ZAGADNIENIA NA EXAM Z GENETYKI 2010-02-06
1. PRAWA MENDLA + ZADANIA
I Prawo Mendla --> prawo czystości gamet: gamety zawierają tylko po jednym allelu danego genu.
- tu wchodzą zadania na dziedziczenie jednogenowe z pełną dominacją (typu Pisum)np. kwiaty białe/czerwone u koniczyny, skrzydła normalne/szczątkowe u muszki owocowej, sierść normalna/albinotyczna u myszy, kolor oczu, mukowiscydoza, anemia sierpowata, fenyloketonuria u człowieka.
Czyli w pokoleniu F1 sa identyczne fenotypowo osobniki a w F2 dochodzi do rozszczepienia cech w stosunku 3:1;
Odchylenia od tego może powodować: niepełna dominacja, kodominacja, geny letalne, subletalne lub wielokrotne
II Prawo Mendla --> prawo niezależnej segregacji dwóch par alleli; allele dwóch różnych genów przechodzą do gamet niezależnie od siebie.
- to dotyczy zadanek dwucechowych o pełnej dominacji np. to z ziarnkami groszku
W pokoleniu F1 wszytskie osobniki SA identyczne fenotypowo a w F2 dochodzi do rozszczepienia fenotypów w stosunku 9:3:3:1.
Odchylenia powoduje: niepełna dominacja, geny letalne, epistaza, sprzężenie genów, niepełna penetracja, ekspresywnośc genów.
2. CHROMOSOMOWA TEORIA DZIEDZICZENIA MORGANA
Chromosomy jako siedliska genów ułożonych liniowo („układy sprzężeń”). Morgan wykazał, że sprzężone są ze sobą te geny, które występują na jednym chromosomie i one nie dziedziczą się niezależnie od siebie (np. u Drosophili są 4samice lub 5samce grupy genów sprzężonych)
3. JEDNOZNACZNE ZDEFINIOWANIE POJĘCIA „GENU”
w sensie strukturalnym jeden gen koduje jeden łańcuch polipeptydowy (lub jeden rodzaj tRNA lub rRNA), decyduje o jego I-rzędowej strukturze
w sensie funkcjonalnym--> to cistron, czyli odcinek DNA, który w teście cis-trans wykazuje efekt pozycji. A po polsku :) - jeżeli dwie mutacje niekomplementują się po skrzyżowaniu czyli nie powstaje dziki fenotyp to znaczy, że są mutacjami allelicznymi i tworzą tzw. grupę komplemenatcji, która w sumie odpowiada pojęciu cistronu.
4. PODSTAWOWE POJĘCIA GENETYCZNE
Gen i cistron jak wyżej :)
Allel --->1. to alternatywna postać danego genu, zajmująca to samo locus w chromosomach homologicznych i mogąca powodować zmiany w wykształceniu danej cechy (to wg książki). 2. to jedna z dwóch lub większej liczby wykluczających się form genu, znajdujących się na tym samym locus chromosomu (od prof.).
Genom --> zespół haploidalnej liczby chromosomów (czyli kobieta 23 a mężczyzna 24!!)
Kariotyp --> morfologiczny obraz haploidalnej liczby chromosomów
Genotyp --> suma całej informacji genetycznej, wszystkich genów we wszystkich chromosomach - w układzie sprzężonym
Fenotyp --> zespól ujawnionych cech danego osobnika warunkowanych przez genotyp i wpływ środowiska
Diploidalność --> zjawisko występowania w komórkach somatycznych dwóch chromosomów homologicznych każdego rodzaju, dotyczy ogólnie Eucaryota. Taki sukces ewolucyjny, że mamy kopię zapasową i możliwość większej zmienności (crossing over!). Dotyczy też grzybów i drożdży.
Haploidalność --> zjawisko u Procaryota, gdzie w komórkach somatycznych organizmu jest tylko po jednym chromosomie każdego typu. Dotyczy też grzybów i drożdży.
Allel (gen) dominujący --> to ten, który ujawnia się fenotypowo zarówno w homozygocie jak i heterozygocie. Jest tak samo wydajny jak dwie kopie genu.
Allel (gen) recesywny --> to ten, który ujawnia się w homozygocie ale nie w heterozygocie.
Krzyżówka testowa --> krzyżówka „mutanta” (czegoś w pokoleń potomnych) z homozygota recesywna w celu zidentyfikowania genotypu rodziców.
Krzyżówka wsteczna --> skrzyżowanie F1 lub F2 z jednym z rodziców.
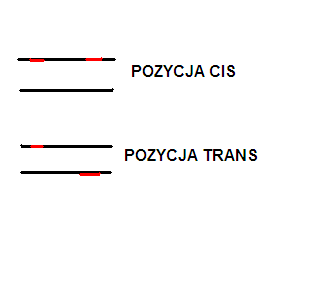
5. TEST KOMPLEMENTACJI CIS-TRANS
Pozwala na określenie alleliczności mutacji. Dotyczy oczywiście tylko organizmów diploidalnych
Po kolei ma się to tak:
Mamy dwie mutacji, które pojawiają się niezależnie i dają taki sam fenotyp mutanta i tu problem - czy to mutacje tego samego czy różnych genów.Krzyżujemy homozygoty obu mutantów (jeden ze względu na jedna mutacje a drugi ze względu na druga mutację) i patrzymy czy w pokoleniu potomnych powstał jakiś osobnik dziki --> czyli czy doszło do komplementacji (uzupełniania się) u heterozygoty.
Jeśli jest komplementacja to: są to mutacje niealleliczne
Jesli brak komplementacji i mamy same mutanty: są to mutacje alleliczne a zmutowane allele to tzw. allele funkcjonalne (cistrony).
6.SENS BIOLOGICZNY MITOZY I MEJOZY
mitoza --> zachowanie stałej liczby chromosomów i genów w komórkach tego samego organizmu
mejoza --> zachowanie stałej liczby chromosomów organizmów danego gatunku (czyli znaczenie w zakresie populacji) + podstawa zmienności organizmów przez crossing over, niezależną segregację chromosomów i losowe łączenie się gamet
7. EPISTAZA
Jest to zjawisko dotyczące współdziałania genów nieallelicznych. Jeden gen (epistatyczny) maskuje fenotypową ekspresję innych genów (hipostatycznych), zmieniając w ten sposób fenotyp.
Mamy 5 rodzajów epistazy:
epistaza recesywna (9:3:4) --> maskowanie przez allele recesywne jednego genu efektu genu w innym locus np. determinacja koloru sierści u gryzoni. Mechanizm tego zjawiska: epistatyczny gen recesywny koduje niefunkcjonalny enzym i nie powstaje barwnik więc homozygoty względem tego genu sa albinosami.
podwójna epistaza recesywna --> komplementacja genów (9:7) --> współdziałanie genów dominujących z różnych loci wymaga ich równoczesnej obecności (komplementacji), aby ujawnił się określony fenotyp np. barwa kolb u Zea mays. Mechanizm: oba geny zaangażowane są w jeden szlak metaboliczny a dominujące formy obu genów są konieczne, aby ujawnił się barwny fenotyp.
epistaza dominująca (12:3:1) --> alle dominujący w jednym locus maskuje efekt genu w innym mlocus, np.dziedziczenie barwy owocu u dyni lub barwy upierzenia u kurcząt. Mechanizm: oba geny zaangażowane w jeden szlak metaboliczny, gdzie ekspresja allelu dominującego jednego genu prowadzi do zahamowania tego szlaku a allel dominujący drugiego genu prowadzi do wytworzenia barwnego produktu.
podwójna epistaza dominująca(15:1) --> wystarczy tylko jeden allel dominujący jednego z dwóch współdziałających genów, aby ujawnił się fenotyp dominujący, np. kolor ziarniaków pszenicy. Mechanizm nieznany.
epistaza dominująca i recesywna (13:3) -->zarówno allel domiujący jedego z genów , jak i allel recesywny drugiego genu maskują efekt genu z innego locus, np. zabarwienie cebuli. Mechanizm sobie podarujemy :)
8. ODSTĘPSTWA OD PRAWA MENDLA (czyli dotyczą cechy jednogenowej
Niepełna dominacja (=dziedziczenie typu Zea) -->gdy heterozygota ma fenotyp pośredni w stosunku do homozygot rodzicielskich a stosunek fenotypowy w F2 wynosi 1:2:1, ponieważ tu genotyp odzwierciedla fenotyp!
Np. barwa kwiatów u wyżlinu --> krzyżując białą homozygotę z czerwoną homozygotą mamy w F1 100% różowe kwiatki.
W F2 natomiast 25% czerwonych (oczywiście homozygot!), 50% różowych (to heterozygoty) i 25% białych (homozygoty).
Inne przykłady tego to: szurpatość u kur, umaszczenie bydła rasy shorthorn, upierzenie kur andaluzyjskich
Wspóldominowanie (=kodominacja, dopełnianie się) --> tutaj w przypadku heterozygot cechy uwarunkowane przez dwa allele danego genu ujawniają się niezależnie od siebie. Oba allele są równorzędne a heterozygota prezentuje fenotyp obojga rodziców.
Np. dziedziczenie u ludzi grupi krwi z układu AB0 (allele IA oraz IB sa współdominujące), anemia sierpowata (heterozygoty HbS/HbA - nie są ciężko chore, mają tylko część krwinek zdeformowanych a do tego są bardziej odporne na malarię),
Epistaza --> patrz pyt. 7
Allele wielokrotne --> taki wybryk natury, że zamiast jak zwykle dwóch alternatywnych form danego genu w populacji jest więcej. Oczywiście dany osobnik ma ZAWSZE tylko dwa allele danego genu ale jak mu sie trafią np. dwa dominujące to będzie jak we wspóldominowaniu.
Np. trzy allele warunkujące grupę krwi w układzie AB0, allele głównego kompleksu zgodności tkankowej, allele genu barwy sierści u królika (agouti, szynszyla, albinos i himalajski typ ubarwienia) oraz allele genu white (w) u Drosophili.
Allele letalne i subletalne -->
Allele letalne dominujące powodują śmierć homozygot i heterozygot. Allele letalne recesywne powodują smierć homozygot, heterozygoty spokojnie sobie żyją zazwyczaj. Mamy tutaj dwie wazne choroby!: Taya-Sachsa (gromadzenie się gangliozydów w neuronach przez defekt ich degradacji w lizosomach; smierć ok 2-4 roku życia), Pląsawica Huntingtona (przez obumieranie komórek jądra ogoniastego mózgu, choroba od momentu ujawnienia do smierci trwa ok 15 lat)
Allele subletalne --> podobne efekty do alleli subletalnych, ponieważ obniżają żywotność i płodność u homozygot recesywnych. Np. allel recesywny genu vestigal (vg) u Drosophili warunkujący szczątkowe skrzydła oraz allel szurpatego upierzenia u kur, który zwiększa ich śmiertelność.
Sprzężenie genów --> patrz pytanie 2
Penetracja i ekpresywność genów -->
Penetracja określa procent osobników ujawniających fenotyp charakterystyczny dal danego genotypu (całkowita = 100%, niezupełna < 100%)
Ekspresywność określa stopień nasilenia cech w fenotypie osobników o tym samym genotypie.
Na obydwie te sprawy mają wpływ: tło genetyczne, współdziałanie z innymi genami, wiek, płeć, warunki środowiska.
9. WPŁYW ŚRODOWISKA NA FENOTYP
Dotyczy to tzw. zmienności fluktuacyjnej organizmów, czyli zdolności wytworzenia różnych fenotypów u organizmów o tym samym genotypie w zależności od warunków środowiska. Np. kwiaty hortensji przez dodanie siarczanu glinu lub żelaza zmieniają barwę płatków z czerwonego na niebieski itp. warunki środ. jak temperatura, pH gleby, światło, dieta mają wpływ na roślinki i zwierzęta.
Patrz też --> penetracja i ekspresywnośc genów z pytania 8.
10. DETERMINACJA PŁCI
Chromosomowa --> związana z obecnością chromosomów płci
system XY - u większości zwierząt, XX to samice, XY to samce (u ssaków ważna rola genu SRY na chromosomie Y, który odgrywa głowna rolę w maskulinizacji osobnika)
system X0 - u większości owadów, samice XX produkują parzystą liczbę chromosomów a samce X0 nieparzystą
system ZW - u ptaków, ciem, niektórych ryb, płazów i gadów; samce sa homogametyczne a samice heterogametyczne
Genowa --> pojedyncze geny autosomalne determinują płeć, występuje u wielu gatunków żab, ryb i roślin
Cytoplazmatyczna --> u niektórych roślin mtDNA zawiera geny męskiej sterylności, ale mogą to przywrócić jądrowe geny męskiej płodności
Haploidalna determinacja płci --> u błonkoskrzydłych, samice są diploidalne a samce haploidalne
Środowiskowa (fenotypowa) -->u wielu ryb, gadów; płeć uwarunkowana zadziałaniem odpowiedniego czynnika środowiskowego np. temperatury
Brak zróżnicowania płciowego --> u wielu roślin i np. płazińców, pierścienic
U drosophili --> płec determinuje stosunek liczby chromosomów X do liczby zespołów autosomów (1=samica, <1=samiec)
11. CECHY SPRZĘŻONE Z PŁCIĄ I UWARUNKOWANE PRZEZ PŁEĆ
sprzężone z płcią (czyli leżą na chromosomach płci) np. dystrofia mięśniowa Duchenne'a, hemofilia A i B, daltonizm, zespół kruchego chromosomu X, gen white (w) u Drosophili itp.
uwarunkowane przez płeć --> nie leża na chromosomach płci lecz na autosomach jednak ich ekspresja fenotypowa zależy od płci - czyli głównie od stężenia męskich hormonów płciowych, np. łysienie u ludzi lub obecność poroża u bydła
12. MAPOWANIE GENÓW
dotyczy chromosomowej teorii dziedziczenia Morgana i grup sprzężeń
Geny sprzężone --> leżące na tym samym chromosomie
Mapowanie --> polega na ustalaniu wzajemnego położenia i odległości genów w chromosomie
Sprzężenie całkowite --> gdy między genami znajdującymi się na jednym chromosomie nie zachodzi crossing-over, czyli nie dochodzi do rekombinacji
Sprzężenie częściowe --> gdy między genami znajdującymi się na jednym chromosomie dochodzi do crossing-over, czyli dochodzi do rekombinacji i powstają nowe kombinacje genów na chromosomach. Na podstawie częstości crossing-over można oszacować względne odległości między loci na chromosomie --> Określa sie to jako:
Żeby ustalić względne położenie pary genów na chromosomie wykonujemy krzyżówkę dwupunktową (podwójna heterozygota * podwójna homozygota recesywna) i jeśli nie otrzymamy w F1 stosunku 1:1:1:1 to są to geny sprzężone i z ilości rekombinacji ustalamy odległość (odsyłam do zadań). Jest jeszcze krzyżówka trójpunktowa, która pozwala na tworzenie map genetycznych, czyli określenia jak po kolei leżą geny na chromosomie oraz umożliwia ocenę wpływu wielokrotnych crossing-over. Krzyżujemy potrójna heterozygotę z potrójna homozygota recesywną. C.d odsyłam do zadań :)
Dochodzi tu także MAPOWANIE MEJOTYCZNE na podstawie analizy worków S. cerevisiae i N. crassa --> patrz: materiały do ćwiczeń
13. BUDOWA DNA I RNA
DNA --> cząsteczka składająca się z dwóch nici biegnących antyrównolegle i owiniętych wokół wspólnej osi tworząc prawoskrętna podwójną helisą. Rdzeń tworzą nukleotydy, które łączą się wiązaniami wodorowym z nukleotydem naprzeciwko w drugiej nici (Adenina z Tyminą oraz Guanina z Cytozyną).
RNA --> jednoniciowa lun czasem dwuniciowa (głównie wirusy, wiroidy) helikalnie skręcona cząsteczka, zbudowana także przez 4 rodzaje nukleotydów (Adenina+ URACYL oraz Guanina + Cytozyna). Występuje też ryboza a nie deoksyryboza jak w DNA!
14. REPLIKACJA DNA
to proces endoenergetyczny, w którym podwójna nić DNA (podwójna helisa) ulega skopiowaniu. Replikacja jest semikonserwatywna (półzachowawcza) - w każdej z dwóch uzyskanych podwójnych nici DNA będzie jedna nić macierzysta i jedna nowa.
U bakterii replikacja zaczyna się w ustalonym miejscu i postępuje bardzo szybko, z prędkością rzędu 1000 nukleotydów/sek. U eukariotów replikacja jest o wiele wolniejsza, ok. 50 nukleotydów/sek., jednak zachodzi równocześnie w wielu miejscach.
Zachodzi na zasadzie komplementarności zasad.
Polimeraza DNA działa jedynie w kierunku od końca 3' do końca 5' (czyli syntetyzuje nową nić w kierunku od 5' do 3'). Z tego powodu jedna z nici jest syntezowana w sposób ciągły (Watsona), druga (Cricka) (ta, którą chcielibyśmy zsyntezować w przeciwną stronę) fragmentami (tzw. fragmenty Okazaki)- które są syntetyzowane przez prymazę a powstające odcinki DNA łączy ligaza.
miejsce inicjacji replikacji (=miejsce ori) - CACTAT
helikazy - rozrywają wiązania wodorowe między nićmi matrycowego DNA, rozkręcając helisę i umożliwiając rozpoczęcie procesu;
prymaza - syntetyzuje starter;
polimerazy DNA - polimeryzuje zgodnie z zasadą komplementarności fosforany deoksyrybonukleotydów;
topoizomerazy - rozplatają podwójną helisę DNA, udostępniając w ten sposób matrycę dla enzymów replikacyjnych lub transkrypcyjnych.
Gyraza DNA - enzym występujący u Procaryota (szczególnie u bakterii), należący do topoizomeraz typu II. Przy użyciu energii pochodzącej z hydrolizy ATP katalizuje on wprowadzanie ujemnych skrętów w helisie DNA.
egzonukleaza - usuwa startery RNA z nici;
ligaza DNA - uzupełnia brakujące wiązania fosfodiestrowe w szkielecie nowozsyntetyzowanej nici DNA;
15. EKSPRESJA GENÓW I JEJ REGULACJA U PROCARYOTA I EUCARYOTA.
proces, w którym informacja genetyczna zawarta w genie zostaje odczytana i przepisana na jego produkty, które są białkami lub różnymi formami RNA.
U bakterii geny są zwykle zorganizowane w grupy genów zwane operonami (np. operon laktozowy), które są regulowane jako grupa i przepisywane na zawierający kilka genów, mRNA.
U eukariotów regulacja oraz przepisywanie na mRNA odnosi się do pojedynczego genu. Proces ten zachodzi w kilku etapach
zainicjowanie transkrypcji genu przez czynniki transkrypcyjne --> moment najważniejszy!!!!!!!!
synteza pre-mRNA przez polimerazę RNA
obróbka posttranskrypcyjna, dzięki której powstaje dojrzały mRNA
transport mRNA z jądra komórkowego do cytoplazmy
rozpoznanie mRNA przez rybosom i translacja białka
degradacja mRNA
fałdowanie białka (nabywanie struktury trzeciorzędowej białka)
modyfikacje posttranslacyjne, np. glikozylacja, fosforylacja
przemieszczenie białka do właściwej pozycji (np. błony komórkowej, mitochondrium, etc.) (może poprzedzać poprzedni proces)
funkcjonowanie białka - często najbardziej długotrwały i praktycznie jedyny etap w którym uwidacznia się biologicznie, fenotypowo, informacja genu.
degradacja białka
16. MODEL OPERONU JAKO PROKARIOTYCZNA JEDNOSTKA TRANSKRYPCJI
to grupa dwóch lub więcej genów strukturalnych tj. kodujących białka lub RNA, poprzedzonych wspólną sekwencją promotora (miejscem wiązania polimerazy RNA) i wspólnym operatorem (sekwencją DNA, do której się wiąże białko regulatorowe umożliwiające lub blokujące przyłączanie się polimerazy RNA do promotora). Kodowane przez osobny gen białko regulatorowe pełni odpowiednio rolę aktywatora lub represora transkrypcji podległego mu operonu. Np. operon laktozowy (szlak kataboliczny), tryptofanowy (szlak anaboliczny).
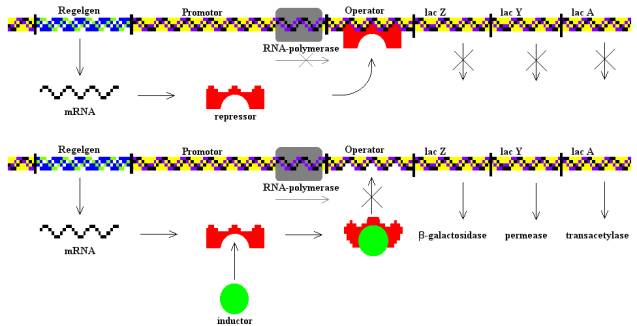
17. TRANSKRYCJA I JEJ ROLA W EKSPRESJI GENU
Etap najważniejszy!!
Podlega największej kontroli
Zachodzi tu: splicing (wycinanie intronów), tzw. redagowanie RNA (zmiany sekwencji nukleotydów przez inwersję lud delecję), tasowanie egzonów (duplikacja lub insercja egzonów)
Produktami są mRNA, rRNA i tRNA.
Geny tzw. HOUSEKEEPING --> stale aktywne nie podlegają regulacji lub ewentualnie bardzo małej (geny związane z cyklem Krebsa lub tworzeniem rybosomów) natomiast geny wyciszone lub hamowane podlegają ścisłej regulacji na poziomie zarówno transkrypcji jak i translacji.
18. SPOSOBY PRZENOSZENIA MATERIAŁU GENETYCZNEGO
transformacja bakteryjna --> komórka AKTYWNIE pobiera DNA z otoczenia (może być naturalna lub indukowana)
transdukcja -->wymiana DNA między bakteriami za pośrednictwem bakteriofagów
transfekcja --> wprowadzania obcego DNA do komórki
w obecności fosforanu wapnia lu dekstranu
drogą elektroporacji
drogą bombardowania kulkami złota lub wolframu
drogą mikroiniekcji
drogą lipofekcji
przez infekcję zmodyfikowanym wirusem
przez fuzję protoplastów --> tworzenie hybryd nawet między równymi gatunkami
19. KOD GENETYCZNY
trójkowy --> odczytywany na ślepo trójkami (=kodonami)
uniwersalny --> dla Procaryota i Eucaryota (wyj.! -> UGA to zwykle kodon STOP ale w mitDNA to Trp)
bezprzecinkowy --> od miejsca początku odczytywania - AUG (metionina) leci ciągle
niezachodzący --> poszczególne trójki polimeraza odczytuje tylko raz i po kolei...wiadomo :)
jednoznaczny --> dane kodon = zawsze ten sam aminokwas
kolinearny --> liniowe kodony w DNA odpowiadają w mRNA kolejności aminokwasów zakodowanych
KODONY STOP --> UAG, UGA, UAA
REDUNDACJA --> oznacza, że istnieje nadmiar info. genetycznej w rzeczywistym kodzie, czyli wiele kodonów ma to samo znaczenie (patrz wyżej :))
20. MUTACJE
def --> to nagłe, skokowe, trwałe, dziedziczne zmiany informacji genetycznej. Mogą zachodzić w kodującej i niekodującej części genomu.
M. spontaniczne --> zachodzą bez udziału środowiska zewn., np. przez pomyłkę polimerazy DNA
M. indukowana --> przez zadziałanie mutagennych czynników środowiska
M. mutatorowe --> zwiększające prawdopodobieństwo następnych mutacji np. takie, które dotyczą mutacji polimerazy
GENOMOWE (aberracje ilościowe)
Euploidie --> zmiana liczby całego zestawu chromosomów. Gdy mamy więcej niż dwa genomy w komórce mówimy i poliploidalności (haploidy n, diploidy 2n, triploidy 3n, tetraploidy 4n itd.)
- autopoliploidy --> zwielokrotnienie tego samego homologicznego zestawu chromosomów u osobnika np. 2n+2n=4n, u człowieka letalne
- allopoliploidy --> na skutek podwojenia liczby zestawu chromosomów niehomologicznych, 2n1+2n2=4n, u człowieka nie występuje to, dotyczy głownie krzyżowaniu roślin
Aneuploidie --> zwiększenie lub zmniejszenie liczby chromosomów w pojedynczej parze chromosomów chomologicznych przez nondysjunkcję chromosomów w podziale.
- nullisomia --> brak chromosomów jednej pary 2n-2
- monosomia --> brak jednego chromosomu danej pary 2n-1
- trisomia --> dodatkowy chromosom danej pary 2n+1
- tetrasomia --> dodatkowe dwa chromosomydanej pary 2n+2
--> u człowieka:
- zespól Pataua 47 XX (lub 47 XY) + 13 -> trisomia
- zespól Edwardsa -----//------- + 18 -> trisomia
- zespół Downa ------//------ + 21 -> trisomia
- zespól Turnera 45 X -> monosomia X u kobiet
- zespól Klinefertera 47 XXY -> trisomia X u mężczyzn
PUNKTOWE (dotyczące pojedynczych zasad) I CHROMOSOMOWE (aberracje strukturalne)
substytucje --> podstawienie zasady purynowej (A i G) na inną purynową = TRANZYCJA lub podstawienie purynowej przez pirymidynową (T i C, U) i na odwrót = TRANSWERSJA
insercje --> wstawienie jednego lub kilku nukleotydów
delecje --> pominięcie jednego lub kilku nukleotydów lub w przypadku chromosomów - pęknięcie i wypadnięcie części chromosomu
duplikacja --> zwielokrotnienie fragmentu chromosomu
inwersja --> pęknięcie fragmentu chromosomu w dwóch miejscach i złej naprawie przez obrót o 1800 i wstawieniu w to miejsce.
translokacje --> przeniesienie odcinków chromosomów pomiędzy chromosomami niehomologicznymi
EFEKTY MUTACJI (wg Prof. lecę)
letalne recesywne lub dominujące --> nie trzeba tłumaczyć
ciche--> czyli produkt genu jest bez zmian lub brak patologii
- synonimiczne -> gdzie nowy kodon koduje ten sam aminokwas
- neutralne -> nie maja wpływu na negatywna zmianę fenotypu
niesynonimiczna (zmiany sensu) --> nowy kodon koduje nowy aminokwas i białko mamy inne
nonsensowna --> powstaje w nowym miejscu kodon terminacyjny (STOP) i powstaje krótsze zwykle niefunkcjonalne białko
Najgorzej, gdy mutacja zajdzie w środkowym -> 2 nukleotydzie kodonu, ponieważ nukleotydy 1 i 3 są częściej naprawiane w procesie rewersji i jest powrót do typu dzikiego/półdzikiego.
Szybkie i wolne kodony dotyczą sekwencji nukleotydów w DNA --> dużo GC w DNA powoduje dużo szybkich genów (musiała być jakaś ewolucyjna selekcja w tym kierunku)
MUTAGENY
promieniowanie jonizujące X --> pękanie chromosomów
promieniowanie UV --> dimeryzacja pirymidyn (T z C lub T z T), w świetle naprawia to FOTOLIAZA, w ciemności system SOS
zw. chemiczne tj.:
- HNO2 --> powoduje deaminację zasad i np.
- EMS (tlenki etylenu) i in. czynniki alkilujące --> mutacje typu tranzycji lub transwersji oraz typu zmiany ramki odczytu
- NH2OH --> jednokierunkowa tranzycja C w T
analogi zasad np. 5-bromouracyl to T, 2-aminopuryna to A
barwniki akrydynowe --> płaskie pierścienie powodujące m.in. delecje lub insercje zasad
21. DROŻDZE JAKO DOSKONAŁY :) OBIEKT BADAŃ GENETYCZNYCH
łatwość hodowli w czystych kulturach w warunkach laboratoryjnych
krótki cykl życiowy
szybki wzrost
zdolność do rozmnażania bezpłciowego --> ogromne populacje identyczne genetycznie się uzyskuje i płciowego --> do krzyżowania
stabilny stan haploidalny i diploidalny
mały genom i poznana funkcja 66% genów!
niepatogenność
organelle komórkowe, regulacja ekspresji genów, białka komórki -> typowe dla Eucaryota --> przełożenie na człowieka!
wydajna rekombinacja
proste wykonanie testów komplementacji na alleliczność mutacji przez krzyżowanie form haploidalnych
można indukować heterologiczną ekspresje genów --> wydajna transformacja plazmidami
oddychanie tlenowe (jak człowiek :)) i beztlenowe (jak komórki nowotworowe)
łatwość manipulacji genetycznych i indukcji mutacji
doświadczenia nie są etycznie niepewne :) i wiele wiele innych cudownych cech ;)
22. APARAT GENETYCZNY KOMÓREK DRODŻY SACCHAROMYCES CEREVISIAE
GENOM JĄDROWY -->
12 200 kB
16 chromosomów, ok. 6,5 tys. otwartych ramek odczytu,
występują też w chromosomach ruchome fragmenty tzw. -> retrotranspozony
60-100 kopii 2-mikronowego plazmidu o funkcji nieznanej, dziedziczą się niezgodnie z Mendlem.
GENOM CYTOPLAZMATYCZNY -->
głównie RNA dwuniciowe wirusowe, koduje np. toksynę KIL do niszczenia innych grzybów,
dziedziczenie niezgodne z Mendlem
GENOM MITOCHONDRIALNY
to ok. 10% info. genetycznej drożdży
kolista (bywa liniowa), dwuniciowa cząsteczka DNA
75-85 kB
35,5 % kodujące z tego -> 16 % egzony a 15 % introny (8-9 genów koduje białka, 25 koduje tRNA, i 3 geny podzielone
15,2 % - ORF-y intronów (?)
bogate w AT
szybko ewoluuje (brak systemów naprawczych)
UGA to tutaj nie kodon STOP lecz koduje on tryptofan!
3 geny są PODZIELONE --> maja INTRONY!! (tzw. mobilne)
- introny gr.I --> przenoszony w miejsce genu bezintronowego dzięki kodowanej przez ten intron endonukleazie i naprawę tego naciętego miejsca na matrycy genu z intronem -> INTRON HOMING
- intron gr.II --> kodują w sobie mutarazę, endonukleazę, odwrotna transkryptazę -> RETROHOMING
dziedziczenie niezgodne z Mendlem
23. GENETYCZNA DETERMINACJA ENZYMÓW ŁAŃCUCHA ODDECHOWEGO
Mitochondralne mutanty oddechowe : (dziedziczenie niemendlowskie!!!!)
- rho0, rho-, mit- (wywoływana przez mangan)--> oddychają tylko beztlenowo więc NIE są zdolne do wzrostu na niefermentowalnym źródle węgla np. mleczan, etanol, glicerol
- rho+ --> dziki typ mtDNA --> oddychają tlenowo
Jądrowe mutanty oddechowe (dziedziczenie Mendlowskie!!!!)
- pet --> uszkodzona biogeneza i funkcja mitochondriów, odychanie beztlenowe
- PET --> typ dziki, tlenowiec
--> CHLORAMFENIKOL -> hamuje jądrowa biosyntezę białka
--> CYKLOHEKSIMID -> hamuje cytoplazmatyczna biosyntezę
24. DZIEDZICZENIE CYTOPLAZMATYCZNE NA PRZYKŁADZIE DROŻDŻY I INNYCH ORGANIZMÓW
u S. cerevisiae --> patrz pyt. 23 -> mutanty oddechowe
Euglene viridis --> samożywna lub heterotroficzna w zależności czy ma chloroplasty czy nie (dziedziczenie mateczne!!)
Chlamydomonas .......... --> brak chloroplastów jest letalny
Paramecium aurelia (pantofelek) --> ma w cytoplazmie jakieś cząstki kappa odpowiedzialne za produkcję paramycyny (dziedziczenie infekcyjne) lub cząstki „mu” odpowiedzialne za zabicie partnera podczas koniugacji
przejaśnienia liści u roślin ozdobnych (związane z funkcjonalnością chloroplastów)
Zea mays (kukurydza) --> gen męskiej sterylności, który współpracuje z jakimś jądrowym genem w wytworzeniu tej cechy (dziedziczenie infekcyjne)
Drosophila melanogaster --> dziedziczenie infekcyjne związane z wirusem sigma -> dotyczy nadwrażliwości na obecność CO2
Limneae peregra (błotniarka) --> skręt muszli tego ślimaka jest dziedziczony mateczne -jest determinowany przez genotyp matki, która wyposaża cytoplazmę komórki jajowe w mRNA
Drosophila melanogaster --> rozwój przedniej części ciała kontrolowany jest przez transkrypty matczynego genu bicoid
25. TYPY MUTACJI MITOCHONDRIALNYCH ??????????
matczyne --> u organizmów, u których komórka jajowa jest duża a plemnik mały -> zwierzęta, rośliny kwitnące
matczyne mitt. i plastydów. u kukurydzy
matczyne -mitt i ojcowskie lub oburodzicielskie plastydów
ojcowskie mitt i matczyne plastydów u banana
ojcowskie mitt. i plasydów u sekwoi wiecznozielonej
selektywne niszczenie, np. u Chlamydomonas w zygocie selektywnie niszczone są mitochondria tylko jednego typu koniugacyjnego
selektywna transmisja --> u omułka osobniki dziedziczą mitt. od obojga rodziców, ale do męskich potomków przekazują jedynie ojcowskie mitt. a do żeńskich jedynie matczyne mitt.
26. MITOCHONDRIALNE CHOROBY GENETYCZNE
--> efekty mutacji narastają z wiekiem!!
ZESPÓL LHON (=LEBERA) --> ślepota w 30-40 roku życia
ZESPÓL KSS --> multisymptomatyczny paraliż mięśni zewn. oka, wada serca, wada pigmentu siatkówki
ZESPÓŁ MELAS-tRNA --> encefalopatia z kwasicą mlczanową, częściowa ślepota
ZESPÓŁ MERRF --> padaczka miokloniczna z obecnością włókien poszarpanych
ZESPÓL NARP dot. podjednostki w ATP-azie --> neuropatia, ataksja, barwnikowe zwyrodnienie siatkówki
Czyli głownie dotyczą układu mięśniowego i nerwowego i mimo, że jest to dziedziczenie mateczne to najczęściej chorują mężczyźni
27. TRANSPOZYCJA I JEJ MECHANIZMY NA PRZYKŁADZIE BAKTERII
def. --> wewnątrzgenomowe przeniesienie części materiału genetycznego międzu chromosomami niehomologicznymi dzięki specjalnym strukturom DNA --> transpozonom (np. tzw. skaczące geny u kukurydzy); możliwe dzięki transpozazie, która nacian dany odcinek DNA. Zmiana info. genetycznej może być na - lub +. Rola --> szubka adaptacja do zmian środowiska --> ewolucja molekularna.
rodzaj rekombinacji nieuprawnionej
model transpozycji Shapiro --> zakłada istnienie układu enzymatycznego transpozazy, który nacina sekwencje CTGG - powtórzenia na obu końcach transpozonów
28. REKOMBINACJA IN VITRO I IN VIVO
in vivo --> w komórce “na żywo” :)
- crossing-over
- losowe ułożenie chromosomów w płytce metafazowej i tym samym ich losowe rozchodzenie sie do komórek potomnych
- losowe łączenie się gamet
- transpozycja
in vitro --> dotyczy inżynierii genetycznej, to łączenie poza komórką fragmentów DNA pochodzących z różnych organizmów i wprowadzenie ich do układu, gdzie może zachodzić replikacja tego DNA. Np. oczywiście komórki drożdży mogą służyć jako gospodarz dal takich rekombinantów :).
29. MOLEKULARNE PODSTAWY REKOMBINACJI DNA IN VITRO
poznanie struktury przestrzennej DNA i danych sekwencji genowych
poznanie mechanizmów ekspresji + jej regulacja (ważna rola promotora w heterologicznej rekombinacji!)
wykrycie, izolacja i charakterystyka enzymów tj. polimerazy, endonukleazy, ligazy...itd
uzyskiwanie kolejnych mutantów mikroorganizmów, owadów + opracowanie map genetycznych
opracowanie sposobu przenoszenia genów in vitro
- przy transformacji trzeba zapewnić stan kompetencji; u Procaryota niezbyt duże stęż. DNA w środowisku a u Eucaryota odwrotnie -> wysokie stęż. DNA, aby zwiększyć wydajność procesu.
- mikroiniekcje --> służą nawet do wnikania do organelli
- ...itd --> patrz pyt. 18
rozwój wiedzy o chemii białek, ich strukturze i funkcji jako produktów genowych w komórce
30. SYSTEM RESTRYKCJI I MODYFIKACJI W KOMÓRCE BAKTERYJNEJ
stwierdzony tylko u Procaryota
niezbędne 2 typy enzymów:
RESTRYKTAZA -> jest to endonukleaza rozpoznająca odpowiednie frgamenty i nacina wzdłuż ich osi symetrii (czasem powstają tzw. lepkie końce --> dot. klasy II restryktaz, ale nie zawsze, zmienna jest tez długość rozpoznawanych fragmentów w zależności od rodzaju restryktazy); służy to do obrony komórki przed fagowym DNA
METYLAZA -> uniemożliwia strawienie ciętych nici i chroni tym samym własne DNA przed strawieniem go po cięciu przez restryktazy. Ale któż to wie jak rozpoznaje własne od fagowego :)
np. EcoR I, Hind III --> enzymy restrykcyjne
def --> transportery obcego DNA do komórki biorcy w celu replikacji
31. WEKTORY
układ cis i trans dwóch mutacji u heterozygoty
1 jednostka mapowa =1% crossing-over = 1% rekombinacji = 1 centimorgan (cM)
Wyszukiwarka
Podobne podstrony:
wybrane zagadnienia na exam - sciaga - mac, Studia, Zastosowanie statystyki w zarzadzaniu
Kopia od Piskurewicza podane zagadnienia na exam (pelne), Pedagogika UKSW
SOP-zagadnienia na exam, Dokumenty STUDIA SKANY TEXT TESTY, ADMINISTRACJA UNIWEREK WROCŁAW MAGISTER,
Zagadnienia na egzamin Genetyka i Biotechnologia, Genetyka
Hopej zagadnienia na egzamin 10 r
Zagadnienia na kolokwium z podstaw marketingu i badan rynkowych 10, socjologia 3 rok, Marketing
zagadnienia na egzamin lekarski 2009-10, BIOFIZYKA
Zagadnienia na zaliczenie wykładów, semestr IV, genetyka, wykłady plus zagadnienia na test u Ciesiel
maszyny, aaa, studia 22.10.2014, Materiały od Piotra cukrownika, Maszyny elektryczne, zagadnienia na
Prawo karne, Egzamin ustny z prawa karnego(1), Zagadnienia na egzamin ustny z prawa karnego (9 i 10
10 LIRYKA, Filologia polska UWM, Teoria literatury, zagadnienia na egzamin
Zagadnienia na egzamin z Biologii i Genetyki 2012, biologia wyklady
Genetyka. 201415 Zagadnienia na zaliczenie wykładów, Studia Ochrona Środowiska, Genetyka, Zaliczeni
Dziedzic Letka, inżynieria genetyczna L, opracowane zagadnienia na zaliczenie
Zestaw 10, Opracowane zagadnienia na egzamin
Zagadnienia na konstytucjyne (10 16)
zagadnienia na ang EXAM
2011 WSEH 10 Miedzynarodowa olimpiada jezyka angielskiego zagadnienia na III etap
Opracowanie Zagadnień na egzamin Mikroprocki
więcej podobnych podstron