Skurcz komórki mięśnia
poprzecznie prążkowanego
Błona komórki mięśniowej
Depolaryzacja błony
przekraczająca wartość
progową
Potencjał
czynnościowy
Synapsy nerwowo-mięśniowe
Inne komórki mięśniowe
Pobudzenie
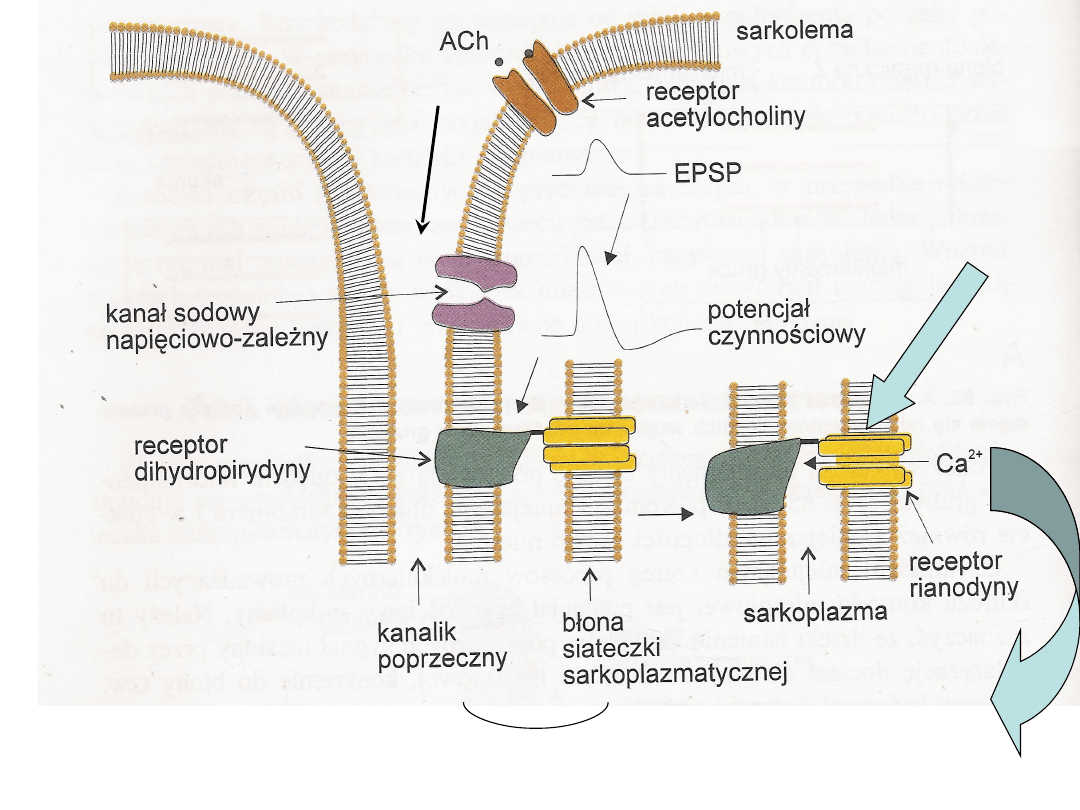
RR –
kanał
wapniow
y
sprzężenie
depolaryzacja
Wzrost
stężenie
wapnia
w
komórce
Jony wapnia
Łączą się z podjednostkami C
troponiny, wchodzącej w skład
miofilamentów cienkich
Procesy metaboliczne
Połączenie cząsteczki aktyny z
główką miozyny
Przesunięcie miofilamentu
cienkiego względem grubego
Zwiększenie
powinowactwa miozyny do
ATP
Połączenie miozyny z ATP
Rozłączenie kompleksu miozyna- aktyna
Hydroliza ATP
Rozpada się kompleks troponina i jony
wapnia
Regulacja siły skurczu mięśnia
sercowego
Siła skurczu
Poziom wolnych
jonów wapnia w
sarkoplazmie
Ilość wiązań miozyna-aktyna
Czynniki zwiększające
ilość wolnych jonów
wapnia w sarkoplazmie
będą zwiększały siłę
skurczu zaś czynniki
obniżające to stężenie
będą osłabiały skurcz.
CZYNNIKI
INOTROPOWE
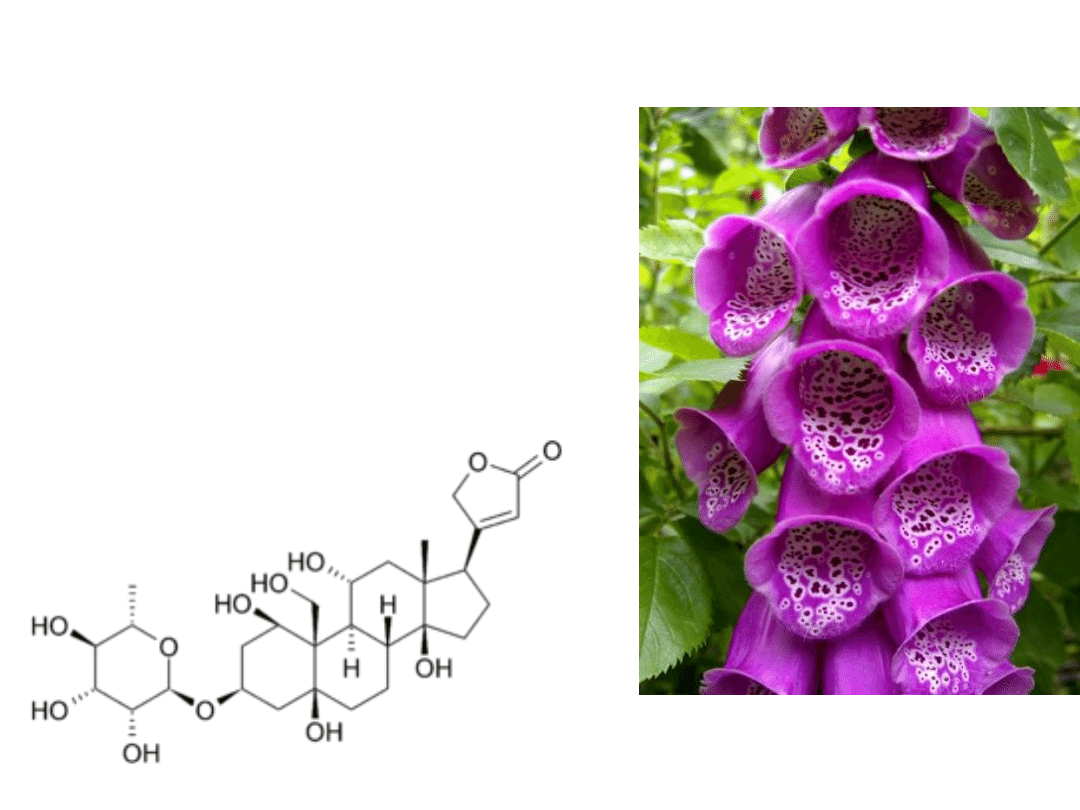
Glikozydy nasercowe
- np. z naparstnicy
- Wykazują dodatnie działanie
inotropowe- zwiększają siłę
skurczu.
- Hamują działanie pompy sodowo-
potasowej
- Przykładem jest strofantyna G
Glikozydy nasercowe
Blokują zewnątrzkomórkowe
miejsce wiązania jonów
Proces bardzo szybki i odwracalny.
Umożliwia poprzez znakowaną
radioaktywnie strofantynę oznaczyć
pompy sodowo-potasowe.
Zmniejszenie różnicy
potencjałów
elektrochemicznych miedzy
wnętrzem a zewnętrzem
komórki.
Zmniejszenie dokomórkowego
transportu jonów sodu z którym
sprzężony jest odkomórkowy
transport jonów wapnia.
Zwiększenie stężenia
wewnątrzkomórkowego jonów sodu oraz
stężenia wewnątrzkomórkowego jonów
wapnia
Zwiększenie prawdopodobieństwa
połączenia jonów wapnia z
podjednostką C troponiny.
WZROST SIŁY SKURCZU MIĘŚNIA
Zewnątrzkomórkowy
sygnał chemiczny
inicjujący skurcz
sarkomeru
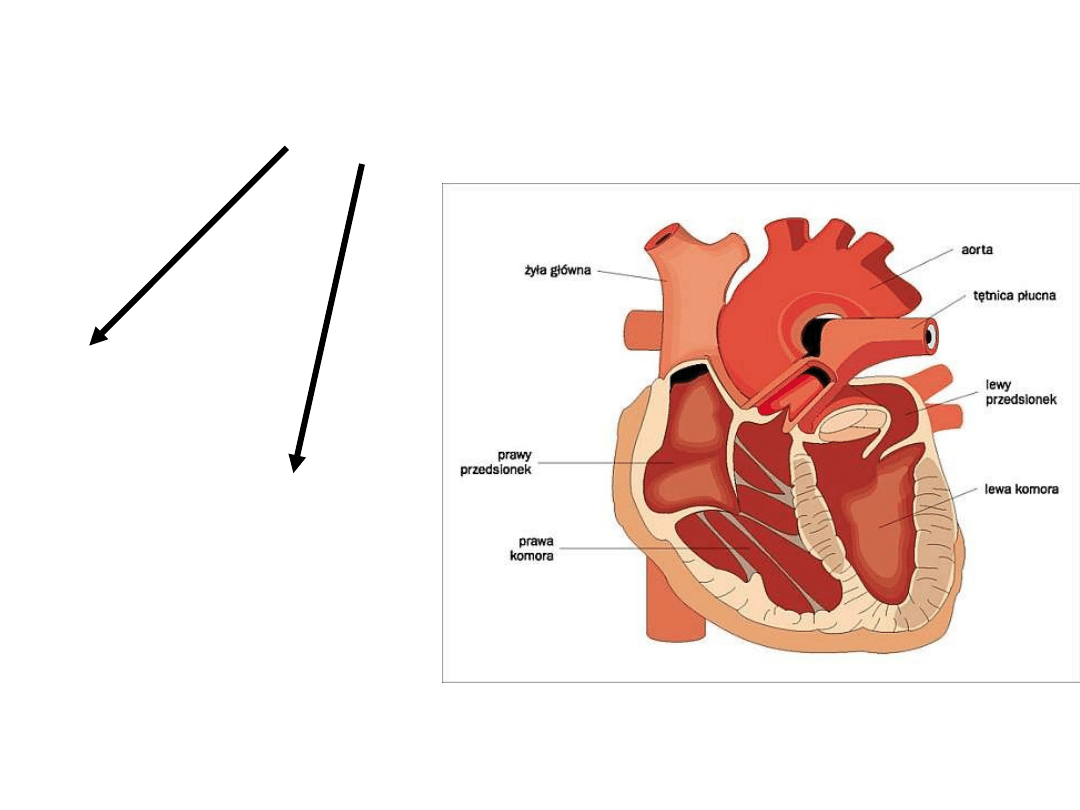
Komórki robocze serca
Mięśnie komór
Mięśnie
przedsionkó
w
Potencjał czynnościowy
mięśniowej komórki roboczej
serca
Bodziec z węzła
zatokowo-
przedsionkowego
spolaryzowana komórka
mięśnia sercowego
zmiana
przepuszczalności
błony dla jonów sodu
Zmniejszenie
potencjału do wartości
ok. –65 mV (potencjał
progowy)
depolaryzacja
depolaryzacja
Przekroczenie potencjału progowego
Otwarcie kanałów
sodowych - gwałtowny
napływ jonów sodu do
wnętrza komórki
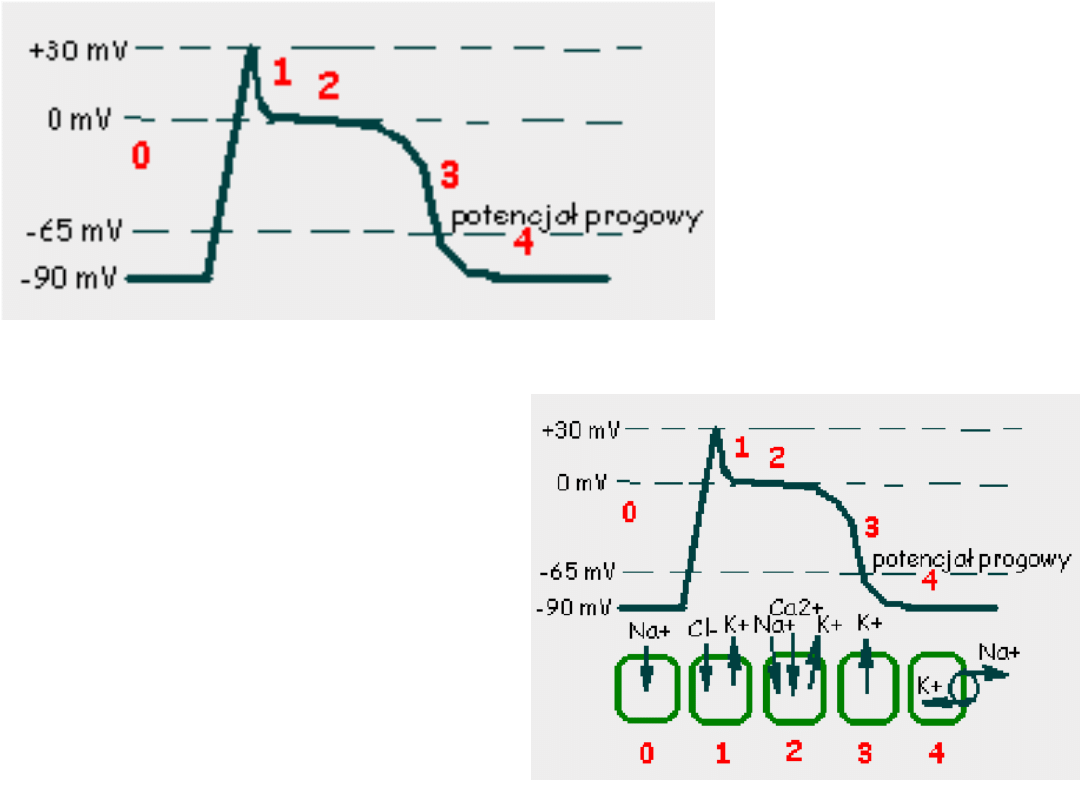
Faza 0 –potencjał błonowy przekracza
wartość progową, dochodzi do
otwarcia
napięciowo-
zależnych
kanałów sodowych
.
Następuje
gwałtowna
depolaryzacja błony
spowodowana
napływem jonów
sodu.
Do wnętrz komórki zaczyna
płynąć prąd chlorkowy.
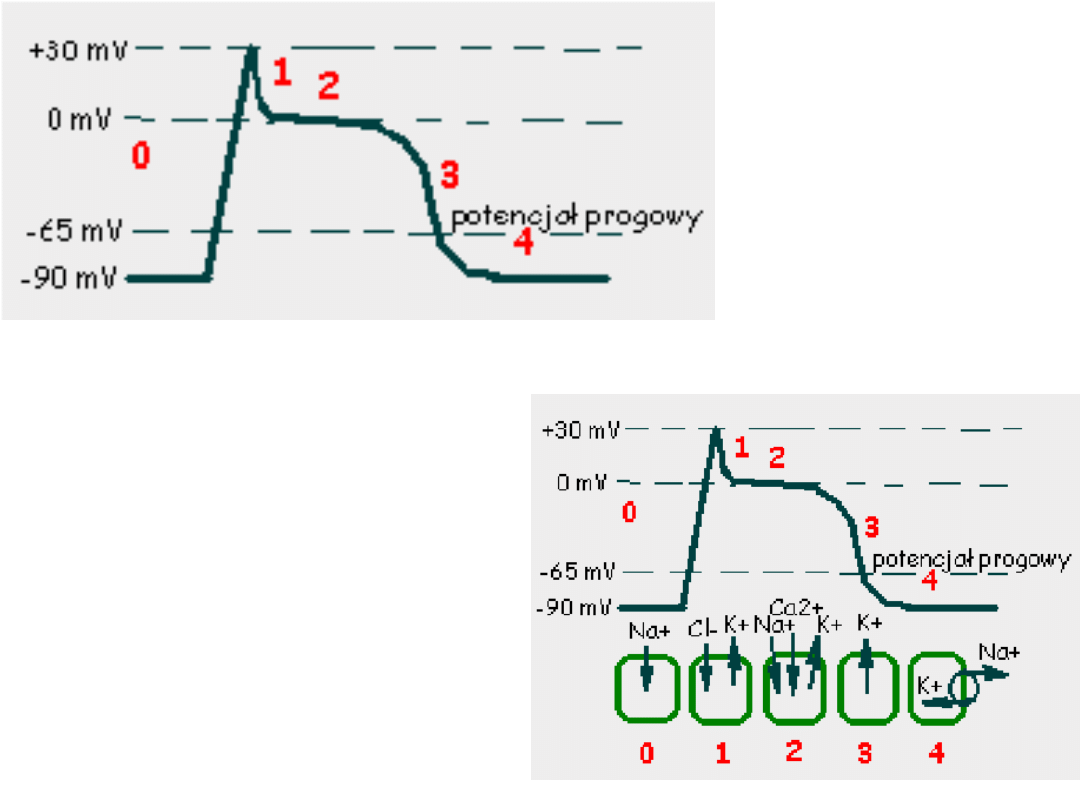
Faza 1 -obecność prądu chlorkowego
i
zmniejszenie prądu sodowego
powoduje spadek potencjału
błonowego.
Zbytniemu spadkowi
potencjału
przeciwdziała
dokomórkowy prąd
wapniowy –
płynie przez
aktywowane
napięciem kanały
wapniowe
typu L.
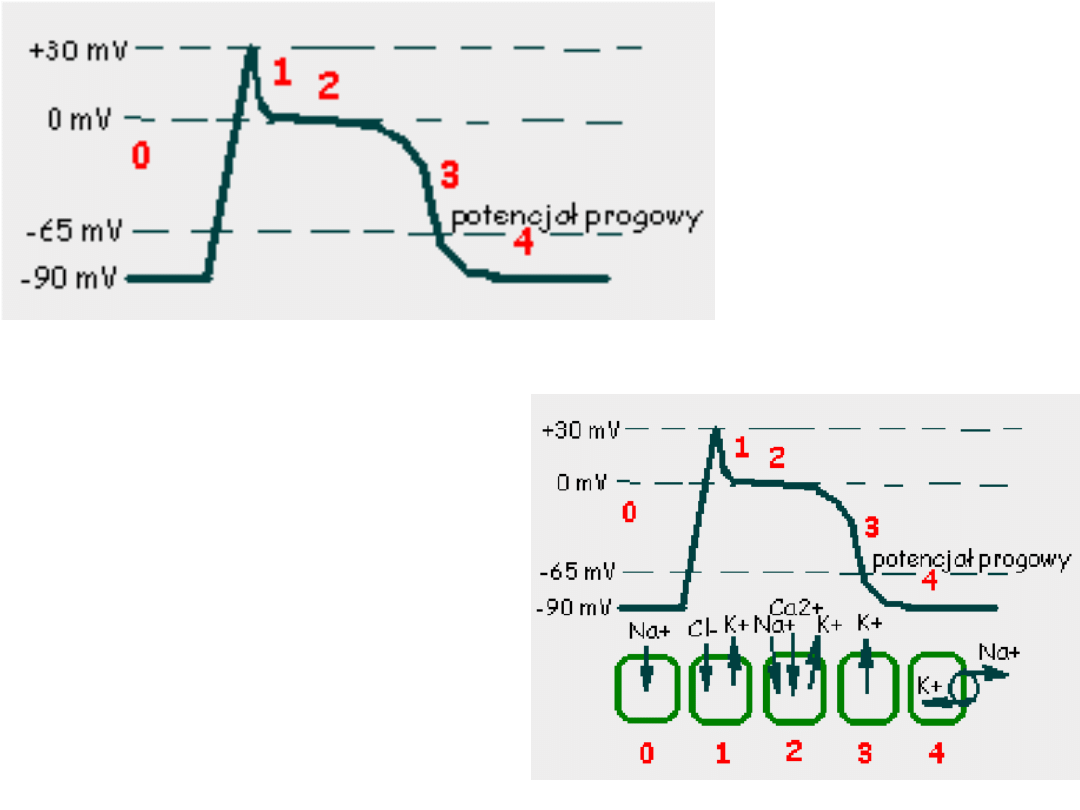
Faza 2 –płynący prąd wapniowy
równoważy
prąd chlorkowy i
potasowy dzięki
czemu przez
około 150ms
potencjał błonowy
pozostaje na
stałym poziomie.
Pod koniec fazy 2 kanały
wapniowe ulegają inaktywacji.
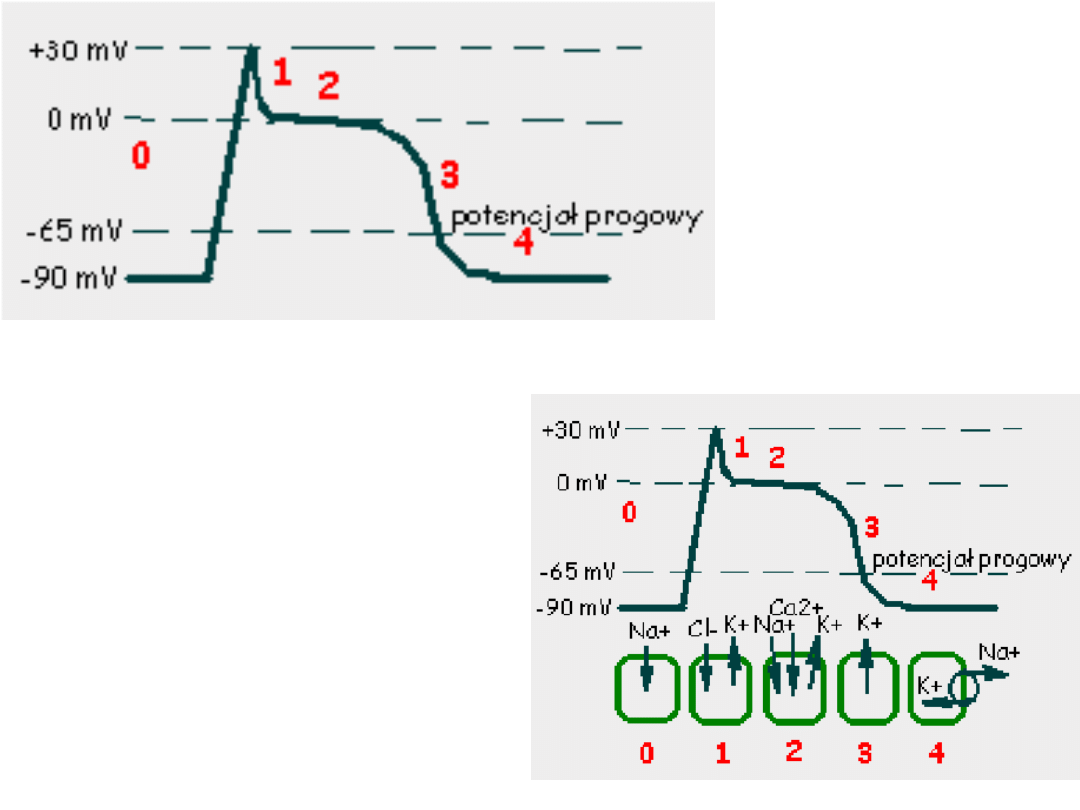
Faza 3 –rosnąca przewaga prądów
repolaryzacyjnych
(chlorkowego i potasowego)
powoduje
repolaryzacje błony do
wartości
zmierzającego do
-90mV.
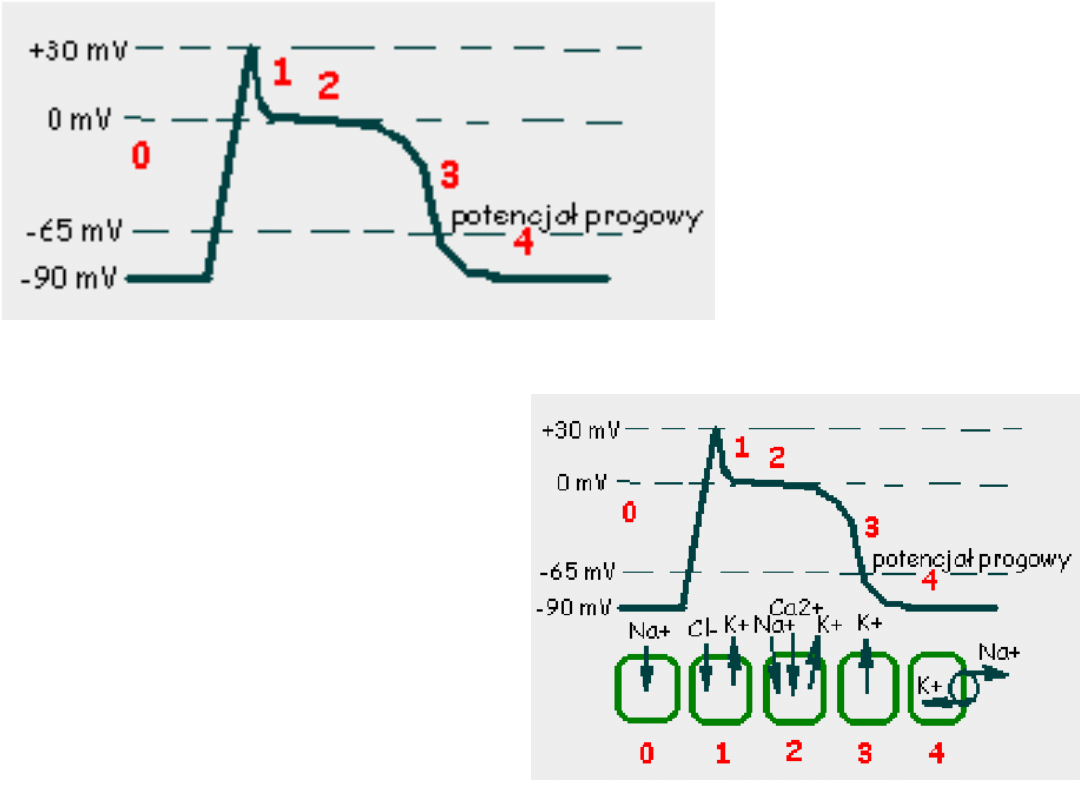
Faza 4 – następuje w momencie
osiągnięcia przez błonę
potencjału
spoczynkowego.

Cechy pobudzenia komórki
roboczej mięśnia sercowego:
- Dużo większy stosunek czasu
depolaryzacji do czas spoczynku niż w
komórkach nerwowych warunkuje
ograniczony przepływ jonów –
oszczędność energii która musiałaby
być spożytkowana na transport
aktywny w celu wyrównania stężeń.
- Małemu przepływowi jonów sprzyja
mała ilość kanałów jonowych w
sarkolemie

-
Prąd potasowy w komórce roboczej jest większy
w stanie spoczynku niż w stanie
zdepolaryzowanym. Odpowiedzialne są za to:
a) Prostownicze dokomórkowe kanały potasowe K
ir
–zamykają się przy potencjale błonowym
równym potencjałowi spoczynkowemu a
otwierane są przez hiperpolaryzację.
b) Opóźnione kanały prostownicze Ka
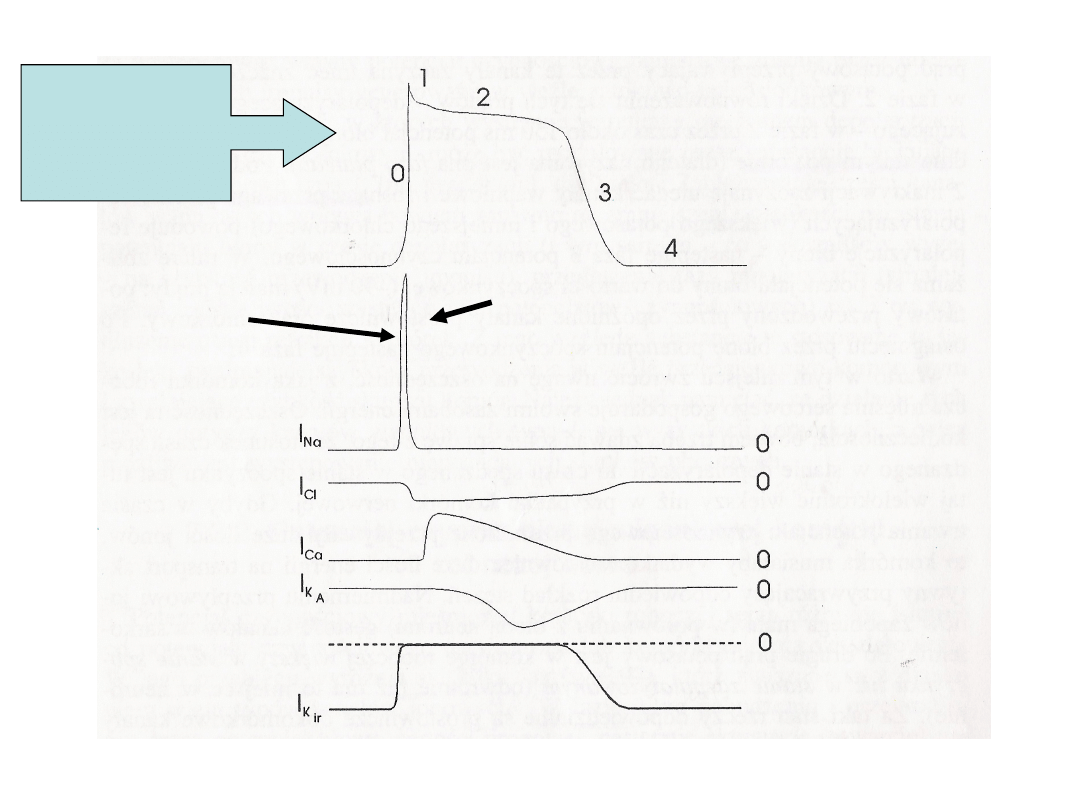
Fazy
potencjału
czynnościoweg
o komórki
roboczej serca
Prąd
depolaryzacyjn
y
Kanały sodowe otwarte -
faza 0 i 1, faza 2,3
–stan inaktywacji- niemożliwe jest
następne pobudzenie komórki –OKRES
REFRAKCJI BEZWZGLĘDNEJ, koniec fazy 3
i 4 następuje OKRES REFRAKCJI
WZGLĘDNEJ.
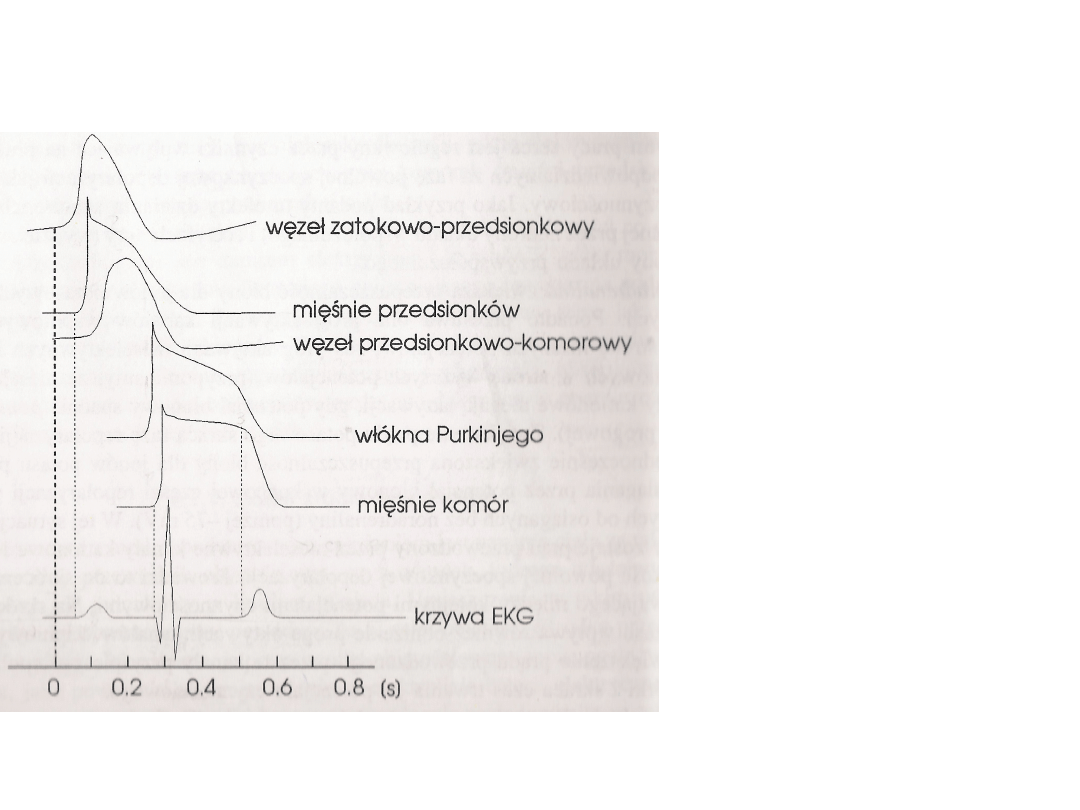
Porównanie przebiegów potencjałów
czynnościowych różnych typów komórek
Włókna odpowiedzialne za
przewodzenie impulsów
mają stosunkowo wolną
falę depolaryzacyjną.
Brak fazy plateau w
potencjałach
czynnościowych komórek
przewodzących skraca
czas trwania potencjałów.

RYTM PRACY SERCA
Noradrenalina
Noradrenalina –
zwiększa
przepuszczalność błon
dla jonów potasu i
wapnia, przesuwa próg
aktywacji kanałów
wapniowych typu L w
stronę niższych
potencjałów a próg
aktywacji
nieselektywnych
kanałów kationowych w
stronę wyższych
potencjałów.
Acetylocholina
Acetylocholina –wpływa na
kanały jonowe odpowiedzialne
za częstotliwość generacji
potencjałów czynnościowych,
działa antagonistycznie do
noradrenaliny, zmniejszając
częstotliwość potencjałów
czynnościowych komórek.
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
Wyszukiwarka
Podobne podstrony:
Środki zwiotczające mięśnie poprzecznie prążkowane
Fizjologia, fizjol cw, Budowa mięśnia poprzecznie prążkowanego
Budowa mięśnia poprzecznie prążkowanego, weterynaria, Anatomia
Diagnostyka laboratoryjna chorób serca i mięśni poprzecznie prążkowanych
wpływ wysiłku fizycznego na mieśnie poprzecznie prązkowane 2
FIZJOLOGIA CZŁOWIEKA ĆWICZENIA, Fizjologia mini poprzecznie prkowanych, Fizjologia mięśni poprzeczni
fizjologia miesni poprzecznie prazkowanych, Studia, Neurobiologia
wpływ wysiłku fizycznego na mieśnie poprzecznie prązkowane 6
wpływ wysiłku fizycznego na mieśnie poprzecznie prązkowane 3
wpływ wysiłku fizycznego na mieśnie poprzecznie prązkowane 1
WPŁYW WYSIŁKU FIZYCZNEGO NA MIĘŚNIE POPRZECZNIE PRĄŻKOWANE
1-3a Zanik mięśnia poprzecznie prążkowanego, Naika, stomatologia, Patomorfologia, LEKARSKI, Patomorf
Fizjologia mięśni poprzecznie prążkowanych
Środki zwiotczające mięśnie poprzecznie prążkowane
15 sarkomery w komorkach miesoniowych poprzecznie prazkowanych i gladkich roznice ultrastrukturalne
Środki zwiotczające mięśnie poprzecznie prążkowane
Budowa komórki mięśniowej i skurcz mięśnia
więcej podobnych podstron