ZESTAW 13
1. Cykl kwasy cytrynowego (cykl Krebsa)
Główną funkcją cyklu Krebsa jest utlenianie pirogronianu do Co2 i H2O z jednoczesnym uzyskiwaniu energii. Cykl ten również odgrywa rolę w wytwarzaniu prekursorów dla szlaków biosyntez. Składa się on z ośmiu etapów:
I. Utworzenie cytrynianu ze szczawiooctanu i acetylo-CoA. Reakcja katalizowana przez cyntazę cytrynianową.
II. Izomeryzacja cytrynianu do izocytryniany. Reakcja katalizowana przez akonitazę.
III. Utlenianie izocytrynianu do α - ketoglutaranu. Reakcja katalizowana przez
dehydrogenazę izocytrynianową, wymaga NAD+.
IV. Utlenianie α - ketaglutaranu do bursztynylo-CoA. Reakcja katalizowana przez kompleks dehydrogenazy α - ketaglutaranowej; eymaga NAD+
V. Przekształcenie bursztynylo- CoA w bursztynian. Reakcja katalizowana przez syntetazę bursztynylo-CoA, reakcja wymaga fosforanów nieorganicznych i GDP lub ATP
VI. Utlenianie bursztynianu do fumaranu. Reakcja katalizowana przez dehydrogenazę bursztynianową; uczestniczy FAD)
VII.
Uwodnienie fumaranu do jabłczanu. Reakcja katalizowana przez fumarazę.
VIII.
Utlenienie jabłczanu do szczawiooctanu. Reakcja katalizowana przez dehydrogenazę
jabłczanową; wymaga NAD+
Podczas każdego obrotu cyklu, powstaje 12 cząsteczek ATP.
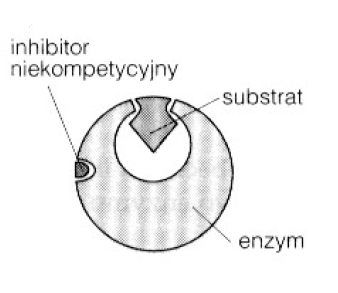
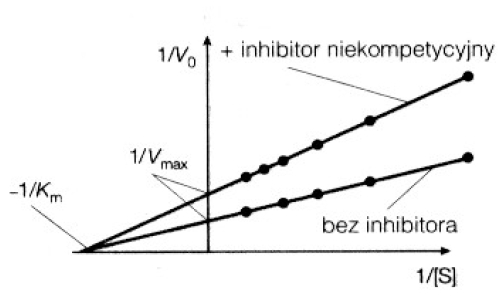
2. Inhibitor niekompetycyjny + wykres.
Inhibitor niekompetycyjny- wiąże się w enzymie z miejscami innymi niż miejsce aktywne i zmniejsza szybkość katalityczną enzymu, powodując konfirmacyjną zmianę w jego kształcie przestrzennym. Wpływu tego inhibitora nie można przezwyciężyć przez duże tężenie substratu.
Wykres Lineweara- Burke'a ukazuje, że inhibitor niekompetycyjny zmniejsza Vmax, ale nie zmienia wartości Km. Wynika to z faktu, że efektu działania tego inhibitora nie da się przezwyciężyć przez zwiększenie stężenia substratu, a powinowactwo enzymu do substratu nie zmienia się.
Inhibitor niekompetycyjny wiąże się z enzymem odwracalnie w innym miejscu niż substrat.
Powodując zmianę przestrzennego kształtu enzymu, co prowadzi do zmniejszenia aktywności katalitycznej. Ponieważ inhibitor wiąże się w innym miejscu niż substrat, enzym może wiązać albo inhibitor, albo substrat lub obydwa naraz.
β - oksydacja acetylo- CoA
Reakcje β - oksydacji zachodzą w matrix mitochondrium u eukariota i cytozolu u prokariota.
Polegają na takich przemianach, aby rozszczepić „dłuższy”acylo-CoA na acetylo-CoA i acylo-CoA
„krótszy”. Obejmuje ona cztery reakcje zachodzące cyklicznie.
I. Utlenienie acylo-CoA do trans−Δ2 - anilo- CoA. Reakcja katalizowana przez dehydrogenazę acylo-CoA z wytworzeniem FADH2.
II. Uwodnienie trans−Δ2 - anilo- CoA do 3-hydroksyacylo-CoA. Reakcja katalizowana przez hydratazę enoilo-CoA.
III. Utlenienie 3-hydroksyacylo-CoA do 3- ketocylo-CoA. Reakcja katalizowana przez dehydrogenazę hydroksylo-CoA; z wytworzeniem NADH.
IV. Tioliza 3-ketoacylo-CoA przez drugą cząsteczkę CoA i stworzenie acylo-CoA „krótszego”
oraz acetylo-CoA.
4. Szlak glukoneogenezy i jej znaczenie.
Glukoneogeneza to proces, w którym zachodzi synteza glukozy z prekurorów nie będących cukrami. Ma duże znaczenie dla podtrzymania zawartości glukozy we krwi podczas głodowania lub dużego wysiłku fizycznego. Zachodzi głównie w wątrobie. W mniejszym stopniu w nerkach, Można go podzielić na 6 etapów:
I. Przekształcenie pirogronianu w szczawiooctan. Reakcja katalizowana przez karboksylazę pirogronianową.
II. Szczawiooctan ulega przemianie do fosfoenolopirogronianu (PEP). Reakcja katalizowana przez karboksykinazę PEP.
III. PEP jest przekształcane do fruktozo-1,6-bisfosforanu. Reakcja zachodzi dzięki przekształceniom kilku reakcji glikolitycznych
IV. Zdefosforylowanie fruktozo-1,6-bifosforanu do fruktozo-6-fosforanu.
V. Przekształcenie fruktozo-6-fosforanu w glukozo-6-fosforan. Reakcja katalizowana przez izomerazę glukozofosforanową.
VI. Zdefosforylowanie glukozo-6-fosforanu do glukozy.
Synteza 1 cząsteczki glukozy z dwóch cząsteczek pirogronianu wymaha 6 ATP.
Wyszukiwarka
Podobne podstrony:
Zestaw 13
Zestaw 13, 4 semestr, matlab, testy
biofizyka egzamin zestawy 13
planimetria zestawy 13 i 14 rozwiązane- Aksjomat ToruĹ„
zestaw 13, AiR, Semestr 2, Grafika inżynierska, zadania grafika
Zestaw 13 InzB
Zestaw 13, Ćwiczenia
opracowane zestawy 13
Zestaw 13 - Świat przyrody, Matura ustna podstawowa
Zestaw 13, Politechnika Lubelska, Studia, Studia, Sprawozdania, studia, Matematyka, MATEMATYKA WYKŁA
zestaw 13 ALzG
zestaw 13 kinetyczna teoria gazów, SEMESTR I, MECHANIKA I FIZYKA STATYSTYCZNA, zadania
Zestawy 13 diagnoza
Geometria krzywych i powierzchni, zestaw 13
ZESTAW 13
Zestaw 13 do ko-ca, Zestaw 13
zestawy maturalne, ZESTAW 13, ZESTAW 13
więcej podobnych podstron