

WĘGLOWODANY
WĘGLOWODANY
•
Węglowodany dostarczane z pokarmem w większości są
polimerami hektoz, z których najważniejszymi są:
glaktoza,
fruktoza,
glukoza
•
Węglowodany dostarczane z pokarmem w większości są
polimerami hektoz, z których najważniejszymi są:
glaktoza,
fruktoza,
glukoza
•
Większość monosacharydów występujących w ustroju należy do
izomerów D.
•
Głównym produktem trawienia węglowodanów i głównym
krążącym we krwi cukrem jest glukoza.
•
Normalne stężenie glukozy na czczo, w obwodowej krwi żylnej
wynosi 3,9 – 5,6 mmol/L (70 – 100 mg/dL), we krwi tętniczej
stężenie glukozy jest o 0,56 – 1,67 mmol/L (15 – 30 mg/dL)
wyższe.
•
Większość monosacharydów występujących w ustroju należy do
izomerów D.
•
Głównym produktem trawienia węglowodanów i głównym
krążącym we krwi cukrem jest glukoza.
•
Normalne stężenie glukozy na czczo, w obwodowej krwi żylnej
wynosi 3,9 – 5,6 mmol/L (70 – 100 mg/dL), we krwi tętniczej
stężenie glukozy jest o 0,56 – 1,67 mmol/L (15 – 30 mg/dL)
wyższe.

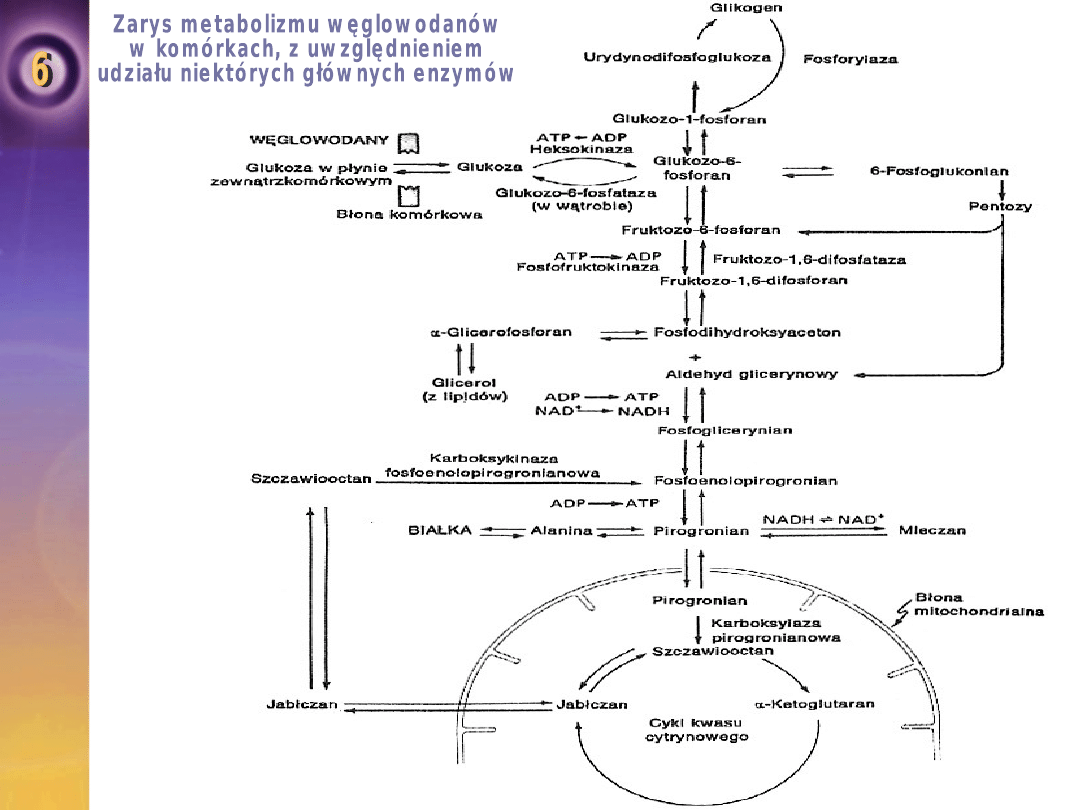
Metabolizm węglowodanów
Metabolizm węglowodanów
•
Gdy glukoza wnika do komórek, ulega fosforylacji do
glukozo – 6 – fosforanu.
Enzymem, który kształtuje tę reakcję, jest heksokinaza.
•
W wątrobie występuje dodatkowy enzym glukokinaza,
która wykazuje większa specyficzność wobec glukozy i w
odróżnieniu od heksokinazy podlega aktywującemu
działaniu insuliny.
W okresie głodzenia i w cukrzycy następuje spadek
aktywności tego enzymu.
•
Glukozo – 6 – fosforan ulega polimeryzacji do glikogenu
albo ulega przemianom katabolicznym.
•
Synteza glikogenu nazywana jest glokogenezą, a rozpad
glikogenu glikogenolizą.
Glikogen jest formą magazynowania glukozy, obecny jest w
większości tkanek ustrojowych, ale największe jego zapasy
występują w wątrobie i mięśniach szkieletowych.
•
Gdy glukoza wnika do komórek, ulega fosforylacji do
glukozo – 6 – fosforanu.
Enzymem, który kształtuje tę reakcję, jest heksokinaza.
•
W wątrobie występuje dodatkowy enzym glukokinaza,
która wykazuje większa specyficzność wobec glukozy i w
odróżnieniu od heksokinazy podlega aktywującemu
działaniu insuliny.
W okresie głodzenia i w cukrzycy następuje spadek
aktywności tego enzymu.
•
Glukozo – 6 – fosforan ulega polimeryzacji do glikogenu
albo ulega przemianom katabolicznym.
•
Synteza glikogenu nazywana jest glokogenezą, a rozpad
glikogenu glikogenolizą.
Glikogen jest formą magazynowania glukozy, obecny jest w
większości tkanek ustrojowych, ale największe jego zapasy
występują w wątrobie i mięśniach szkieletowych.

Metabolizm węglowodanów
Metabolizm węglowodanów
•
Przemiana glukozy w pirogronian lub mleczan nazywana jest
glikolizą.
•
Rozpad glukozy zachodzi poprzez rozszczepienie na triozy lub
przez procesy utleniania i dekarboksylacji do wytworzenia pentoz.
•
Przemiana glukozy poprzez triozy do uzyskania pirogronianu
nazywana jest drogą Embdena – Mayerhofa, a przemian z
pośrednim wytworzeniem 6-fosfoglukonianu i petoz jest drogą
bezpośredniego utleniania (cykl
heksozomonofosforanowy).
•
Pirognian jest przekształcony do acetylo-CoA.
•
Wzajemne przekształcanie węglowodanów, białek i tłuszczy
polega na przekształceniu uzyskanego z lipidów glicerolu w
fosforan dihydroksyacetonu.
•
Przemiana glukozy w pirogronian lub mleczan nazywana jest
glikolizą.
•
Rozpad glukozy zachodzi poprzez rozszczepienie na triozy lub
przez procesy utleniania i dekarboksylacji do wytworzenia pentoz.
•
Przemiana glukozy poprzez triozy do uzyskania pirogronianu
nazywana jest drogą Embdena – Mayerhofa, a przemian z
pośrednim wytworzeniem 6-fosfoglukonianu i petoz jest drogą
bezpośredniego utleniania (cykl
heksozomonofosforanowy).
•
Pirognian jest przekształcony do acetylo-CoA.
•
Wzajemne przekształcanie węglowodanów, białek i tłuszczy
polega na przekształceniu uzyskanego z lipidów glicerolu w
fosforan dihydroksyacetonu.

Metabolizm węglowodanów
Metabolizm węglowodanów
•
Przemiana białek polega na przekształceniu na drodze
deaminacji pewnej ilości aminokwasów ze szkieletami
węglowymi, podobnymi do produktów pośrednich cyklu
przemian Embdena-Mayrehofa i cyklu kwasu cytrynowego w te
właśnie produkty pośrednie.
W ten sposób i poprzez konwersję mleczanu do glukozy cząsteczki
nie będące glukozą mogą być przekształcone w glukozę
(glukoneogeneza).
•
Glukoza może być przekształcana w tłuszcze poprzez acetylo-
CoA, ponieważ przemiana pirogranianu do acetylo-CoA jest
nieodwracalna. w przeciwieństwie do większości reakcji glikozy,
to jednak tłuszcze nie są ta drogą przekształcane na glukozę.
Całkowita przemiana tłuszczów w węglowodany w ustroju jest
bardzo ograniczona, ponieważ poza nieistotnym ilościowo
tworzeniem węglowodanów z glicerolu nie istnieje żadna inna
droga dla takiej przemiany.
•
Przemiana białek polega na przekształceniu na drodze
deaminacji pewnej ilości aminokwasów ze szkieletami
węglowymi, podobnymi do produktów pośrednich cyklu
przemian Embdena-Mayrehofa i cyklu kwasu cytrynowego w te
właśnie produkty pośrednie.
W ten sposób i poprzez konwersję mleczanu do glukozy cząsteczki
nie będące glukozą mogą być przekształcone w glukozę
(glukoneogeneza).
•
Glukoza może być przekształcana w tłuszcze poprzez acetylo-
CoA, ponieważ przemiana pirogranianu do acetylo-CoA jest
nieodwracalna. w przeciwieństwie do większości reakcji glikozy,
to jednak tłuszcze nie są ta drogą przekształcane na glukozę.
Całkowita przemiana tłuszczów w węglowodany w ustroju jest
bardzo ograniczona, ponieważ poza nieistotnym ilościowo
tworzeniem węglowodanów z glicerolu nie istnieje żadna inna
droga dla takiej przemiany.

Cykl kwasu cytrynowego
Cykl kwasu cytrynowego
•
Cykl kwasu cytrynowego (cykl Krebsa, cykl kwasów
trikarboksylowych) jest szeregiem reakcji, w których
acetylo-CoA jest metabolizowany do CO
2
i atomów wodoru.
•
Acetylo-CoA ulega kondensacji z anionem 4 – węglowego
kwasu szczawiooctowego, tworząc cytrynian i zredukowany
CoA (HS – CoA).
•
W ciągu siedmiu następujących po sobie reakcji 2
cząsteczki CO
2
odłączają się i regeneruje się cząsteczka
szczawiooctanu.
•
Cztery pary atomów H przenoszone są na łańcuch
flawoproteinowo-cytochromowy, tworząc 12 cząsteczek
ATP i 4 cząsteczki H
2
O, z których dwie są zużyte w cyklu.
•
Cykl kwasu cytrynowego (cykl Krebsa, cykl kwasów
trikarboksylowych) jest szeregiem reakcji, w których
acetylo-CoA jest metabolizowany do CO
2
i atomów wodoru.
•
Acetylo-CoA ulega kondensacji z anionem 4 – węglowego
kwasu szczawiooctowego, tworząc cytrynian i zredukowany
CoA (HS – CoA).
•
W ciągu siedmiu następujących po sobie reakcji 2
cząsteczki CO
2
odłączają się i regeneruje się cząsteczka
szczawiooctanu.
•
Cztery pary atomów H przenoszone są na łańcuch
flawoproteinowo-cytochromowy, tworząc 12 cząsteczek
ATP i 4 cząsteczki H
2
O, z których dwie są zużyte w cyklu.
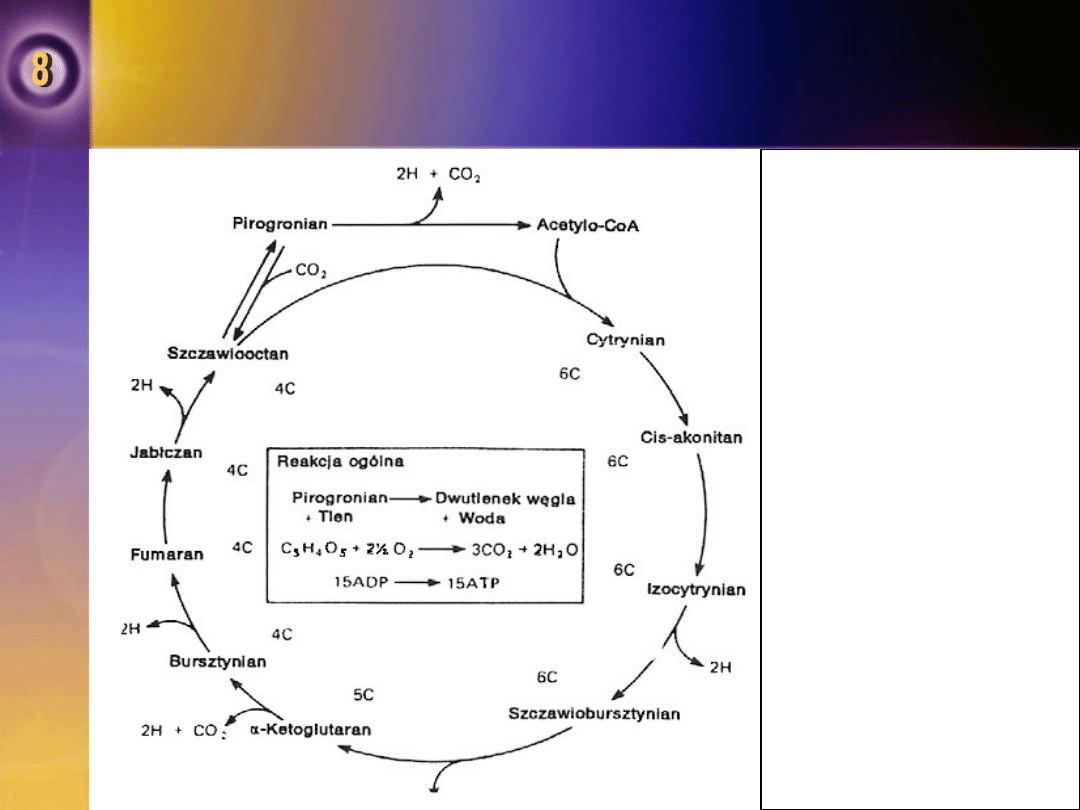
Cykl kwasu cytrynowego
Cykl kwasu cytrynowego
Numery w kole (6C, 5C,4C)
wskazują liczbę atomów
węgla w każdym intermediacie
kwasu.
Dwa atomy H uzyskiwane są
w reakcji:
•pirogronian → acetylo-CoA,
a 8 atomów H uzyskuje się
odpowiednio w każdym obrocie
cyklu, co powoduje utworzenie
w wyniku fosforylacji
oksydacyjnej 15 cząsteczek
ATP.

Cykl kwasu cytrynowego
Cykl kwasu cytrynowego
•
Cykl kwasu cytrynowego jest wspólną drogą utleniania
węglowodanów, tłuszczów i niektórych aminokwasów do
CO
2
i H
2
O.
•
Większość tych związków włącza się przez acetylo-CoA,
ale pirogronian może również włączać się przez
przyłączenie CO
2
(karboksylacja) tworząc szczawiooctan.
•
Niektóre aminokwasy mogą być przekształcane na drodze
deaminacji w związki pośrednie cyklu kwasu cytrynowego.
•
Połączenie pirogronianu z CO
2
w szczawiooctan jest
zaledwie jedną z wielu możliwych reakcji metabolicznych,
w których CO
2
jest substancją budulcową, nie zaś
zbędnym produktem.
•
Cykl kwasu cytrynowego wymaga obecności O
2
i nie
przebiega w warunkach beztlenowych.
•
Cykl kwasu cytrynowego jest wspólną drogą utleniania
węglowodanów, tłuszczów i niektórych aminokwasów do
CO
2
i H
2
O.
•
Większość tych związków włącza się przez acetylo-CoA,
ale pirogronian może również włączać się przez
przyłączenie CO
2
(karboksylacja) tworząc szczawiooctan.
•
Niektóre aminokwasy mogą być przekształcane na drodze
deaminacji w związki pośrednie cyklu kwasu cytrynowego.
•
Połączenie pirogronianu z CO
2
w szczawiooctan jest
zaledwie jedną z wielu możliwych reakcji metabolicznych,
w których CO
2
jest substancją budulcową, nie zaś
zbędnym produktem.
•
Cykl kwasu cytrynowego wymaga obecności O
2
i nie
przebiega w warunkach beztlenowych.

Wytwarzanie energii
Wytwarzanie energii
•
Całkowite wytwarzanie związków fosforanowych bogatych w
energię w czasie metabolizmu glikozy i glikogenu do
pirogranianu zależy od drogi rozkładu i zachodzi przez
przemianę Embdena- Mayerhofa lub drogą cyklu pentozo-
fosforanowego.
•
Przemiana 1 mola aldehydu 3 – fosfoglicerynowego do 3 –
fosfoglicerynianu powoduje wytworzenie 1 mola ATP, a
przemiana 1 mola fosfoenolopirogrogronianu, do
pirogronianu – następnego 1 mola ATP.
•
Ponieważ 1 mol glikozo–6 fosforanu rozpada się w cyklu
Embdena-Mayerhofa na dwa mole aldehydu 3 –
fosfoglicerynowego, powstają zatem 4 mole ATP z 1 mola
glikozy rozłożonej do pirogranianu.
•
Wszystkie te reakcje zachodzą bez udziału O
2
, stanowią
zatem beztlenową drogę udziału O
2
, stanowią zatem
beztlenową drogę wytwarzania energii.
•
Całkowite wytwarzanie związków fosforanowych bogatych w
energię w czasie metabolizmu glikozy i glikogenu do
pirogranianu zależy od drogi rozkładu i zachodzi przez
przemianę Embdena- Mayerhofa lub drogą cyklu pentozo-
fosforanowego.
•
Przemiana 1 mola aldehydu 3 – fosfoglicerynowego do 3 –
fosfoglicerynianu powoduje wytworzenie 1 mola ATP, a
przemiana 1 mola fosfoenolopirogrogronianu, do
pirogronianu – następnego 1 mola ATP.
•
Ponieważ 1 mol glikozo–6 fosforanu rozpada się w cyklu
Embdena-Mayerhofa na dwa mole aldehydu 3 –
fosfoglicerynowego, powstają zatem 4 mole ATP z 1 mola
glikozy rozłożonej do pirogranianu.
•
Wszystkie te reakcje zachodzą bez udziału O
2
, stanowią
zatem beztlenową drogę udziału O
2
, stanowią zatem
beztlenową drogę wytwarzania energii.

Wytwarzanie energii
Wytwarzanie energii
•
Jeden mol ATP jest zużywany dla utworzenia fruktozo -1 , 6
difosforanu z fruktozo–6–fosforanu i 1 mol w procesie
fosforylacji glikozy na glikozo – 6 – fosforan po wniknięciu do
komórki.
•
W konsekwencji, kiedy w warunkach anaerobowych z
glikogenu tworzy się pirogronian, „czysty” zysk energetyczny
wynosi 3 mole ATP na 1 mol glikozo–6–fosforanu.
•
Jeśli jednak pirogronian powstaje z 1 mola glikozy
występującej we krwi, uzyskuje się tylko 2 mole ATP.
•
Do przekształcenia aldehydu fosfoglicerynowego w
fosfoglicerynian konieczne jest dostarczenie NAD
+
.
•
W warunkach tlenowych NADH jest utleniany przez łańcuch
flawoproteinowo-cytrochromowy, odtwarzając NAD
+
i tworząc
ponadto 6 moli ATP na każde 2 mole utlenianego
fosfoglicerynianu.
•
Jeden mol ATP jest zużywany dla utworzenia fruktozo -1 , 6
difosforanu z fruktozo–6–fosforanu i 1 mol w procesie
fosforylacji glikozy na glikozo – 6 – fosforan po wniknięciu do
komórki.
•
W konsekwencji, kiedy w warunkach anaerobowych z
glikogenu tworzy się pirogronian, „czysty” zysk energetyczny
wynosi 3 mole ATP na 1 mol glikozo–6–fosforanu.
•
Jeśli jednak pirogronian powstaje z 1 mola glikozy
występującej we krwi, uzyskuje się tylko 2 mole ATP.
•
Do przekształcenia aldehydu fosfoglicerynowego w
fosfoglicerynian konieczne jest dostarczenie NAD
+
.
•
W warunkach tlenowych NADH jest utleniany przez łańcuch
flawoproteinowo-cytrochromowy, odtwarzając NAD
+
i tworząc
ponadto 6 moli ATP na każde 2 mole utlenianego
fosfoglicerynianu.

Wytwarzanie energii
Wytwarzanie energii
•
W warunkach beztlenowych (glikoza anaerobowa) może nastąpić
zablokowanie procesu glikozy na etapie aldehydu 3 – fosfoglicerynowego.
•
Nastąpi to wtedy, kiedy dostępny NAD
+
, będzie przekształcany w postać
zredukowaną NADH.
•
Pirogronian może przyjmować atomy wodoru z NADH tworząc mleczan i
NAD
+
.
•
W ten sposób metabolizm glikozy i wytwarzanie energii może się
odbywać przez pewien czas bez udziału O2.
•
Mleczan akumuluje się i jest przekształcany w pirogronian, jeśli
przywrócone zostaje dostarczanie O2.
•
Natomiast powstały NADH przenosi swe atomy wodoru na łańcuch
flawoproteinowo – cytrochromowy.
•
W warunkach beztlenowych (glikoza anaerobowa) może nastąpić
zablokowanie procesu glikozy na etapie aldehydu 3 – fosfoglicerynowego.
•
Nastąpi to wtedy, kiedy dostępny NAD
+
, będzie przekształcany w postać
zredukowaną NADH.
•
Pirogronian może przyjmować atomy wodoru z NADH tworząc mleczan i
NAD
+
.
•
W ten sposób metabolizm glikozy i wytwarzanie energii może się
odbywać przez pewien czas bez udziału O2.
•
Mleczan akumuluje się i jest przekształcany w pirogronian, jeśli
przywrócone zostaje dostarczanie O2.
•
Natomiast powstały NADH przenosi swe atomy wodoru na łańcuch
flawoproteinowo – cytrochromowy.
Pirogronian + NADH + H
+
Mleczan
+ NAD
+
Pirogronian + NADH + H
+
Mleczan
+ NAD
+

Wytwarzanie energii
Wytwarzanie energii
•
W czasie glikozy tlenowej ilość wytworzonego ATP jest 19
razy wyższa w porównaniu z 2 molami ATP wytworzonymi
w warunkach beztlenowych.
•
Powstaje nie tylko dodatkowych 6 moli ATP wytworzonych
przez utlenianie NADH droga łańcucha flawoproteinowo -
cytrochromowego , ale utlenianie 2 moli NADH
utworzonych przez przekształcenie 2 moli pirogronianu w
acetylo – CoA powoduje powstanie 6 moli ATP.
•
Każdy obrót cyklu kwasu cytrynowego w warunkach
tlenowych wytwarza 12 moli ATP.
•
•
Zatem całkowite wytwarzanie ATP z 1 mola glikozy krwi
rozłożonej w warunkach tlenowych drogą Embdena-
Mayerhofa i w cyklu kwasu cytrynowego wynosi 38 moli
ATP (2 + [2 x 3] + [2 x3] + [2 x 12])
•
W czasie glikozy tlenowej ilość wytworzonego ATP jest 19
razy wyższa w porównaniu z 2 molami ATP wytworzonymi
w warunkach beztlenowych.
•
Powstaje nie tylko dodatkowych 6 moli ATP wytworzonych
przez utlenianie NADH droga łańcucha flawoproteinowo -
cytrochromowego , ale utlenianie 2 moli NADH
utworzonych przez przekształcenie 2 moli pirogronianu w
acetylo – CoA powoduje powstanie 6 moli ATP.
•
Każdy obrót cyklu kwasu cytrynowego w warunkach
tlenowych wytwarza 12 moli ATP.
•
•
Zatem całkowite wytwarzanie ATP z 1 mola glikozy krwi
rozłożonej w warunkach tlenowych drogą Embdena-
Mayerhofa i w cyklu kwasu cytrynowego wynosi 38 moli
ATP (2 + [2 x 3] + [2 x3] + [2 x 12])

Wytwarzanie energii
Wytwarzanie energii
•
Utlenianie glikozy droga heksozomobofosforanową
dostarcza dużych ilości NADPH.
•
Uzyskanie tego zredukowanego koenzymu ma
podstawowe znaczenie dla wielu procesów
metabolicznych.
•
Wytworzone w tym procesie pentozy są elementami
budowy nukleotydów.
•
Ilość wytworzonego ATP zależy, od ilości NADPH
przekształcanego w NADH, a następnie utlenianego w
łańcuchu flawoproteinowo-cytrochromowym.
•
Utlenianie glikozy droga heksozomobofosforanową
dostarcza dużych ilości NADPH.
•
Uzyskanie tego zredukowanego koenzymu ma
podstawowe znaczenie dla wielu procesów
metabolicznych.
•
Wytworzone w tym procesie pentozy są elementami
budowy nukleotydów.
•
Ilość wytworzonego ATP zależy, od ilości NADPH
przekształcanego w NADH, a następnie utlenianego w
łańcuchu flawoproteinowo-cytrochromowym.
„Zastawki
jednokierunkowego
przepływu”
„Zastawki
jednokierunkowego
przepływu”
•
Metabolizm jest regulowany
przez wiele hormonów oraz inne
czynniki.
•
Aby uzyskać jakiekolwiek zmiany
w poszczególnych procesach
metabolicznych, czynniki
regulujące muszą poprowadzić
reakcję chemiczną w jednym
kierunku.
•
Większość reakcji w przemianie
pośredniej jest dowolnie
odwracalna, ale istnieje pewna
liczba reakcji stanowiących
„zastawki jednokierunkowego
przepływu”, przebiegających w
jednym kierunku pod wpływem
jednego enzymu lub
mechanizmu transportującego, a
w przeciwnym kierunku pod
działaniem innych enzymów lub
czynników.
•
Metabolizm jest regulowany
przez wiele hormonów oraz inne
czynniki.
•
Aby uzyskać jakiekolwiek zmiany
w poszczególnych procesach
metabolicznych, czynniki
regulujące muszą poprowadzić
reakcję chemiczną w jednym
kierunku.
•
Większość reakcji w przemianie
pośredniej jest dowolnie
odwracalna, ale istnieje pewna
liczba reakcji stanowiących
„zastawki jednokierunkowego
przepływu”, przebiegających w
jednym kierunku pod wpływem
jednego enzymu lub
mechanizmu transportującego, a
w przeciwnym kierunku pod
działaniem innych enzymów lub
czynników.
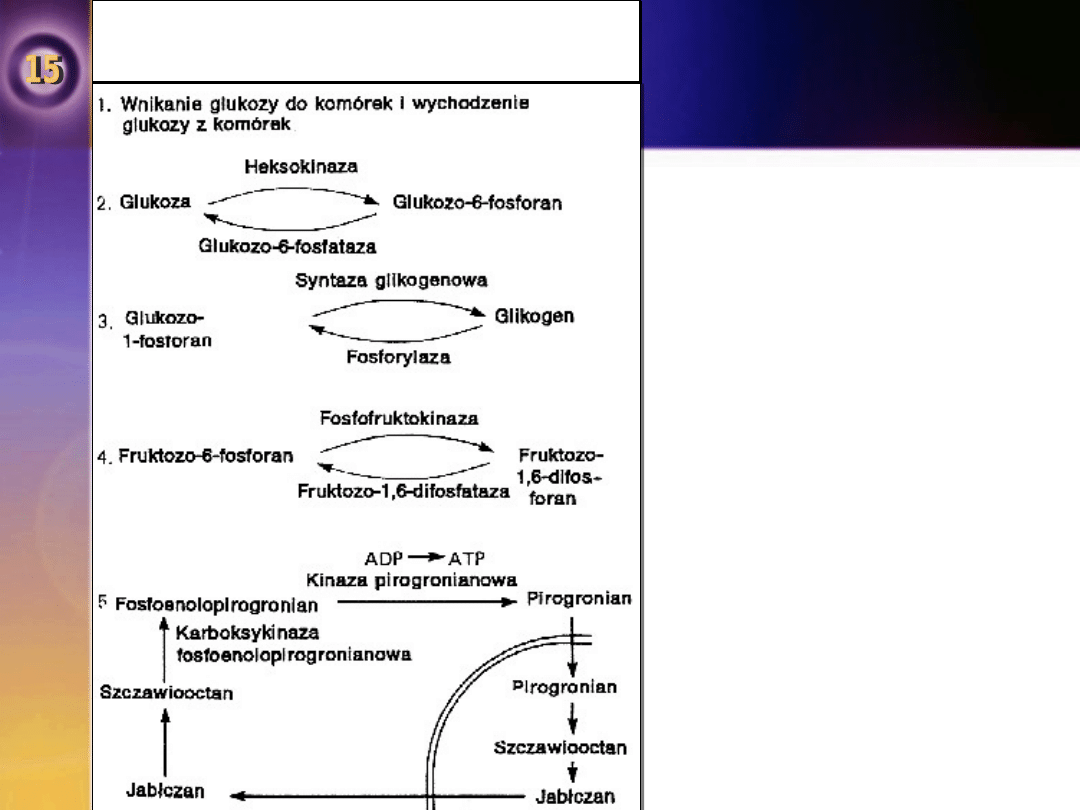
Pięć przykładów „jednokierunkowego krążenia”
w metabolizmie węglowodanów
Pięć przykładów „jednokierunkowego krążenia”
w metabolizmie węglowodanów
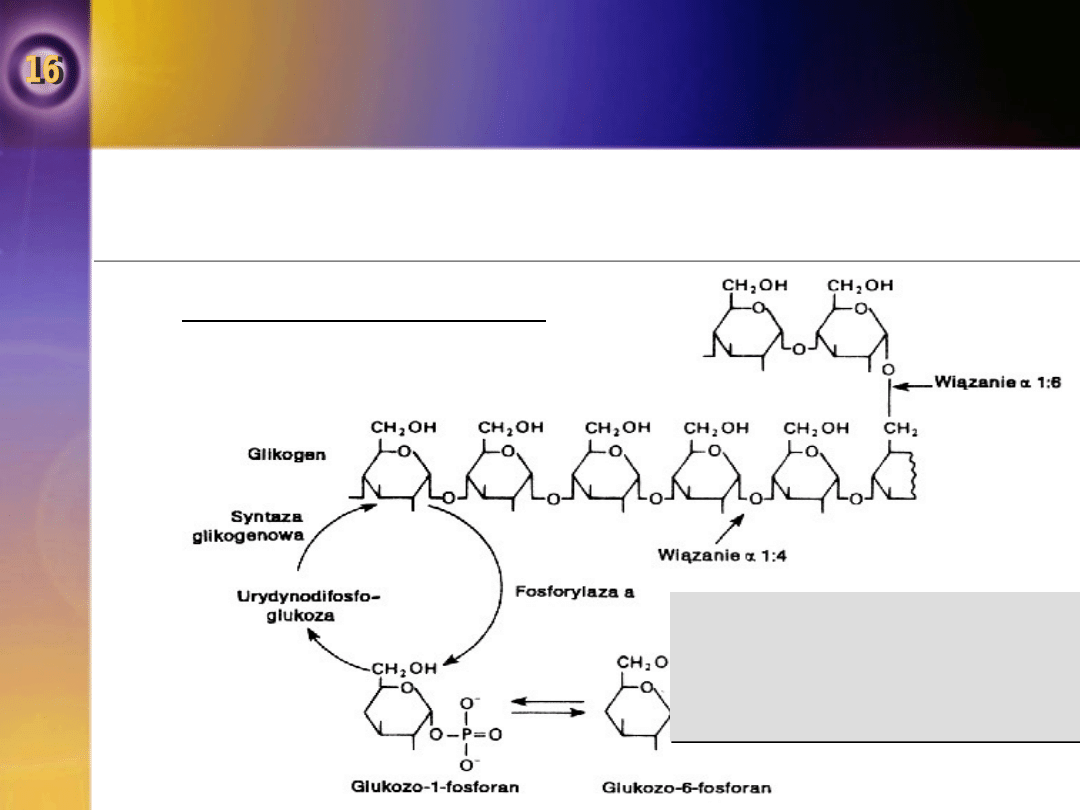
Fosforylaza
Fosforylaza
•
Rozpad glikogenu regulowany jest przez kilka hormonów.
•
Glikogen syntetyzowany jest z glukozo-1-fosforanu drogą urydynodifosfoglukozy
(UDPG) z udziałem enzymu syntezy glikogenowej katalizującej końcowy etap.
•
Rozpad glikogenu regulowany jest przez kilka hormonów.
•
Glikogen syntetyzowany jest z glukozo-1-fosforanu drogą urydynodifosfoglukozy
(UDPG) z udziałem enzymu syntezy glikogenowej katalizującej końcowy etap.
Synteza i rozpad glikogenu
Glikogen jest rozgałęzionym
polimerem glikozy z dwoma
rodzajami wiązań glikozydowych.
Cięcie wiązania alpaha 1:6 w
punktach rozgałęzienia katalizowane
jest przez inny enzym.
Glikogen jest rozgałęzionym
polimerem glikozy z dwoma
rodzajami wiązań glikozydowych.
Cięcie wiązania alpaha 1:6 w
punktach rozgałęzienia katalizowane
jest przez inny enzym.
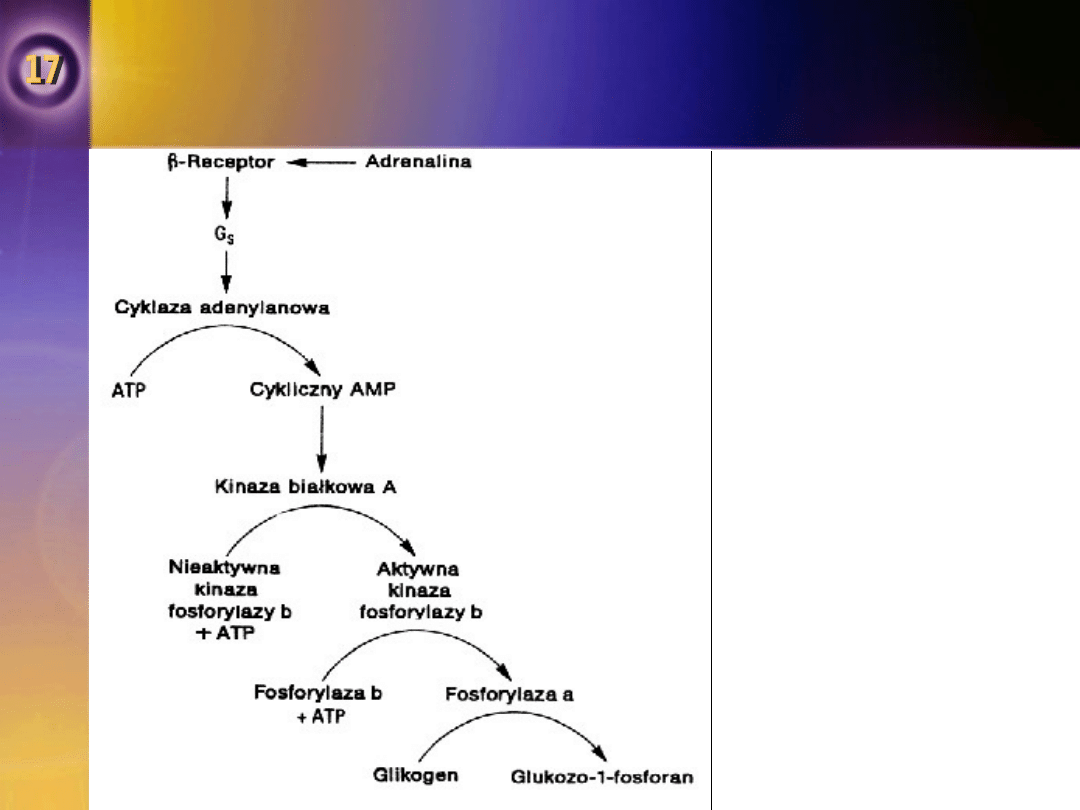
Fosforylaza
Fosforylaza
•
W wątrobie
fosforylaza
aktywowana jest
częściowo przez
adrenalinę za
pośrednictwem
receptorów beta-
adrenergicznych.
•
Zapoczątkowuje to
ciąg reakcji, które
stanowią klasyczny
przykład działania
hormonalnego przy
udziale cyklicznego
AMP
.
•
W wątrobie
fosforylaza
aktywowana jest
częściowo przez
adrenalinę za
pośrednictwem
receptorów beta-
adrenergicznych.
•
Zapoczątkowuje to
ciąg reakcji, które
stanowią klasyczny
przykład działania
hormonalnego przy
udziale cyklicznego
AMP
.
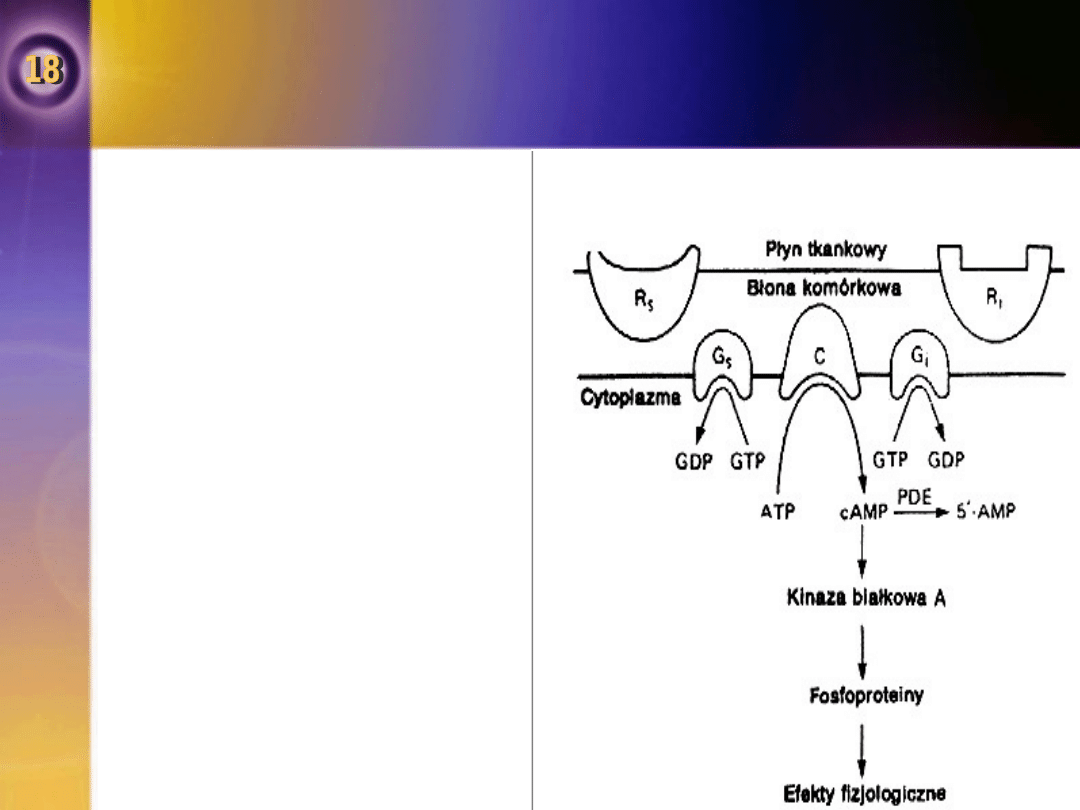
Fosforylaza
Fosforylaza
•
Kinaza białkowa A
aktywowana jest przez
cykliczny AMP i katalizuje
przeniesienie grupy
fosforanowej na kinazę
fosforylazy,
przekształcając ją w
formę aktywną.
•
Z kolei kinaza fosforylazy
katalizuje fosforylację
aktywując fosforylazę.
•
Nieaktywna fosforylaza
znana jest jako
fosforylaza b
(defosforylaza) a
fosforylaza aktywna jako
fosforylaza a
(fosfosforylaza).
•
Kinaza białkowa A
aktywowana jest przez
cykliczny AMP i katalizuje
przeniesienie grupy
fosforanowej na kinazę
fosforylazy,
przekształcając ją w
formę aktywną.
•
Z kolei kinaza fosforylazy
katalizuje fosforylację
aktywując fosforylazę.
•
Nieaktywna fosforylaza
znana jest jako
fosforylaza b
(defosforylaza) a
fosforylaza aktywna jako
fosforylaza a
(fosfosforylaza).
Fosforylaza
Fosforylaza
•
Duże dawki
wazopresyny i
angiotensyny II mogą
również powodować
glikogenolizę poprzez
ten mechanizm, ale jest
wątpliwe, czy te
hormony odgrywają
jakakolwiek fizjologiczna
rolę w homeostazie
glukozy.
•
Duże dawki
wazopresyny i
angiotensyny II mogą
również powodować
glikogenolizę poprzez
ten mechanizm, ale jest
wątpliwe, czy te
hormony odgrywają
jakakolwiek fizjologiczna
rolę w homeostazie
glukozy.
•
Aktywacja kinezy białkowej A przez cykliczny AMP nie tylko nasila
proces glikogenu, ale także hamuje jego syntezę.
•
Rozpad glikogenu następuje również w wątrobie po działaniu
katecholamin za pośrednictwem receptorów alpha-adrenergicznych.
•
W rozpadzie tym uczestniczy wewnątrzkomórkowy Ca
2+
, który
powoduje aktywację kinazy fosforylazy, niezależnie od cyklicznego
AMP.
•
Aktywacja kinezy białkowej A przez cykliczny AMP nie tylko nasila
proces glikogenu, ale także hamuje jego syntezę.
•
Rozpad glikogenu następuje również w wątrobie po działaniu
katecholamin za pośrednictwem receptorów alpha-adrenergicznych.
•
W rozpadzie tym uczestniczy wewnątrzkomórkowy Ca
2+
, który
powoduje aktywację kinazy fosforylazy, niezależnie od cyklicznego
AMP.

Fosforylaza
Fosforylaza
•
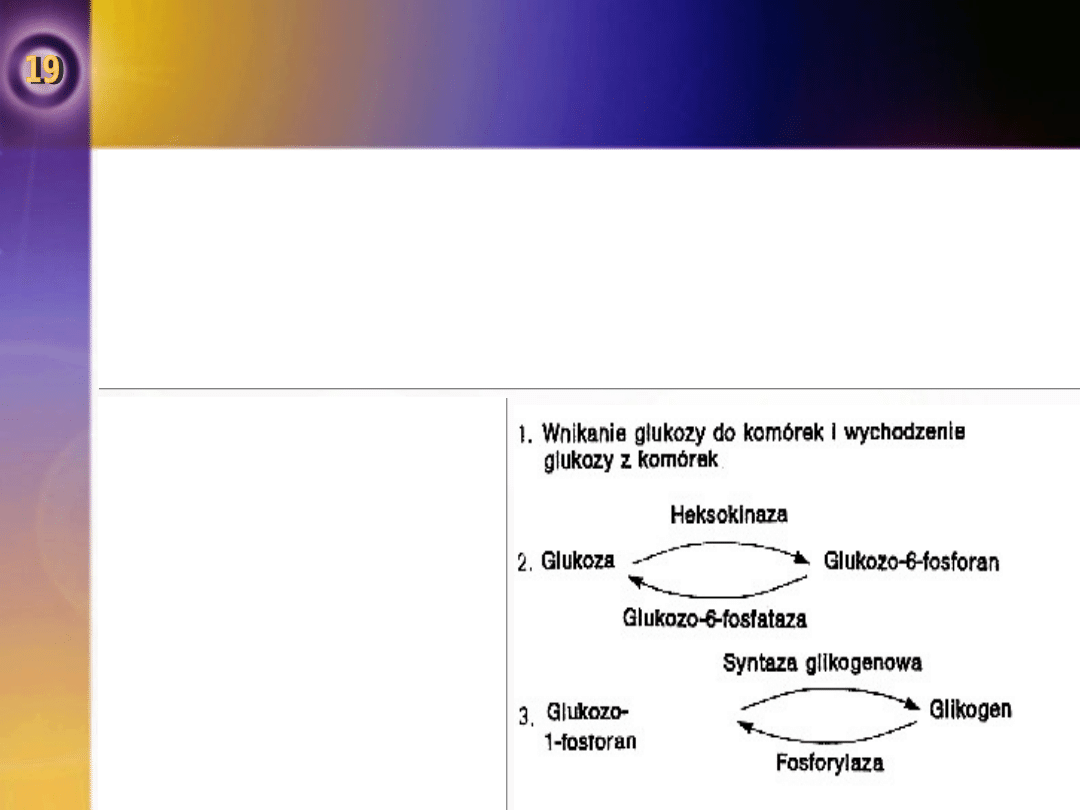
Ponieważ wątroba zawiera enzym glukozo–6–fosfotazę,
znaczna część glikozo-6-fosforanu wytwarzanego w tym
narządzie może być przekształcana w glukozę i dostawać
się do krwi krążącej, podwyższając w ten sposób stężenie
glukozy we krwi.
•
W procesie tym mogą brać udział również nerki.
•
Inne tkanki nie zawierają tego enzymu, mimo to duża
część glukozo-6-fosforanu jest w nich katabolizowana w
dwóch procesach:
Embdena-Meyerhofa,
przemianie heksomonofosforanowej.
•
Tak podwyższony katabolizm glukozy w mięśniach
szkieletowych powoduje zwiększenie stężenia mleczanu
we krwi.
•
Ponieważ wątroba zawiera enzym glukozo–6–fosfotazę,
znaczna część glikozo-6-fosforanu wytwarzanego w tym
narządzie może być przekształcana w glukozę i dostawać
się do krwi krążącej, podwyższając w ten sposób stężenie
glukozy we krwi.
•
W procesie tym mogą brać udział również nerki.
•
Inne tkanki nie zawierają tego enzymu, mimo to duża
część glukozo-6-fosforanu jest w nich katabolizowana w
dwóch procesach:
Embdena-Meyerhofa,
przemianie heksomonofosforanowej.
•
Tak podwyższony katabolizm glukozy w mięśniach
szkieletowych powoduje zwiększenie stężenia mleczanu
we krwi.

Fosforylaza
Fosforylaza
•
Poprzez stymulację cyklazy adenylanowej
adrenalina powoduje aktywację fosforylazy w
wątrobie i mięśniach szkieletowych.
•
Konsekwencją tego procesu jest zwiększenie
stężenia we krwi zarówno glukozy, jak i
mleczanu.
•
Glikagon działa podobnie, ale podwyższa
aktywność fosforylazy tylko w wątrobie.
•
W konsekwencji glikagon wywołuje zwiększenie
stężenia glukozy we krwi bez jakichkolwiek zmian
w stężeniu mleczanu.
•
Poprzez stymulację cyklazy adenylanowej
adrenalina powoduje aktywację fosforylazy w
wątrobie i mięśniach szkieletowych.
•
Konsekwencją tego procesu jest zwiększenie
stężenia we krwi zarówno glukozy, jak i
mleczanu.
•
Glikagon działa podobnie, ale podwyższa
aktywność fosforylazy tylko w wątrobie.
•
W konsekwencji glikagon wywołuje zwiększenie
stężenia glukozy we krwi bez jakichkolwiek zmian
w stężeniu mleczanu.

Choroba McArdle’a
Choroba McArdle’a
•
Glikogenoza z niedoboru miofosforylazy znana jest w
patologii jako choroba McArdle’a.
•
Glikogen gromadzi się w mięśniach szkieletowych z
powodu niedoboru fosforylazy mięśniowej.
•
Chorzy na tę chorobę uskarżają się na ból mięśni,
usztywnienie w czasie wysiłku i wykazują bardzo obniżoną
tolerancję na wysiłek fizyczny.
•
W ich mięśniach glikogen nie może być rozkładany w celu
dostarczenia energii dla skurczu mięśni, glukoza zaś
dostarczona z krwi krążącej do mięśni wystarcza tylko na
bardzo ograniczony wysiłek fizyczny.
•
Po wstrzyknięciu glukagonu lub adrenaliny we krwi tych
pacjentów dochodzi do prawidłowego zwiększenia stężenia
glikozy, co wskazuje, że aktywność fosforylazy wątrobowej
jest prawidłowa.
•
Glikogenoza z niedoboru miofosforylazy znana jest w
patologii jako choroba McArdle’a.
•
Glikogen gromadzi się w mięśniach szkieletowych z
powodu niedoboru fosforylazy mięśniowej.
•
Chorzy na tę chorobę uskarżają się na ból mięśni,
usztywnienie w czasie wysiłku i wykazują bardzo obniżoną
tolerancję na wysiłek fizyczny.
•
W ich mięśniach glikogen nie może być rozkładany w celu
dostarczenia energii dla skurczu mięśni, glukoza zaś
dostarczona z krwi krążącej do mięśni wystarcza tylko na
bardzo ograniczony wysiłek fizyczny.
•
Po wstrzyknięciu glukagonu lub adrenaliny we krwi tych
pacjentów dochodzi do prawidłowego zwiększenia stężenia
glikozy, co wskazuje, że aktywność fosforylazy wątrobowej
jest prawidłowa.

„Glukostat wątrobowy”
„Glukostat wątrobowy”
•
Jeśli stężenie we krwi jest duże, to zachodzi bezpośrednie
pobieranie jej prze wątrobę, a gdy stężenie jej we krwi jest
małe, następuje uwalnianie glukozy z wątroby do krwi.
•
Wątroba zatem funkcjonuje jako rodzaj „glukostatu”
(regulator stężenia glukozy”), utrzymującego stałe stężenie
glukozy we krwi krążącej.
•
Czynność ta nie jest automatyczna, pobieranie i uwalnianie
glikozy z wątroby zachodzi pod wpływem wielu hormonów.
Wyspy trzustkowe (Langerhansa) w trzustce wydzielają co
najmniej cztery hormony peptydowe.
insulina i glukagon – spełniają istotną rolę w regulacji
pośredniej przemiany węglowodanów, białek i tłuszczów.
somatostyna – bierze udział w regulacji czynności
wydzielniczej komórek wysp trzustkowych,
polipeptyd trzustkowy – fizjologiczna rola tego hormonu
nie jest do końca poznana
•
Jeśli stężenie we krwi jest duże, to zachodzi bezpośrednie
pobieranie jej prze wątrobę, a gdy stężenie jej we krwi jest
małe, następuje uwalnianie glukozy z wątroby do krwi.
•
Wątroba zatem funkcjonuje jako rodzaj „glukostatu”
(regulator stężenia glukozy”), utrzymującego stałe stężenie
glukozy we krwi krążącej.
•
Czynność ta nie jest automatyczna, pobieranie i uwalnianie
glikozy z wątroby zachodzi pod wpływem wielu hormonów.
Wyspy trzustkowe (Langerhansa) w trzustce wydzielają co
najmniej cztery hormony peptydowe.
insulina i glukagon – spełniają istotną rolę w regulacji
pośredniej przemiany węglowodanów, białek i tłuszczów.
somatostyna – bierze udział w regulacji czynności
wydzielniczej komórek wysp trzustkowych,
polipeptyd trzustkowy – fizjologiczna rola tego hormonu
nie jest do końca poznana

Przemieszczanie glukozy w nerkach
Przemieszczanie glukozy w nerkach
•
W nerkach glukoza jest swobodnie przesączana, lecz przy
normalnym stężeniu we krwi, prawie cała, poza niewielka
ilością, jest resorbowana w kanalikach nerkowych bliższych.
•
Jeśli ilość przesączanej glikozy zwiększa się, wzrasta też
resorpcja, ale istnieje limit ilości glukozy, którą kanaliki
bliższe mogą resorbować.
•
Gdy transport maksymalny dla glukozy (Tmg) jest
przekroczony, znaczne ilości glukozy pojawiają się w moczu
(glikozuria).
•
Nerkowy próg dla glukozy, czyli stężenie glukozy we krwi
tętniczej, przy którym pojawia się glikozuria, osiągany jest
wtedy, kiedy stężenie glukozy we krwi żylnej wynosi około
10,0 mmol/L (180 mg/dL), ale może być wyższe, jeżeli
wielkość filtracji kłębuszkowej jest niska.
•
W nerkach glukoza jest swobodnie przesączana, lecz przy
normalnym stężeniu we krwi, prawie cała, poza niewielka
ilością, jest resorbowana w kanalikach nerkowych bliższych.
•
Jeśli ilość przesączanej glikozy zwiększa się, wzrasta też
resorpcja, ale istnieje limit ilości glukozy, którą kanaliki
bliższe mogą resorbować.
•
Gdy transport maksymalny dla glukozy (Tmg) jest
przekroczony, znaczne ilości glukozy pojawiają się w moczu
(glikozuria).
•
Nerkowy próg dla glukozy, czyli stężenie glukozy we krwi
tętniczej, przy którym pojawia się glikozuria, osiągany jest
wtedy, kiedy stężenie glukozy we krwi żylnej wynosi około
10,0 mmol/L (180 mg/dL), ale może być wyższe, jeżeli
wielkość filtracji kłębuszkowej jest niska.

Glikozuria
Glikozuria
•
Glikozuria pojawia się wtedy, kiedy zwiększa się stężenie
glukozy we krwi w wyniku względnego niedoboru insuliny
(diabetes melitus) lub w wyniku nadmiernej glikogenolizy
po urazie fizycznym lub psychicznym.
•
U niektórych ludzi mechanizm transportu glukozy w
kanalikach nerkowych jest wadliwy z przyczyn wrodzonych i
dlatego glikozuria występuje przy prawidłowym stężeniu
glukozy we krwi.
•
Stan ten nosi nazwę glikozurii nerkowej. Natomiast po
posiłku bogatym w węglowodany u zdrowych ludzi pojawia
się tzw. glikozuria pokarmowa.
•
U wielu z tych osób stwierdza się łagodna postać cukrzycy.
•
maksymalny wskaźnik absorpcji glukozy z jelita wynosi
około 0,67 mol mol/h (120 g/h).
•
Glikozuria pojawia się wtedy, kiedy zwiększa się stężenie
glukozy we krwi w wyniku względnego niedoboru insuliny
(diabetes melitus) lub w wyniku nadmiernej glikogenolizy
po urazie fizycznym lub psychicznym.
•
U niektórych ludzi mechanizm transportu glukozy w
kanalikach nerkowych jest wadliwy z przyczyn wrodzonych i
dlatego glikozuria występuje przy prawidłowym stężeniu
glukozy we krwi.
•
Stan ten nosi nazwę glikozurii nerkowej. Natomiast po
posiłku bogatym w węglowodany u zdrowych ludzi pojawia
się tzw. glikozuria pokarmowa.
•
U wielu z tych osób stwierdza się łagodna postać cukrzycy.
•
maksymalny wskaźnik absorpcji glukozy z jelita wynosi
około 0,67 mol mol/h (120 g/h).

Czynniki warunkujące stężenie glukozy we
krwi
Czynniki warunkujące stężenie glukozy we
krwi
•
Stężenie glukozy we krwi w dowolnym czasie
jest zależne od równowagi pomiędzy ilością
glukozy opuszczającą krew krążącą.
•
Podstawowymi czynnikami warunkującymi
stężenie glukozy we krwi są:
przyjmowanie pokarmu,
szybkość wnikania glukozy do;
miocytów,
do tkanki tłuszczowej,
do innych narządów,
aktywność glukostatyczna wątroby.
•
Stężenie glukozy we krwi w dowolnym czasie
jest zależne od równowagi pomiędzy ilością
glukozy opuszczającą krew krążącą.
•
Podstawowymi czynnikami warunkującymi
stężenie glukozy we krwi są:
przyjmowanie pokarmu,
szybkość wnikania glukozy do;
miocytów,
do tkanki tłuszczowej,
do innych narządów,
aktywność glukostatyczna wątroby.
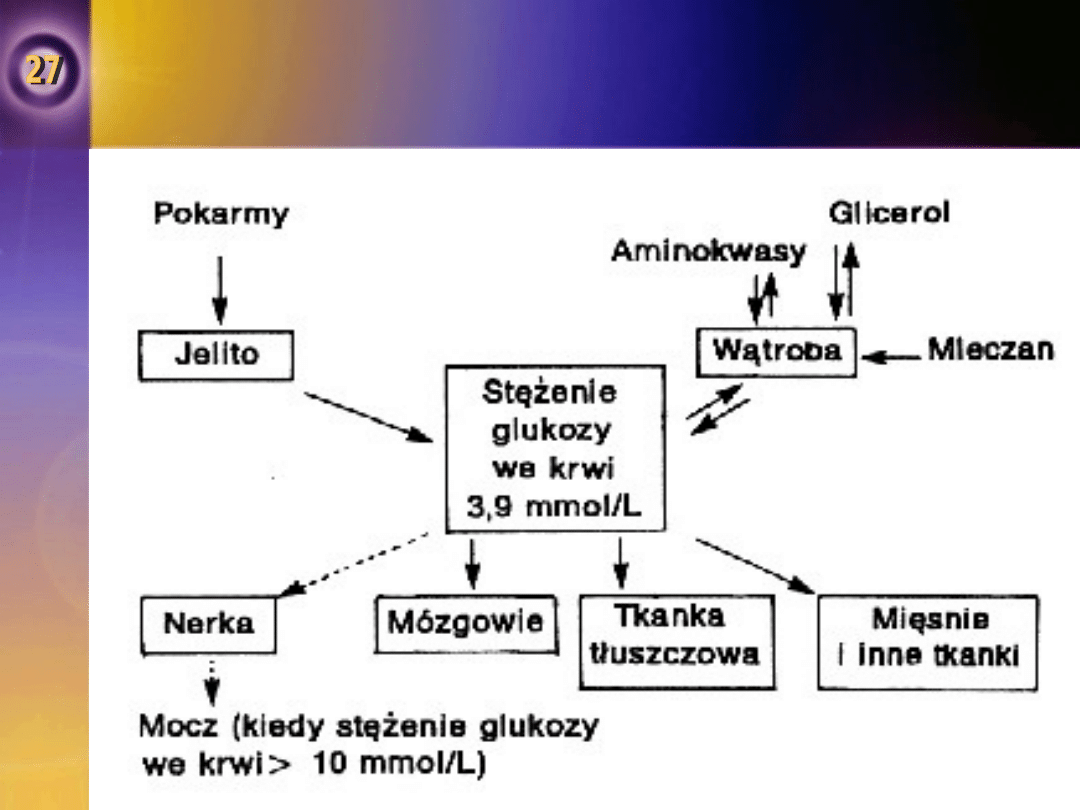
Rola wątroby w utrzymaniu stałego
stężenia glukozy we krwi
Rola wątroby w utrzymaniu stałego
stężenia glukozy we krwi

Czynniki warunkujące stężenie glukozy we
krwi
Czynniki warunkujące stężenie glukozy we
krwi
•
Pięć procent spożytej glukozy jest natychmiast zamieniane w
wątrobie na glikogen, a 30 – 40% jest zamienione na tłuszcze.
•
Pozostała część jest metabolizowana w mięśniach i w innych
tkankach.
•
Na czczo glikogen wątrobowy jest rozkładany i wątroba uwalnia
glukozę do krwi. Jeśli okres na czczo przedłuża się i w wątrobie
następuje wzrost glukogenezy z aminokwasów i glicerolu.
•
Dochodzi do umiarkowanego zmniejszania stężenia glukozy we
krwi do około 3,61 mmol/L 965 mg/dL) u mężczyzn i do około
2,22 mmol/L (40 mg/dL) u kobiet w okresie przed pokwitaniem,
a glukogeneza zapobiega pojawianiu się objawów cięższej
hipoglikemii, nawet w okresie przedłużonego głodzenia.
•
Przyczyna mniejszego stężenia glukozy we krwi na czczo u
kobiet nie jest znana, ale podobne wartości występują również
u chłopców w okresie dojrzewania.
•
Pięć procent spożytej glukozy jest natychmiast zamieniane w
wątrobie na glikogen, a 30 – 40% jest zamienione na tłuszcze.
•
Pozostała część jest metabolizowana w mięśniach i w innych
tkankach.
•
Na czczo glikogen wątrobowy jest rozkładany i wątroba uwalnia
glukozę do krwi. Jeśli okres na czczo przedłuża się i w wątrobie
następuje wzrost glukogenezy z aminokwasów i glicerolu.
•
Dochodzi do umiarkowanego zmniejszania stężenia glukozy we
krwi do około 3,61 mmol/L 965 mg/dL) u mężczyzn i do około
2,22 mmol/L (40 mg/dL) u kobiet w okresie przed pokwitaniem,
a glukogeneza zapobiega pojawianiu się objawów cięższej
hipoglikemii, nawet w okresie przedłużonego głodzenia.
•
Przyczyna mniejszego stężenia glukozy we krwi na czczo u
kobiet nie jest znana, ale podobne wartości występują również
u chłopców w okresie dojrzewania.

Homeostaza węglowodanowa podczas
wysiłku fizycznego
Homeostaza węglowodanowa podczas
wysiłku fizycznego
•
U mężczyzny o masie ciała 70 kg całkowita rezerwa
węglowodanów wynosi około 10 467 kJ (2500 kcal);
zgromadzona jest w:
400g glikogenu mięśni;
100 g glikogenu wątroby;
20 g glukozy płynu zewnątrzkomórkowego.
•
Dla porównania 496 MJ (112 000 kcal) (około 80% dostarczanej
energii) gromadzone jest w tłuszczu, a pozostałość w białku.
•
W spoczynku mięśnie zużywają kwasy tłuszczowe
metabolizując je.
•
U ludzi w spoczynku na czczo mózgowie odpowiedzialne jest
za zużycie 70-80% glukozy, za pozostałą część w
przeważającej części erytrocyty.
•
U mężczyzny o masie ciała 70 kg całkowita rezerwa
węglowodanów wynosi około 10 467 kJ (2500 kcal);
zgromadzona jest w:
400g glikogenu mięśni;
100 g glikogenu wątroby;
20 g glukozy płynu zewnątrzkomórkowego.
•
Dla porównania 496 MJ (112 000 kcal) (około 80% dostarczanej
energii) gromadzone jest w tłuszczu, a pozostałość w białku.
•
W spoczynku mięśnie zużywają kwasy tłuszczowe
metabolizując je.
•
U ludzi w spoczynku na czczo mózgowie odpowiedzialne jest
za zużycie 70-80% glukozy, za pozostałą część w
przeważającej części erytrocyty.
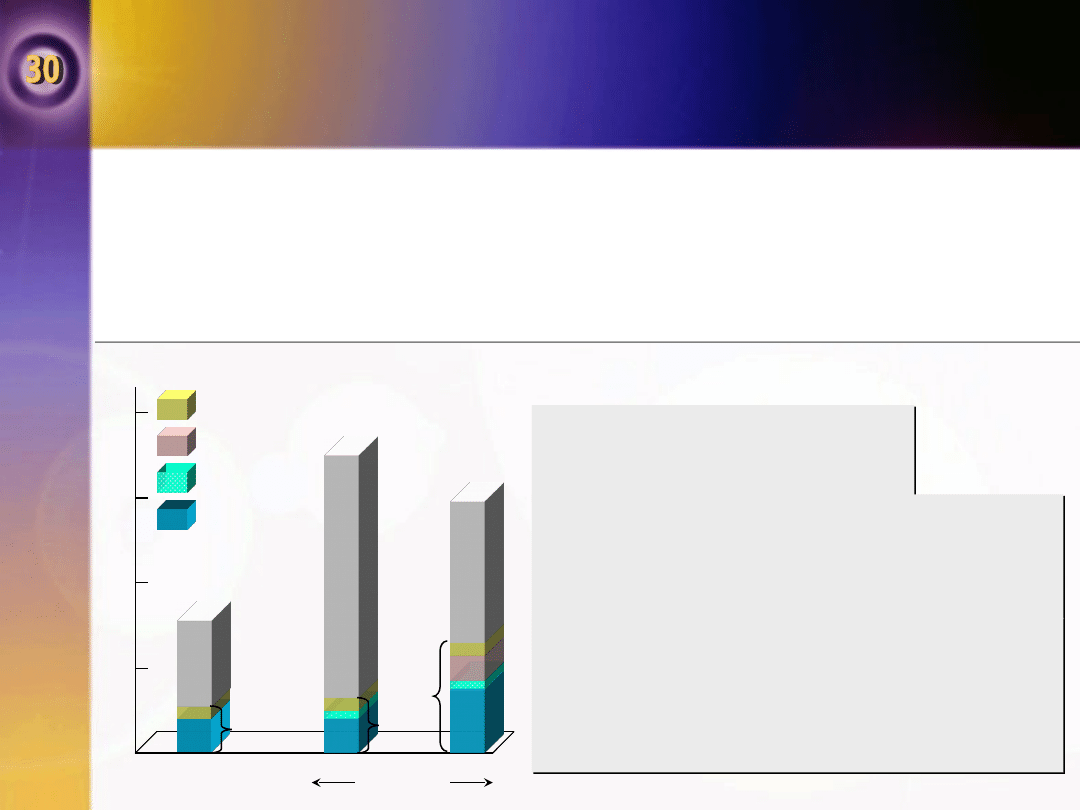
Homeostaza węglowodanowa podczas
wysiłku fizycznego
Homeostaza węglowodanowa podczas
wysiłku fizycznego
•
W czasie wysiłku zapotrzebowanie energetyczne mięśni jest
początkowo pokrywane przez glikogenolizę w mięśniach i wzrastające
pochłanianie glukozy przez mięśnie.
•
Stężenie glukozy we krwi początkowo zwiększa się wraz ze wzrostem
glikogenolizy w wątrobie, ale może zmniejszać się podczas wytężonego,
przedłużonego wysiłku fizycznego. Występuje nasilenie glukogenezy.
•
W czasie wysiłku zapotrzebowanie energetyczne mięśni jest
początkowo pokrywane przez glikogenolizę w mięśniach i wzrastające
pochłanianie glukozy przez mięśnie.
•
Stężenie glukozy we krwi początkowo zwiększa się wraz ze wzrostem
glikogenolizy w wątrobie, ale może zmniejszać się podczas wytężonego,
przedłużonego wysiłku fizycznego. Występuje nasilenie glukogenezy.
0
0,5
1,0
1,5
2,0
Spoczynek
Wysił
ek
fizycz
ny
Aminokwasy
Glicerol
Pirogronian
Mleczan
4
0
240 min
16%
45%
25%
Stężenie insuliny w
osoczu zmniejsza się, a
glikagonu zwiększa się
Stężenie insuliny w
osoczu zmniejsza się, a
glikagonu zwiększa się
Po wysiłku fizycznym glikogen
wątroby jest ponownie uzupełniany
przez dodatkową glukogenezę i
obniżenie uwalniania glukozy z
wątroby.
Po wysiłku fizycznym glikogen
wątroby jest ponownie uzupełniany
przez dodatkową glukogenezę i
obniżenie uwalniania glukozy z
wątroby.
Stężenie insuliny wyraźnie się
zwiększa, zwłaszcza we krwi żyły
wrotnej. Insulina wnikająca do
wątroby przypuszczalnie przyczynia
się do odkładania glikogenu.
Stężenie insuliny wyraźnie się
zwiększa, zwłaszcza we krwi żyły
wrotnej. Insulina wnikająca do
wątroby przypuszczalnie przyczynia
się do odkładania glikogenu.
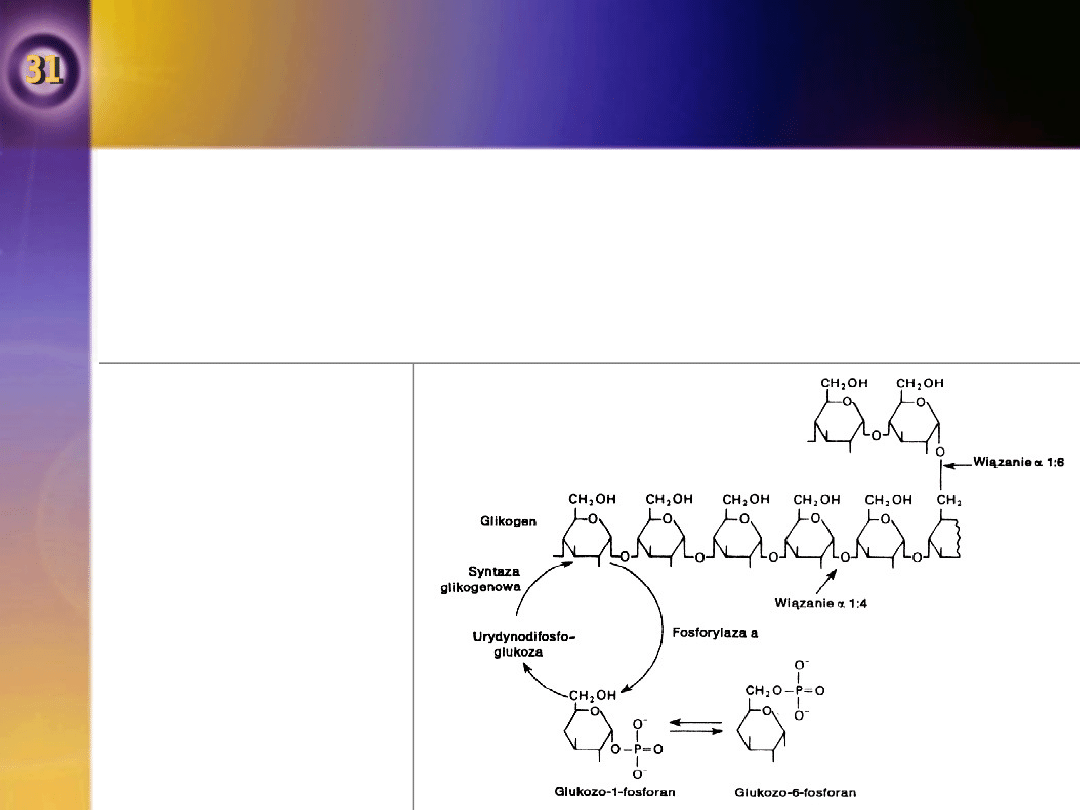
Metabolizm heksoz innych niż
glukoza
Metabolizm heksoz innych niż
glukoza
•
Ta ostatnia reakcja
jest odwracalna i
dostarcza glaktozę
niezbędną do
tworzenia
glikolipidów i
mukoprotein, gdy
pobieranie glaktozy
z pokarmu jest
niedostateczne.
•
Wykorzystanie
glaktozy – podobnie
jak glukozy –
zależne jest od
obecności insuliny.
•
Ta ostatnia reakcja
jest odwracalna i
dostarcza glaktozę
niezbędną do
tworzenia
glikolipidów i
mukoprotein, gdy
pobieranie glaktozy
z pokarmu jest
niedostateczne.
•
Wykorzystanie
glaktozy – podobnie
jak glukozy –
zależne jest od
obecności insuliny.
•
Innymi heksozami absorbowanymi z przewodu pokarmowego są;
glaktoza, która uwalnia się przez trawienie laktozy i przekształcana jest
w ustroju w glukozę; fruktoza, która w pewnej części dostaje się do
organizmu z pokarmami, a w części pochodzi z hydrolizy sacharozy.
•
Po fosforylacji glaktoza jest przekształacana do postaci
urydynodifosfosfoglukozę, która bierze udział w syntezie glikogenu.
•
Innymi heksozami absorbowanymi z przewodu pokarmowego są;
glaktoza, która uwalnia się przez trawienie laktozy i przekształcana jest
w ustroju w glukozę; fruktoza, która w pewnej części dostaje się do
organizmu z pokarmami, a w części pochodzi z hydrolizy sacharozy.
•
Po fosforylacji glaktoza jest przekształacana do postaci
urydynodifosfosfoglukozę, która bierze udział w syntezie glikogenu.

Metabolizm heksoz innych niż
glukoza
Metabolizm heksoz innych niż
glukoza
•
We wrodzonej wadzie metabolicznej znanej jako
glaktozemia występuje wrodzony niedobór transferazy
urydylofosfogalaktozy – enzymu odpowiedzialnego za
reakcję między glaktozo-1-fosforanem i
urydynodifosfoglukozą
.
•
Dostarczana wówczas z pożywieniem glaktoza gromadzi
się we krwi krążącej. Prowadzi to do poważnych zaburzeń
wzrostu i rozwoju dziecka.
•
Stosowanie diety pozbawionej galaktozy poprawia te
warunki, bez wywołania objawów glaktozy, ponieważ są
obecne enzymy konieczne do tworzenia
urydynodifosfogalaktozy z urydynodifosoglukozy.
•
We wrodzonej wadzie metabolicznej znanej jako
glaktozemia występuje wrodzony niedobór transferazy
urydylofosfogalaktozy – enzymu odpowiedzialnego za
reakcję między glaktozo-1-fosforanem i
urydynodifosfoglukozą
.
•
Dostarczana wówczas z pożywieniem glaktoza gromadzi
się we krwi krążącej. Prowadzi to do poważnych zaburzeń
wzrostu i rozwoju dziecka.
•
Stosowanie diety pozbawionej galaktozy poprawia te
warunki, bez wywołania objawów glaktozy, ponieważ są
obecne enzymy konieczne do tworzenia
urydynodifosfogalaktozy z urydynodifosoglukozy.

Metabolizm heksoz innych niż
glukoza
Metabolizm heksoz innych niż
glukoza
•
Fruktoza jest przekształcania częściowo w fruktozo-6-fosforan, a
następnie metabolizowana, przez fruktozo – 1,6-difosforan (
slajd – 6
).
•
Enzym katalizujący powstawanie fruktozo-6-fosforanu to
heksokinaza – ten sam enzym, który katalizuje przekształcenie
glukozy w glukozo-6-fosforan.
•
Znacznie więcej fruktozy jest przekształcane w fruktozo-1-fosforanu
jest następnie rozszczepiana na fosforan dihydroksyacetonu i
aldehyd glicerynowy.
•
Aldehyd glicerynowy jest fosforylowany i wraz z fosforanem
dihydroksyacetonu wchodzi w szlak przemian glukozy.
•
Ponieważ reakcje przebiegające w trakcie fosforylacji fruktozy w
pozycji 1 atomu węgla mogą zachodzić z normalną szybkością, pod
nieobecność insuliny, zaleca się podawanie fruktozy chorym na
cukrzycę dla uzupełnienia zapasów węglowodanów.
•
Fruktoza jest przekształcania częściowo w fruktozo-6-fosforan, a
następnie metabolizowana, przez fruktozo – 1,6-difosforan (
slajd – 6
).
•
Enzym katalizujący powstawanie fruktozo-6-fosforanu to
heksokinaza – ten sam enzym, który katalizuje przekształcenie
glukozy w glukozo-6-fosforan.
•
Znacznie więcej fruktozy jest przekształcane w fruktozo-1-fosforanu
jest następnie rozszczepiana na fosforan dihydroksyacetonu i
aldehyd glicerynowy.
•
Aldehyd glicerynowy jest fosforylowany i wraz z fosforanem
dihydroksyacetonu wchodzi w szlak przemian glukozy.
•
Ponieważ reakcje przebiegające w trakcie fosforylacji fruktozy w
pozycji 1 atomu węgla mogą zachodzić z normalną szybkością, pod
nieobecność insuliny, zaleca się podawanie fruktozy chorym na
cukrzycę dla uzupełnienia zapasów węglowodanów.

Metabolizm heksoz innych niż
glukoza
Metabolizm heksoz innych niż
glukoza
•
Większość fruktozy jest metabolizowana w jelicie i w
wątrobie, z zatem jej znaczenie w zaopatrywaniu w
węglowodany w ustroju ograniczone.
•
Fruktozo-6-fosforan może także być fosforylowany przy 2
atomie węgla, tworząc fruktozolatorem glukoneogenezy
zachodzącej w wątrobie.
•
Kiedy stężenie fruktozo – 2,6-difosforanujest duże,
przekształcenie fruktozo-6-fosforanu i wzrasta dzięki temu
rozpad glikozy do pirogronianu.
•
Zmniejszenie się stężenia fruktozo-2,6-difosforanu ułatwia
reakcję w odwrotnym kierunku i wspomaga w następstwie
glukoneogenezę.
•
Jedną z funkcji kinazy białkowej, wytworzonej w wyniku
działania glukagonu, jest zmniejszenie stężenia fruktozo-
2,6-difosforanu w wątrobie.
•
Większość fruktozy jest metabolizowana w jelicie i w
wątrobie, z zatem jej znaczenie w zaopatrywaniu w
węglowodany w ustroju ograniczone.
•
Fruktozo-6-fosforan może także być fosforylowany przy 2
atomie węgla, tworząc fruktozolatorem glukoneogenezy
zachodzącej w wątrobie.
•
Kiedy stężenie fruktozo – 2,6-difosforanujest duże,
przekształcenie fruktozo-6-fosforanu i wzrasta dzięki temu
rozpad glikozy do pirogronianu.
•
Zmniejszenie się stężenia fruktozo-2,6-difosforanu ułatwia
reakcję w odwrotnym kierunku i wspomaga w następstwie
glukoneogenezę.
•
Jedną z funkcji kinazy białkowej, wytworzonej w wyniku
działania glukagonu, jest zmniejszenie stężenia fruktozo-
2,6-difosforanu w wątrobie.
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
Wyszukiwarka
Podobne podstrony:
Węglowodory aromatyczne (2) ppt
01 Wybrane zaburzenia gospodarki węglowodanowejid 2675 ppt
Lekcja II Witaminy jako skladniki koenzymow i ich udzial w przemianiach weglowodanow lipidow i biale
3 Metabolizm Przemiana białek
Biochemia wykład 13 Metabolizm węglowodanów
Metabolizm węglowodanów
Zaburzenia przemian metabolicznych
17 Wybrane aspekty metabolizmu ostatnie osiągnięcia naukiid 17370 ppt
Powiązanie metabolizmu weglowodanow i tluszczy w normie i patologii
42 Moc elektryczna w obwodach prądu przemiennego ppt
Zaburzenia przemian metabolicznych
11 Procesy i przemianyid 12239 ppt
więcej podobnych podstron