
Wykład 4
Metabolizm węgla
Fotosynteza i
fotooddychanie
1

Metabolizm węgla
• Bilans reakcji świetlnych fotosyntezy
• Wiązanie i redukcja CO
2
(rośliny C3, C4, CAM)
• Aktywność fotosyntetyczna roślin (woda, światło,
CO
2
, temperatura)
• Fotooddychanie
2
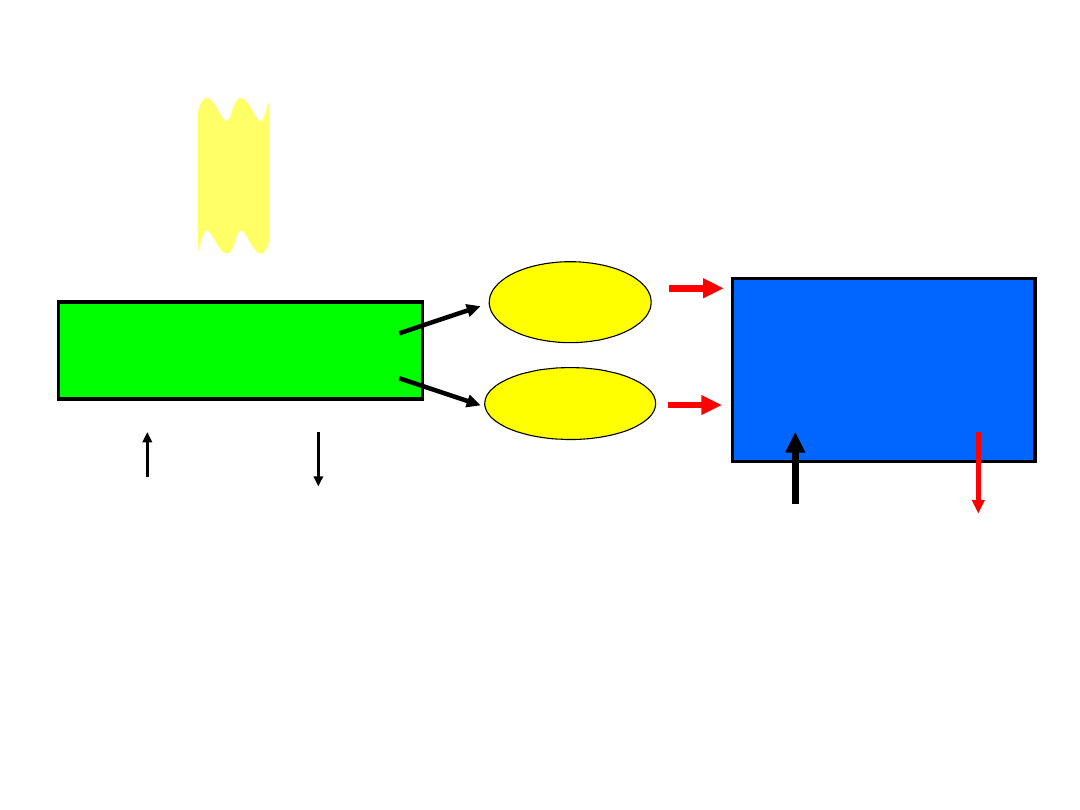
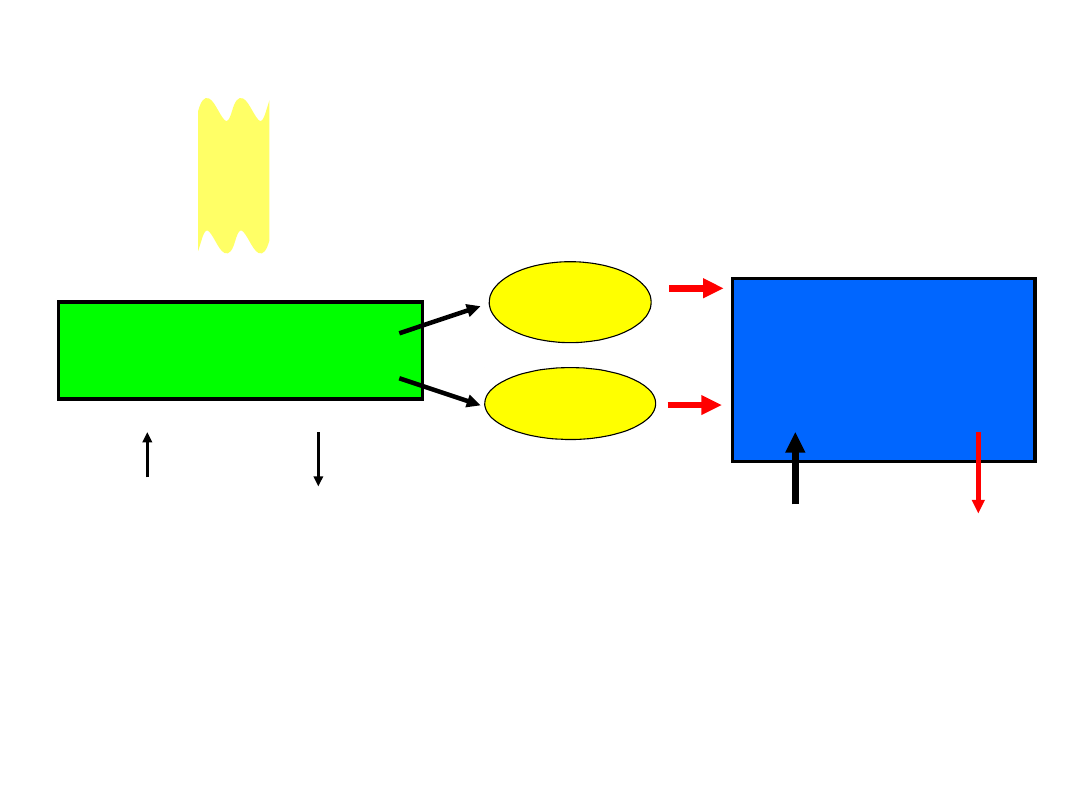
Reakcje świetlne
błony tylakoidów
ATP
NADP
H
Reakcje
węgla
[ciemniowe]
enzymy stromy
H
2
O
O
2
CO
2
CH
2
O
cuk
ry
światło
Reakcje świetlne fotosyntezy
i
reakcje wiązania
węgla
zachodzą w oddzielnych częściach chloroplastu
3

Fotony (kwanty światła)
są absorbowane przez
barwniki fotosyntetyczne
, co oznacza przekształcenie
energii świetlnej fotonów w energię wzbudzenia
elektronowego cząsteczek barwników. Energia
wzbudzenia dociera do specjalnych cząsteczek
chlorofilu
a
, określanych jako P680 (PSII) i P700 (PSI), będących
pierwotnymi donorami elektronów dwóch funkcjonalnych
kompleksów białkowo-barwnikowych - PSI i PSII
.
CHLOROFIL
to barwnik zbudowany z tetrapirolowego
pierścienia (podobnego do porfiryny) z centralnie
umieszczonym atomem
Mg
. Dołączony jest do niego (C-
20) 20-węglowy, hydrofobowy łańcuch zwany „ogonem
fitolowym”, który kotwiczy cząsteczkę chlorofilu w błonie
tylakoidu.
Tylakoidy roślin i glonów zawierają chlorofil a i b.
Chlorofil a
- w centrach aktywnych PSI i PSII i w
antenach
Chlorofil b
- jedynie w kompleksach antenowych
4
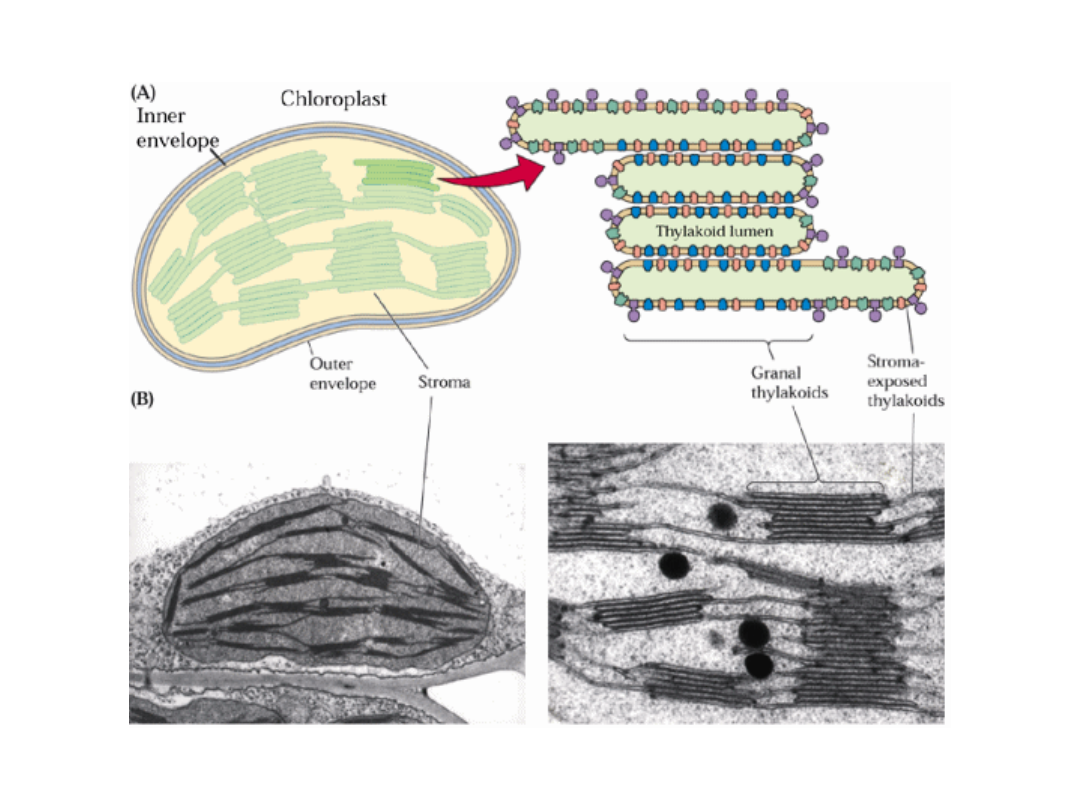
5
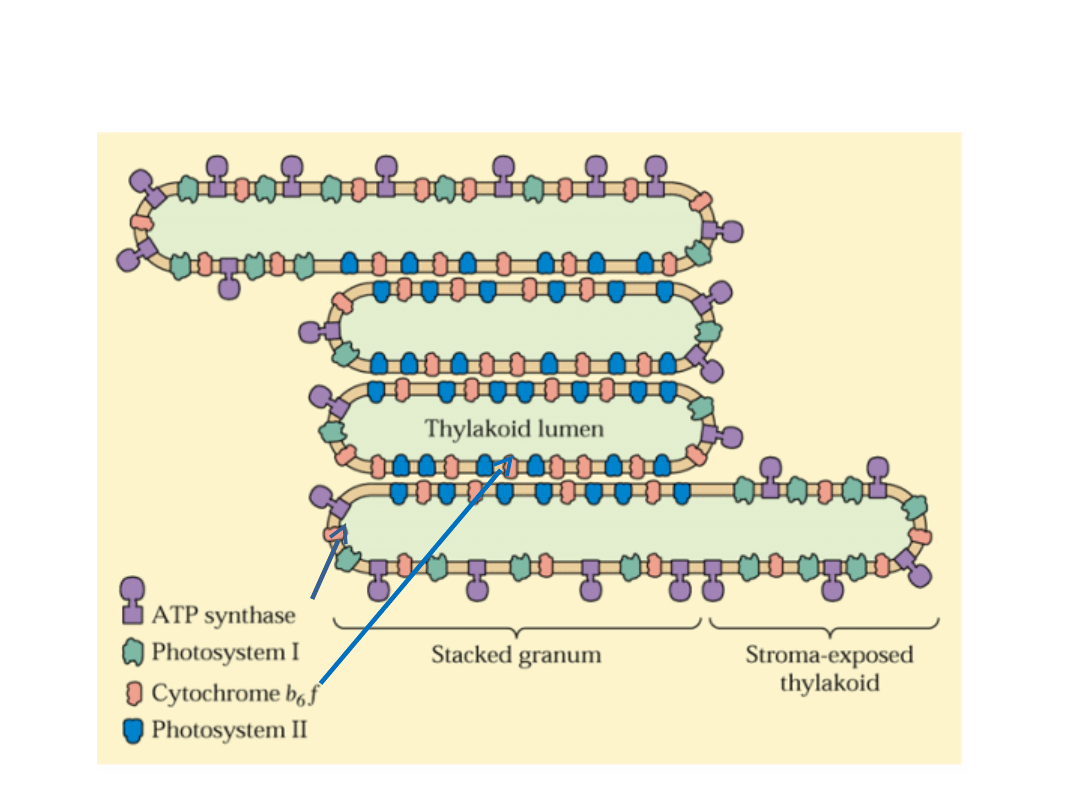
Przestrzenne rozdzielenie PSI i PSII w błonach tylakoidów umożliwia
sprawne regulowanie absorpcji światła przez obydwa fotosystemy
oraz zapobiega fotoinhibicji PSII.
6
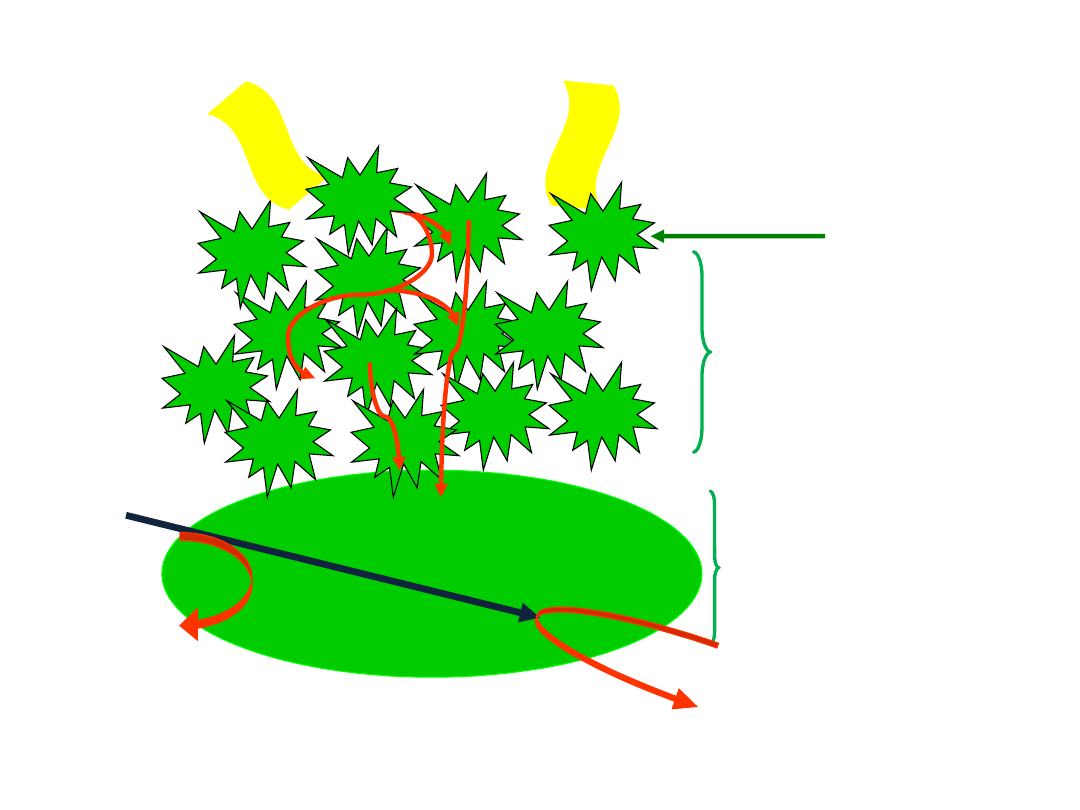
Schemat budowy fotosystemu - PSI lub PSII
Q-
Cząsteczki chlorofilu absorbują fotony, energia wzbudzenia
przekazywana jest do centrum reakcji, w którym zachodzą reakcje redox (utleniania i redukcji)
akceptor
elektronów
anteny
centrum reakcji
A
A+
Q
donor
elektronów
cząsteczki
chlorofilu
h
ē
h
h
h
h
h
h
7
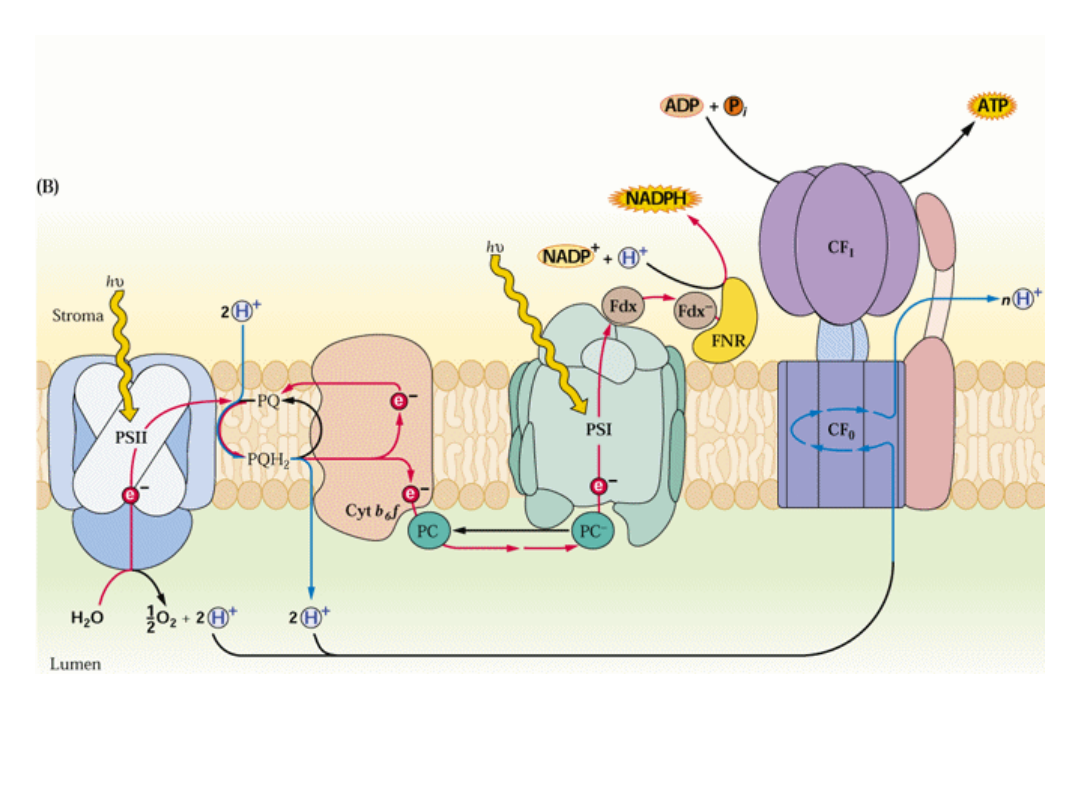
Budowa i rozmieszczenie kompleksów białkowych
fotosyntetycznego transportu elektronów w błonie tylakoidów
Utlenienie wody i redukcja NADP zachodzą po przeciwnych stronach błony tylakoidu
Protony zgromadzone po stronie lumen błony tworzą gradient niezbędny do syntezy ATP
(fosforylacja niecykliczna), jednocześnie zachodzi redukcja NADP
+
.
zmod. Fig. 12.22. Buchanan 2000
8
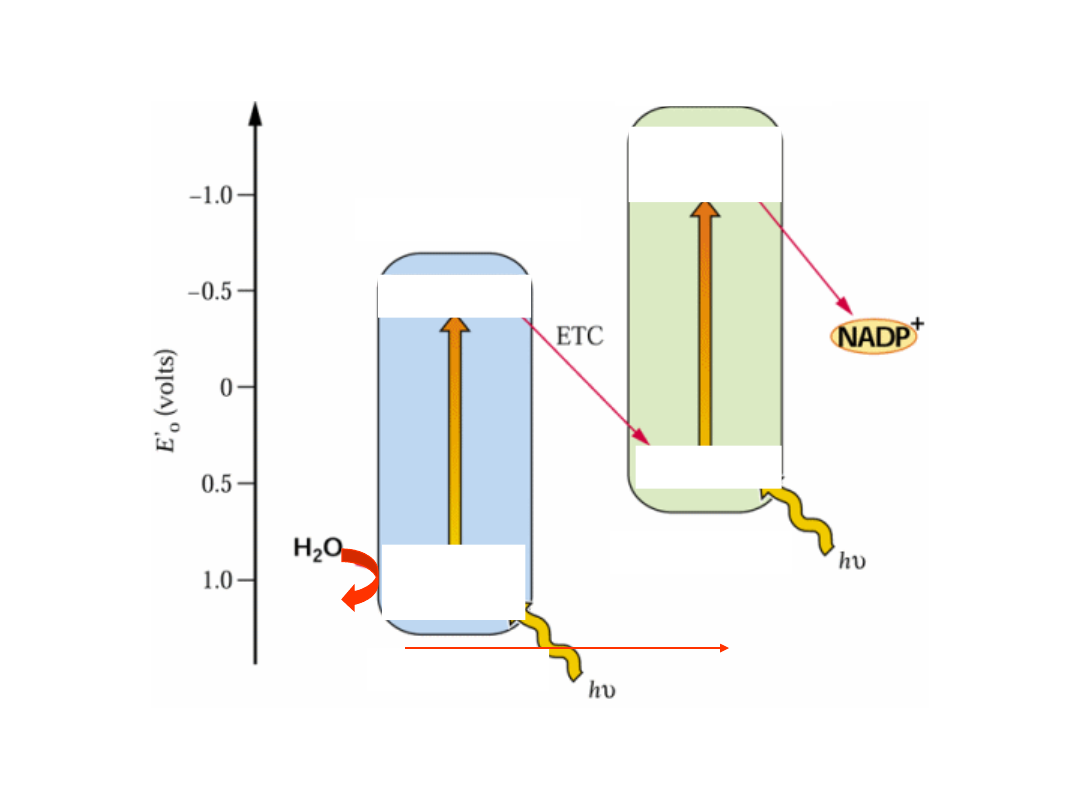
Schemat układu „Z” przenośników Fotosystemu (PS) II i I
Na świetle PS II wytwarza silny utleniacz (utleniający wodę) oraz reduktant.
Pod wpływem światła PS I wytwarza silny reduktant (redukujący NADP
+
) oraz
słaby utleniacz.
ETC przekazuje elektrony z PSII (z reduktanta) na PSI (na utleniacz).
Silny
utleniacz
reduktant
Silny
reduktant
Utleniacz
Fotosystem II
Fotosystem I
½ O
2
+ 2H
+
+
_
przepływ elektronów
Poziom energetyczny
wysoki
niski
e
-
e
-
9
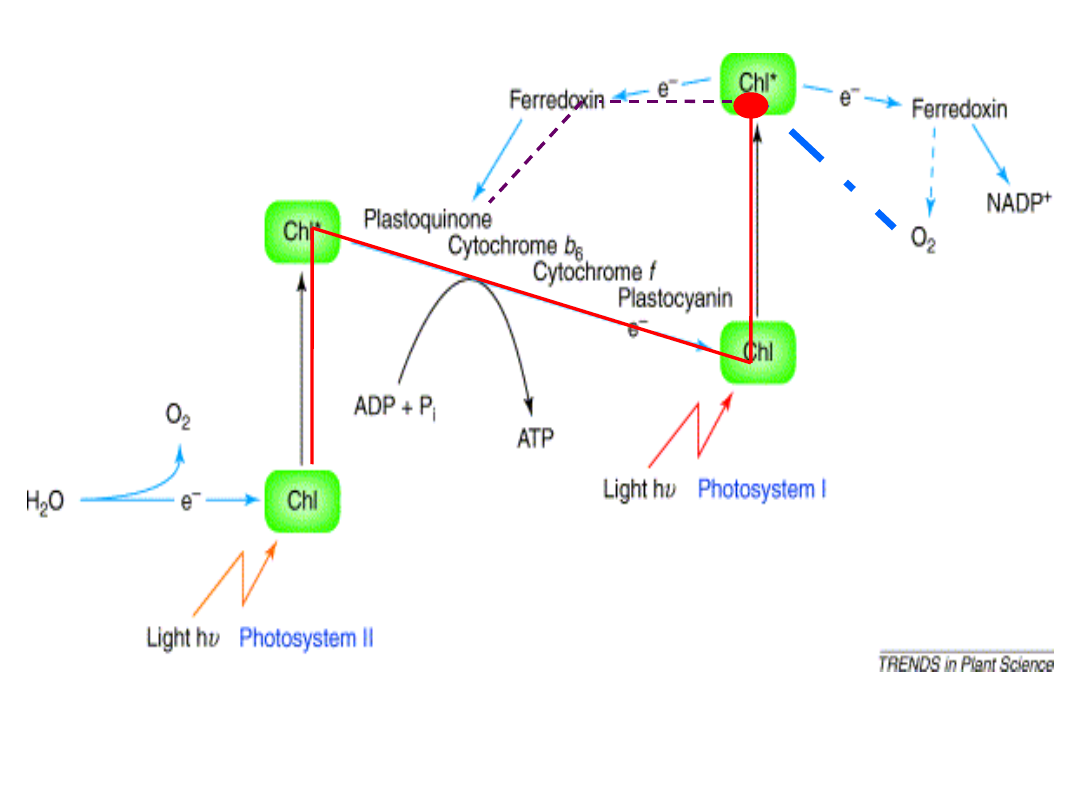
Z-schemat fotosyntezy -fotofosforylacja niecykliczna - synteza ATP i
redukcja NADP
+
fotofosforylacja cykliczna – tylko synteza ATP
fotofosforylacja pseudocykliczna – tylko synteza ATP
1 0
O
2
2 O
2
.-
H
2
O
2
2H
2
O
Ferredoksyn
a
PSI
2H
+
, 4ē,
O
2
2H
+
,
SOD
O
2
APX
Transport
elektronów
PSII
4H
+
2
askorbinian
2
monodehydroaskorbini
an
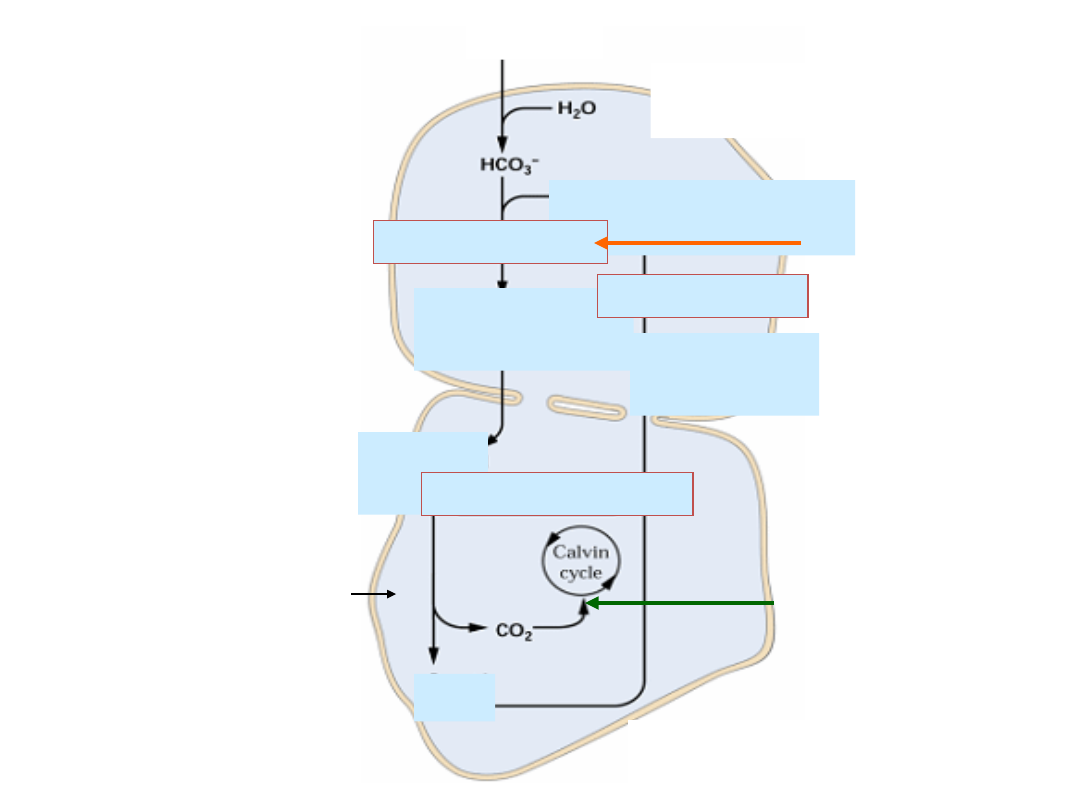
Fosforylacja pseudocykliczna - REAKCJA
MEHLERA
- cykl woda-woda
1 1
Reakcje świetlne
błony tylakoidów
ATP
NADP
H
Reakcje
węgla
[ciemniowe]
enzymy stromy
H
2
O
O
2
CO
2
CH
2
O
cuk
ry
światło
Reakcje świetlne fotosyntezy
i
reakcje wiązania
węgla
zachodzą w oddzielnych częściach chloroplastu
3
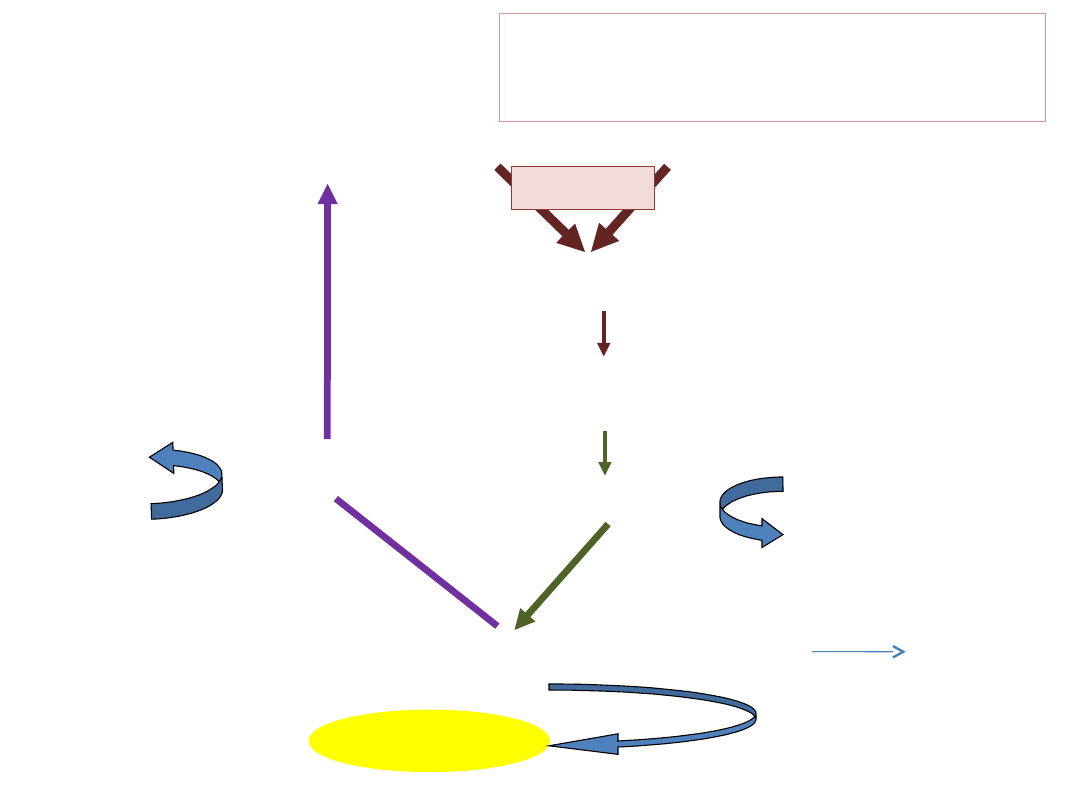
Rybulozo-1,5-bisfosforan
(RuBP) (6C5)
6CO
2
KARBOKSYLACJA
3-fosfoglicerynian PGA (12 x C3)
REDUKCJA
ATP, NADPH
ADP+Pi, NADP
aldehyd 3-fosfoglicerynowy
(2 x C3)
ASYMILATY
REGENERACJ
A
ADP+P
i
ATP
Zasadnicze etapy cyklu C3
6C1+6C5=12C3
[2C3=ZYSK NETTO FOTOSYNTEZY]
[10C3=6C5]
(10C3)
(6C5)
RUBISCO
heksoza
1 3
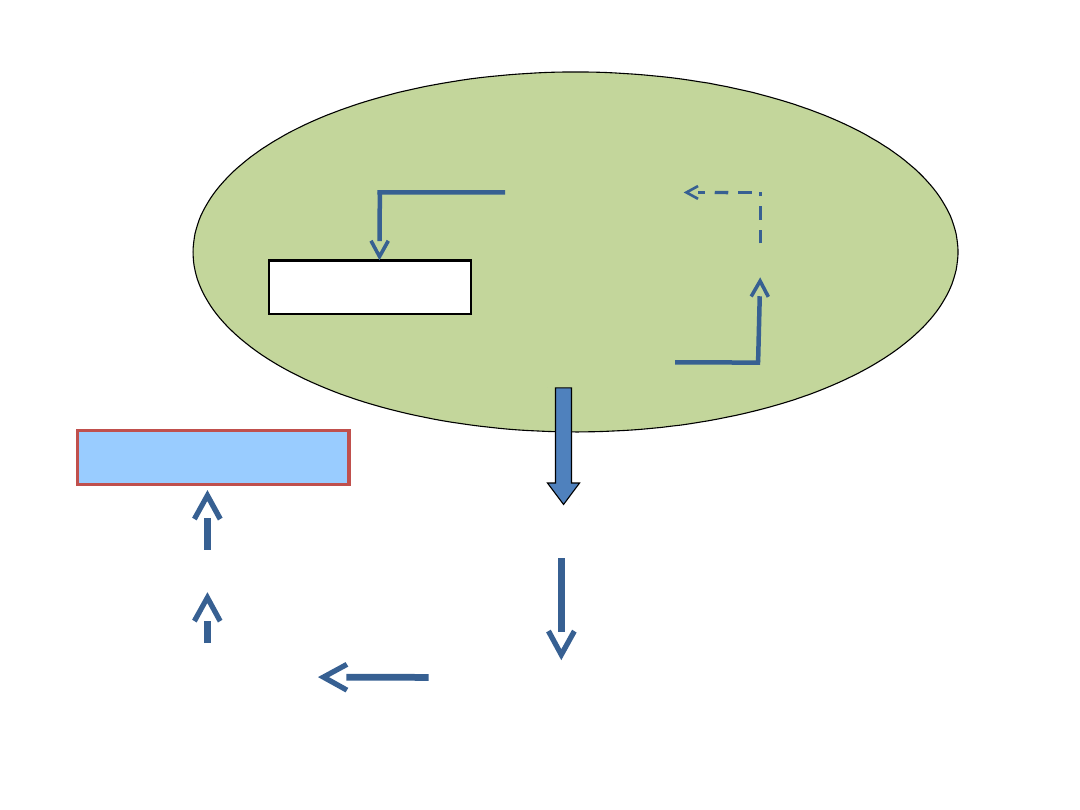
Chloroplast
FOSFOTRIOZY
1,6-bisfosfofruktoza
ADP-glukoza
SKROBIA
1,6-bisfosfofruktoza
Cytosol
SACHAROZA
6-fosfofruktoza
6-
fosfosachar
oza
UDP-
glukoza
Synteza sacharozy zachodzi w cytosolu
Synteza skrobi jest na terenie chloroplastu
Synteza sacharozy i skrobi
1 4

Aktywność fotosyntetyczna roślin
• Światło
(świetlny punkt kompensacyjny, rośliny światło-
i cieniolubne)
• CO
2
(stężenie kompensacyjne, stężenie CO
2
może
ograniczać fotosyntezę przy silnym oświetleniu)
• Temperatura
(może ograniczać fotosyntezę przy wysyceniu
światłem i dobrym zaopatrzeniu w CO
2
)
• H
2
O
(zamknięcie aparatów szparkowych)
1 5
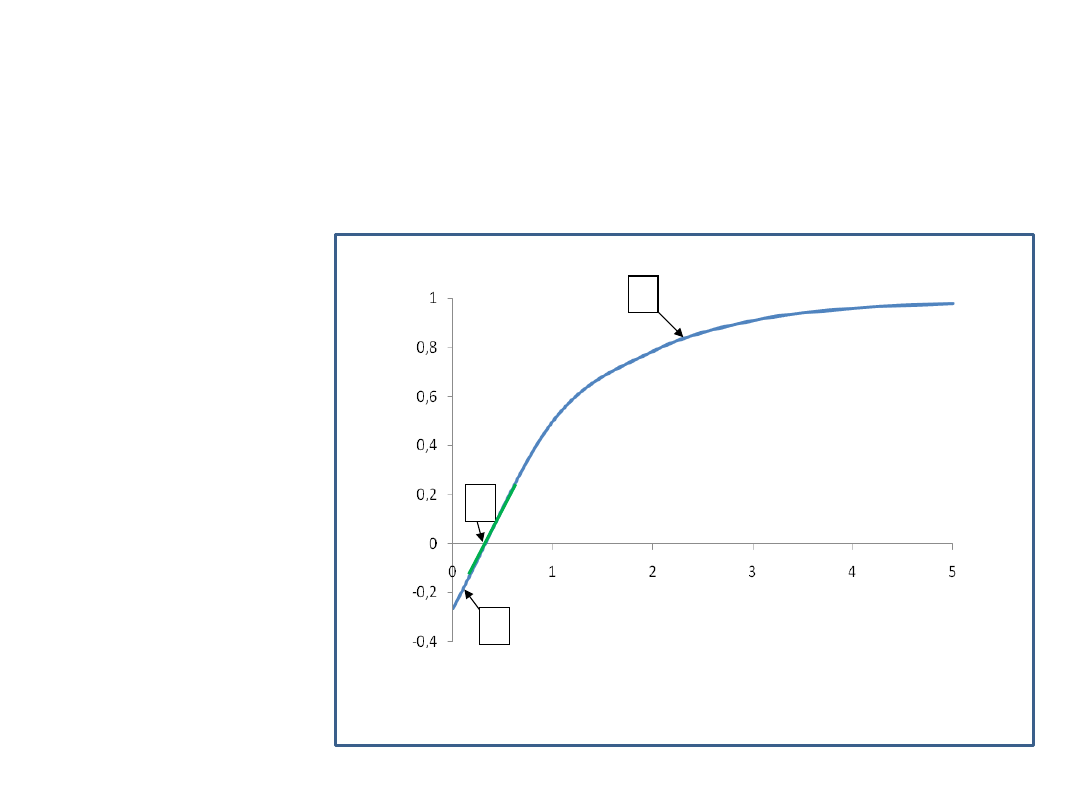
Wpływ natężenia światła na fotosyntezę
Tylko 5% energii wykorzystuje fotosynteza, gdyż barwniki absorbują tylko
w zakresie pomiędzy 400 a 700 nm (PhAR)
Po
b
ie
ra
n
ie
C
O
2
Natężenie światła
W
y
d
zi
e
la
n
ie
C
O
2
1
2
3
[jednostki
umowne]
Krzywa świetlna fotosyntezy
–
zmiany intensywności fotosyntezy,
mierzone jako pobieranie CO
2
w zależności od natężenia światła.
1 - oddychanie komórkowe
(oddychanie przeważa nad fotosyntezą)
2 -
świetlny punkt kompensacyjny
(zrównoważenie procesów)
3 – punkt wysycenia fotosyntezy
zależne od gatunku
1 6
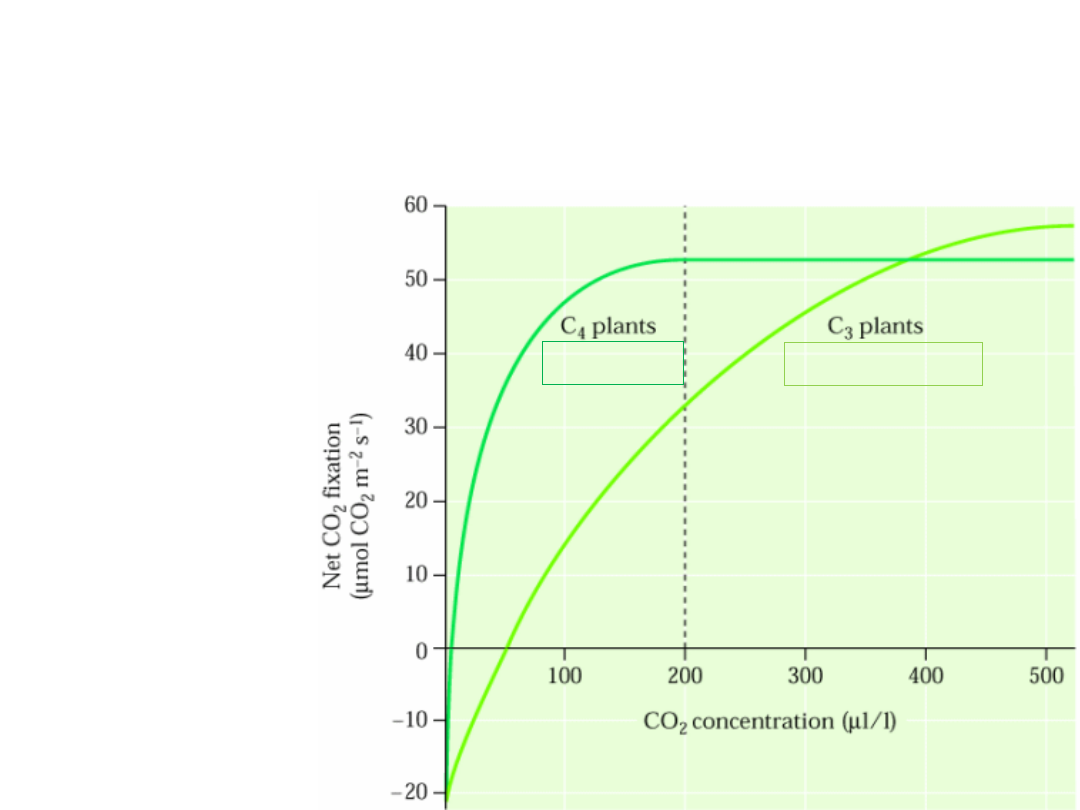
Wpływ stężenia CO
2
w atmosferze na
asymilację CO
2
u roślin
punkt kompensacyjny CO
2
=
stężenie
CO
2
,
przy którym wiązanie CO
2
osiąga wartość 0 przy
wysycającym świetle
wiązanie CO
2
w trakcie fotosyntezy jest zrównoważone przez jego
wydzielanie podczas oddychania
*
0-5 µl/l
*
20-100 µl/l
C
4
– saturacja wiązania CO
2
przy ok. 200 µl/l CO
2
C
3
– saturacja wiązania CO
2
przy ok. 400 µl/l CO
2
* *
zmod. Fig. 14.44. Buchanan 2000
1 7
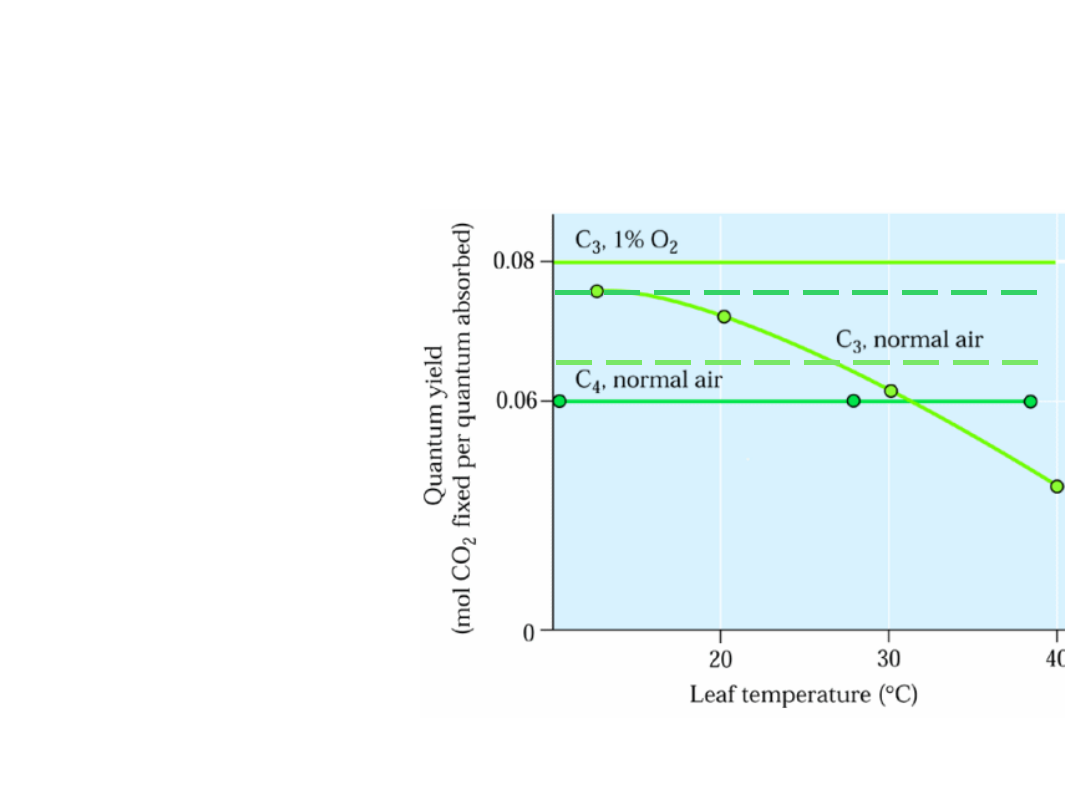
Wpływ temperatury na wydajność kwantową
fotosyntezy
u roślin C3 i C4
Wydajność kwantowa () = mol CO
2
/ kwant absorbowanego światła
C
4
– 21% O
2
wydajność kwantowa stała i
niezależna
od temp. =
brak
fotooddychania
C
3
–
21% O
2
fotooddychanie
wzrost fotooddychania
wraz ze wzrostem temp.
spadek wydajności kwantowej
-1% O
2
ograniczenie fotooddychania
wydajność kwantowa stała i niezależna
od temp.
-1% O
2
wydajność kwantowa stała i niższa
niż przy 21% O
2
(koszt wiązania CO
2
w pochwach okołowiązkowych)
w niższych temp. wyd. kw. u
C3 › C4
w 21% O
2
Ze względu na ograniczenie
fotooddychania
C
4
, 1% O
2
C
3
, niska temp.
zmod. Fig. 14.49. Buchanan 2000
1 8
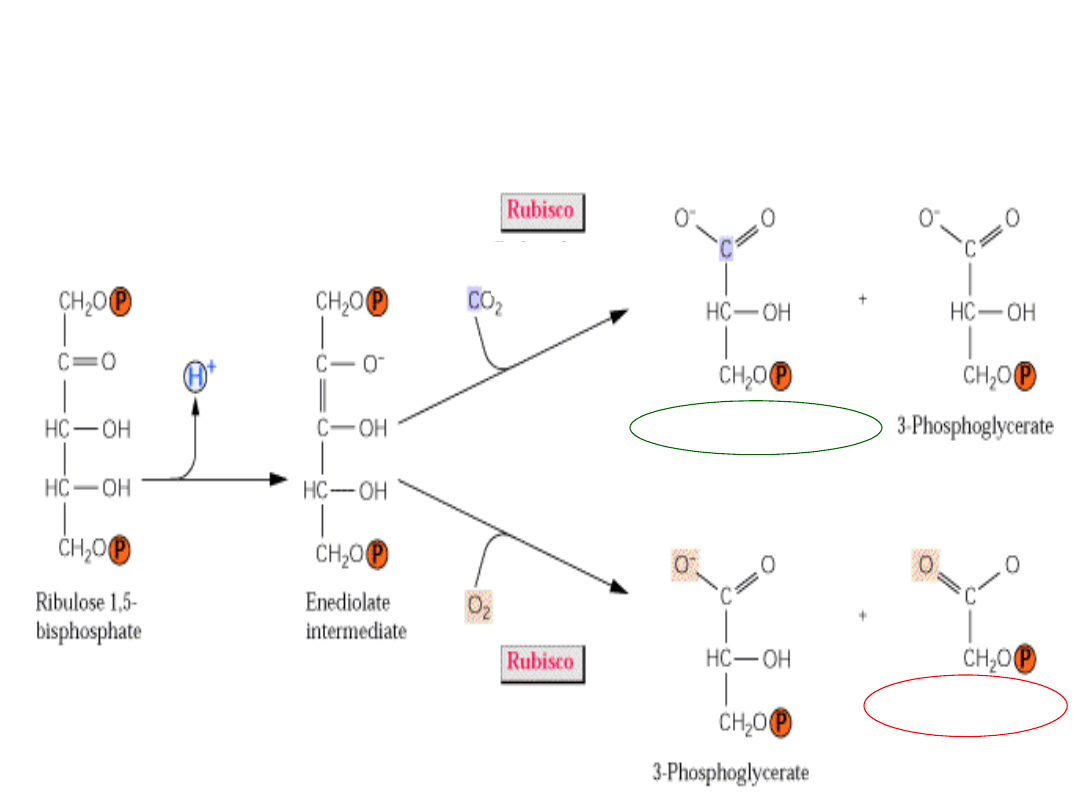
U roślin C3 Rubisco katalizuje
karboksylację
i
oksygenację
RuBP
(fotooddychanie)
3-
fosfoglicerynia
n
2-
fosfoglikola
n
fotosynte
za
fotooddych
anie
aktywność
karboksylaz
owa
aktywność
oksygenaz
owa
zmod. Fig. 14.45. Buchanan 2000
1 9
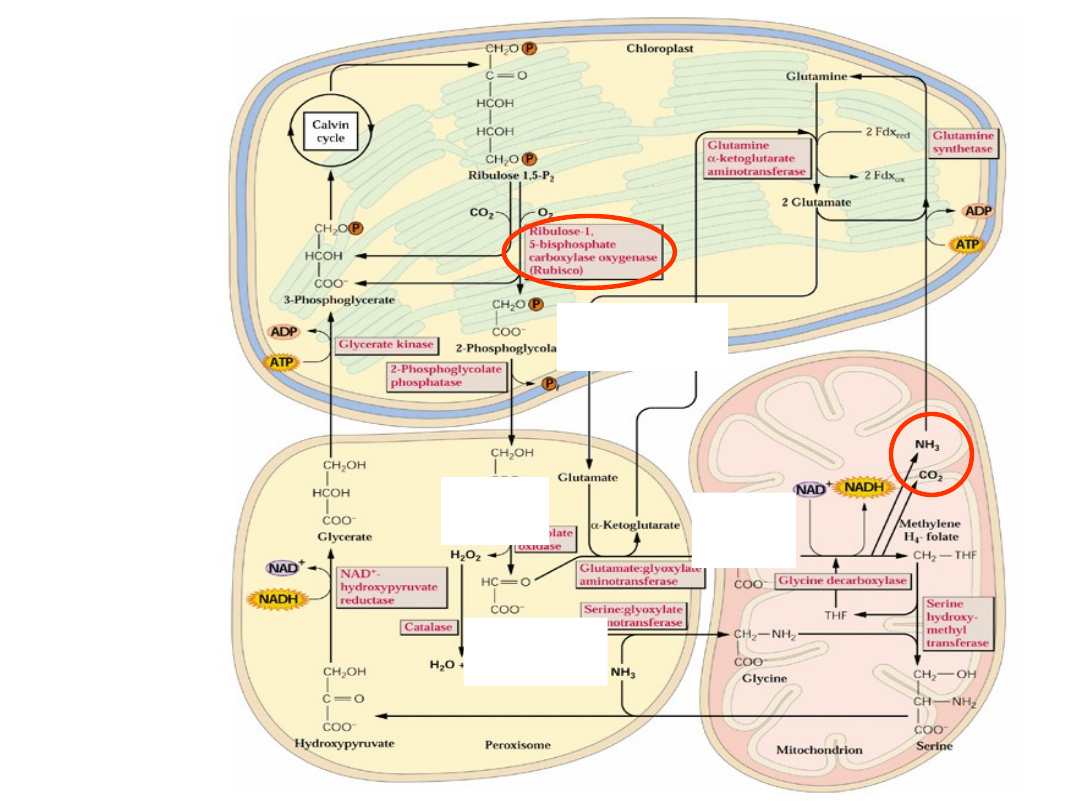
chloroplast
peroksyzom
Mitochondr
ium
glikol
an
glicy
na
fosfoglikol
an
glioksala
n
Fotooddychanie
zmod. Fig. 14.47. Buchanan 2000
2 0
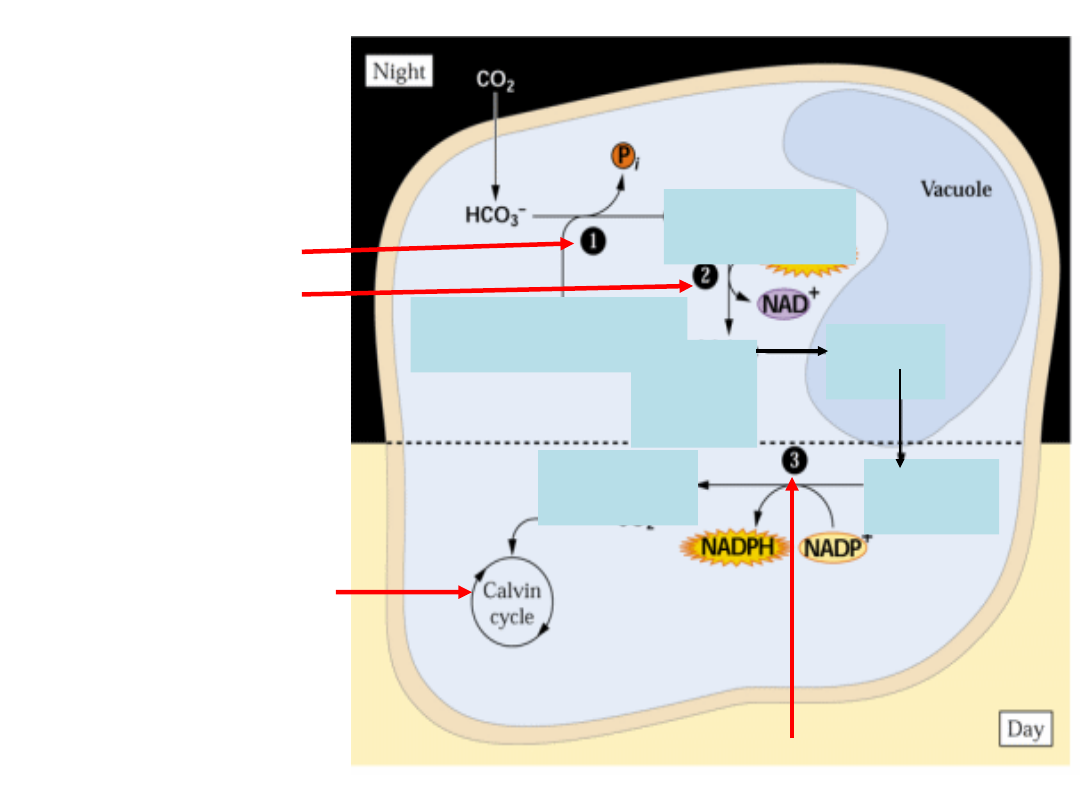
Fosfoenolo
pirogronian
szczawiooc
tan
jabłcz
an
pirogroni
an
Schemat cyklu C
4
i jego powiązanie z cyklem
C
3
Karboksylacja
Regeneracja
C
3
Dekarboksylacja
Komórka pochwy
okołowiązkowej (mezofil
wieńcowy)
Komórka
mezofilu
CO
2
u traw pierwszymi produktami
są 4-węglowe kwasy organiczne
etapy asymilacji
CO
2
są rozdzielone
przestrzennie
Karboksylaza PEP
RuBisCO
Wysokie stężenie CO
2
Rośliny C4
zmod. Fig. 12.46. Buchanan 2000
21
jabłcz
an
pirogron
ian
fosfoenolopirogr
onian
szczawiooc
tan
jabłcza
n
Asymilacja CO
2
u
roślin
kwasowych
(CAM)
jabłcz
an
karboksylaza PEP
dehydrogenaza
jabłczanowa
dehydrogenaza jabłczanowa dekarboksylująca
= malic enzyme
RuBisCO
etapy asymilacji
CO
2
są rozdzielone
w czasie
22

•W wyższej temperaturze wzrasta aktywność
oksygenazowa Rubisco
i
wzrasta fotooddychanie, bo spada stosunek
CO
2
/O
2
•Fotooddychanie w wyższej temperaturze jest
wyższe i obniża wydajność kwantową fotosyntezy
(CO
2
/kwant światła)
•U roślin C4 mechanizm zatężania CO
2
eliminuje
fotooddychanie
ale
•Wydajność kwantowa fotosyntezy u roślin C4 jest
niższa niż u C3 w warunkach nie-fotooddechowych
(1% O
2
).
Związane jest to z dodatkowym kosztem działania
cyklu C4 (2 ATP/CO
2
)
Fotooddychanie
23

Oddychanie na świetle
•Pomiar wymiany gazowej na świetle jest
utrudniony przez proces fotosyntezy – w ciemności
oddychanie stanowi 5-10% wymiany gazowej
fotosyntezy
•W nocy w oddychaniu zużywane jest 50-70% węgla
zasymilowanego podczas dnia
•Sądzono, że oddychanie na świetle jest bardzo
ograniczone, bo
cykl Krebsa na świetle jest częściowo zahamowany
(10-20% aktywności w ciemności)
ale:
•Udział fotosyntezy w budowaniu cytosolowej puli
ATP jest niewielki
•Mitochondria (fosforylacje łańcucha oddechowego)
uczestniczą w budowaniu cytosolowej puli ATP
•Synteza sacharozy w cytosolu zależy od
mitochondrialnego ATP
•Oddychanie mitochondrialne redukuje nadmiar
NADPH wytworzony w czasie fotosyntezy
24
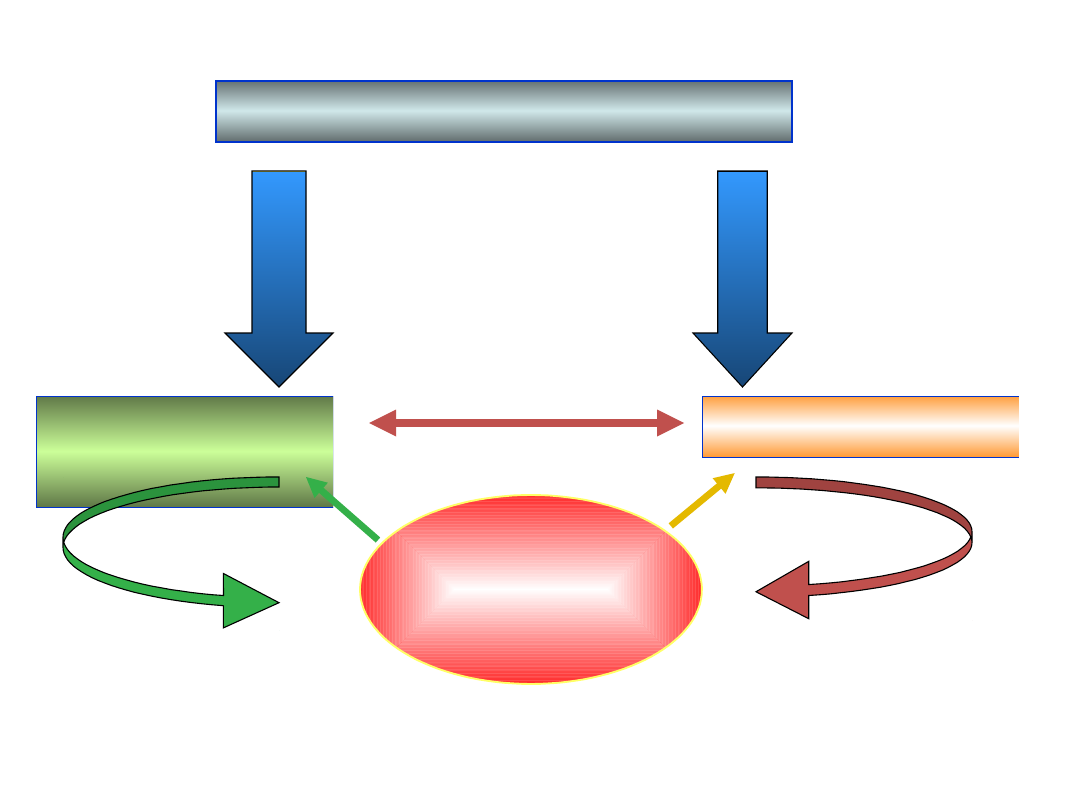
Czynniki środowiskowe
Fotosyntez
a
Oddychanie
ATP
25
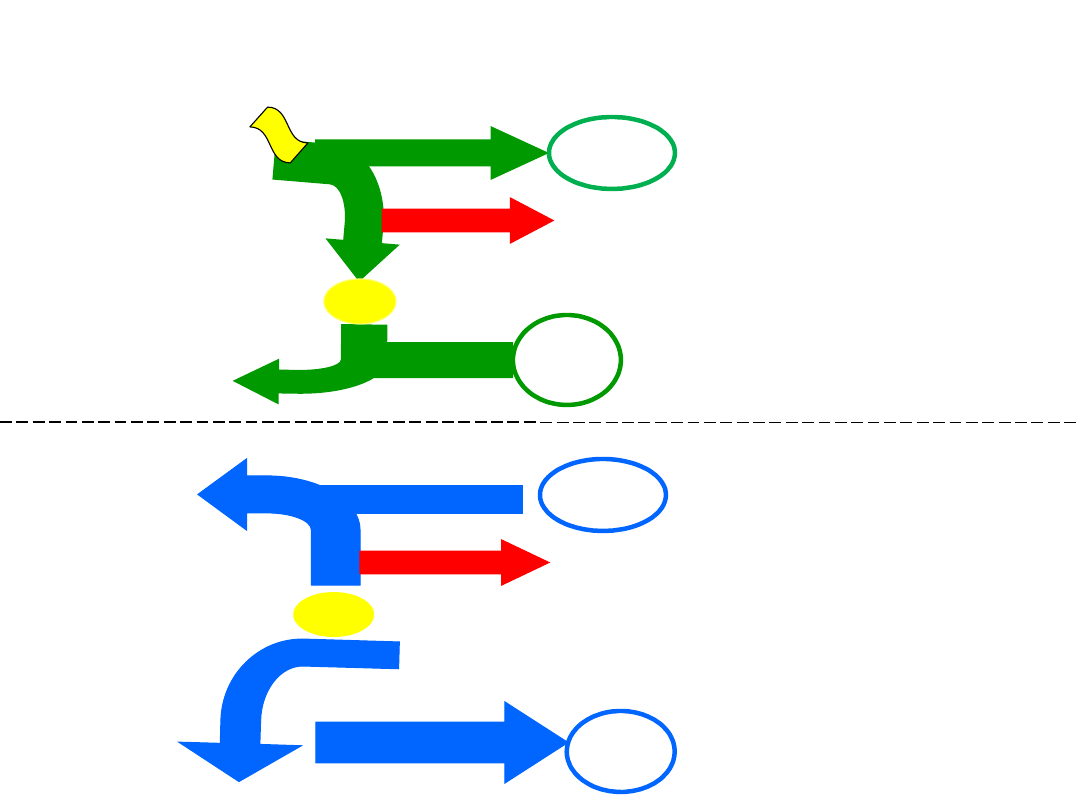
Fotosynteza
i
oddychanie
– przeciwstawność
procesów
FOTOSYNTEZA
światło
2e
-
0. 5
O
2
H
2
O
ATP
CO
2
CH
O
ODDYCHANIE
0. 5 O
2
H
2
O
ATP
2e
-
CH
O
CO
2
26
Fotosynteza
Rubisco
karboksylaza
PEP (ok.5%)
dekarboksylaza
glicyny
cykl TCA
CO
2
O
2
Fotooddychanie
Rubisco
Łańcuch oddechowy
(COX, AOX)
Reakcja Mehlera
(PSI)
Reakcje świetlne (PSII)
Wymiana gazowa na świetle
przewaga procesu fotosyntezy nad oddychaniem
(ok. 50% zasymilowanego CO
2
jest uwalniane w procesie oddychania)
27
Document Outline
- Slide 1
- Metabolizm węgla
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Aktywność fotosyntetyczna roślin
- Wpływ natężenia światła na fotosyntezę
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
Wyszukiwarka
Podobne podstrony:
wyklad 14 15 2010
Zarzadzanie strategiczne w organizacjach publicznych wyklad III listopad 2010
3 wykład (21 10 2010)
do wykladni prawa z 01 2010
10 Wykład (15 12 2010)
Komunikacja Bielicka wykład 1 24 04 2010
Makroekonomia Wykład 17 01 2010
9 wyklad 07 12 2010
wyklad 21.02.2010
organizacja uslug hotelarskich wyklad 07.03.2010, GWSH, organizacja usług w hotelarstwie
Wykłady bromatologia 2009-2010, Płyta farmacja Poznań, IV rok, bromatologia 2, wykłady
Wykład 02.10.2010 (sobota) A. Bandyra, UJK.Fizjoterapia, - Notatki - Rok I -, Kliniczne podstawy fiz
Wykład 08.05.2010
zarządzanie wykłady, 11.12.2010
Historia kultury, historia kultury- wyklad 20.02.2010, Historia kultury
KINEZJOLOGIA WYKŁAD 12 03 2010, Fizjoterapia CM UMK, Kinezjologia
więcej podobnych podstron