
Trawienie i
wchłanianie
węglowodanów
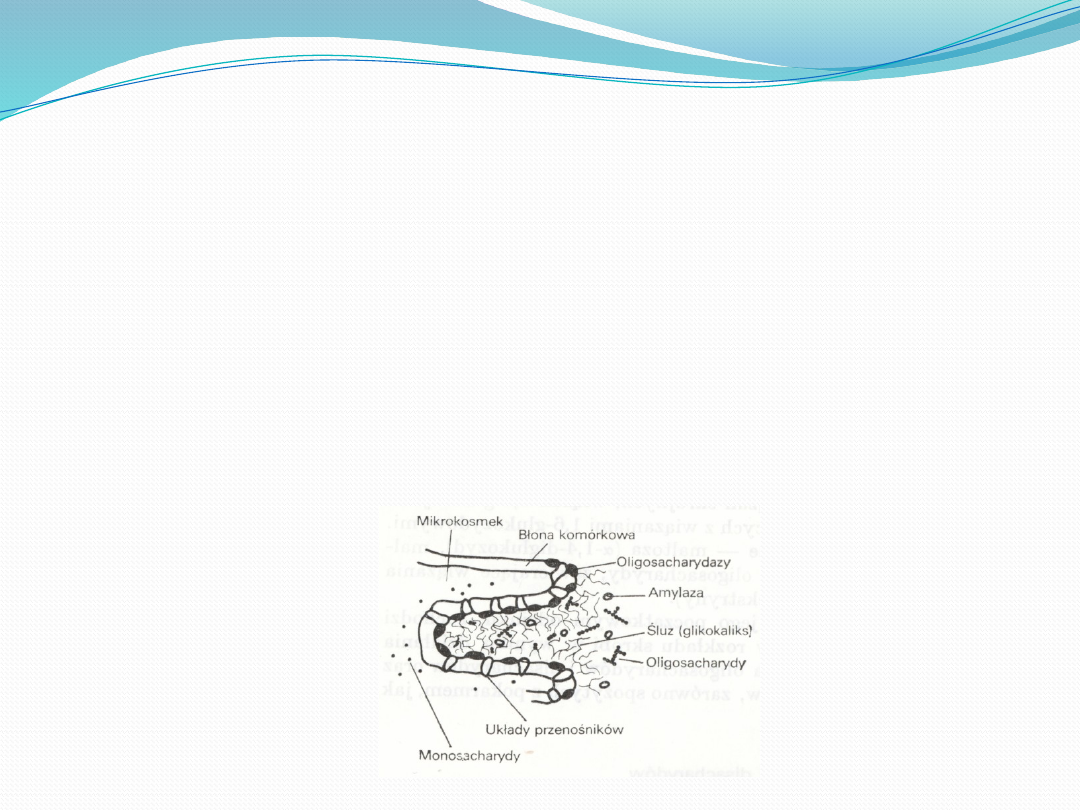
Podstawowymi czynnościami układu pokarmowego jest
trawienie i wchłanianie substancji pokarmowych.
Węglowodany są podstawowym składnikiem pokarmowym w
pożywieniu. Trawienie węglowodanów w przewodzie
pokarmowym zwierząt wykazuje różnice gatunkowe. W
zależności od pobieranego pokarmu, podstawą pożywienia
mogą być różne rodzaje cukrów. Podstawowymi cukrami
pokarmowymi zwierząt wszystkożernych, a także
mięsożernych, są skrobia i glikogen, natomiast zwierząt
roślinożernych błonnik i różne heteroglikany. Z tego względu
trawienie tych składników pokarmowych przez zwierzęta jest
bardzo zróżnicowane.

Trawienie skrobi i glikogenu rozpoczyna amylaza
ślinowa. Największą aktywność amylolityczną wykazuje
ślina człowieka. U świni aktywność ta jest około 100-
krotnie mniejsza. Ślina psa oraz zwierząt trawożernych
w ogóle nie zawiera amylazy. Działanie amylolityczne
tego enzymu jest najsilniejsze przy pH=6,7, w
obecności jonów wodorowych i chlorkowych. W ślinie
występuje przede wszystkim α-amylaza. Pod jej
wpływem następuje dekstrynizacja skrobi połączona z
odrywaniem cząsteczek maltozy i niewielkiej liczby
cząsteczek glukozy. Ślina rozpoczyna proces trawienia,
gdyż w żołądku następuje jego zahamowanie. Wzrost
stężenia kwasu solnego powoduje denaturację amylazy.

Dalsze etapy trawienia skrobi zachodzą w jelicie cienkim. Do
dwunastnicy wydzielana jest z sokiem trzustkowym amylaza
trzustkowa, działająca podobnie do amylazy ślinowej jednak w
wyższym pH. Enzym ten kończy trawienie skrobi i glikogenu,
przekształcając niestrawione dotychczas fragmenty tych cukrów w
maltozę. W soku jelitowym występują liczne glikozydazy, takie jak
maltaza, laktaza i sacharaza, które powodują rozszczepienie
dwucukrów do cukrów prostych. Laktaza umożliwia hydrolizę laktozy
do cukrów prostych: glukozy i galaktozy. Obecna jest ona w życiu
płodowym i aktywność jej spada razem z wiekiem. Maltaza powoduje
hydrolizę maltozy do glukozy. Sacharoza powoduje hydrolizę
sacharozy do glukozy oraz fruktozy
Cukry proste są wchłaniane do krwiobiegu głównie w początkowym
odcinku jelita cienkiego.
Cukry proste nie podlegają biernej dyfuzji, lecz transportowane są za
pomocą tzw. „carriera”- przenośnika, znajdującego się wewnątrz
błony i w sposób specyficzny transportującego cząsteczki
monosacharydów do wnętrza komórek nabłonka jelitowego

Przenośniki zostały ponumerowane w
kolejności ich odkrywania.
GluT1 znajduje się w wysokich stężeniach w
komórkach śródbłonka, które wyściełają naczynia
krwionośne oraz wytwarzają barierę między mózgiem
a krwią.
GluT2 występuje w narządach które uwalniają glukozę
do krwi takich jak wątroba jelito nerki.
GluT3 znajduje się w komórkach nerwowych mózgu,
zapewnia stały transport glukozy do neuronów.
GluT4 jest głównym przenośnikiem w mięśniach i
komórkach tłuszczowych.
GluT5 znajduje się głównie w jelicie cienkim i nerkach

Białka te składają się z odcinków Hydrofilowych
powinowatych ze środowiskiem wodnym i hydrofobowych
powinowatych z środowiskiem lipidowego błony komórkowej.
Wyżej opisana struktura przenoszenia glukozy do komórki
musi utworzyć kanaliki w błonie komórkowej .

Pięciokątny kanalik według założenia skład się
z pięciu spiralnych odcinków cząsteczki
przenośnika, takich jak odcinek numer 8. Jeśli
te spirale zwracały by swe lipidofilne strony
(niebieskie) w kierunku błony, a strony
wiążące glukozę (czerwone) do wewnątrz,
tworzyły by one kanalik, który mógłby
pomieścić cząsteczkę glukozy.
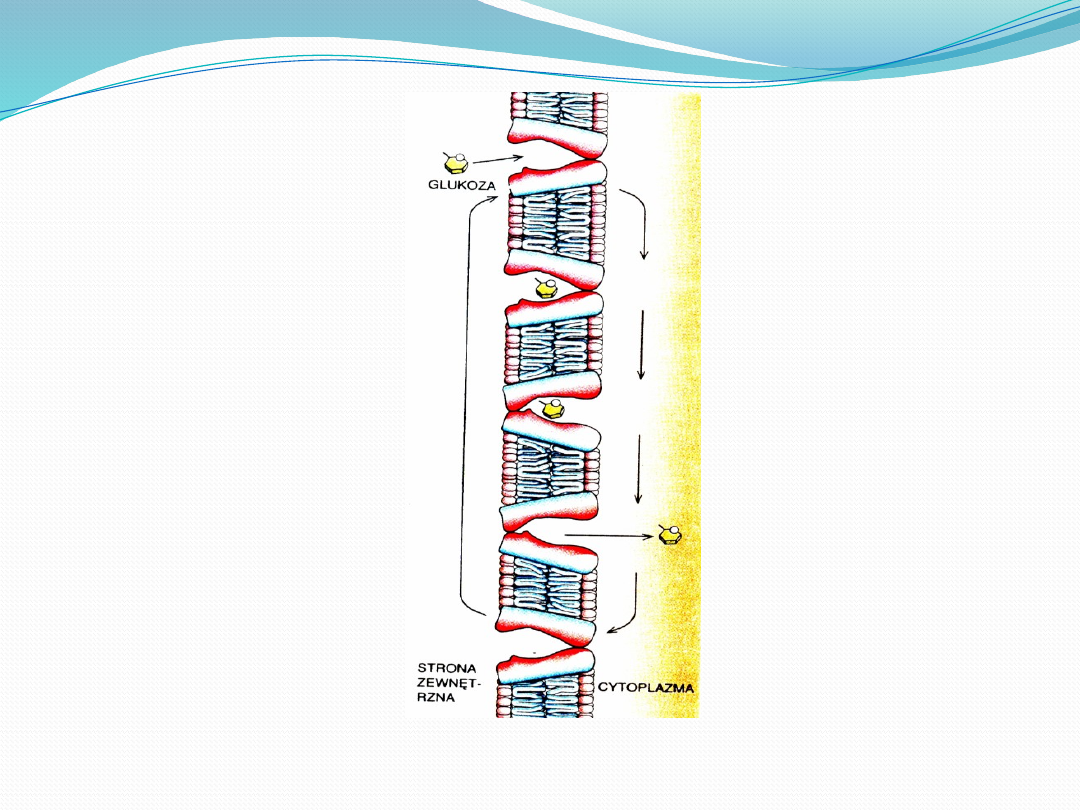

Białko przenośnikowe przybiera dwie formy: jedna wiąże
glukozę po zewnętrznej stronie błony komórkowej,
druga- po stronie wewnętrznej. Cząsteczka glukozy jest
wchłaniana przez komórkę w 4 etapach. W pierwszym
zajmuje ona skierowane na zewnątrz miejsce wiążące.
W drugim kompleks przenośnika i glukozy zmienia
konformacje w taki sposób, ze miejsce wiążące wraz z
glukoza kieruje się do wnętrza komórki. W trzecim
etapie przenośnik uwalnia glukozę do cytoplazmy
komórki. W czwartym wolny przenośnik zmienia
konformacje na taka, w której miejsce wiążące dla
glukozy jest skierowane na zewnątrz. Utworzony kanalik
jest otwarty na jednym końcu i zwężony na drugim oraz
ze glukoza wiąże się w kieszeni przy otwartym końcu.

Wszystkie cukry proste podlegające transportowi w jelicie
można podzielić na dwie grupy. Do jednej należą glukoza i
galaktoza, a do drugiej fruktoza, mannoza i pentozy. Cukry
drugiej grupy resorbowane są zgodnie z gradientem stężeń, a
transport ich odbywa się bez zużycia energii. Cukry pierwszej
grupy resorbują się znacznie szybciej, a co ważniejsze
resorbują się także wbrew gradientowi stężeń, a więc i
wówczas gdy ich stężenie w świetle jelita jest mniejsze niż w
krwi. Transport ten jest związany z użyciem energii, a ponadto
zależy od obecności jonu sodowego w środowisku. Jest to tak
zwany transport aktywny. Przy dużym stężeniu w jelicie, do
krwi mogą być wchłaniane dwucukry: maltoza, sacharoza i
laktoza. Spośród nich organizm zwierzęcy potrafi wykorzystać
maltozę przez jej rozszczepienie we krwi na cząsteczki
glukozy. Powstałe dwucukry są eliminowane z krwiobiegu
przez nerki.

Zwierzęta roślinożerne, przede wszystkim przeżuwacze i
koniowate, trawią przyjmowane przez nich wielocukry za
pośrednictwem enzymów bakterii i wymoczków żwacza i jelita
cienkiego. Trawienie celulozy odbywa się przy udziale celulaz
bakteryjnych. Powodują one rozszczepienie celulozy na
fragmenty w postaci prostych łańcuchów tzw. celodekstryn,
które następnie są degradowane do oligosacharydów, a w
dalszej kolejności do celobiozy i glukozy. Rozkład celobiozy
odbywa się z udziałem celobioazy, która jest β-glikozydazą,
bądź też za pośrednictwem fosforylazy celobiozowej,
prowadzącej do wytworzenia glukozo-1-fosforanów. Obydwa
te enzymy są wytwarzane przez mikroorganizmy, bytujące w
przewodzie pokarmowym tych zwierząt. Skrobia i dekstryna
są rozkładane przez amylazę do maltozy, a następnie przez α-
glukozydazy do glukozy. Fosforylaza maltozy może drogą
fosforylizy uwolnić glukozo-1-fosforan.

Heksozy pochodzące z różnych wielocukrów roślinnych ulegają
procesom prowadzącym do wytworzenia kwasów: pirogronowego i
mlekowego, a dalej przekształceniu ich w lotne kwasy
tłuszczowe(LKT). Procesy te zachodzą przy udziale enzymów
bakteryjnych i są określane jako fermentacje. Lotne kwasy
tłuszczowe są następnie transportowane przez ścianę żwacza do
krwi. Błony komórek nabłonka żwacza są przepuszczalne dla
kwasów zarówno zdysocjowanych jak i nie zdysocjowanych. Do
krwi mogą jednak przechodzić tylko kwasy nie zdysocjowane.
Wchłanianie lotnych kwasów tłuszczowych zachodzi
prawdopodobnie w drodze dyfuzji, zależnej od metabolizmu
komórek nabłonkowych. Na dyfuzję wywierają wpływ pH oraz
gradienty elektryczne treści żwacza oraz krwi.
Trawienie węglowodanów, a zwłaszcza skrobi, w jelicie cienkim
przeżuwaczy nie ma większego znaczenia, ponieważ sok
trzustkowy u tych zwierząt wykazuje słabe działanie amylolityczne.
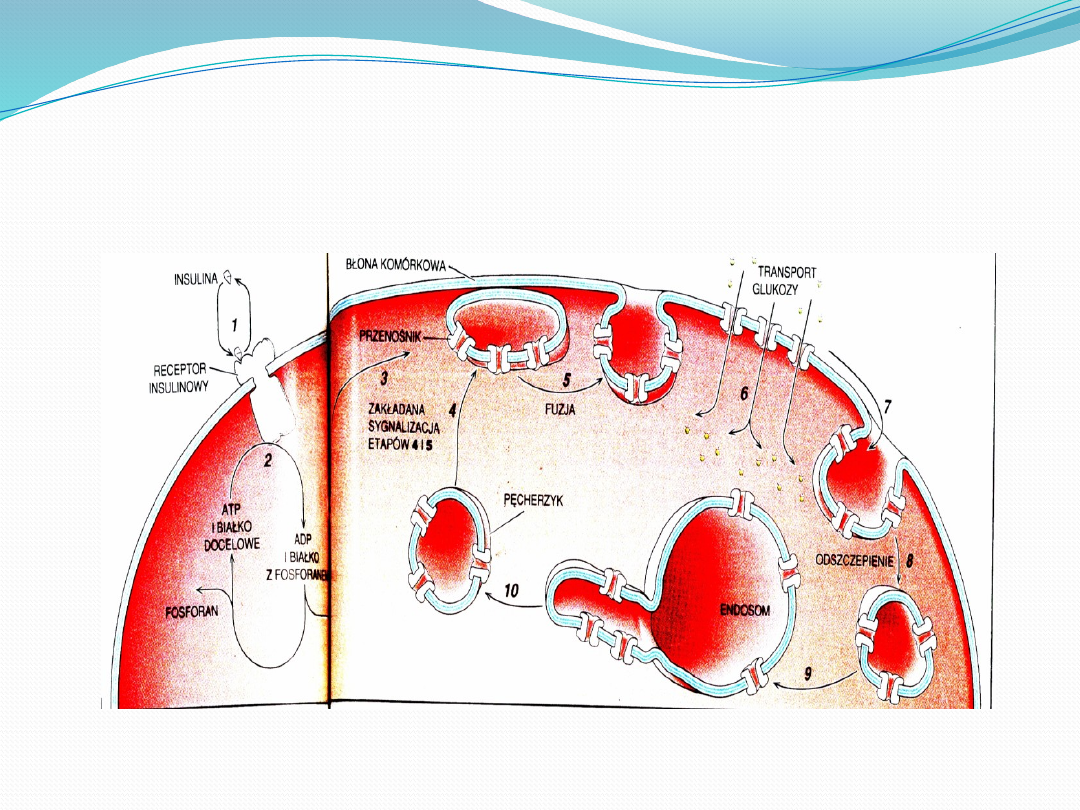
INSULINA-WSPOMAGANIE KOMORKI W
TRANSPORCIE GLUKOZY

Glukoza jest głównym cukrem we krwi, a wzrost jej poziomu po
wchłonięciu z jelita pobudza trzustkę do wydzielania insuliny, która
zwiększa pobieranie glukozy, przez komórki, tym samym stężenie
glukozy we krwi szybko powraca do poziomu fizjologicznego.
Zwrócona na zewnątrz część receptora insulinowego wiąże insulinę(1)
sprawiając ze część receptora zwrócona do wewnątrz przenosi grupę
fosforytowa z adenozynotrojfosforanu(2) na jedno lub więcej białek
docelowych tworząc ADP. Docelowe białko niosące fosforan wysyła
sygnał do przemieszczenia przenośników glukozy, wprawiając
pęcherzyki z przenośnikami w ruch w kierunku błony(3). Pęcherzyki te
stapiają się z błona komórkowa(5), przyspieszając transport
glukozy(6). Przenośniki trafiają z powrotem do wnętrza komórki(7),
kiedy małe pęcherzyki, utworzone przez wpuklenie błony i
odszczepienie, stapiają się z większymi endosomami(8), gdzie
przenośnik rozdziela się na rurkowate uwypuklenia(9), które odrywają
się tworząc nowe pęcherzyki(10). W obecności insuliny pęcherzyki
będą ciągle stapiać się z błona komórkowa(4), lecz obniżenie poziomu
insuliny przerywa opisany cykl, a przenośniki glukozy gromadzą się
wówczas w pęcherzykach wewnątrzkomórkowych
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Przenośniki zostały ponumerowane w kolejności ich odkrywania.
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- INSULINA-WSPOMAGANIE KOMORKI W TRANSPORCIE GLUKOZY
- Slide 14
Wyszukiwarka
Podobne podstrony:
Trawienie i wchlanianie weglowodanow AW
Trawienie i wchlanianie weglowodanow AW
18. Trawienie i wchłanianie białek i węglowodanów, licencjat(1)
trawienie i wchłanianie tłuszczów
Procesy trawienia i wchłaniania w układzie pokarmowym człowieka
trawienie i wchłanianie białek
trawienie i wchłanianie cukrów
Trawienie i wchłanianie
Trawienie i wchłanianie antastic pl
TRAWIENIE I WCHŁANIANIE, TRAWIENIE I WCHŁANIANIE:
Trawienie i wchlanianie substancji odzywczych
W2 Trawienie wchlanianie
19. Trawienie i wchłanianie lipidów, licencjat(1)
Trawienie i wchłanianie substancji odżywczych
trawienie i wchłanianie
Wykład 9 trawienie i wchłanianie
5 Trawienie i wchlanianie substancji odzywczych
TRAWIENIE I WCHŁANIANIE POKARMÓW
TRAWIENIE I WCHŁANIANIE WERSJA OSTATECZNA
więcej podobnych podstron