prowa-
dz¹ce do pojawiania siê w nastêpstwie selekcji nowych
wariantów antygenowych uprzednio wystêpuj¹cych
podtypów. Przesuniêcie antygenowe jest procesem sta-
³ym i ci¹g³ym, natomiast skok antygenowy pojawia siê
nagle (19, 22).
W wietle przedstawionych danych mimo ¿e obec-
ne szczepy podtypu H5N1 ci¹gle, poza wspomnia-
nymi wyj¹tkami, nie wykazuj¹ zdolnoci transmisji
z cz³owieka na cz³owieka, nie mo¿na wykluczyæ, ¿e
to mo¿e nast¹piæ. Przemawiaæ mog¹ za tym pandemie
grypy ludzkiej od ptasiej z XX wieku. Z drugiej stro-
ny tak staæ siê nie musi, a w zapobieganiu tej ewentu-
alnoci du¿a rola przypada weterynarii. G³ównymi ele-
mentami zapobiegania i zwalczania grypy ptasiej,
a porednio zachorowañ na tê chorobê ludzi s¹ kli-
niczne i laboratoryjne przegl¹dy stad drobiu i badania
pad³ych ptaków nieudomowionych, oparty na interne-
cie system informacji i ostrzegania, zorganizowany
w skali globalnej przez wiatow¹ Organizacjê Zdro-
wia Zwierz¹t (OIE), bioasekuracja ferm i stad przy-
zagrodowych, wybijanie z koniecznoci (czyli wszyst-
kich ptaków) w ognisku choroby oraz prewencyjne
(czyli ptaków zdrowych) znajduj¹cych siê w s¹siedz-
twie ogniska choroby (9, 29, 34). W nawi¹zaniu do
tego nale¿y dodaæ, ¿e najskuteczniejsz¹ obron¹ przed
pojawianiem siê chorobotwórczych dla cz³owieka
mutantów wirusa influenzy jest szybkie i sprawne ogra-
niczanie oraz likwidacja zwierzêcego rezerwuaru wi-
rusa influenzy, w³anie poprzez wybijanie ptaków za-
ka¿onych (9, 24). Rola immunoprofilaktyki zosta³a
omówiona w nastêpuj¹cych publikacjach (7, 29). Jej
stron¹ ujemn¹ jest utrzymywanie siê w populacjach
szczepionego drobiu wirusa patogennego, co daje szan-
se na pojawianie siê nowych odmian chorobotwór-
czych nie tylko dla ptaków, ale równie¿ dla cz³owie-
ka. Dodatkowo, ptaki immunizowane, mimo infekcji,
nie wykazuj¹ objawów klinicznych, co utrudnia wy-
krycie obecnoci wirusa w stadzie.
Zoonotyczny charakter BSE
Mimo ¿e BSE przenosi siê z osobnika na osobnika
(z krowy na krowê lub z krowy na cz³owieka) za po-
rednictwem nie do koñca poznanego, pasa¿owalne-
go (transmissible) czynnika etiologicznego, w zwi¹z-
ku z czym mo¿e byæ zaliczona do chorób zakanych,
to jednak czynnik ten ró¿ni siê zasadniczo swymi w³a-
ciwociami i mechanizmem wywo³ywania choroby od
wirusa grypy, wszystkich innych wirusów oraz bakte-
rii, czyli drobnoustrojów stanowi¹cych typow¹ przy-
czynê infekcji i chorób zakanych. BSE wywo³uje nie-
konwencjonalny czynnik, którego g³ównym sk³adni-
kiem jest oporne na proteolizê zakane bia³ko, okre-
lane jako prion (16, 27, 35). Czynnik etiologiczny BSE
nie zawiera kwasów nukleinowych i nie podlega re-
plikacji, natomiast szerzy siê w tkance nerwowej w na-
stêpstwie kontaktu z form¹ fizjologiczn¹ bia³ka prio-
nowego, wywo³uj¹c u niego, na zasadzie swego rodza-
ju efektu domina, zmiany jego struktury trzeciorzêdo-
wej, charakterystyczne dla prionu (1, 10). Efektem jest
degeneracja komórek nerwowych, uwidaczniaj¹ca siê
g¹bczastoci¹ w badaniu histopatologicznym tkanki
nerwowej, a klinicznie zwiêkszon¹ pobudliwoci¹ i za-
burzeniami motorycznymi oraz ataksj¹ (5, 28). Brak
reakcji zapalnej i odpowiedzi immunologicznej ze stro-
ny zaka¿onego osobnika równie¿ stanowi ró¿nicê
w stosunku do typowych chorób zakanych. Choroba
zawsze koñczy siê mierci¹ (35).
Pimiennictwo na temat BSE stwierdzonej po raz
pierwszy w Wielkiej Brytanii w 1986 r. jest bardzo
obszerne (8, 30). Sporód tych danych przedmiotem
niniejszego artyku³u s¹ jednak g³ównie prace wskazu-
j¹ce na prze³amanie progu gatunkowego: krowa ®
cz³owiek i na wspó³zale¿noæ wystêpowania wariantu
choroby Creutzfeldta-Jakoba (vCJD) jako efektu prze-
niesienia chorobotwórczych prionów z byd³a na cz³o-
wieka (6, 12). Tabela 1 prezentuje dane epidemiolo-
giczne dotycz¹ce BSE i vCJD wg danych OIE z 2005 r.
i 2006 r. (33, 34). Wyp³ywaj¹ z niej, w kontekcie stop-
nia zagro¿enia zdrowia cz³owieka ze strony konsumo-
wanych produktów pochodzenia bydlêcego, zw³asz-
cza zawieraj¹cych tkankê mózgow¹, nastêpuj¹ce
stwierdzenia. W Wielkiej Brytanii, gdzie wykazano
najwiêksz¹ liczbê zachorowañ na BSE, w ci¹gu 20
minionych lat (182 620 sztuk byd³a), na vCJD zmar³o
160 osób. W Portugalii, gdzie liczba przypadków BSE
na 1 mln krów w wieku powy¿ej 24. miesi¹ca ¿ycia
by³a nawet wy¿sza ni¿ w Wielkiej Brytanii, na vCJD
zmar³a dotychczas 1 osoba. W Niemczech z 398 przy-
Medycyna Wet. 2007, 63 (3)
274
padkami BSE i w Szwajcarii z 463 przypadkami BSE
nie stwierdzono ani jednego zachorowania na vCJD,
a s¹ to przecie¿ kraje o wysokim poziomie diagnosty-
ki chorób zakanych cz³owieka i zwierz¹t. Nie nale¿y
siê zatem spodziewaæ b³êdnych wyników. O spadku
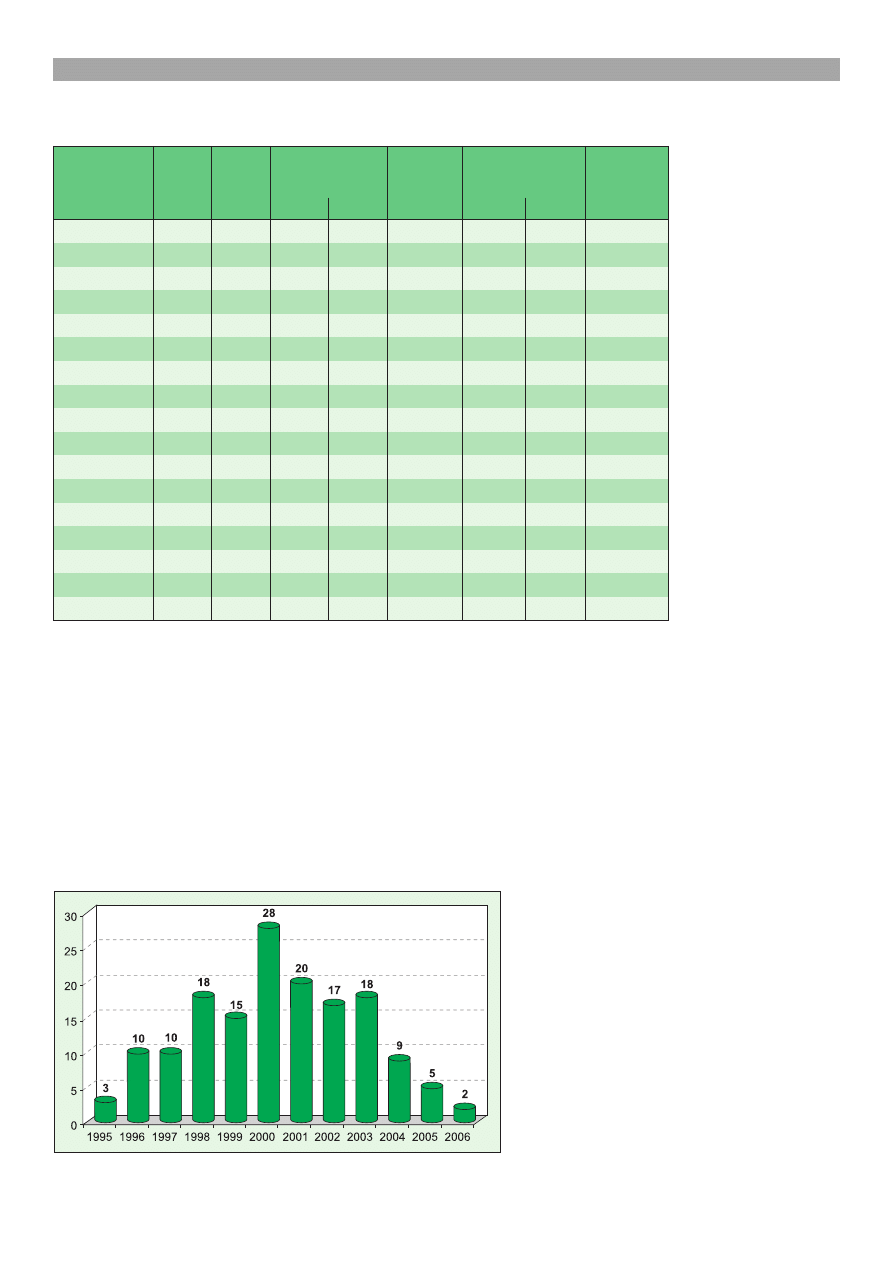
zachorowañ na vCJD w Wielkiej Brytanii, jak wynika
z ryc. 3, wiadczy stwierdzenie tam w 2000 r. 28
przypadków, a w 2005 r. tylko 5 (3). Wykazano
te¿ tam przesuniêcie redniej wieku wystêpowania
BSE u byd³a z 86 miesiêcy w 2001 r. do 108 miesiêcy
w 2004 r. Zmniejsza
siê zatem rezerwuar
chorobotwórczych
dla cz³owieka prio-
nów, poniewa¿ star-
sze krowy s¹ elimino-
wane ze stad dostar-
czaj¹cych produkty
¿ywnociowe dla
cz³owieka. Dodatko-
wo na 41 mln sztuk
byd³a zbadanych la-
boratoryjnie w kie-
runku BSE w latach
2001-2005 jedynie
u 4 krów w wieku po-
ni¿ej 35 miesiêcy,
czyli niespe³na 3 lat,
uzyskano wynik do-
datni. Równie¿ ten
rezultat wskazuje na
stopniowo zmniej-
szaj¹cy siê rezerwuar
odzwierzêcych prio-
nów chorobotwór-
czych dla cz³owieka
(2). Kszta³tuj¹ca siê
aktualnie coraz bar-
dziej korzystna sy-
tuacja w odniesieniu
do BSE jest efektem
wprowadzonego w latach 90. zakazu podawania byd-
³u jako dodatków paszowych m¹czek miêsno-kost-
nych, zw³aszcza od prze¿uwaczy, mog¹cych zawieraæ
chorobotwórcze priony. Jest ona te¿ wynikiem w³a-
ciwie opracowanych i realizowanych programów mo-
nitorowania i zwalczania BSE (2).
Wy³ania siê zatem podstawa do z³agodzenia obec-
nych restrykcji, ustanowionych w celu zapobiegania
vCJD u ludzi. W konsekwencji podjêto dzia³ania zmie-
rzaj¹ce w tym kierunku. Nie wybija siê, przeciwnie
ni¿ poprzednio, wszystkich krów w stadzie,
w którym rozpoznano przypadek BSE, a wy-
³¹cznie tzw. kohortê, czyli krowy w przedzia-
le wiekowym ± 1 rok w stosunku do krowy,
która zachorowa³a. Chodzi bowiem o osob-
niki, które ewentualnie równoczenie z ni¹
spo¿ywa³y tê sam¹ karmê, zawieraj¹c¹ prio-
ny (17). Przesuniêto te¿ wiek poubojowego
badania tkanki mózgowej krów ubijanych
w rzeni z 24 miesiêcy do 30 miesiêcy, co
istotnie zmniejsza koszt tego rodzaju drogich
badañ (4). Istniej¹ ze strony g³ównych leka-
rzy weterynarii szeregu pañstw naciski, by
granicê tê przesun¹æ do 36 miesiêcy. Ze
wzglêdu jednak na zwiêkszone tym samym
ryzyko zagro¿enia zdrowia cz³owieka decy-
zja ta, na razie, nie zosta³a podjêta (14).
j
a
r
K
S
E
S
B
S
D
J
C
v
E
S
B
w
ó
r
k
a
b
z
c
i
L
.
s
e
i
m
4
2
>
a
³
d
y
b
u
E
S
B
a
b
z
c
i
L
a
n
.
s
e
i
m
4
2
>
w
ó
k
i
n
b
o
s
o
0
0
0
0
0
0
1
k
e
d
a
p
S
¯
t
s
o
r
z
W
y
c
i
n
¿
ó
r
k
a
r
B
.r
4
0
0
2
.r
5
0
0
2
.r
4
0
0
2
.r
5
0
0
2
a
i
g
l
e
B
2
3
1
0
1
1
1
1
2
1
4
4
5
9
3
1
1
2
8
8
,
7
1
0
3
4
,
1
¯
a
d
a
n
a
K
2
1
5
1
2
1
1
1
1
0
0
0
0
0
7
6
1
9
4
1
,
0
1
9
4
1
,
0
a
k
s
e
z
C
a
k
il
b
u
p
e
R
1
4
2
0
2
1
7
1
8
2
0
0
0
8
7
6
4
2
3
,
0
1
0
9
7
,
1
1
a
i
n
a
D
1
5
1
0
2
1
1
1
1
3
1
6
5
1
7
7
1
6
9
2
,
1
1
6
9
2
,
1
a
j
c
n
a
r
F
8
7
9
5
1
1
1
4
5
1
3
0
0
0
0
0
4
1
1
1
1
6
3
7
,
4
1
9
1
7
,
2
¯
y
c
m
e
i
N
8
9
3
0
1
5
6
2
3
0
0
0
5
5
9
5
0
2
9
,
0
1
1
0
7
3
,
5
¯
a
i
d
n
a
lr
I
3
6
5
1
1
4
6
2
1
9
6
5
9
0
8
0
9
2
7
2
3
,
3
4
0
2
7
,
3
2
¯
y
h
c
o
³
W
4
3
1
1
2
1
7
1
8
0
0
0
0
8
9
2
1
8
4
3
,
2
1
4
8
6
,
2
a
i
n
o
p
a
J
1
3
2
1
2
1
5
1
7
0
0
8
6
0
0
2
1
1
9
4
,
2
1
8
8
4
,
3
a
i
d
n
a
l
o
H
1
1
8
1
2
1
6
1
3
0
0
0
5
6
7
1
1
9
9
3
,
3
1
9
9
6
,
1
¯
a
k
s
l
o
P
1
6
4
0
1
1
1
0
2
4
9
5
3
7
0
3
1
8
7
5
,
3
1
7
0
5
,
6
a
il
a
g
u
tr
o
P
6
0
0
1
1
1
1
1
9
1
5
3
2
2
4
9
6
9
0
7
8
,
3
9
8
0
6
,
2
5
¯
a
j
c
a
w
o
³
S
1
0
2
0
2
1
7
1
2
2
1
4
1
4
8
2
5
3
6
,
4
2
1
8
3
0
,
7
¯
a
i
n
e
w
o
³
S
1
1
6
0
2
1
1
1
1
3
0
8
0
8
1
2
1
5
8
5
,
4
1
5
8
5
,
4
a
i
n
a
p
z
s
i
H
9
3
6
1
7
3
1
8
9
6
3
7
7
1
5
3
5
4
9
,
8
3
8
5
8
,
7
2
¯
a
ir
a
c
j
a
w
z
S
3
6
4
0
1
1
3
1
3
3
0
0
0
0
0
8
1
0
5
7
,
3
1
0
5
7
,
3
a
i
n
a
t
y
r
B
a
k
l
e
i
W
0
2
6
2
8
1
0
6
1
1
1
8
3
3
4
2
2
1
5
9
4
5
8
9
4
6
9
7
,
7
6
0
3
9
,
4
4
¯
Tab. 1. Dane epidemiologiczne dotycz¹ce g¹bczastej encefalopatii byd³a (BSE) i wariantu choroby
Creutzfeldta-Jakoba (vCJD) (33, 34)
Objanienia: S BSE = liczba przypadków g¹bczastej encefalopatii byd³a; S vCJD = liczba przypadków
wariantu choroby Creutzfeldta-Jakoba; ¯ = spadek; = wzrost; = brak ró¿nic
Ryc. 3. Liczba przypadków vCJD w Wielkiej Brytanii (dane z 3 kwiet-
nia 2006 r.) (3)

Medycyna Wet. 2007, 63 (3)
275
Podsumowuj¹ca ocena
zagro¿enia zdrowia cz³owieka
W wietle przedstawionych danych istnieje stale
du¿e niebezpieczeñstwo ze strony odzwierzêcych,
a zw³aszcza odptasich wirusów influenzy, pojawiania
siê odmian wysoce chorobotwórczych dla cz³owieka.
Nie wydaje siê jednak prawdopodobne, by powtórzy-
³y siê pandemie, które mia³y miejsce w wieku XX.
Dotyczy to zw³aszcza krajów rozwiniêtych, dysponu-
j¹cych sprawnie dzia³aj¹c¹ s³u¿b¹ weterynaryjn¹ w te-
renie oraz kompetentn¹ diagnostyk¹ laboratoryjn¹, jak
równie¿ funkcjonuj¹cym w skali globalnej interneto-
wym systemem ostrzegania przed zbli¿aj¹cym siê za-
gro¿eniem.
Ustosunkowuj¹c siê do wspó³zale¿noci BSE
i vCJD, mo¿na wnioskowaæ, ¿e zagro¿enie to, dziêki
aktualnym kwalifikacjom przedstawicieli nauki i prak-
tyki weterynaryjnej maleje, na co dowody zosta³y przy-
toczone.
Pimiennictwo
1.Aguzzi A., Heppner F. L.: Pathogenesis of prion diseases: a progress report.
Cell Death Differ. 2000, 7, 889-902.
2.Anon.: Report on the monitoring and testing of ruminants for the presence of
Transmissible Spongiform Encephalopathy (TSE) in the EU in 2004. Euro-
pean Communities 2005.
3.Anon.: Monthly Creutzfeldt-Jakob disease statistics, The Department of
Health of England, April 2006.
4.Anon.: Rozporz¹dzenie MRiRW z dnia 1 lutego 2002 r., Dz. U. nr 10, poz.
103, 2002.
5.Braun U., Schicker E., Hornlimann B.: Diagnostic reliability of clinical
signs in cows with suspected bovine spongiform encephalopathy. Vet. Rec.
1998, 143, 101-105.
6.Bruce M. E., Will R. G., Ironside J. W., McConnell I., Drummond D., Suttie A.,
McCardle L., Chree A., Hope J., Birkett C., Cousens S., Fraser H., Bostock C. J.:
Transmissions to mice indicate that new variant CJD is caused by the BSE
agent. Nature 1997, 389, 498-501.
7.Capua I., Marangon S.: The use of vaccination as an option for the control of
avian influenza. 71 General Session International Committee World Organi-
sation for Animal Health (OIE), Paris 18-23 May 2003, 1-10.
8.Collee J. G., Bradley R.: BSE: a decade on part 2. Lancet 1997, 349, 715-
-721.
9.Dyrektywa Rady 2005/94/WE z dnia 20 grudnia 2005 r. w sprawie wspólno-
towych rodków zwalczania grypy ptaków i uchylaj¹ca dyrektywê 92/40/
EWG. Dz. U. UE 2006, L 10, 16-65.
10.Eghiaian F.: Structuring the puzzle of prion propagation. Curr. Opin. Struct.
Biol. 2005, 15, 724-730.
11.Hayden F., Croisier A.: Transmission of avian influenza viruses to and be-
tween humans. J. Infect. Dis. 2005, 192, 1-4.
12.Hill A. F., Desbruslais M., Joiner S., Sidle K. C., Gowland I., Collinge J.,
Doey L. J., Lantos P.: The same prion strain causes vCJD and BSE. Nature
1997, 389, 448-450.
13.Kawaoka Y., Krauss S., Webster R. G.: Avian to human transmission of the
PB1 gene of influenza A virus in the 1957 and 1968 panademics. J. Virol.
1989, 63, 4603-4608.
14.Knight R.: The relationship between new variant Creutzfeldt-Jakob disease
and bovine spongiform encephalopathy. Vox Sang. 1999, 76, 203-208.
15.Kuiken T., Holmes E. C., McCauley J., Rimmelzwaan G. F., Williams C. S.,
Grenfell B.: Host species barriers to influenza virus infections. Science 2006,
312, 394-397.
16.Prusiner S. B.: Novel proteinaceous infectious particles cause scrapie Scien-
ce 1982, 216, 136-144.
17.Rabenau H. F., Cinatl J., Doerr H. W.: Prions. A challenge for science, medi-
cine and the public health system. Karger AG, Basel 2004, s. 186.
18.Samorek-Salamonowicz E., Truszczyñski M., Kozdruñ W.: Ptasia grypa
wiatowy problem epidemiologiczny. Kosmos 2005, 54, 321-330.
19.Scholtisek C.: Molecular evolution of influenza viruses. Virus Genes 1995,
11, 209-215.
20.Shinya K., Ebina M., Yamada S., Ono M., Kasai N., Kaeaoka Y.: Avian flu:
influenza virus receptors in the human airway. Nature 2006, 440, 435-436.
21.Shortridge K. F., Gao P., Guan Y., Jto T., Kawaoka Y., Markwell D., Taka-
da A., Webster R. G.: Interspecies transmission of influenza viruses: H5N1
virus and a Hong Kong SAR perspective. Vet. Microbiol. 2000, 74, 141-147.
22.Shortridge K. F.: The influenza conundrum. J. Med. Microbiol. 1997, 46,
813-815.
23.Stevens J., Blixt O., Tumpey T. M., Taubenberger J. K., Paulson J. C., Wil-
son J. A.: Structure and receptor specificity of hemagglutinin from an H5N1
influenza virus. Science 2006, 312, 404-410.
24.Swayne D. E., Halvorson D. A.: Avian Influenza, [w:] Diseases of Poultry.
Iowa State Press a Blackwell Publishing Company 2003, 135-160.
25.Van Riel D., Munster V. J., de Wit E., Rimmelzwaan G. F., Fouchier R. A. M.,
Osterhaus Ab D. M. A., Kuiken T.: H5N1 virus attachment to lower respira-
tory tract. Science 2006, 312, 399-403.
26.Webster R. G., Hulse D. J.: Microbial adaptation and change: avian influenza.
Res. Sci. tech. Off. int. Epiz. 2004, 23, 453-465.
27.Weissmann C.: The state of the prion. Nature Rev. 2004, 2, 1-11.
28.Wells G. A., Hancock R. D., Cooley W. A., Richards M. S., Higgins R. J.,
David G. P.: Bovine spongiform encephalopathy: diagnostic significance of
vacuolar changes in selected nuclei of the medulla oblongata. Vet. Rec. 1989,
125, 521-524.
29.Wijaszka T., Truszczyñski M.: Rozwa¿ania dotycz¹ce zgody na szczepienia
przeciw wysoce patogennej influenzie ptasiej. ¯ycie Wet. 2006, 81, 165-
-167.
30.Wilesmith J. W., Hoinville L. J., Ryan J. B., Sayers A. R.: Bovine spongiform
encephalopathy: aspects of the clinical picture and analyses of possible changes
1986-1990. Vet. Rec. 1992, 130, 197-201.
31.World Health Organisation of Health. Data Source: Word Organisation of
Animal Health (OIE) and national governments. Map Production Public
Health Mapping and GIS. Communicable Diseases, 26 June 2006.
32.World Health Organisation. Map Production Public Health Mapping and GIS.
Communicable Diseases, 20 June 2006.
33.World Organisation for Animal Health (OIE). 73 General Session, Paris 22-27
May 2005.
34.World Organisation for Animal Health (OIE). 74 General Session, Paris 21-26
May 2006.
35.¯mudziñski J. F., Truszczyñski M., Macio³ek H.: G¹bczaste encefalopatie ze
szczególnym uwzglêdnieniem g¹bczastej encefalopatii byd³a (Bovine Spon-
giform Encephalopathy, BSE). Wyd. PIWet, Pu³awy 1995, 4-42.
Adres autora: prof. dr hab. Marian Truszczyñski, Pañstwowy Instytut
Weterynaryjny Pañstwowy Instytut Badawczy, Al. Partyzantów 57,
24-100 Pu³awy; e-mail: mtruszcz@piwet.pulawy.pl
Wyszukiwarka
Podobne podstrony:
gąbczasta encefalopatia bydła
Gabczaste encefalopatie kotow i Nieznany
Gąbczaste encefalopatie dr n, Pomoce naukowe, studia, medycyna
Gabczaste encefalopatie kotow i Nieznany
Przesłanie ptasiej grypy dla ludzkości
Budowa, wystepowanie i znaczenie biologiczne disacharydow
Cechy funkcjonalne i ich znaczenie w hodowli bydla
GOSPODARCZE ZNACZENIE BYDŁA 1
Budowa, wystepowanie i znaczenie biologiczne disacharydow
Cechy funkcjonalne i ich znaczenie w hodowli bydla
Rzadko występujące choroby jajników u bydła
ROLA I ZNACZENIE WSKAŹNIKOWE ORGANIZMÓW WYSTĘPUJĄCYCH W OSADZIE CZYNNYM
Białka występowanie, podział, znaczenie
więcej podobnych podstron