Zestaw zagadnień na egzamin z Botaniki Leśnej
1.Co to jest histologia, embriologia, kariologia, cytologia, palynologia, morfologia, fizjologia roślin
Histologia (z gr. histos - utkanie) - nauka o rozwoju, budowie i funkcjach tkanek. W rzeczywistości zakres zainteresowań histologii bywa traktowany szerzej, poza histologią ogólną obejmuje też histologię szczegółową (anatomię mikroskopową) - naukę o mikroskopowej budowie narządów oraz cytologię i embriologię.
dział botaniki, nauka wyodrębniona z morfologii roślin, badająca budowę i rozwój zarodka roślinnego w okresie jego życia w osłonie tkanek organizmu macierzystego; obejmuje również proces zapłodnienia, tworzenie się, budowę i rozwój narządów generatywnych i (u okrytonasiennych) bielma. Zasadnicze dla współczesnej e.r. odkrycia zapoczątkował G.B. Amici, wyjaśniając ok. 1824 rolę pyłku kwiatowego w tworzeniu się nasienia; następnie W. Hofmeister wyjaśnił (po kilkunastu latach prac) pojęcie gametofitu; 1870 J. Hanstein opisał segmentację zarodków, 1884 E. Strasburger zapłodnienie komórki jajowej, 1898 S.G. Nawaszyn podwójne zapłodnienie u roślin okrytonasiennych. Współczesna e.r. dzieli się na porównawczą (opisową) i eksperymentalną, zajmującą się m.in. kontrolą zapłodnienia i hodowlą zarodków in vitro.
Kariologia (z greckiego karyon "jądro"), dział cytologii zajmujący się budową jądra komórkowego, a szczególnie budową chromosomów.
Cytologia, inaczej biologia komórki (z gr. kytos i logos) - nauka o budowie wewnętrznej i funkcji podstawowej jednostki budulcowej organizmów żywych jaką jest komórka. Każdy organizm roślinny i zwierzęcy składa się z komórek. Istnieją organizmy zbudowane z jednej tylko komórki - jednokomórkowce i organizmy składające się z wielkiej liczby komórek - wielokomórkowce.Cytologia jest blisko związana z nauką o tkankach - histologią.Cytologia zajmuje się organellami komórkowymi: jądrem, mitochondrium, wakuolami, retikulum endoplazmatycznym i innymi.
Palinologia jest nauką o pyłkach roślin i zarodnikach grzybów. palinologia, palynologia [gr. palýnein `sypać', `przyprószyć', lógos `nauka'], bot. najmłodszy dział botaniki, zajmujący się badaniem ziaren pyłku i zarodników roślin, zarówno współcześnie żyjących, jak i kopalnych; wzięła pocz. z pyłkowej analizy (palinologicznej analizy).
Morfologia roślin (gr. morphē = kształt, logos = nauka) - jedna z najstarszych gałęzi wiedzy o roślinach, nauka zajmująca się badaniem zewnętrznej budowy roślin, zależnościami jakie między nimi występują, zachodzącymi w nich przemianami, a także badaniem ich pokroju, ukształtowania poszczególnych organów.Podczas gdy morfologia zajmuje się badaniem zewnętrznej budowy roślin, anatomia roślin bada ich budowę wewnętrzną.Morfologia roślin posiada wydzielone działy:
Fizjologia roślin, fitofizjologia - nauka zajmująca się badaniem procesów życiowych roślin oraz funkcjami ich organów. Badane procesy to m.in.: wzrost, odżywianie, oddychanie i rozmnażanie. Jest jedną z gałęzi botaniki i fizjologii. Fizjologia roślin zajmuje się badaniem zjawisk i procesów zachodzących na róznych poziomach organizacji biologicznej:
na poziomie molekularnym - syntezy i łańcuchy reakcji biochemicznych zachodzących w komórce
na poziomie struktur komórkowych - transport związków chemicznych i przekazywanie sygnałów w organizmach roślinnych
na poziomie organów
na poziomie organizmu
na poziomie zbiorowiska roślinnego.
Znajomość procesów zachodzących w roślinach wykorzystywana jest praktycznie w takich dziedzinach jak: rolnictwo, leśnictwo i ogrodnictwo.
2. Budowa komórki roślinnej - podstawowe organele
Opis: Budowa komórki roślinnej: A) ściana komórkowa, B) plasmodesma, C) błona komórkowa, D) chloroplast, E) błona tylakoidu, F) mitochondrium, G) lizosom, H) aparat Golgiego, I) wakuola, J) retikulum endoplazmatyczne gładkie, K) retikulum endoplazmatyczne szorstkie, L) jądro, M) błona jądrowa, N) otwór w błonie jądrowej, O) jąderko
Opisy organeli :
Kształt komórki determinuje ściana komórkowa, która dodatkowo chroni komórkę przed pęknięciem w wyniku zwiększonego napływu wody do jej wnętrza. U bakterii właściwych (czyli także sinic) zbudowana jest z biopolimeru peptydowo-wielocukrowego - mureiny, zaś u archeanów (archeabakterii) głównym jej składnikiem jest pseudomureina lub białka ułożone w tzw. warstwę S. Część archeanów i wszystkie mikoplazmy (grupa bakterii) nie posiadają ściany komórkowej.U bakterii grubość ściany komórkowej warunkuje, jaki będzie rezultat barwienia metodą Grama i de facto jest podstawą klasyfikacji bakterii na Gram-dodatnie i Gram-ujemne. Te pierwsze (G+) mają ścianę o grubości 15-50 nm, zaś drugie (G-) kilkukrotnie cieńszą, 2-10 nm. Różnica ta pociąga za sobą także odmienności w fizjologii i wrażliwości na leki między obiema grupami bakterii.
Plasmodesma, plazmodesma - połączenie międzykomórkowe występujące w komórce roślinnej, pasma cytoplazmy i siateczki śródplazmatycznej (retikulum endoplazmatyczne) przechodzące przez szczeliny (jamki) w ścianie komórkowej. Plasmodesmy łączą ze sobą protoplasty komórek, które sąsiadują ze sobą. Dzięki nim komórki w łatwy sposób mogą wymieniać pomiędzy sobą różne substancje. Plazmodesmy są odpowiednikami złączy szczelinowych w komórkach zwierzęcych.
Błona komórkowa (plazmolemma) otacza całą komórkę. U eukariontów posiadających ścianę komórkową zawsze występuje po stronie wewnętrznej tej ściany. Plazmolemma zbudowana jest podobnie, jak u bakterii właściwych: składa się z dwóch warstw fosfolipidów oraz zanurzonych w nich białek. W budowie lipidów błonowych komórek jądrowców dominują nienasycone kwasy tłuszczowe. Znaczny (5-25%) jest także udział cholesterolu[10].
Białka zanurzone w plazmolemmie pełnią funkcje receptorów, białek kanałowych, czy enzymatycznych, które odpowiadają za pobieranie wody, soli mineralnych i substancji odżywczych, wydzielanie substancji na zewnątrz (np. produktów przemiany materii), obieranie bodźców ze środowiska zewnętrznego, itd. Ponadto struktury białek wraz z skoordynowanymi innymi cząsteczkami, tak zwany glikokaliks, komórkom bardziej złożonym organizmów nadaje tożsamość antygenową, co warunkuje m.in. występowanie różnych grup krwi[11].
Chloroplast - otoczone podwójną błoną białkowo-lipidową organellum komórkowe występujące u roślin i glonów eukariotycznych). Są rodzajem plastydów. Zawierają zielone barwniki chlorofile pochłaniające energię światła słonecznego potrzebną do fotosyntezy. W nich zachodzi przemiana dwutlenku węgla oraz wody z wykorzystaniem energii świetlnej w glukozę oraz tlen.
Chloroplasty są otoczone dwiema błonami o różnej przepuszczalności, otaczającymi stromę wypełniającą wnętrze chloroplastu. Błona zewnętrzna dobrze przepuszcza jony. Wewnętrzna błona jest natomiast słabo przepuszczalna i tworzy liczne woreczki (zwane tylakoidami). W chloroplastach granalnych ułożone są one w płaskie stosy zwane tylakoidami gran. U chloroplastów bezgranalnych natomiast występują jedynie lamelle - tylakoidy stromy, czyli tylakoidy rozciągnięte wzdłuż całego chloroplastu (w komórkach pochew okołowiązkowych niektórych roślin przeprowadzających fotosyntezę C4[1] i glonów.
Wnętrze chloroplastu wypełnia białkowa substancja - stroma - koloid białkowy. W jej skład wchodzą m.in. niewielkie ilości DNA, enzymy biorące udział w fotosyntezie oraz rybosomy typu prokariotycznego (tzw. rybosomy małe, o stałej sedymentacji 70s), które biorą udział w produkcji białek, są one jednak mniejsze od rybosomów eukariotycznych o stałej sedymentacji 80s, znajdujących się w cytoplazmie.
Chloroplasty zaliczamy do organelli samoreplikujących się (obecność chloroplastowego DNA oraz zdolność do samoreplikacji uważane są przez wielu naukowców za dowody na endosymbiotyczne pochodzenie tych organelli od innych organizmów, które zostały pochłonięte przez komórkę, lecz zamiast ulec strawieniu, przekształciły się w symbionty).
Tylakoid - pęcherzykowata struktura, podstawowy element budowy wewnętrznej chloroplastu komórki roślinnej lub podobna struktura wchodząca w skład samożywnej komórki prokariotycznej. W chloroplastach tylakoidy tworzą tzw. system lamellarny, w którego skład wchodzą grana, czyli zwarte stosy spłaszczonych tylakoidów oraz intergrana (tylakoidy stromy), czyli pojedyncze pęcherzyki tylakoidów łączące grana.
W błonach tylakoidów zachodzi faza jasna fotosyntezy, podczas gdy w stromie zachodzi ciemna faza fotosyntezy.Błona tylakoidu zbudowana jest głównie z glikolipidów, a dokładniej galaktolipidów takich jak monogalaktozylodiacyloglicerol i digalaktozylodiacyloglicerol, w których skład wchodzi wiele kwasów nienasyconych. Fosfolipidy stanowią tylko 15% procent wszystkich lipidów chloroplastowych. W błonie występuje także duża ilość różnego rodzaju białek. Charakterystycznym elementem budowy błon tylakoidów są barwniki fotosyntetyczne, np. chlorofile.Podobieństwo tylakoidów prokariotów i tych, które wchodzą w skład chloroplastów, sugeruje, że chloroplasty to prokarioty, które wniknęły do wnętrza przodków dzisiejszych eukariotów i weszły z nimi w związek symbiotyczny.
Mitochondrium (w liczbie mnogiej mitochondria; dawniej chondriosom[1]) - organellum komórki eukariotycznej pochodzenia endosymbiotycznego, w którym zachodzą procesy będące głównym źródłem energii (w postaci ATP) dla komórki, w szczególności proces fosforylacji oksydacyjnej, zachodzący w błonie wewnętrznej mitochondriów.
Mitochondria posiadają własny genom. Genom mitochondriów jest nieduży - koduje tylko od kilkunastu do kilkudziesięciu białek z kilkuset białek niezbędnych do funkcjonowania mitochondrium.
Przeciętna komórka eukariotyczna zawiera od kilkuset do kilku tysięcy mitochondriów. W komórkach ameby Chaos chaos L. stwierdzono 500.000 mitochondriów[2]. Przyrost ich liczby jest możliwy dzięki zdolności tego organellum do podziału przebiegającego podobnie jak podział wolno żyjących bakterii.
Mitochondrium otaczają dwie błony białkowo-lipidowe, obie podobne w budowie do zwykłej błony komórkowej, ale o bardzo różnych właściwościach. Błona zewnętrzna otaczająca całe organellum jest naszpikowana białkami zwanymi porynami. Poryny są w istocie dużymi (około 2-3 nm średnicy) kanałami, przez które mogą się przedostawać wszystkie cząsteczki o masie nie przekraczającej 6000 daltonów. Większe cząsteczki mogą pokonać zewnętrzną błonę tylko przy pomocy transportu aktywnego. Połowę masy zewnętrznej błony stanowią fosfolipidy. Wśród białek ją budujących są enzymy odpowiadające za bardzo rozmaite reakcje, jak np. wydłużanie łańcuchów kwasów tłuszczowych, utlenianie adrenaliny i rozkład tryptofanu.Enzymem markerowym (markerem) błony zewnętrznej jest oksydaza monoaminowa (MAO). Enzymem markerowym (markerem) przestrzeni międzybłonowej jest kinaza adenilanowa.Błona wewnętrzna nie zawiera poryn i jest nieprzepuszczalna - transport jonów i innych cząsteczek do jej wnętrza wymaga specjalnych transporterów błonowych. Umożliwia to wytworzenie gradientu protonowego niezbędnego do działania łańcucha oddechowego. Błona wewnętrzna zawiera ponad 100 rozmaitych polipeptydów, a białka stanowią ponad ¾ jej masy. Są to białka łańcucha oddechowego, syntaza ATP wytwarzająca ATP w macierzy mitochondrialnej oraz białka transportujące metabolity do wnętrza macierzy i na zewnątrz. Szczególnym fosfolipidem w błonie jest kardiolipina, lipid typowy dla komórek bakteryjnych. Enzymem markerowym (markerem) błony wewnętrznej jest oksydaza cytochromu c.Błona wewnętrzna tworzy wpuklenia - grzebienie mitochondrialne, w których zakotwiczone są enzymy łańcucha oddechowego. Wpuklenia zwiększają powierzchnię błony - i tak np. w mitochondriach wątroby powierzchnia wewnętrznej błony mitochondrialnej jest pięciokrotnie większa od powierzchni zewnętrznej błony mitochondrialnej.Wnętrze mitochondrium wypełnia macierz (matriks) mitochondrialna. Jest to rodzaj żelu - wodny roztwór białek i metabolitów zużywanych na potrzeby mitochondrium. Białkami wewnętrznymi mitochondrium są wszystkie enzymy β-oksydacji kwasów tłuszczowych, cyklu Krebsa, syntezy steroidów itp. Macierz zawiera również mitochodndrialny DNA (mtDNA), rybosomy mitochondrialne i tRNA mitochondrialne. Enzymem markerowym (markerem) matrix mitochondrialnej jest syntetaza cytrynianowa.
Lizosom - (występuje wyłącznie w komórkach eukariotycznych) niewielkie (0,05-0,5 μm) pęcherzyki zawierające enzymy rozkładające białka, kwasy nukleinowe, węglowodany i tłuszcze. Łącznie w lizosomach jest obecnych ok. 40 różnych hydrolaz. W lizosomie zachodzi nie tylko proces trawienia komórkowego wchłoniętych pokarmów, ale także rozkład niepotrzebnych już cząsteczek. Nazwę lizosom wprowadził de Duve w roku 1955, wcześniej opisane były także przez Miecznikowa (1865)
Substancje, które mają ulec strawieniu, docierają do lizosomu w pęcherzykach, w które zostają "ubrane" w trakcie pinocytozy i fagocytozy.
Błona lizosomów zawiera liczne monotopowe glikoproteidy (o długich łańcuchach i dużej ilości kwasów sialowych), liczne pompy protonowe oraz liczne białka przenośnikowe.
Rodzaje lizosomów:
trawienne - rozkład substancji,
magazynujące - magazynowanie substancji,
W lizosomach rozkładane są na ogół białka długowieczne w odróżnieniu od proteosomów.
Enzymem markerowym (markerem) lizosomów jest kwaśna fosfataza.
Aparat Golgiego - organellum występujące niemal we wszystkich komórkach eukariotycznych, służące chemicznym modyfikacjom substancji zużywanych przez komórkę, bądź wydzielanych poza nią. Podstawową jednostką strukturalną aparatu Golgiego jest diktiosom.
W strukturach Golgiego odbywa się:
Struktury błoniaste są strukturami dynamicznymi, odbywa się między nimi przepływ substancji zawartych wewnątrz kanałów i pęcherzyków (tutaj opatrzonych płaszczem koatomerowym z białek COPI) oraz błon.
W obrębie diktiosomu wyróżnia się dwa bieguny:
biegun cis (formowania)
biegun trans (dojrzewania)
Wakuole (wodniczki) - struktury komórkowe występujące u roślin i niektórych pierwotniaków i w komórkach zwierzęcych. W komórkach zwierzęcych występuje wiele małych wodniczek (wakuol), natomiast w roślinnych jedna lub kilka dużych. U pierwotniaków występują wodniczki tętniące, odpowiedniki wakuoli, ale od wakuoli o wiele mniejsze. Zajmują do 90% komórki. Ze starzeniem się komórki zachodzi proces rozrośnięcia się i zamienienia wakuoli w jedną wodniczkę.
Funkcje wakuol
magazyn wody w komórce
czasami trawienie wewnątrzkomórkowe - wakuole mogą zawierać enzymy hydrolityczne biorące udział w procesie trawienia
Retikulum endoplazmatyczne, siateczka śródplazmatyczna, siateczka wewnątrzplazmatyczna, ER - wewnątrzkomórkowy i międzykomórkowy system kanałów odizolowanych od cytoplazmy podstawowej błonami (membranami) biologicznymi. Tworzy nieregularną sieć cystern, kanalików i pęcherzyków. Siateczka śródplazmatyczna jest szczególnie rozbudowana w komórkach, w których zachodzi intensywna synteza białek.
W procesie wirowania frakcjonującego lizatu komórkowego błony retikulum endoplazmatycznego tworzą frakcję mikrosomalną.
Rozróżnia się dwa typy retikulum:
Funkcje ER
Jądro komórkowe - organellum znajdujące się w każdej komórce eukariotycznej (wyjątek to komórki tracące jądro w procesie dojrzewania, np. erytrocyty ssaków, czyli ciałka czerwone lub zrogowaciałe komórki naskórka; takie komórki nie są w stanie rozmnażać się i szybko tracą zdolność do życia).
Zwykle w komórce znajduje się jedno jądro (monokariocyty), ale spotykane są też komórki dwujądrzaste (dikariocyty), np. hepatocyty oraz wielojądrzaste (polikariocyty), np. osteoklasty. Większość jąder ma średnicę 3,5 - 20 mikrometrów[1].
Jądro komórkowe posiada własną otoczkę, mieści się zwykle w pobliżu środka komórki i przeważnie ma kształt kulisty, może jednak być również podłużne, owalne, itp. Występuje w nim w postaci ziarenek substancja silnie barwiąca się barwnikami zasadowymi, zwana chromatyną. Wewnątrz jądra znajduje się maleńkie ciałko zwane jąderkiem.Rolą jądra komórkowego jest przechowywanie informacji zawartej w DNA, jej powielanie w procesie podziału komórki, a także kontrolowanie całości metabolizmu komórki dzięki kopiowaniu fragmentów DNA (kopiami są tu odcinki RNA) odpowiednich dla syntezy potrzebnych enzymów czy innych cząsteczek. Jądro otoczone jest podwójną białkowo-lipidową błoną. Poprzez pory w tejże błonie do cytoplazmy przenoszone są tylko fragmenty RNA; DNA nie opuszcza jądra.
Błona jądrowa, otoczka jądrowa, kariolemma - podwójna błona białkowo-lipidowa odgraniczająca wnętrze jądra komórkowego od cytoplazmy.
W obrębie otoczki jądrowej wyróżnia się:
wewnętrzną błonę jądrową,
zewnętrzną (cytoplamzatyczną) błonę jądrową,
przestrzeń okołojądrową (perynuklearną), która znajduje się pomiędzy dwoma błonami,
pory jądrowe - umożliwiają bierną i czynną wymianę makromolekuł z cytoplazmą. Z jądra do cytoplazmy przenikają, między innymi, kompleksy rybonukleoproteinowe mRNA oraz dojrzałe rybosomy. Do jądra wnikają, między innymi, białka syntetyzowane w cytoplazmie. Transport molekuł o masie atomowej powyżej ~60 kilodaltonów odbywa się aktywną drogą z zużyciem energii poprzez hydrolizę ATP. Cząsteczki mniejsze mogą przenikać do jądra drogą swobodnej dyfuzji.
blaszkę jądrową (lamina) - ma grubość 10-100 nm, przylega do wewnętrznej błony jądrowej i składa się z sieci delikatnych włókienek białkowych, utworzonych przez białka klasy lamin. U ssaków wyróżnia się laminę A, B oraz C. Struktura i skład aminokwasowy lamin jest podobny do filamnetów pośrednich. Blaszka jądrowa nadaje odpowiedni kształt jądru komórkowemu oraz uczestniczy w organizacji strukturalnej chromatyny - jest miejscem umocowania pętli chromatynowych. Laminy zaangażowane są również w proces fragmentacji i odbudowy otoczki w czasie podziału mitotycznego.
Pory jądrowe - otwory w podwójnej błonie jądrowej, służące do transportu cząsteczek z jądra komórkowego do cytoplazmy albo w przeciwnym kierunku.
W obrębie poru jądrowego znajduje się jądrowy kompleks porowy o średnicy 120-150 nm. Liczba jądrowych kompleksów porowych jest zależna od wieku, aktywności metabolicznej i typu komórki. W hepatocytach znajduje się 3000-7000 porów, a w komórkach o wysokiej aktywności metabolicznej liczba ta sięga 50 mln. Przeciętnie w komórkach eukariotycznych jest 10-20 porów/mm².
Jąderko - ultraelement jądra komórkowego odpowiedzialny za syntezę RNA, głównie rRNA. Jakościowo stanowi zagęszczenie chromatyny. W trakcie podziału komórkowego jąderko zanika. Można to uzasadnić zablokowaniem transkrypcji genów kodujących rRNA, ponieważ wtedy chromosomy ulegają kondensacji.
Jąderko tworzone jest z obszaru jąderkotwórczego (NOR), u człowieka występuje 10 NOR-ów (znajdujących się na ramionach krótkich chromosomów par 13, 14, 15, 21 i 22), a np. u świni 4 NOR-y.
3. Ściana komórkowa a błona komórkowa (różnice w budowie i funkcji)
Funkcje ściany komórkowej
Ogranicza wzrost komórki
Chroni przed urazami mechanicznymi
Zabezpiecza przed nadmiernym parowaniem
Nadaje kształt i sztywność komórce
Chroni przed utratą wody
Przepuszcza substancje
układają się równolegle do siebie i prostopadle do osi podziału komórki
Pęcherzyki zlewają sie ze sobą, tworząc trój-warstwową przegrodę w obrębie dzielącej sie komórki, zwanej przegrodą pierwotną. Warstwa środkowa otoczona jest 2 błonami plazmatycznymi.
Celuloza zostaje wbudowana przez aparat Golgiego do środka przegrody pierwotnej. Jest ona odkładana od wewnętrznej strony warstwy środkowej i po zewnętrznej stronie błon plazmatycznych. Tak powstaje ściana komórkowa pierwotna. Ma ona wbudowaną do środka blaszkę środkową, która spaja 2 części ściany.
Występowanie błon komórkowych jest istotne dla funkcjonowania i życia komórki. Najstarszš ewolucyjnie jest funkcja izolacyjna błony. Uformowana błona oddzielała różne substancje organiczne od otaczajšcego rodowiska. Umożliwiało to stopniowš organizację komórki oraz przebieg reakcji chemicznych między czšsteczkami odseparowanymi od rodowiska zewnętrznego.
Błony wewnštrzkomórkowe umożliwiajš z kolei istnienie organelli komórkowych oraz pełnienie różnych funkcji przez te organelle. Błony wewnštrzkomórkowe powodujš istnienie przedziałów wewnštrz komórki, różnišcych się składem chemicznym i właciwociami. Dzięki temu komórki wykazujš przystosowanie do okrelonych czynnoci w obrębie narzšdów lub tkanek.
Błony komórkowe cechuje wybiórcza przepuszczalnoć, zatem izolacja komórki od rodowiska zewnętrznego oraz utrzymanie stałego składu chemicznego poszczególnych przedziałów nie jest zupełne. Błony jedynie ograniczajš wpływ oddzielajšcych je rodowisk.
Poznawanie budowy i funkcji błon komórkowych stanowi jeden z ważniejszych problemów współczesnej biochemii. W ostatnich latach powięcono wiele uwagi błonom komórkowym i przeprowadzono wiele badań dotyczšcych zarówno struktury, jak i transportu przez błony. Sš to badania z pogranicza chemii i fizyki, bardzo trudne, a jednoczenie bardzo doniosłe. Skażenie rodowiska naturalnego, wprowadzanie do organizmu ksenobiotyków oraz proces starzenia się powodujš uszkodzenie struktury czšsteczek tworzšcych błony komórkowe, a tym samym uszkodzenie błon. Znajomoć tych zagadnień jest niezbędna we współczesnej medycynie i biologii.
Budowa blony komorkowej:
Zazwyczaj inne białka występują po wewnętrznej, a inne po zewnętrznej stronie błony. Cząsteczki należące do błony mogą z łatwością poruszać się wewnątrz swojej warstwy (dyfuzja lateralna, o ile nie są związane na przykład od wewnątrz z białkami cytoszkieletu), jednak napotykają duże trudności z przejściem do warstwy przeciwnej.Podobna asymetria dotyczy także samej błony, która ma zazwyczaj odmienny skład (różne proporcje, ale i jakość) lipidowy w swojej monowarstwie wewnętrznej i zewnętrznej.Asymetria jakościowa rozmieszczenia lipidów może także dotyczyć płaszczyzny błony - w monowarstwach istnieją lokalne obszary o składzie odbiegającym od rozkładu przypadkowego. Są to tak zwane "rafty". Są one bogatsze od sąsiednich obszarów monowarstwy w specyficzne lipidy, cholesterol czy białka. Lipidy znajdujące się w takich domenach mogą być poniżej temperatury głównego przejścia fazowego i nie mieć struktury ciekłokrystalicznej, co powoduje ich agregację. Funkcje "raftów" oraz sposoby ich powstawania w błonach nie są jeszcze dokładnie znane, ale nie wyklucza się ich interakcji i wpływania na aktywność białek błonowych czy udział w fuzjach błon.
4. Jądro komórkowe, jego budowa i funkcje
Funkcje jądra
zawiera informację genetyczną (informację o budowie wszystkich białek jakie komórka jest w stanie wytworzyć) zakodowaną w DNA
steruje biosyntezą białka
przekazuje informację genetyczną komórkom potomnym w procesach podziałów
Struktura wewnętrzna jądra
Zasadniczymi elementami budującymi jądro komórkowe to skomplikowana i ważna funkcjonalnie otoczka jądrowa, całe wnętrze zwane nukleoplazmą, a składa się na nią całe wnętrze jądra (z wyjątkiem odrębnego jąderka), gdzie znajduje się materiał genetyczny i dzieli się na kilka rejonów o odmiennej morfologii. Są to: region skondensowanej chromatyny, region okołochromatynowy oraz przestrzeń międzychromatynowa. Wyróżnia się dodatkowo w obrębie nukleoplazmy jąderko, które traktowane jest jako osobna struktura. Odbywa się tam synteza rybosomowego RNA oraz dojrzewanie i montowanie podjednostek rybosomalnych.
W jądrze komórkowym odbywają się procesy replikacji DNA, transkrypcja i procesy potranskrypcyjne. Wszystkie te procesy wiążą się ze strukturą wewnętrzną jądra. W nukleoplazmie syntezowany jest także heterogenny jądrowy RNA oraz mały jądrowy RNA. To ostatnie badania wykazują, że ultrastruktura jądra komórkowego jest znacznie bardziej skomplikowana niż wydawało się to do tej pory. Wraz z pojawieniem się mikroskopu elektronowego możliwe było opisanie ziarnistości międzychromatynowych i okołochromatynowych. W 1969 roku wyróżniono w jądrze nowe regiony, odrębne morfologicznie: włókna perichromatynowe (okołochromatynowe) zawierające RNP (rybonukleoproteina) i ciała zwinięte.
Ultrastruktura jądra komórkowego w różnych komórkach może się nieco różnić, ale zasadnicze elementy budowy pozostają niezmienione. Można w nim wyróżnić cztery takie regiony:
- otoczka jądrowa, składająca się z podwójnej błony białkowo-lipidowej z kompleksami porowymi i blaszką jądrową, która podściela wewnętrzną błonę jądrową i ma postać delikatnej, włóknistej warstwy
- heterochromatyna, która znajduje się pod otoczką w postaci splątanych nici oraz heterochromatyna okołojąderkowa,
- jąderko, które stanowi kupienie specyficznych rejonów chromatynowych
- przestrzeń międzychromatynowa
Jądro komórkowe przechodzi liczne zmiany struktury, które związane są z fazą cyklu komórkowego. Cykl komórkowy można podzielić na dwie zasadnicze fazy. Jest to podział komórki (mitoza) oraz interfaza, czyli ten czas, kiedy komórka nie dzieli się. Dodatkowo interfazę dzieli się jeszcze na trzy fazy. Pierwszą z nich jest faza G1, w której komórka dojrzewa i intensyfikacji ulegają wszystkie procesy metaboliczne. Druga z nich to faza S, w której następuje replikacja materiału genetycznego zawartego w jądrze. Ostatnią fazą przed podziałem komórkowym jest faza G2, w której komórka przygotowuje się do podziału i sprawdza, czy wszystkie procesy związane z replikacją zaszły prawidłowo. Po fazie G2 następuje podział komórki.
Kiedy rozpoczyna się podział komórki zmienia się także struktura wewnętrzna jądra. Zanika otoczka jądrowa, rozproszeniu ulega także jąderko, chromatyna ulega kondensacji, w wyniku czego powstają charakterystyczne struktury, czyli chromosomy. Ich liczba jest charakterystyczna dla gatunku. Podczas podziału ułożone są parami, po dwa takie same chromosomy, które później rozchodzą się do dwóch różnych komórek. Najlepiej widoczne są one w metafazie podziału mitotycznego, kiedy tworzy się tzw. płytka metafazowa (chromosomy ułożone są parami w płaszczyźnie równikowej komórki). Chromatyna w czasie interfazy nie jest dobrze widoczna w jądrze, ponieważ jej nici są bardzo cieniutkie, ale DNA jądrowe nigdy nie wydostaje się poza jądro.
5. Chromosomy, ich rola
Chromosom - forma organizacji materiału genetycznego wewnątrz komórki. Nazwa pochodzi z greki, gdzie χρῶμα (chroma, kolor) i σῶμα (soma, ciało). Chromosomy rozróżniano poprzez wybarwienie.
Chromosomy występują w formie mikroskopijnej struktury najlepiej widocznej w metafazie podziału komórkowego, kiedy to są najbardziej skondensowane. Chromosomy są zbudowane z dwóch chromatyd siostrzanych (podłużnych jego części) połączonych w jednym punkcie centromerem (wyjątkiem są chromosomy powstałe po pęknięciu centromeru w trakcie podziału jądra komórkowego - pod koniec metafazy). U organizmów prokariotycznych chromosom stanowi pojedyncza, kolista cząsteczka DNA, natomiast u organizmów eukariotycznych liniowa cząsteczka DNA. Każda cząsteczka DNA buduje jedną chromatydę. Zarówno u prokariotów, jak i eukariontów chromosomy zbudowane są z kompleksu DNA i białek histonowych lub histonopodobnych (u prokariotów). W komórkach organizmów prokariotycznych i niektórych eukariotycznych (drożdże, pierwotniaki) występują również nieosłonięte, koliste cząsteczki DNA zwane plazmidami.
U organizmów eukariotycznych chromosomy z obu stron zakończone są powtarzającą się sekwencją nukleotydów tworzących telomer. Skracanie telomerów podczas podziałów komórki być może prowadzi do starzenia się komórki.
Struktura chromosomu nie jest niezmienna, podlega on bowiem zmianom zwanym mutacjami. Mutacje dotyczące bezpośrednio chromosomów to aberracje chromosomowe lub mutacje genomowe.
Chromosomy dzielą się na autosomy - zawiadujące dziedziczeniem cech nie sprzężonych z płcią, oraz chromosomy płciowe - czyli allosomy lub heterosomy, których obecność przejawia się u konkretnej płci i w wielu przypadkach determinuje ją.
Liczba chromosomów może być różna (np. 1 para) aż do 100 par, ale zazwyczaj wynosi kilka do kilkudziesięciu par (4 pary u muszki owocowej, 20 par u myszy, 23 pary u człowieka, 24 pary u szympansa, 39 u psa). Liczba autosomów jest cechą charakterystyczną gatunku, a jej fluktuacje prowadzą do powstawania nowych gatunków (patrz: specjacja).
Komórki mogą być:
U gatunków rozmnażających się bezpłciowo każda komórka organizmu ma tę samą liczbę chromosomów.
U gatunków rozmnażających się płciowo występują komórki zarówno haplo- jak i diploidalne. W przypadku wielu organizmów, w tym zdecydowanej większości kręgowców, liczba chromosomów w komórkach somatycznych jest dwa razy większa (diploidalna) niż w gametach (haploidalna). Do powstania haploidalnych gamet dochodzi w wyniku mejozy. Podział komórek somatycznych (diploidach) zachodzi na drodze mitozy, w której najpierw dochodzi do podwojenia materiału genetycznego.
W przypadku innych organizmów, takich jak np. rośliny lądowe, występuje przemiana pokoleń - pokolenie haploidalne występuje po pokoleniu diploidalnym. Są one przeważnie bardzo od siebie odmienne.
Chromosomy determinują płeć ,
7. Tkanki roślinne, podział
Tkanki roślinne - zespoły komórek o podobnej budowie, określonych czynnościach i wspólnym pochodzeniu występujące u roślin naczyniowych. Tu też tradycyjnie zaliczane są niektóre tkanki wielokomórkowych brunatnic zaliczanych obecnie do protistów.
Tkanki roślinne dzieli się na dwie podstawowe grupy: tkanki twórcze (tzw. merystemy), w których komórki dzielą się intensywnie i są niezróżnicowane, oraz tkanki stałe o komórkach zróżnicowanych funkcjonalnie.
Substancje, z których powstają tkanki roślin pobierane są zarówno z powietrza jak i z gleby. Atomy węgla i tlenu, będące składnikami wszystkich tkanek roślinnych, pobierane są z powietrza w postaci dwutlenek węgla. Inne niezbędne pierwiastki wśród których jest azot, fosfor, potas, siarka, wapń, magnez oraz pewna ilość pierwiastków śladowych, korzenie rośliny pobierają z gleby w postaci związków mineralnych. Roślina przetwarza te substancje nieorganiczne w związki organiczne, z których buduje swoje tkanki.
Histologia roślin wyróżnia co najmniej kilkadziesiąt szczegółowych typów tkanek roślinnych. Poniższa lista nie wyczerpuje zagadnienia:
pierwotne:
wtórne:
8. Tkanki twórcze, funkcje i podział
Tkanka twórcza, merystem to tkanka roślinna, w której zachodzi intensywny proces podziału komórek. U większości roślin takie organy, jak łodyga, liście, rozwijają się z małego, centralnego skupiska komórek - merystemu. Każdy zawiązek nowego organu wyrasta w innym kierunku w stosunku do zawiązka, który pojawił się wcześniej.
Budowa
zbudowana jest z komórek żywych, które przez całe życie zachowują zdolność podziału,
komórki tej tkanki są niewielkie, ściśle do siebie przylegające, z dużymi centralnymi jądrami komórkowymi, małymi wodniczkami i cienkimi celulozowymi ścianami,
Merystemy dzielimy na:
9. Tkanki okrywające
Tkanka okrywająca - jedna z tkanek roślinnych, dzieli się na skórkę (tkanka żywa) i korek (tkanka martwa). Skórka występuje na liściach i łodygach (w postaci epidermy) oraz na korzeniach (w postaci epiblemy inaczej zwanej ryzodermą). Tkankę okrywającą dzieli się na pierwotną i wtórną.
Budowa
Tkanka okrywająca składa się z żywych, prostokątnych komórek, ściśle przylegających do siebie. Występuje w niej kutykula, włoski mechaniczne oraz aparaty szparkowe. Nie posiada chloroplastów, dzięki czemu światło przenika do wewnątrz. Fellogen składa się z komórek merystematycznych,które dzielą się odkladając na zewnątrz nowe komórki korka (fellem) i od czasu do czasu również do wnętrza jedną lub kilka warstw żywych komórek o cechach komórek miękiszowych (felloderma). Owe trzy warstwy : felloderma, fellogen i martwe komórki korka tworzą korkowicę.
Funkcje
Funkcją tej tkanki jest oddzielenie środowiska zewnętrznego od środowiska wewnętrznego, ochrona przed czynnikami zewnętrznymi i utratą wody oraz udział w wymianie gazowej.
Formy ochrony:
wytworzenie kolców, kutneru,
wydzielanie substancji drażniących, kłujących, zapachowych.
10. Tkanki przewodzące
Tkanka przewodząca - tkanka roślinna, w której odbywa się transport wody wraz z rozpuszczonymi w niej substancjami do wszystkich części roślin, zbudowana jest z niejednorodnych komórek.
Dzieli się ją na:
martwy ksylem (drewno), przewodzący wodę i rozpuszczone w niej sole mineralne z korzeni do innych części rośliny. W funkcji tej wyspecjalizowały się dwa rodzaje elementów drewna: cewki (tracheidy), charakterystyczne dla paprotników i roślin nagonasiennych, i naczynia (tracheje), występujące u okrytonasiennych.
U roślin wyższych występują naczynia:protoksylem - składa się z naczyń o umocnieniach pierścieniowych i spiralnych;
metaksylem - (drabinkowate) 90% transportu wody (jeśli roślina nie ma przyrostu wtórnego)
ksylem wtórny - mają ściany jajkowate (w roślinie, która posiada przyrost wtórny)
żywy floem (łyko), przewodzący rozpuszczone produkty asymilacji z liści w głąb rośliny, zbudowany z bezjądrowych komórek sitowych tworzących rurki sitowe (sitowe elementy roślin), a także z komórek miękiszowych i włókien wzmacniających. U roślin okrytonasiennych występują ponadto wyspecjalizowane tzw. komórki towarzyszące, pełniące rolę pomocniczą w przewodzeniu substancji pokarmowych przez rurki sitowe.
11. Tkanki asymilacyjne
Miękisz asymilacyjny (inaczej: miękisz zieleniowy, chlorenchyma) - tkanka roślinna, wyspecjalizowany miękisz, złożony z komórek zawierających liczne soczewkowate chloroplasty. Jest tkanką żywą. Zachodzi w nim proces fotosyntezy, a przez to odżywianie rośliny. Występuje głównie w liściach i młodych łodygach. W liściach występuje najbliżej jego powierzchni. Jego komórki nie są jednorodne. Wyróżnić można w jego obrębie miękisz palisadowy, miękisz gąbczasty i miękisz wieloramienny.
12. Tkanki miękiszowe
Miękisz, tkanka miękiszowa, parenchyma[1] (gr. parénchyma - miąższ) - jednorodna tkanka roślinna, która wypełnia znaczną część organizmów roślin. Zbudowana z żywych, zwykle dużych (0,05 - 0,5 mm) i cienkościennych komórek, o ścianach celulozowych, rzadko drewniejących, z dużą wakuolą otoczoną cytoplazmą. Protoplast jest mało wyspecjalizowany. Charakterystyczną cechą miękiszu jest występowanie przestworów międzykomórkowych. Komórki miękiszu zachowują zdolność do podziałów i odróżnicowania, dzięki czemu odgrywają istotną rolę w zjawiskach regeneracyjnych.
Miękisz jest tkanką występującą we wszystkich częściach ciała rośliny i wchodzi w skład wszystkich tkanek złożonych. Tkanki miękiszowe pełnią w roślinie zasadnicze czynności fizjologiczne przemiany materii, uczestniczą w fotosyntezie, oddychaniu, osmozie, transpiracji, gromadzą także substancje zapasowe i wodę. Mimo że miękisz zbudowany jest z cienkościennych komórek, w stanie turgoru pełni jednak w roślinach istotną funkcję mechaniczną.
Istnieje kilka kryteriów podziału tkanek miękiszowych. Ze względu na strukturę tkanki wyróżnia się:
Ze względu na kryterium pochodzenia wyróżnia się miękisz pierwotny, powstający z merystemu pierwotnego oraz miękisz wtórny, powstający z merystemu wtórnego.
Ze względu na pełnione funkcje wśród tkanek miękiszowych wyróżnia się:
Poza tym w zależności od specjalizacji mówić można o miękiszu wydzielniczym, taninowym, wyścielającym itp.
W końcu biorąc pod uwagę to, jakich tkanek złożonych miękisz jest częścią wyróżnia się:
13. Tkanki mechaniczne
Tkanki mechaniczne, lub inaczej wzmacniające, nadają roślinie sztywność, chronią przed działaniem wiatrów, ulewnych deszczy i śniegu, jak również utrzymują ją w pozycji pionowej. Tkanki mechaniczne mają błony komórkowe zgrubiałe. Wyróżniamy następujące trzy rodzaje tkanek: twardzicę, zwarcicę i komórki kamienne. Twardzica jest zbudowana z silnie wydłużonych martwych komórek, przybierających postać włókien. Nadaje ona sztywność oraz dużą wytrzymałość wielu roślinom, (len, konopie, drzewa). Zwarcica jest tkanką żywą nie drewniejącą. Komórki jej mają błony z celulozy tylko częściowo zgrubiałe. Występuje ona tylko u niektórych roślin, głównie u dwuliściennych. Komórki kamienne wchodząc w skład ochrony okryw owoców i nasion, np. pestek brzoskwiń i śliw. Czasem występują one pojedynczo lub w małych grupach wśród tkanki miękiszowej (np. w owocu gruszy).
14. Tkanki i utwory wydzielnicze
Tkanka - zespół komórek o podobnej budowie, określonych czynnościach, wspólnym pochodzeniu i ich wytworów (substancja międzykomórkowa) o podobnym pochodzeniu, budowie, przemianie materii i przystosowanych do wykonywania określonej funkcji na rzecz całego organizmu. Tkanki występują u zwierząt tkankowych (tkanki zwierzęce) oraz u roślin wyższych (tkanki roślinne). Tkanki są elementami składowymi narządów i ich układów. Dział biologii zajmujący się tkankami to histologia.
Komórki odpowiedzialne za wykonywanie funkcji zwane są miąższem, zaś komórki będące rusztowaniem - zrębem tkanki/narządu.
Uwzględniając cechy morfologiczne i fizjologiczne zespołów komórkowych tworzących tkanki, wszystkie tkanki dzieli się na cztery grupy:
Zarówno u zwierząt, jak i u roślin, występuje tkanka nabłonkowa oraz łączna i podporowa. Do tkanek wyłącznie zwierzęcych należy tkanka mięśniowa i nerwowa.
Twory wydzielnicze. Pojedyncze komórki i ich zespoły, które wydzielają w dużych ilościach specyficzne substancje, mogą występować na powierzchni roślin - są to wytwory skórki - lub w jej wnętrzu - różnicują się z komórek miękiszowych. Mimo różnego pochodzenia, maja wspólne cechy, jak np. gęstą cytoplazmę i dobrze rozwiniętą siateczkę sródplazmatyczną, co świadczy o intensywnej przemianie materii, produkowane substancje są wydzielane ma zewnątrz komórki drogą egzocytozy albo gromadzą się w wodniczkach na terenie komórki.
Wytworami skórki są włoski wydzielnicze, np. produkujące silnie pachnące związki lotne (olejki eteryczne), które mają znaczenie obronne, odstraszają roślinożerców, a czasem są dla niech trujące. Inne działanie maja włoski parzące, np. u pokrzywy. Rośliny okrytonasienne mają w kwiatach miodniki - utwory produkujące słodki płyn, zwabiający zapylaczy. Błyszczące włoski na liściach roślin owadożernych wydzielają enzymy trawiące ciała owadów, rośliny te, żywiąc się owadami, uzupełniają niedobór azotu. Utwory wydzielnicze znajdują się także wewnątrz ciała roślin, jak np. rury mleczne i kanały żywiczne. Rury mleczne to żywe, wielojądrowe komórki, a sok mleczny to zawartość ich wodniczek, jest on zawiesiną białek, cukrów, żywic, gum, olejków eterycznych itd., z soku mlecznego kauczukowca (Hevea brasiliensis) otrzymuje się kauczuk naturalny, a z pigwicy (Achras sapota) - gumę chicle, używaną do produkcji gumy do żucia. Różne substancje, np. żywice, mogą być także wydzielane do przestworów miedzykomórkowych, w ten sposób powstają kanały żywiczne u sosny albo jodły, sok mleczny i żywica mają znaczenie obronne - zawarte w nich związki chronią roślinę przed roślinożercami.
15. Łodyga, jej budowa i funkcje
Łodyga - nadziemna, osiowa część rośliny naczyniowej, która wraz z umiejscowionymi na niej liśćmi, pąkami, kwiatami i owocami stanowi pęd. Powstaje w wyniku podziałów i różnicowania komórek stożka wzrostu.
bierze udział w procesie fotosyntezy.
Warunki bytowania determinują w znacznej mierze funkcje łodygi, w tym głównie stosunek funkcji mechanicznej do transportowej, np. układ mechaniczny łodyg roślin wodnych jest przystosowany do falowania i prądów, brak lub znacznie ograniczony jest natomiast transportu wody; w przypadku łodyg podziemnych (kłącza, rozłogi) zarówno funkcja mechaniczna jak i transportowa jest znacznie ograniczona. Łodygi nadziemne (zarówno pnące jak i wolno stojące) muszą być przystosowane zarówno do sprawnego transportu wody i substancji organicznych, jak i do znoszenia naprężeń, wygięć i skręceń mechanicznych
Anatomia
{kind=link}
Przekrój przez łodygę: 1 - rdzeń, 2 - protoksylem, 3 - drewno (ksylem), 4 - łyko, 5 - sklerenchyma, 6 - kora pierwotna, 7 - epiderma
Zewnętrzną warstwę łodygi stanowi pokryta kutykulą skórka pędu (epiderma), w której znajdują się aparaty szparkowe. Przykładem rośliny jednoliściennej o łodydze pełnej jest kukurydza zwyczajna Zea mays. Pod skórką znajduje się cienka warstwa sklerenchymy, a pod nią cienki miękisz asymilacyjny. Większość jednoliściennych nie ma wtórnych tkanek twórczych (kambium i felogenu), w związku z czym zachowują przez całe życie budowę pierwotną. Tkanki zwykle nie są zróżnicowane na korę pierwotną i walec osiowy. Wnętrze łodygi wypełnia miękisz zasadniczy (bezzieleniowy), w którym rozproszone nieregularnie są wiązki przewodzące (taki rozproszony układ wiązek to ataktostela). Są to wiązki kolateralne zamknięte (bez przyrostu wtórnego.
Inny typ budowy anatomicznej łodygi reprezentuje np. pszenica zwyczajna Triticum aestivum. Pod epidermą znajdują się niewielkie kupienia miękiszu asymilacyjnego, poprzedzielane skupieniami sklerenchymy, w obrębie których znajdują się wiązki przewodzące kolateralne zamknięte (na przemian większe i mniejsze). Wnętrze młodej łodygi wypełnia miękisz zasadniczy, który w miarę starzenia się rośliny stopniowo zamiera i kruszy się, pozostawiając pusty kanał wewnątrz łodygi (w międzywęźlu). Pozostałe tkanki drewnieją. Taka konstrukcja łodygi sprawia, że jest ona giętka i lekka[2].
Łodyga roślin dwuliściennych może mieć budowę pierwotną i wtórną. Budowa pierwotna występuje u wszystkich gatunków, budowa wtórna - u niektórych. W tym drugim przypadku budowa pierwotna ograniczona jest tylko do najmłodszych, wierzchołkowych części łodygi, budowa wtórna cechuje starsze części. Budowę pierwotną łodygi tworzą tkanki pierwotne, powstające w merystemie wierzchołkowym (stożku wzrostu pędu. Zewnętrzną warstwę łodygi stanowi pokryta kutykulą epiderma z aparatami szparkowymi. Pod skórką znajduje się kora pierwotna. Jej zewnętrzną część stanowi zazwyczaj tkanka wzmacniająca - kolenchyma albo sklerenchyma. Pod nią znajduje się warstwa miękiszu, w obwodowej części bogatego w chloroplasty (stąd zielony kolor łodygi). Ostatnią stanowi endoderma, której komórki zawierają ziarna skrobi lub wytwarzają pasemka Caspary'ego. Centralną część łodygi zajmuje walec osiowy. U niektórych roślin jego zewnętrzną część stanowi okolnica (perycykl). W walcu osiowym znajduje się tkanka przewodząca tworząca zwarty pokład lub ułożona w postaci wiązek poprzedzielanych tkanką miękiszową. Środek walca osiowego wypełnia rdzeń, najczęściej miękiszowy. Przyrost wtórny łodygi następuje wskutek działania merystemów bocznych, tj. miazgi i fellogenu. Miazga wytwarza na zewnątrz łyko wtórne, a do wnętrza - drewno wtórne.
16. Łodyga u traw i zbóż
Źdźbło - zielna łodyga roślin z rodziny wiechlinowatych (traw). Cechuje się dużą wytrzymałością mechaniczną, pomimo delikatnej i lekkiej budowy. Składa się z pustych międzywęźli oraz pełnych węzłów (kolanek).
Morfologia
Źdźbło jest obłe (w odróżnieniu od kanciastej łodygi wielu turzycowatych) i zwykle nagie. W dolnej części, w najniższych węzłach może się rozgałęziać i tu też wyrastają korzenie przybyszowe. W części środkowej i górnej źdźbło nie rozgałęzia się z wyjątkiem kwiatostanu (do rozgałęziających się wyjątków należy np. trzcinnik lancetowaty Calamagrostis canescens i bambusowe. Liście na źdźble wyrastają naprzeciwlegle w dwóch prostnicach. Liście obejmują węzły źdźbła swoimi pochwowatymi nasadami tworząc tzw. kolanko i chroniąc w ten sposób delikatną tkankę twórczą. W plemieniu bambusowych łodygi są zdrewniałe, u innych traw zielne.
Anatomia
Składa się z pustych międzywęźli oraz węzłów. W dolnych częściach międzywęźli znajduje się tkanka twórcza, dzięki której źdźbło może bardzo szybko rosnąć. Jest to tzw. merystem interkalarny. Jest to słaby punkt źdźbła - w okresie kłoszenia łatwo można rozerwać łodygę u nasady międzywęźli i wyciągnąć z pochwy liściowej. Wiązki przewodzące umieszczone są zwykle w źdźbłach w dwóch pierścieniach. Są to wiązki kolateralne zamknięte, w związku z czym łodygi nie mają możliwości przyrostu na grubość. Wiązki otoczone są dużą ilością tkanki mechanicznej - sklerenchymy, tworzącej wokół zewnętrznej części łodygi walec, nadający pędom traw dużą elastyczność i wytrzymałość. W plemieniu palczatkowych i kukurydzowych łodygi są pełne a wiązki przewodzące rozmieszczone są bezładnie. Epiderma (skórka) łodygi jest podobna do skórki liści. Niezróżnicowany miękisz asymilacyjny w młodych częściach źdźbła tworzy zwykle pasma biegnące wzdłuż łodygi.
17. Liść, jego budowa i funkcje
Liść - element budowy części osiowej (pędowej) roślin telomowych. Wyrastające z węzłów końcowe elementy rozgałęzień pędu, wyodrębniające się ze względu na funkcję i budowę od łodygi (nie mają np. zdolności do nieprzerwanego wzrostu). Pełnią głównie funkcje odżywcze i z tego powodu mają zwykle dużą powierzchnię umożliwiającą ekspozycję na odpowiednią ilość promieniowania słonecznego. Poza tym liście biorą udział w transpiracji i gutacji oraz wymianie gazowej. Nierzadko liście pełnią także funkcje spichrzowe, czepne, ochronne, obronne i pułapkowe, w takich przypadkach ulegając daleko idącym przystosowaniom w zakresie funkcji i budowy.
Morfologia
Większość liści składa się z różnokształtnej, zwykle spłaszczonej blaszki liściowej osadzonej bezpośrednio (liść siedzący, bezogonkowy) lub za pomocą ogonka liściowego (liść ogonkowy) na łodydze. Ogonek liściowy lub bezpośrednio blaszka liściowa umieszczone są na łodydze za pomocą tzw. nasady liścia. U niektórych roślin (np. u przedstawicieli rodzin wiechlinowate, rdestowate, selerowate) dolna część liścia rozrasta się w pochwę liściową. Pełni ona funkcję wzmacniającą łodygę (wiechlinowate), zwiększa powierzchnię asymilacyjną (selerowate) lub pełni funkcje spichrzowe i ochronne (cebula zwyczajna). U traw i turzycowatych liście na granicy pochwy i blaszki liściowej mają języczek liściowy. U nasady liści, zwłaszcza tych zaopatrywanych przez trzy ślady liściowe, wykształcają się przylistki (zwykle dwa). U niektórych roślin tropikalnych na końcu blaszki liściowej wykształcają się tzw. wylistki służące do odprowadzania wody z powierzchni liścia.
Liście asymilacyjne są zwykle spłaszczone grzbieto-brzusznie. Do wyjątków należą liście spłaszczone bocznie (np. u kosaćca Iris) lub cylindryczne (np. u wielu gatunków z rodzaju czosnek Allium). Dolna strona liścia zwana jest odosiową lub grzbietową (w pąku kwiatowym znajduje się na górze). Górna strona liścia zwana jest doosiową lub brzuszną[1].
Cechy takie jak nerwacja (użyłkowanie), czyli układ wiązek przewodzących i usytuowanie liścia na łodydze są często cechami charakterystycznymi gatunków, podobnie jak: kształt liścia, jego brzegu, nasady i zakończenia.
Inne funkcje i przystosowania liścia
U niektórych roślin liście, oprócz swojej podstawowej roli spełniają również inne funkcje:
Czepne. Liście niektórych roślin (np. groszku) przekształcone są w wąsy, które podobnie, jak ogonki liściowe powojników mogą owijać się wokół różnych przedmiotów.
Spichrzowe. Na przykład łuski cebuli.
Magazynujące wodę. Występują np. u "żywych kamieni" - Lithops.
Obronne. Liście przekształcone w ciernie (np. u kaktusa) lub kolce (np. u róży).
Wabiące. Zwabiają zwierzęta zapylające kwiaty.
Pułapkowe. Występują u roślin owadożernych, "trawiących" owady służące im za źródło az Liście są bardzo ważnymi dla rośliny organami. Ich główne funkcje to: przeprowadzanie fotosyntezy, transpiracja (czynne wyparowywanie wody) i wymiana gazowa. Niektóre liście dzięki przekształceniom mogą pełnić również inne funkcje.
18. Aparaty szparkowe, funkcje i zasada działania
Aparat szparkowy, szparka (gr. stoma jama, szpara) - struktury wielokomórkowe (aparat) lub dwukomórkowe (szparka) pochodzenia epidermalnego roślin służące do kontrolowanej wymiany gazowej między tkankami wewnętrznymi rośliny i atmosferą.
Funkcja i mechanizm działania
Szparki mają decydujące znaczenie dla funkcjonowania układu wentylacyjnego roślin pozwalającego na dostarczanie niezbędnego dla procesu fotosyntezy dwutlenku węgla. Dzięki nim rośliny mogą skutecznie pobierać ten gaz chroniąc się równocześnie przed szkodliwymi stratami wody w wyniku transpiracji. Funkcjonowanie szparek wiąże się ze zdolnością do zmiany kształtu komórek szparkowych. Komórki te poprzez zmianę turgoru, wykorzystując zjawisko osmozy, powodują zamykanie (obniżenie turgoru) lub otwieranie (podwyższenie turgoru) szparek. Skuteczność tego działania wspomagana jest przez nierównomierne zgrubienia ścian komórkowych, które są grubsze w środkowych odcinkach komórek szparkowych. Jeśli komórki mają dużą ilość wody, nadymają się i odciągają od siebie zgrubiałe ściany. Gdy następuje utrata wody, szparki się zamykają. Dawniej sądzono, że zmiany turgoru komórek szparkowych są skutkiem hydrolizy i syntezy skrobi w chloroplastach tych komórek. Obecnie wiadomo, że ruchy szparek powodowane są zmianą ciśnienia osmotycznego kationów potasu pobieranych przez komórki szparkowe przy jednoczesnym wydzielaniu protonów. Pobieranie jonów K+ zależne jest od stężenia dwutlenku węgla wewnątrz liścia (od intensywności fotosyntezy, czyli pośrednio od światła, zaopatrzenia w wodę, temperatury). Stopień otwarcia aparatów szparkowych jest także bezpośrednio regulowany przez obecność światła niebieskiego - receptorem jest zeaksantyna tylakoidów gran, która na niepoznanej dokładnie drodze powoduje fosforylację H+-ATPazy aktywując ten enzym. H+-ATPaza, dzięki energii z ATP wypompowuje protony wodoru na zewnątrz komórki, co umożliwia napływ kationów potasu do komórki poprzez kanały wpustowe. Równowaga jonowa utrzymywana jest przez aniony jabłczanowe i Cl-. Anion jabłczanowy powstaje przez hydrolizę skrobi, natomiast Cl- transportowany jest z komórek sąsiednich. Wzrastające stężenie jonów obniża potencjał wodny komórki szparkowej wywołując napływ wody z komórek o większym potencjale, napływająca woda zwiększa turgor, duży turgor jest przyczyną wyginania się komórek szparkowych i otwierania szparek.
19. Korzeń, jego budowa i funkcje
Korzeń - fragment sporofitu niemal wszystkich roślin naczyniowych, nieobecny jedynie u psylotowych posiadających chwytniki i zanikły wtórnie u niektórych roślin (np. u pływaczy i wolffii bezkorzeniowej). Dostarcza roślinom wodę i substancje odżywcze (sole mineralne), utrzymuje rośliny w glebie i służy do gromadzenia substancji zapasowych. Korzenie tworzą silnie rozczłonkowany układ penetrujący podłoże nazywany systemem korzeniowym. Stanowi on dolny biegun osiowych organizmów roślinnych, u których górny biegun to pęd. Rośliny zakorzenione są zwykle w glebie, u epifitów wykształcają się korzenie powietrzne, u hydrofitów korzenie zanurzone. Korzeń często mylony jest z kłączem stanowiącym podziemny fragment pędu. Korzenie w odróżnieniu od pędu nie wykształcają liści i rosną w dół (geotropizm), zwykle są niezielone, choć chlorofil mogą zawierać niektóre korzenie powietrzne.
Ze względu na sposób powstawania wyróżnia się korzenie pierwotne rozwijające się z zawiązka korzenia zarodkowego oraz korzenie przybyszowe powstające w części pędowej rośliny (na łodydze, kłączu lub nawet liściach).
Funkcje korzeni
Najważniejsze i najczęściej spotykane funkcje korzeni to dostarczanie roślinie wody wraz z substancjami odżywczymi oraz przytwierdzenie rośliny. Czasem pełnią one dodatkowo funkcję spichrzową, służą do rozmnażania wegetatywnego, rzadko odgrywają ważną rolę w układzie wentylacyjnym roślin lub nawet pełnią funkcję asymilacyjną.
Korzenie stosunkowo nieczęsto odgrywają istotną rolę w rozmnażaniu wegetatywnym, w naturze ustępując wśród organów podziemnych rozłogom i kłączom. Rośliny potomne mogą powstawać z odrostów korzeniowych (np. u osiki i robinii akacjowej), z bulw korzeniowych (np. dalia), czasem przystosowanych do rozmnażania w formie łatwo odpadających od rośliny macierzystej rozmnóżek (np. ziarnopłon wiosenny). Inaczej jest w przypadku działalności ogrodniczej, podczas której wykorzystywana jest powszechnie zdolność wytwarzania korzeni przybyszowych z fragmentów pędów do tworzenia sadzonek
FUNKCJE
- zaopatrywanie rośliny w wodę
- rozpuszczanie w niej sole mineralne czerpane z gleby
- przytwierdzanie rośliny do podłoża
- utrzymanie rośliny wpozycji pionowej
- starsze partie korzeni pełnią funkcje magazynów związków zapasowych
- niektórym roślinom korzenie służą do wegetatywnego rozmnażania
- korzenie oddechowe
- korzenie chłonące wodę i parę wodną z powietrza
- u niektórych roślin pełni jeszcze inne dodatkowe funkcje, np.: u burak, marchwi lub dalii jest organem spichrzowym, w którym roślina gromadzi substancje zapasowe
- korzeń pewnych storczyków mających bardzo silnie zredukowane liście spełnia rolę organu asymilacyjnego
- u lian bywa on organem czepnym
- korzenie kurczliwe, spotykane, np.: u lilii, mają zdolność- kurczenia się, wskutek czego mogą wciągać- roślinę głębiej w glebę
1. Budowa morfologiczna korzenia
Korzeń jest pierwszym organem, który rozwija się podczas kiełkowania
nasienia. Zawiązek korzenia w kiełkującym nasieniu wydłuża się i zagłębia
w ziemię. Daje on początek korzeniowi głównemu. Po pewnym czasie gdy
korzeń główny osiągnie odpowiednią długość i grubość, wyrastają korzenie
boczne, rozrastające się poziomo lub ukośnie w glebie (geotropizm dodatni).
Od korzeni bocznych wyrastają kolejno odgałęzienia dalszych rzędów.
Oprócz korzenia głównego i jego odgałęzień rośliny mogą wytwarzać tzw.
korzenie przybyszowe o pochodzeniu pozazarodkowym, wyrastające
z podziemnych lub nadziemnych części łodyg. Rozrastające się w glebie
korzenie tworzą system korzeniowy. Wyróżnia się dwa podstawowe typy
systemu korzeniowego: palowy i wiązkowy. System palowy (ekstensywny)
utworzony jest z korzenia głównego rosnącego pionowo w dół oraz
wyrastających z niego cieńszych i zwykle krótszych korzeni bocznych.
System wiązkowy utworzony jest z wielu korzeni o podobnej grubości,
mogących się rozgałęziać. W systemie wiązkowym brak korzenia głównego,
który wcześnie zanika, a jego miejsce zajmuje pęk korzeni przybyszowych.
Korzeń roślin nasiennych wydłuża się dzięki intensywnie dzielącym się
komórkom tkanki twórczej (merystematycznej) wierzchołka wzrostu
korzenia. Wierzchołek wzrostu korzenia okryty jest ochronną warstwą
komórek miękiszowych - czapeczką korzeniową, której zadaniem jest
ochrona konieczna w trakcie przeciskania się przez podłoże. Zewnętrzne
komórki czapeczki nieustannie obumierają i złuszczają się, a ich miejsce
zajmują nowe. W wyniku podziału komórek merystemu wierzchołkowego
powstają wszystkie tkanki korzenia. Korzeń wykazuje wyraźną budowę
strefową. Bezpośrednio za wierzchołkiem wzrostu (o długości ok.1mm),
który stanowi strefę podziałową korzenia, znajduje się strefa wydłużania
(elongacyjna). Strefa ta o długości 3 - 5 mm. Utworzona jest przez
niezróżnicowane, szybko rosnące komórki. Obie te strefy warunkują stałe
wydłużanie się korzenia. Za strefą wydłużania znajduje się
kilkucentymetrowej długości strefa dojrzewania komórek, zwana również
strefą włośnikową. W tej strefie następuje różnicowanie się komórki na
tkanki stałe. Strefa włośnikowa pełni główną funkcję korzenia - pobiera
wodę z rozpuszczonymi substancjami mineralnymi. Liczne komórki tkanki
okrywającej (skórki) mają tu kilkumilimetrowej długości nitkowate
uwypuklenia - włośniki. Włośniki okrywają korzeń widocznym gołym
okiem białym nalotem. W miarę dojrzewania włośniki górnej, starszej części
strefy obumierają i zanikają, na ich miejscu tworzą się nowe włośniki,
powstające na młodych dojrzewających komórkach. Za strefą włośnikową
rozpoczyna się strefa wyrośnięta korzenia z której wyrastają korzenie
boczne. Nie ma ona określonej długości, a u wielu roślin przyrasta również
na grubość. Strefa ta ma ostatecznie uformowane tkanki.
2. Budowa anatomiczna korzenia
Młode korzenie , począwszy od strefy włośnikowej wykazują w swojej
budowie trzy podstawowe systemy tkanek: okrywająca , leżąca pod nią kora
pierwotna oraz walec osiowy, zawierający wiązki przewodzące.
Skórka korzenia (epiblema) stanowi osłonę jego głębiej położonych
tkanek i tworzy włośniki. Zbudowana jest z jednej warstwy cienkościennych,
prostopadłościennych komórek
przystosowanych do pobierania wody.
Czynność te usprawniają włośniki. W starszych partiach korzeni roślin
dwuliściennych skórka obumiera i ulega złuszczeniu, a jej miejsce zajmuje
korek. Jest to martwa tkanka nie pobierająca wody pełniące funkcje
mechaniczne. Pod skórką znajduje się kora pierwotna, zbudowana
z komórek miękiszowych luźno ułożonych wskutek czego występują liczne
przestwory międzykomórkowe ułatwiające wymianę gazową. W młodych
korzeniach komórki kory pierwotnej przewodzą wodę z solami mineralnymi
ze skórki do wiązek przewodzących walca osiowego; w starszych korzeniach
pełnia one funkcje spichrzowe. Najbardziej wewnętrzna warstwa kory
pierwotnej granicząca z walcem osiowym to śródskórnia (entoderma).
Ściany jej komórek są zgrubiałe i skorkowaciałe, a same komórki są martwe,
ściśle do siebie przylegające. Tylko niektóre komórki pozostają żywe
i cienkościenne , są to komórki przepustowe umożliwiające transport wody
z kory pierwotnej do walca osiowego.
Pierwszą zewnętrzną warstwa walca osiowego jest okolnica (perycykl).
Z okolnicy biorą początek korzenie boczne. Wiązki przewodzące
zbudowane są z pasm wydłużonych komórek przewodzących wodę (drewno)
i asymilaty (łyko). Ułożone są one w walcu pierścieniowo. Pierścień tkanki
przewodzącej złożony jest z na przemian leżących pasm drewna i łyka.
Wiązki przewodzące asymilaty zbudowane są głównie z silnie wydłużonych,
żywych komórek, tzw. rurek sitowych. Stykają się one poprzecznymi
ścianami posiadającymi otworki, przez które przechodzą pasma cytoplazmy
łączące sąsiednie protoplasty i przewodzące substancje organiczne. Wiązki
przewodzące wodę utworzone są głównie z wydłużonych, martwych
komórek, miedzy którymi zanikły ściany poprzeczne. W ten sposób powstały
długie cienkie rurki zwane naczyniami. Ściany naczyń opatrzone są
spiralnymi lub pierścieniowatymi zgrubieniami, które nadają naczyniom
znaczną sztywność. Przedstawiona charakterystyka odpowiada budowie
pierwotnej korzenia. U roślin jednoliściennych korzenie zachowują przez
całe życie tkanki pierwotne dlatego nie wykazują przyrostu na grubość.
U roślin dwuliściennych przyrost korzenia na grubość jest wynikiem
funkcjonowania tkanek twórczych wtórnych. W starszych partiach korzenia
tworzą się tkanki twórcze wtórne: miazga (kambium) i miazga korkotwórcza
(felogen). Miazga powstaje miedzy wiązkami drewna i łyka. Jej warstwa
przebiega faliście, tak że pasma drzewne znajdują się po wewnętrznej stronie
miazgi, pasma łykowe zaś na zewnątrz. Falista początkowo linia miazgi
z czasem wyrównuje się i formuje pierścień. W kolejnych latach miazga
odkłada ku środkowi korzenia nowe słoje drewna wtórnego, na zewnątrz zaś
nowe warstwy łyka wtórnego. Równocześnie miedzy wiązkami
przewodzącymi miazga odkłada pasma miękiszu, tworząc tzw. promienie
rdzeniowe. Miękisz ten pełni rolę tkanki spichrzowej. Druga tkanka twórcza
korzenia - miazga korkotwórcza zawiązuje się w okolnicy. Wytwarza ona
warstwy korka czyli wtórnej tkanki okrywającej. Korek który powstaje
wokół walca osiowego, odcina korę pierwotną od odżywiających ją wiązek
przewodzących. Powoduje to obumarcie i złuszczanie się kory pierwotnej.
Korek staje się w ten sposób zewnętrzną tkanką korzenia. Korek, łyko i część
miękiszu stanowią korę wtórną. Korzeń o budowie wtórnej składa się
z dwóch warstw: kory wtórnej i drewna wtórnego. U różnych roślin te dwie
warstwy są rozwinięte w różnym stopniu. Wieloletnie i dwuletnie rośliny
zielne mają stosunkowo silnie rozbudowaną korę pierwotną. Ich korzeń jest
przeważnie organem zimującym, zawiera znaczne ilości materiałów
zapasowych
20. Wiązki łyko-drzewne otwarte a wiązki łyko-drzewne zamknięte (rola, różnice w budowie)
Wiązki przewodzące (w. łykodrzewne, w. sitowo-naczyniowe) - pasmo pierwotnej tkanki przewodzącej u roślin naczyniowych, składające się z części sitowej zwanej łykiem i części naczyniowej zwanej drewnem. Ich system rozciąga się od korzeni do łodyg i liści. U roślin wykazujących przyrost wtórny na grubość drewno i łyko są oddzielone cienką warstwą miazgi twórczej, która daje początek nowej tkance przewodzącej.
Łyko składa się z żywych komórek, jego zadaniem jest przewodzenie asymilatów od organów asymilujących do całej rośliny. Drewno składa się z komórek martwych i przewodzi wodne roztwory soli mineralnych od korzenia do liści, pełni ponadto funkcje wzmacniające.
Wiązki przewodzące mogą mieć różną budowę, w łodygach najczęstsze są tzw. wiązki kolateralne, w których łyko występuje po stronie zewnętrznej, a drewno - od wewnątrz w stosunku do osi łodygi.
Wiązki dzieli się także na otwarte (charakterystyczne dla roślin dwuliściennych), gdy między drewnem i łykiem występuje warstwa miazgi twórczej (kambium), umożliwiająca wtórny przyrost na grubość, oraz zamknięte, pozbawione kambium, występujące u roślin jednoliściennych.
U niektórych dwuliściennych wiązki przewodzące zawierają dwie warstwy łyka rozdzielonego drewnem (wiązki bikolateralne). Przy czym między łykiem zewnętrznym, a drewnem występuje miazga.
W korzeniach występują zwykle wiązki promieniste (radialne), o naprzemiennym układzie drewna i łyka, rozdzielonych pasmami miękiszu. W liściach wiązki przewodzące tworzą unerwienie liścia.
U roślin jednoliściennych wiązki rozrzucone są na całym przekroju łodygi, u roślin dwuliściennych i nagonasiennych ułożone są w regularny pierścień.
Tkanka przewodząca
- To tkanka stała, niejednorodna, która umożliwia roślinie przenoszenie substancji odżywczych: wody, soli mineralnych oraz produktów fotosyntezy na znaczne odległości - transport daleki.
- Komórki budujące tę tkankę mają wydłużony kształt, niewielką liczbę elementów komórkowych, których obecność utrudniałaby transport (niektóre pozbawione są ich w ogóle); tworzą zwarte układy biegnące wzdłuż całej rośliny.
- Wyróżniamy dwa rodzaje tkanki przewodzącej: ksylem (drewno, naczynia) oraz floem (łyko, rurki sitowe).
- Ksylem, inaczej naczynia lub tracheje, przewodzą wodę i sole mineralne z korzeni do liści. U roślin starszych ewolucyjnie: paprotników i nagozalążkowych, zamiast naczyń występują prymitywniejsze cewki (tracheidy). Ściany cewek mają jamki proste i lejkowate, kształt silnie wydłużony i ostro zakończone końce. Komórki naczyń są martwe, pozbawione protoplastu, tworzą zespoły stykających się osiowo i bocznie rurek biegnących nieprzerwanie od korzeni aż po liście. W ścianach rurek drzewnych
Rys. Ksylem - tkanka przewodząca
Rys. Cewki - tkanka przewodząca
w różnym stopniu zanikają ściany poprzeczne, a w ich obrębie powstają perforacje (jamki) ułatwiające przepływ wody. Naczynia są zgrubiałe i inkrustowane ligniną, dzięki czemu, jako sztywne i wytrzymałe, stanowią szkielet rośliny. Naczynia stanowią podstawowy element wiązek przewodzących pierwotnych i wtórnych.
- Floem, inaczej łyko - jego podstawowym elementem strukturalnym są żywe wydłużone komórki ułożone w zwarte zespoły, biegnące wzdłuż organów rośliny; ich ściany poprzeczne są perforowane i tworzą tzw. płytki sitowe. Komórki są silnie zwakuolizowane, połączone pasmami cytoplazmy przechodzącymi przez liczne otworki w ścianach, jądro rurek sitowych zanika. Rurkom sitowym towarzyszą komórki przyrurkowe zawierające protoplast i jądro komórkowe (pełnią funkcje odżywcze). U roślin starszych ewolucyjnie: paprotników i nagonasiennych, występują komórki sitowe, które są wydłużone, wrzecionowate i na całej powierzchni mają skupiska jamek prostych, tzw. pola sitowe. Floem przewodzi asymilaty z liści i zielonych części łodyg w dół rośliny do korzeni. Wyjątkowo w przypadku roślin wieloletnich po okresie spoczynkowym (zima) łyko przewodzi z korzeni spichrzowych do młodych liści.
Rys. Tkanka przewodząca - floem
Łyko stanowi podstawowy element wiązek przewodzących pierwotnych i wtórnych.
- Oprócz cewek, naczyń, rurek sitowych i komórek sitowych w tkance przewodzącej występują: włókna drzewne i łykowe o budowie zbliżonej do włókien sklerenchymatycznych oraz towarzyszący wiązkom przewodzącym miękisz drzewny i łykowy. Ich rolą jest wzmacnianie i odżywianie elementów przewodzących.
- Ze względu na zdolność do rozrastania się wiązki przewodzące dzielimy na:
otwarte - występuje w nich kambium i dzięki temu mogą tworzyć elementy drewna i łyka wtórnego, a roślina przyrasta na grubość (nagozalążkowe i okrytozalążkowe dwuliścienne);
zamknięte - nie występuje w nich kambium i roślina nie przyrasta na grubość (okrytonasienne jednoliścienne, wyjątek stanowią juki, draceny, palmy).
- Ze względu na liczbę i strukturę elementów składowych dzielimy je na:
proste - zbudowane tylko z łyka lub tylko z drewna (w budowie pierwotnej korzeni).
Oddzielnie przebiegające wiązki łykowe i drzewne występują w korzeniu, tworząc układ na przemianległy promienisty.
złożone - drewno i łyko występują obok siebie w różnych układach: wiązki łykodrzewne kolateralne zamknięte, kolateralne otwarte, bikolateralne, hadrocentryczne i leptocentryczne.
- Kolateralna zamknięta - drewno naprzeciw łyka, bez miazgi, np. jednoliścienne.
- Kolateralna otwarta - drewno naprzeciw łyka, między nimi warstwa miazgi, np. dwuliścienne.
- Bikolateralna (dwuobokleżne) - jedno pasmo drewna otoczone z dwóch stron łykiem zewnętrznym i wewnętrznym, występuje miazga, np. dynia, ziemniak.
- Hadrocentryczne (współśrodkowe) - drewno otoczone wokół łykiem, np. paprocie.
- Leptocentryczne (współśrodkowe) - łyko otoczone wokół drewnem, np. konwalia.
Rys. Rodzaje wiązek przewodzących: A - kolateralna zamknięta,
B - kolateralna otwarta, C - bikolateralna, D, E - koncentryczna:
D - kambium
21. Budowa wtórna łodygi (przekrój poprzeczny pnia drzew)
- Budowa wtórna jest wynikiem działalności dwóch merystemów bocznych: kambium i fellogenu; merystemy te wytwarzają nowe tkanki przewodzące: drewno i łyko wtórne oraz tkankę okrywającą - korek.
- Działalność merystemów wtórnych doprowadza do powstania grubego silnego pnia zdolnego do udźwignięcia rozgałęzionej korony. W łodygach zielnych przyrost wtórny jest nieznaczny lub nie występuje w ogóle. U współcześnie żyjących paprotników przyrost wtórny spotyka się zupełnie wyjątkowo, choć występował u form kopalnych. U jednoliściennych występuje tylko w nielicznych przypadkach. Powszechnie przyrost wtórny występuje u dużych wieloletnich form drzewiastych i krzewiastych okrytozalążkowych, dwuliściennych i nagozalążkowych.
- Kambium wiązkowe i międzywiązkowe łączą się ze sobą i tworzą ciągły cylinder tkanki merystematycznej.
Rys. Schemat rozwoju budowy pierwotnej i budowy wtórnej łodygi roślin
22. Na czym polega opadanie liści i jakie jest znaczenie tego zjawiska.
Niezwykłym zjawiskiem jest sposób organizacji anatomicznej która doprowadza do odpadnięcia liścia. Najczęściej składa się on z blaszki liściowej i ogonka. Ogonek który odznacza się większą sztywnością ze względu na duży udział tkanek wzmacniających (przede wszystkim kolenchymy) ma za zadanie przewodniczyć między liściem a pędem i tworzyć swoiste rusztowanie dla organu. Tylko kiedy liść zaczyna obumierać, albo następuje w roślinie stres wodny- liście muszą odpaść. Dzieje się to na skutek aktywności enzymów hydrolitycznych w stosunku do podstawowego budulca roślin (celulozy, pektyn i ligniny) i wytworzenia się formy : warstwy odcinającej. Jest to pasmo cienkościennych komórek, często, zewnętrznie z zaznaczonym wklęśnięciem. Na ten element czynności życiowych hormony roślinne także wpływają, podobnie, w grupach jak to jest w procesie destrukcji chlorofilu. Schematycznie warstwę odcinającą przedstawić można w taki sposób:
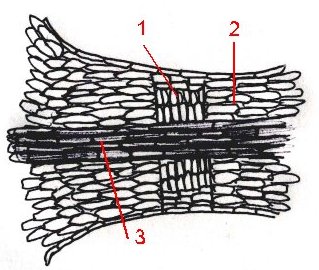
Warstwa odcinająca (1)Miękisz korowy (2)
Wiązki przwodzące(3)
Warstwa ta jest miejscem słabym jeśli chodzi o wytrzymałość na siły fizyczne, dlatego często przy podmuchach wiatru- liście, w jesiennych kolorach od gałęzi odczepiają się (następuje zabliźnienie) i zlatują dając przy tym obraz osobliwy dla tej pory roku co zradza natchnienie u poetów, muzyków i malarzy.
Na koniec zapytać trzeba- dlaczego tak się dzieje? Pytanie dlaczego ma dwa wymiary (zajmuje się tym filozofia) : dlaczego w sensie z jakiej przyczyny i dlaczego czyli po co (dla czego)?
Liście są organami niezwykle aktywnymi, konieczne w tym względzie otwarcie na świat zewnętrzny (cienka skórka, aparaty szparkowe, przewaga tkanek żywych) przypłacają krótkim życiem (mówię o roślinach liściastych z naszej strefy klimatycznej). Drzewo nie może przyjąć innej taktyki jak odrzucać organ który byłby narażony na destrukcję, a wiosną tworzyć nowy. Spodziewać się można, że odbywa się to przy znacznych nakładach energetycznych i ogólnej mobilizacji układu.
Zatem jesień to dla liści zaprogramowana genetycznie i przy udziale gospodarki hormonalnej śmierć. Śmierć mająca swój początek w destrukcji barwników fotosyntetycznych (chlorofili) i ujawniająca istnienie innych (karotenoidów) przez zmiany molekularne błon plazmatycznych i rozpad wewnątrzkomórkowych struktur, po hydrolizę celulozy i pektyn by ostatecznie ulec rozkładowi przy pracy saprofitów i już na ziemi. Dla niektórych ludzi, prawdziwe piękno jesieni właśnie w tym może się ujawnić- świadomości fizjologicznych procesów jednych zawęża do poziomu wyspecjalizowanego analizatora, innych rozwija i wzbogaca.
Opadanie liści
W trakcie przebarwiania, w miejscu styku ogonka z pędem powstaje tkanka odcinająca. Przy tym procesie niemały udział ma niezwykły, gdyż jedyny gazowy hormon roślin - etylen. Jego synteza zachodzi m. in. w starzejących się tkankach jesiennych liści. Działa on antagonistycznie do innego hormonu produkowanego w liściach - auksyny. W miarę starzenia się liści auksyn ubywa, czego rezultatem jest zwiększona produkcja etylenu w strefie odcięcia. Powoduje to kolejne fizjologiczne zmiany. Do dziś proces reakcji wywoływanych przez etylen nie jest do końca poznany.
Strefa odcięcia składa się z cienkościennych komórek miękiszowych i różni się tym samym od otaczających komórek. Od strony łodygi tworzą ją komórki korka. Po rozpuszczeniu przez enzymy blaszki środkowej liść utrzymywany jest na jednej wiązce przewodzącej. Staje się podatny na siły grawitacji i po czasie odpada. U niektórych drzew. np. u dębu i buka wiązka przewodząca jest wzmocniona, a przez to liść jest silniej związany z gałęzią i może pomimo obumarcia pozostać jeszcze na niej długi czas.
Drzewa iglaste
Inaczej procesy te przebiegają u drzew iglastych. Zredukowane liście - igły - pokryte są grubą warstwą skórki i substancji woskowej zabezpieczając je przed niskimi temperaturami, woda zaś jest z nich na okres zimy odprowadzana, by podczas mrozów nie doprowadzić do zamarznięcia i rozsadzenia igły. Dlatego drzewa iglaste, z wyjątkiem modrzewia, nie zrzucają igieł na okres zimy.
23. Rozmnażanie płciowe (rola, zalety)
Rozmnażanie płciowe, generatywne - rozmnażanie za pomocą haploidalnych komórek rozrodczych (gamet) - męskiej i żeńskiej, powstających zazwyczaj w wyspecjalizowanych narządach płciowych (u zwierząt) lub organach generatywnych (u roślin). Po połączeniu się dwóch komórek rozrodczych powstaje diploidalna zygota, z której rozwija się diploidalny zarodek, a następnie nowy organizm.Występują trzy sposoby rozmnażania płciowego: izogamia, anizogamia, oogamia, przy czym u roślin wyższych występuje tylko oogamia. Nietypowym sposobem rozmnażania jest tzw. koniugacja, która u roślin polega na kopulowaniu dwóch komórek stykających się ze sobą a pochodzących od dwóch organizmów poprzez przepłynięcie protoplastu męskiego do żeńskiego i zlanie się w zygotę.
Podstawową zaletą wynikającą z tego sposobu rozmnażania jest różnorodność potomstwa pozwalająca organizmom rozmnażającym się płciowo na szybszą ewolucję niż u rozmnażających się bezpłciowo. Ta różnorodność jest wynikiem kilku zdarzeń losowych, jakie mają miejsce w procesie tworzenia gamet i zapłodnienia:
segregacji chromosomów
losowego doboru gamet
Segregacja chromosomów - tworzące się gamety otrzymują po jednym chromosomie z każdej pary chromosomów homologicznych (a więc podobnych, lecz nie identycznych). Jeśli komórka macierzysta gamet ma tylko jedną parę chromosomów (sytuacja wielce nietypowa, liczba chromosomów zależy od gatunku organizmu), to podczas tworzenia się gamety następuje losowy wybór jednej z dwóch możliwości - gameta odziedziczy chromosom, który jej rodzic u swego zarania otrzymał w gamecie męskiej albo ten który rodzic otrzymał w gamecie żeńskiej.
Wymiana genów pomiędzy chromosomami homologicznymi znana jest lepiej jako zjawisko crossing-over. Dzięki temu mechanizmowi wymiany chromosomy nie są sztywnymi pakietami genów, ale geny które u rodzica należą do różnych chromosomów tej samej pary, mogą u potomka znaleźć się w tym samym chromosomie. Gameta otrzymuje tylko jeden chromosom każdej pary na skutek czego geny położone na różnych chromosomach ulegną rozdzieleniu, natomiast należące do tego samego chromosomu mają szansę odziedziczyć się razem.
Losowy dobór gamet - czynniki o których wspomniano w powyższych punktach prowadzą do dużej różnorodności gamet. Na ogół gamety tworzone są w dużym, a nawet gigantycznym nadmiarze i tylko nieliczne z nich uczestniczą w tworzeniu zygoty i rozwijającego się z niej organizmu potomnego. Gamety, którym dane będzie uczestniczyć w zapłodnieniu są wybierane losowo. Gamety nie wybrane giną bez śladu
Rozmnażanie płciowe zapewnia szybkie rozprzestrzenianie się zmienności genetycznej ,
Podczas rozmnażania płciowego dochodzi do mieszania się materiału genetycznego od ojca i matki, podczas gdy w trakcie rozmnażania bezpłciowego organizm potomny ma taki sam materiał genetyczny, jak organizm matczyny. Mieszanie się materiału genetycznego rodziców powoduje większą różnorodność genetyczną i dlatego stwarza lepszą szansę na powstanie potomstwa lepiej przystosowanego do środowiska. Z kolei rozmnażanie bezpłciowe zapobiega rozprzestrzenianiu się niekorzystnych mutacji - twierdzą Wiliam Rice i Adam K. Chippindale z University of California w Santa Barbara. Według nich, rozmnażanie płciowe pozwala też na pozbycie się niekorzystnego materiału genetycznego, swoistego "balastu".
24. Co to są gamety i zygota? Na czym polega zapylenie zapłodnienie?
Zygota — komórka powstała w wyniku zapłodnienia, czyli połączenia haploidalnej gamety męskiej z haploidalną gametą żeńską w procesie rozmnażania płciowego. Nowo powstała diploidalna komórka ulega dalszym podziałom dając początek organizmowi potomnemu. U protistów jednokomórkowych dwie łączące się komórki nazywają się gametami, a powstająca komórka diploidalna-komórką zygotyczną lub zygotą. Zygota jest tu krótkotrwałą formą życiową i szybko dzieli się mejotycznie, dając cztery haploidalne komórki potomne. Te zaś dojrzewają, stają się normalnymi organizmami zdolnymi do samodzielnego życia i rozmnażania bezpłciowego. Taki cykl życiowy określono jako cykl z mejozą postgamiczną, ponieważ mejoza zachodzi w nim bezpośrednio po gamii.
Gameta - komórka płciowa organizmów żywych.Może uczestniczyć w procesie zapłodnienia łącząc się z gametą innej płci i tworząc zygotę.U większości organizmów wyróżnia się dwa typy komórek płciowych, nazywane + i -, gdy morfologicznie się nie różnią (izogamia) lub męską i żeńską, gdy jedna z nich (plemnik, powstaje w gonadach męskich - jądrach) jest ruchliwa lecz uboga w materiały zapasowe, a druga (komórka jajowa, powstaje w gonadach żeńskich - jajnikach) - przeciwnie (oogamia). Wyróżnia się też sytuację pośrednią, gdy oba rodzaje komórek są ruchliwe, lecz komórki jednego typu są większe - makrogamety - od innych - mikrogamet (anizogamia).Gamety jako jedyny rodzaj komórek przechodzą podział mejotyczny. Powstają w procesie gametogenezy (plemniki podczas spermatogenezy, komórki jajowe - oogenezy).
Zapylenie, proces obejmujący przeniesienie pyłku z pręcika bezpośrednio do okienka zalążka (u roślin nagozalążkowych) lub na znamię słupka (u roślin okrytozalążkowych) oraz rozwój pyłku.
Zapylenie pyłkiem pochodzącym z innej rośliny nazywa się zapyleniem krzyżowym (obcopylność), zapylenie pyłkiem tej samej rośliny - samozapyleniem. Rośliny wytwarzają szereg cech utrudniających samozapylenie (które jest zjawiskiem niekorzystnym), a ułatwiających zapylenie krzyżowe, jak np.: różny czas dojrzewania pręcików i słupków (przedprątność i przedsłupność), różnosłupkowość oraz samopłonność.
Pyłek może być przenoszony przez wiatr (wiatropylność), zwierzęta (zoogamia), przede wszystkim owady (owadopylność), ale także np. przez ptaki (ornitogamia) oraz wodę (wodopylność, czyli hydrogamia).
Zapylenie - zjawisko występujące u roślin kwiatowych, polegające na przeniesieniu ziarna pyłku na znamię słupka. Kwiaty posiadają specjalne przystosowania do tego celu. Znamiona ich słupków są kleiste, a pyłek jest zwykle bardzo niewielkich rozmiarów.
Rodzaje zapylenia
Zapłodnienie - połączenie się jąder komórek rozrodczych (kariogamia komórki rozrodczej męskiej i żeńskiej) w wyniku czego powstaje nowa komórka nazywana zygotą.U ssaków, w tym człowieka, zapłodnienie może być skutkiem kopulacji pomiędzy osobnikami przeciwnych płci. Warunkiem zapłodnienia jest wniknięcie plemnika do komórki jajowej - ma to miejsce w bańce jajowodu, aczkolwiek do zapłodnienia może dojść w innym odcinku układu rozrodczego kobiety. Zapłodnienie może też być wynikiem inseminacji lub metody in vitro. W wyniku zapłodnienia dochodzi do przekazania potomstwu zawartych w gametach czynników dziedziczenia (genów) obydwu organizmów rodzicielskich. Wynikiem zapłodnienia jest ciąża. U ludzi do zapłodnienia dochodzi zwykle pomiędzy 10 a 18 dniem cyklu.
Zapłodnienie, połączenie się gamety żeńskiej i męskiej (czyli komórek rozrodczych) w jedną komórkę, tzw. zygotę, dającą początek nowemu organizmowi. Jeżeli gamety mają haploidalną liczbę chromosomów, w jądrze powstałej zygoty znajdują się dwa niezależne zespoły chromosomowe, jest więc diploidalna (wyjątkiem są np. gamety roślin poliploidalnych, dające w wyniku zapłodnienia poliploidalną zygotę).
Gamety pochodzą zwykle z różnych osobników (tzw. zapłodnienie krzyżowe - allogamia), rzadziej z tego samego osobnika (samozapłodnienie). Dla roślin okrytozalążkowych charakterystycznym zjawiskiem jest zapłodnienie podwójne.
25. Podziały mitotyczne i mejotyczne komórki (ich rola)
Mitoza - proces podziału pośredniego jądra komórkowego, któremu towarzyszy precyzyjne rozdzielenie chromosomów do dwóch komórek potomnych. W jego wyniku powstają komórki, które dysponują materiałem genetycznie identycznym z komórką macierzystą. Jest to najważniejsza z różnic między mitozą a mejozą. Mitoza zachodzi w komórkach somatycznych zwierząt oraz w komórkach somatycznych i generatywnych roślin.
Przebieg mitozy
Interfaza nie jest częścią mitozy. Stanowi część cyklu komórkowego pomiędzy podziałami komórki.[1] Interfaza - najdłuższa faza życia komórki, należąca do cyklu komórkowego. Jest etapem, w którym komórka przygotowuje się do podziału mitotycznego lub mejotycznego. Interfazę stanowią trzy stadia:
Faza G1 (ang. gap1 - przerwa) - poprzedza ją zakończony podział mitotyczny i jest fazą wzrostową komórki. Następuje synteza różnych rodzajów białek, m.in. strukturalnych czy enzymatycznych i zwiększenie organelli takich jak mitochondria, czy lizosomy. Komórka w tej fazie zwiększa swoją masę i objętość, osiągając stadium komórki macierzystej. Pod koniec fazy G1 dochodzi do syntezy specjalistycznych białek regulatorowych, odpowiedzialnych za przejście komórki w fazę S.
Faza S (ang. synthesis - synteza) - dochodzi do replikacji DNA, czyli do podwojenia ilości kwasu (z 2c do 4c, gdzie c oznacza ilość DNA). Poza tym zachodzi synteza histonów, a pod koniec fazy replikacja centriol. Proces ten u człowieka zachodzi zazwyczaj w ciągu 8 godzin.
Faza G2 (ang. gap2 - przerwa)- następuje synteza białek wrzeciona podziałowego, głównie tubuliny jak również składników błony komórkowej potrzebnych do jej wytworzenia po zakończonym podziale. Pod koniec fazy G2 dochodzi do syntezy specjalistycznych białek regulatorowych, odpowiedzialnych za przejście komórki w mitozę
Faza G0 (ang. gap0 - przerwa)- w przypadku, gdy nie dojdzie do wytworzenia białek odpowiedzialnych za przejście faz G1 i G2 do następnego stadium, komórka przechodzi w fazę G0. Interfaza ulega wtedy zatrzymaniu, komórka traci zdolność replikacji DNA i zaczyna się specjalizować. Dotyczy to np. komórek nerwowych czy mięśniowych. W niektórych przypadkach może dojść do powrotu do cyklu komórkowego poprzez stymulację komórek np. hormonami.
Profaza Jest to pierwszy etap podziału komórki eukariotycznej.
b) chromosomy zaczynają być widoczne
c) ujawnia się struktura chromosomu
g) zanika otoczka jądrowa
Metafaza
{kind=link}
{kind=link}
a) rozpad błony jądrowej (w tym momencie rozpoczyna się metafaza)
b) następuje przyczepienie wrzeciona podziałowego do centromerów
c) chromosomy ustawiają się w płaszczyźnie równikowej komórki, tworząc płytkę metafazową.
Anafaza
a) następuje rozdzielenie chromatyd siostrzanych, powstają chromosomy potomne (jest to właściwym początkiem anafazy)
c) podział organelli na równe zespoły
Telofaza
b) wyodrębniają się jądra potomne identyczne z jądrem rodzicielskim
c) chromosomy ulegają despiralizacji do chromatyny
d) dochodzi do cytokinezy (czasami proces ten dokonuje się już w anafazie)
e) powstają dwie diploidalne komórki potomne
Właściwy podział mitotyczny poprzedza przygotowująca do niego interfaza, które razem tworzą cykl komórkowy.
Mejoza, skrót: R! (R - od redukcji) - proces podziału jądra komórkowego. Polega na podziale jądra komórkowego, z którego powstają 4 jądra o połowie chromosomów (po jednym z każdej pary) komórki macierzystej. Podziałowi mejotycznemu ulegają komórki generatywne zwierząt oraz niektóre komórki somatyczne roślin (komórki macierzyste zarodników). W przypadku królestwa protista wyróżnia się 2 rodzaje mejozy: mejozę pregamiczną (poprzedzającą powstanie gamet) oraz mejozę postgamiczną (następującą po powstaniu gamet). Pierwszy podział mejotyczny nazywany jest podziałem redukcyjnym (mejoza I), drugi zaś podziałem zachowawczym, czyli ekwacyjnym (mejoza II). Podczas mejozy zachodzą dwa sprzężone ze sobą podziały:
I podział mejotyczny
II podział mejotyczny (przypominający podział mitotyczny)
Przebieg mejozy I
- Profaza I
wykształcenie się włókienka podziałowego (kariokinetycznego), kondensacja chromatyny do chromosomów jest długa i składa się z 5 stadiów:
Na tym etapie kończy się mejoza u ssaków niepłodnych, np. u muła ze względu na brak chromosomów homologicznych (jest on krzyżówką międzygatunkową)
diploten - pary chromatyd chromosomów siostrzanych rozchodzą się, ale pozostają złączone w punktach zwanych chiazmami. Rozdzielenie chromosomów homologicznych tzw. desynapsis następuje w wyniku rozpuszczenia kompleksu synaptonemalnego. Następuje intensywna synteza RNA i dekondensacja chromosomów. Może zachodzić crossing-over, czyli wymiana odcinków chromatyd chromosomów homologicznych
Zmniejszenie syntezy RNA, kondensacja chromosomów (grubieją i oddalają się od otoczki jądrowej). Kinetochory każdego z dwóch chromosomów tworzących biwalent zlewają się ze sobą. Mikrotubule łączą kinetochor tylko z jednym centromerem. Chromatydy niesiostrzane pozostają połączone w chiazmach, których liczba systematycznie maleje.
Biwalenty ustawione w płaszczyźnie równikowej (gwiazda macierzysta), mikrotubule wrzeciona kariokinetycznego połączone z nimi poprzez kinetochory. Wrzeciono gotowe.
Włókna wrzeciona skracają się i odciągają chromosomy do biegunów komórki - następuje redukcja liczby chromosomów.
Odtwarzanie się otoczek jądrowych. Chromosomy częściowo ulegają despiralizacji, następuje cytokineza i powstają dwie komórki potomne, które mają o połowę mniej chromosomów niż komórka macierzysta.
Przebieg mejozy II
Profaza II
Formowanie nowego wrzeciona podziałowego, zanika otoczka jądrowa.
Metafaza II
Kończy się tworzenie wrzeciona podziałowego. Centromery chromosomów ustawiają się w płaszczyźnie równikowej komórki. Nici białkowe wrzeciona łączą się z centromerami.
Anafaza II
Wrzeciono podziałowe kurczy się, centromery pękają, czego skutkiem jest oddzielenie się chromatyd
Telofaza II
Odtworzenie otoczki jądrowej wokoło skupisk chromosomów potomnych - wyodrębnienie się jąder potomnych, despiralizacja chromosomów do chromatyny.
Cytokineza
Podział cytoplazmy.
W rezultacie mejozy I tworzą się 2 komórki haploidalne(1n), a kolejny podział, już bez redukcji materiału genetycznego, sprawia, że w wyniku całej mejozy z jednej komórki diploidalnej powstają 4 komórki haploidalne.
26. Części składowe kwiatu i ich funkcje
Budowa morfologiczna kwiatu
Pojedynczy kwiat zbudowany jest z częsci płonnych nie związanych bezpośrednio z rozmnażaniem, oraz z części płodnych. Części płodne to pręciki z woreczkami pyłkowymi, są to organy męskie oraz owocolistki z zalążkami - organy żeńskie. Z części płonnych (sterylnych) kwiatu zbudowana jest okrywa kwiatowa, która u roślin nagonasiennych jest bardzo niepozorna a niejednokrotnie nie występuje wcale, u roślin okrytozalążkowych zaś jest różnorodna i czasami bardzo okazała.W przypadku roślin nagonasiennych nieosłonięte zalążki leżą na pojedynczych owocolistkach. Zaś u roślin okrytonasiennych jeden lub kilka zrośniętych owocolistków tworzy słupek, w jego dolnej częsci tworzącej zalążnię, zamknięte są zalążki.
Kwiat rozwija się z pączków kwiatowych jako pęd boczny w pachwinie przysadki (liścia przykwiatkowego, wspierającego). Umieszczony jest na szypułce lub jest siedzący.
Części składowe kwiatu:
dno kwiatowe - wszystkie elementy kwiatu są na nim osadzone.
miodniki - gruczoły występujące w kwiatach owadopylnych.
Słupek (łac. pistyllum, ang. pistil) - żeński organ płciowy w kwiecie okrytonasiennych. Zbudowany jest ze zrośniętych ze sobą lub wolnych owocolistków (carpellae), które są zmodyfikowanymi liśćmi. Słupki zajmują zawsze centralne miejsce w kwiecie. Mogą występować pojedynczo lub po kilka tworząc w tym drugim przypadku słupkowie (gynaeceum).
Słupek zbudowany jest z jednego (słupek apokarpiczny) lub kilku (słupek synkarpiczny) owocolistków (makrosporofilów). Owocolistki mogą zrastać się ze sobą brzegami lub zaginać się do środka słupka i zrastać się zewnętrznymi powierzchniami. W tym ostatnim przypadku owocolistki tworzą wewnątrz słupka tzw. przegrody prawdziwe, dzielące zalążnię słupka na komory. Czasem powstają niepełne przegrody, nie rozdzielające słupka na komory (np. w rodzaju mak Papaver). Niekiedy tworzą się fałszywe przegrody, powstające nie ze ścian owocolistków, ale z wybujałych tkanek owocolistków (np. w rodzinie kapustowatych podzielone są fałszywą przegrodą na dwie komory). W zależności od liczby owocolistków budujących słupek, wyróżnia się słupki jedno-, dwu-, trój- lub wielokrotne, które mogą mieć od jednej do wielu komór.
Morfologicznie słupek składa się z części dolnej - zalążni (ovarium), która na górze zwęża się w szyjkę słupka i zakończona jest znamieniem. Czasem (np. u tulipana) nie wyodrębnia się szyjka - mówimy wtedy o znamieniu siedzącym. W komorach zalążni słupka znajdują się zalążki wyrastające na łożyskach.
{kind=link}
Pręcikowie (gr. androeceum, andros oikia = męski dom) - wszystkie pręciki jednego kwiatu. Kwiaty okółkowe, zawierające jeden okółek pręcików nazywane są haplostemon. Często występują dwa okółki, które nazywane są diplostemon jeżeli zewnętrzny okółek jest międzyległy do wewnętrznego lub obdiplostemon jeśli jest mu nadległy (np. u gruboszowatych i goździkowatych). Pręciki zależnie od umieszczenia nadległego działkom korony nazywane są epipetala, a nadległe działkom kielicha episepala. Pręcikowie o ustawieniu spiralnym składa się zwykle ze zmiennej liczby pręcików.
Okwiat, okrywa kwiatowa (ang. perigon, perianth, łac. perigonium, perianthium) - część kwiatu złożona zwykle z kielicha (kalyx) i korony (corolla), może występować także kieliszek (epikalyx). Okwiat służy ochronie pręcików i słupków, a także jako powabnia u roślin owadopylnych dzięki zapachowi i kolorom, dla zapylających je owadów, ptaków, ssaków i innych zwierząt.
U jednoliściennych występuje okwiat pojedynczy (perigonium), który składa się z dwóch okółków jednakowych listków okwiatu (tepala) - barwnych jak u tulipana lub zielonych jak u kosmatki.
Korona (łac. corolla, ang. corolla) - element kwiatu składający się z okółka barwnych płatków korony (petala), które stanowią powabnię dla owadów, lub innych zwierząt zapylających kwiaty. Korona stanowi wewnętrzną część okwiatu.
Barwa płatków jest spowodowana występowaniem barwników w soku komórkowym (np. antocyjany). Olejki eteryczne, które nadają kwiatom zapach, występują zazwyczaj w komórkach epidermy płatków. Płatki korony składają się z rozszerzonej zwykle blaszki i wąskiego paznokcia, za pomocą którego są przytwierdzone do dna kwiatowego. Na płatkach korony u wielu roślin występują miodniki wytwarzające nektar. Czasami całe płatki przekształcone są w miodniki - tzw. listki miodnikowe.
Korony mogą być:
Kielich (łac., ang. kalyx) - najbardziej zewnętrzna część kwiatu, składająca się z okółka zielonych działek kielicha (łac. sepala, ang. sepals), które rozwinęły się głównie z normalnych liści podkwiatkowych. Zasadniczą funkcją kielicha jest osłanianie wewnętrznych części kwiatu. Zwykle jest zielony. U niektórych roślin kielich pełni dodatkowo funkcję powabni, mającej za zadanie przywabianie owadów. Zazwyczaj ma to miejsce w kwiatach o zredukowanych płatkach korony. Wówczas kielich pełni funkcję płatków korony i jest jaskrawo ubarwiony. Pod kielichem u niektórych roślin występuje kieliszek (epikalyx) np. u truskawki.
Rodzaje kielicha ze względu na jego trwałość:
kielich odpadający. Zasadnicza funkcja kielicha kończy się z chwilą rozwoju kwiatów i wówczas u wielu roślin kielich odpada
kielich trwały - pozostający po przekwitnięciu kwiatu i biorący udział w osłanianiu młodego owocu. Czasami podczas rozwoju owocu kielich znacznie rozrasta się (tzw. kielich powiększony). U niektórych roślin wiatrosiewnych przekształca się w puch kielichowy (pappus), tworzący aparat lotny umożliwiający długie unoszenie się nasion w powietrzu.
Dno kwiatowe (ang. receptacle, łac. receptaculum) - przeważnie mniej lub bardziej rozszerzony koniec szypułki lub łodygi, na którym wyrasta kwiat. W zależności od rośliny dno kwiatowe może być poszerzane, płaskie, wypukłe lub wklęsłe. Boczne części kwiatu są osadzone na nim.
Specyficzną odmianą dna kwiatowego jest tzw. hypancjum - jest silnie wgłębione i otacza słupki znajdujące się wewnątrz niego.
Miodniki, nektaria (łac. nectaria, ang. nectary) - organ gruczołowy roślin wydzielający nektar zwabiający owady. Rośliny posiadające miodniki i wytwarzające nektar nazywamy roślinami miododajnymi.Nektaria występują najczęściej w obrębie kwiatów (nektaria kwiatowe), czasami jednak na innych częściach rośliny; na ogonkach liściowych i głównym nerwie liścia (np. u akacji i śliwy), na liściach przykwiatowych, lub w kątach nerwów liści. Są to tzw. nektaria pozakwiatowe. W obrębie kwiatu miodniki występować mogą u podstawy słupka, na wewnętrznej stronie płatków korony czy u podstawy płatków korony. Czasami funkcję miodników pełnią niektóre przekształcone pręciki tzw. prątniczki. Tak jest np. u dziewięciornika błotnego.
27. Scharakteryzować budowę słupka i określić rolę poszczególnych elementów
Znamię (ang. stigma) - część słupka roślin okrytonasiennych przyjmująca ziarna pyłku w trakcie zapylenia (przed zapłodnieniem). Zazwyczaj znajduje się na szczycie szyjki słupka, gdy szyjki brak - wykształca się znamię siedzące. Liczba znamion w słupku jest różna u różnych gatunków i zwykle zależy od liczby owocolistków budujących słupek.Znamię zbudowane jest z tkanki zwanej transmisyjną (stygmatoidalną), która tworzy także połączenie z wnętrzem zalążni. W prymitywnych owocolistkach znamieniem jest powierzchnia brzegów owocolistka. W bardziej wyspecjalizowanych organach znamię może być specyficznie uformowane w zależności od budowy słupka i biologii zapylania danego gatunku. Rośliny wiatropylne mają znamię w postaci pędzelków (np. pokrzywa), piórek (np. trawy) lub są nitkowato wydłużone (np. leszczyna). Rośliny owadopylne mają znamiona pokryte brodawkami lub włoskami, wykształcone w postaci listka znamieniowego (np. szafran), o kształcie miseczkowatym, kulistym lub języczkowatym. Odpowiedni kształt znamienia i różnego rodzaju włoski, brodawki umożliwiają zatrzymanie i umieszczenie ziarn pyłku na znamieniu.Owłosione znamiona zwykle są suche, podczas gdy znamiona gładkie powleczone są lepką wydzieliną. Wydzielana jest ona przez epidermę gruczołowatą pokrywającą znamię, czasem dodatkowo opatrzoną brodawkami lub włoskami. U niektórych gatunków tkanka wydzielnicza znajduje się pod epidermą. W takim przypadku wydzieliny gromadzą się najpierw w przestworach międzykomórkowych, a następnie wydostają się na zewnątrz przez luźno ułożone komórki skórki i pęknięcia w obrębie kutykuli.Wydzieliny występują na znamieniu zwykle bardzo obficie, chroniąc znamię przed wyschnięciem, utrzymując warstewkę wody koniecznej do skiełkowania pyłku, nie służą natomiast do wabienia owadów. Wydzielina poza tym, że podobnie jak różnego rodzaju włoski, brodawki, ułatwia zatrzymywanie i utrzymywanie się pyłku na znamieniu, dodatkowo umożliwia selektywne kiełkowanie pyłku. Zawiera substancje hormonalne inicjujące jego kiełkowanie oraz wzrost łagiewki pyłkowej. Wydzielinę stanowi mieszanina lipidów, związków fenolowych, glikozydów, antocyjanów, enzymów i kwasów aminowych.
Pyłek kwiatowy, ziarna pyłku - męskie elementy rozrodcze występujące u roślin nasiennych wytwarzane w komorach pyłkowych pylników z tkanki archesporialnej. Pylniki są elementami pręcików. Nowo powstałe ziarno pyłku stanowi mikrosporę, a dojrzałe ziarno pyłku - gametofit męski. Służy ono do zapylania słupków kwiatowych.U roślin nagonasiennych ziarno pyłku posiada pęcherzyki lotne, które umożliwiają wiatropylność. U okrytonasiennych pyłek przenoszony jest zazwyczaj przez owady, ziarno pyłku uległo dalszej redukcji (brak pęcherzyków lotnych).Nauką zajmującą się badaniem pyłku jest palinologia. Aby określić skład pyłku wykonuje się analizę pyłkową.
Łagiewka pyłkowa - wytworzona przez komórkę wegetatywną znajdującą się w mikrosporze (ziarnku pyłku), przenosi komórki plemnikowe do woreczka zalążkowego.
wydłużona wypustka wegetatywnej komórki ziarna pyłku. Zadaniem łagiewki pyłkowej jest przetransportowanie komórek plemnikowych do rodni (u nagozalążkowych) lub woreczka zalążkowego (u okrytozalążkowych). Wykształcenie łagiewki pyłkowej umożliwiło roślinom nasiennym uniezależnienie procesu zapłodnienia od wody.
Łagiewka pyłkowa
SYNERGIDY
dwie komórki aparatu jajowego występujące w woreczku zalążkowym okrytonasiennych. Umieszczone są wraz z komórką jajową przy biegunie mikropylarnym. Synergidy odgrywają rolę w procesie zapłodnienia - przyjmują łagiewkę pyłkową, która wnika do woreczka zalążkowego właśnie poprzez te komórki. woreczek zalążkowy.
Komórka jajowa : Gameta żeńska(komórka jajowa) tworzy się w zalążni.
Woreczek zalążkowy - gametofit żeński u roślin okrytozalążkowych, rozwijający się z jednej z czterech makrospor (powstałych w wyniku mejozy w jednej z komórek ośrodka zalążka).Makrospora rozwija się i przekształca w jednojądrowy woreczek zalążkowy. Woreczek rozrasta się i wypełnia prawie całkowicie zalążek. Jądro woreczka trzykrotnie dzieli się mitotycznie i powstaje osiem haploidlanych jąder potomych. Dwa z nich z czasem łączą się w diploidalne jądro wtórne woreczka. Pozostałe sześć haploidalnych jąder otacza się własną cytoplazmą i tworzy osobne komórki. Z komórek tych powstają: komórka jajowa, dwie synergidy, trzy antypody.U niektórych gatunków liczba antypod w woreczku zalążkowym może sięgać kilkudziesięciu, w wyniku powtarzających się podziałów komórek.
Antypody - komórki wegetatywne woreczka zalążkowego, ustawione w woreczku zalążkowym od strony osadki (w dolnym, chalazalnym biegunie woreczka); najczęściej trzy, ale może być ich więcej (np. u niektórych gatunków z rodziny wiechlinowatych lub astrowatych) lub mogą nie występować wcale (w rodzinie wiesiołkowatych). Zwykle żyją krótko i uczestniczą w odżywianiu woreczka zalążkowego. Czasem mogą być długotrwałe (np. u tojadu), mają wtedy prawdopodobnie funkcję wydzielniczą.
28. Kwiaty obupłciowe, rozdzielnopłciowe i nijakie, rośliny jednopienne i dwupienne - znaczenie rozdzielności płci
Wyróżnia się kwiaty obupłciowe (zawierające jednocześnie słupki i pręciki).
KWIATY są przeważnie obupłciowe (zawierają jednocześnie pręciki i owocolistki) i wykazują ogromną różnorodność budowy. Owocolistki zrastają się i w ten sposób tworzą charakterystyczny dla okrytonasiennych - SŁUPEK. Typowy kwiat oprócz pręcików i owocolistków posiada PŁATKI KORONY i DZIAŁKI KIELICHA. Wszystkie te elementy osadzone są w postaci okółków na osi (silnie skróconej) zwanej DNEM KWIATOWYM.
Obupłciowość, androginia, u roślin występowanie męskich i żeńskich organów rozrodczych (gonady) w tym samym kwiecie lub kwiatów męskich i żeńskich na tej samej roślinie. Obupłciowość występuje u większości roślin wyższych.
Rozdzielnopłciowość, występowanie w obrębie jednego gatunku bądź populacji osobników męskich i żeńskich. Rozdzielnopłciowość jest zjawiskiem powszechnym u zwierząt wyższych.
Wśród roślin wyższych dość częste jest występowanie na jednym osobniku rozdzielnopłciowych kwiatów, tzn. męskich, zawierających tylko pręciki i żeńskich, zawierających tylko słupki. Rzadziej zdarzają się gatunki o rozdzielnopłciowych osobnikach, produkujących tylko męskie lub tylko żeńskie kwiaty.
Rozdzielnopłciowość częstsza jest u roślin nagonasiennych (np. miłorząb, welwiczia, sagowce), bardzo rzadka wśród roślin okrytonasiennych (np. wierzby, topole, pokrzywy, szczawie, palmy). Rozdzielnopłciowe kwiaty występują u roślin z rodziny szpilkowych.
Występują także kwiaty nijakie (bez pręcików i słupków), które służą za powabnie (kwiaty brzeżne u chabra bławatka.
Jednopienne rośliny, rośliny, które jedocześnie posiadają odrębne kwiaty jednopłciowe: słupkowe (żeńskie) i pręcikowe (męskie). Jednopienność (monoecja) występuje np.: u brzozy, buka, leszczyny, sosny.
Dwupienne rośliny, rośliny, których część osobników wytwarza wyłącznie kwiaty żeńskie, a inne wyłącznie kwiaty męskie. Dwupienność występuje u niektórych drzew (topole, wierzby) i nielicznych roślin zielnych (szczaw, chmiel, pokrzywa). rośliny mające na jednych osobnikach tylko kwiaty żeńskie (słupkowe), na drugich - wyłącznie kwiaty męskie (pręcikowe); dwupienność uniemożliwia samozapylenie;
29. Rola bielma (endospermu). Co to są rośliny bezbielmowe
Bielmo (endosperm) - di- lub tri- lub pentaploidalna (2n, 3n, 5n zależnie od rodziny, lub w wyniku endomitoz w dojrzałym bielmie - wielokrotność tych wartości) tkanka miękiszowa w nasionach roślin okrytonasiennych powstająca w trakcie procesu podwójnego zapłodnienia po połączeniu jednego z jąder plemnikowych z wtórnym jądrem woreczka zalążkowego (typowa sytuacja - wtedy bielmo jest triploidalne). Cytoplazmę pierwszej komórki bielma stanowi pozostałość woreczka zalążkowego po utworzeniu i oddzieleniu się błonami komórki jajowej, synergid i antypod. U roślin jednoliściennych bielmo jest głównym źródłem substancji pokarmowych w dojrzałym nasieniu. U roślin dwuliściennych bielmo zostaje zużyte przez rozwijający się zarodek, który następnie gromadzi substancje pokarmowe w liścieniach.
Funkcja bielma
Bielmo jest tkanką spichrzową, gromadzącą materiały zapasowe dla rozwijającego się zarodka. Substancje te powstają w częściach asymilacyjnych rośliny, następnie doprowadzane są do tkanek w sąsiedztwie woreczka zalążkowego. U wielu roślin, w celu wykorzystania tych substancji, powstają w bielmie ssawki (haustoria), które mogą przerastać różne części zalążka (ośrodek, chalazę, osłonki) a także ścianę zalążni i stąd czerpać materiały zapasowe. Bielmo może magazynować:
Rośliny bezbielmowe to np. rośliny strączkowe.
30. Obcopylność i samopylność
Obcopylność, allogamia, zapylenie krzyżowe, zapylenie znamienia słupka kwiatowego pyłkiem obcym, czyli pochodzącym od innej rośliny tego samego gatunku. Obcopylność prowadzi do wzrostu zróżnicowania genetycznego populacji i zwiększenia jej potencjału adaptacyjnego.
zdolność niektórych roślin do zapłodnienia i wytwarzania nasion tylko po zapyleniu pyłkiem pochodzącym z innego kwiatu, zwykle z innej rośliny
Samopylność , samozapylenie (autogamia) - jest to zapylenie słupka pyłkiem pochodzącym z pręcików z tego samego kwiatu, lub innych kwiatów, ale na tej samej roślinie. Dla większości roślin jest to zjawisko niekorzystne gdyż nie następuje wymiana materiału genetycznego przy powstawaniu osobników potomnych. Większość roślin wykształciła więc różne mechanizmy zapobiegające samozapyleniu: przedprątność, przedsłupność, dwupienność, różnosłupkowość , samopłonność.U wielu roślin mechanizmy zabezpieczające przed samozapyleniem nie są całkowicie skuteczne. U roślin tych zawsze część nasion powstaje w wyniku zapylenia pyłkiem pochodzącym z innej rośliny (zapylenie krzyżowe), a część w wyniku samozapylenia. Tak jest np. u berberysu Thunberga, rutewki orlikolistnej i wielu innych. Ilość nasion powstałych w wyniku samozapylenia zależy od różnych czynników, duży wpływ ma pogoda. Przy dobrej pogodzie, gdy kwiaty są licznie odwiedzane przez owady, większość nasion powstaje w wyniku zapylenia krzyżowego, w czasie długotrwałych deszczów większa jest szansa na samozapylenie. U złoci żółtej w czasie deszczu kwiaty zamykają się i wówczas dochodzi w nich do samozapylenia.
31. Dlaczego samopylność jest z reguły niekorzystna dla roślin?
Dla większości roślin jest to zjawisko niekorzystne gdyż nie następuje wymiana materiału genetycznego przy powstawaniu osobników potomnych. Większość roślin wykształciła więc różne mechanizmy zapobiegające samozapyleniu: przedprątność, przedsłupność, dwupienność, różnosłupkowość , samopłonność. W wyniku samozapłodnienia u takich roślin obserwuje się homozygotyczność wszystkich roślin w populacji. Homozygotyczność rośliny wyjściowej, czyli dającej początek populacji powoduje późniejszy brak jej (populacji) zróżnicowania pod względem genetycznym. Natomiast sama homozygotyczność jest efektem powtarzającego się przez kilka kolejnych pokoleń samozapłodnienia, które coraz bardziej eliminuje z populacji heterozygoty. Wszystkie rośliny potomne, mające ten sam genotyp, zwane są linią czystą.
Wynika z tego fakt , że pod względem genetycznym korzystniejszym dla roślin jest obcopylność , ponieważ efektem krzyżowania może być także zjawisko heterozji, czyli tzw. bujności mieszańców.
32. Bariery zapobiegające samozapyleniu
W przyrodzie rośliny z reguły unikają zapylenia własnym pyłkiem. Istnieją różnego rodzaju zabezpieczenia, uniemożliwiające ten proces.
SAMOPŁONNOŚĆ: ziarna pyłku nie kiełkują na znamionach słupków tej samej rośliny. Jest to cecha uwarunkowana genetycznie.
BARIERA CZASOWA: nierównoczesne dojrzewanie słupków i pręcików w tym samym kwiecie:
Protogynia :(przedsłupność) - wcześniejsze dojrzewanie słupków;
Protandria :(przedprątność) - wcześniejsze dojrzewanie pylników i wysypywanie się pyłku.
BARIERA PRZESTRZENNA:
Monoecja (jednopienność) - na tej samej roślinie występują kwiaty rozdzielnopłciowe;
Dioecja (dwupienność) - na jednych osobnikach powstają tylko kwiaty męskie, na innych tylko żeńskie;
Heterostylia (różnosłupkowość) - w obrębie gatunku występują kwiaty różniące się długością szyjki słupka. Słupki są dłuższe lub krótsze od pręcików, co uniemożliwia samozapylenie.
33. Związek budowy kwiatu ze sposobami jego zapylenia (owadopylność, wiatropylność)
Owadopylność (entomogamia, entomofilia) - jeden z zoogamicznych sposobów zapylania oznaczający zapylanie kwiatów przez owady przenoszące pyłek. Owadopylność jest zależnością mutualistyczną, powstała na drodze koewolucji.
Kwiaty roślin wykazują szereg przystosowań zwabiających owady i umożliwiających zapylenie:
barwne, duże kwiaty lub drobne kwiaty zebrane w duże kwiatostany,
wykształcenie pyłku o dużych, lepkich ziarnach, bogatych w substancje odżywcze.
Kolorowe plamy i lśniące włoski wskazują owadom drogę do ukrytych w głębi kwiatu miodników. Jeżeli kwiaty są małe, zebrane są w duży, widoczny z daleka kwiatostan. Tak jest np. u słonecznika i u innych roślin z rodziny astrowatych i innych rodzin. Kwiaty niektórych roślin np. berberysu, mają specjalną budowę, dzięki której pod ciężarem owada pręcik przegina się, obsypując go pyłkiem. Niektóre kwiaty stanowią dla owadów pułapkę. Np. u obuwika pospolitego kwiaty zamykają się, uniemożliwiając owadowi wyjście przez długi czas, co daje pewność zapylenia kwiatu przez wiercącego się w środku owada.
Wiatropylność, anemogamia, rozsiewanie i przenoszenie pyłku przez wiatr. U roślin nagozalążkowych pyłek przenoszony jest bezpośrednio na zalążek, u okrytozalążkowych - na znamiona słupków. Rośliny wiatropylne produkują dużo lekkiego i suchego pyłku.U wielu roślin wiatropylnych, w tym u większości drzew, kwiaty rozwijają się wcześniej niż liście (proanthia), co zwiększa szanse na zapylenie.
Kwiaty roślin wiatropylnych mają prostą budowę, są bezwonne i nie posiadają miodników, nie muszą bowiem zwabiać owadów. Są też z reguły drobne i niepozorne. Wytwarzają za to bardzo dużo lekkiego pyłku, który łatwo wysypuje się z licznych pylników. W jednym kłosie żyta jest ich aż 42 mln[potrzebne źródło]. U niektórych roślin z rodziny pokrzywowatych pyłek jest mechanicznie wyrzucany z pylników w czasie ich gwałtownego rozprostowywania się. Słupki w kwiatach roślin wiatropylnych mają duże znamiona, często podzielone lub piórkowate, by łatwiej mogły wychwycić pyłek. Często pokryte są lepką substancją ułatwiającą przyklejenie się ziaren pyłku.Aby doszło do zapylenia krzyżowego niezbędne jest, by pyłek mógł jak najdłużej utrzymywać się w powietrzu i był odporny na niekorzystne warunki atmosferyczne. Przystosowaniem do utrzymywania się w powietrzu pyłku sosny są dwie komory powietrzne obecne przy każdym ziarnie pyłku.
34. Środowisko a roślina, formy życiowe: hydrofiy, higrofity, kserofity( sklerofity, sukulenty), epifity
Hydrofity, rośliny wodne, rośliny wodnopączkowe (gr. hydrol = woda, fyton = roślina) - rośliny wodne (oprócz planktonu), których pączki odnawiające zimują w wodzie. Wg klasyfikacji Raunkiæra są jedną z grup kryptofitów, czyli roślin skrytopączkowych. W strefach klimatycznych, w których występują okresy niesprzyjające wegetacji roślin (np. zima), zimują pączki znajdujące się w wodzie. Na wiosnę odtwarza się z nich nowy pęd.
Wg klasyfikacji Raunkiaera, zmodyfikowanej przez Braun-Blanqueta, wśród hydrofitów wyróżnia się 3 grupy:
korzeniące się w podłożu - np. Hydrophyta radicantia. Dzielą się na dwie podgrupy:
Hygrofity, higrofity, rośliny bagienne - rośliny zamieszkujące siedliska lądowo-bagienne, mocno uwilgotnione. Są roślinami wilgociolubnymi, wrażliwymi na niedobór wody. Należą do nich np. takie gatunki, jak szczawik błotny, knieć błotna. Posiadają duże liście pokryte cienką kutykulą i aparatami szparkowymi po obu stronach blaszki. Występują u nich duże przestwory międzykomórkowe wypełnione aerenchymą. System korzeniowy jest słaby, ciśnienie osmotyczne wynosi 8-11 atmosfer. Szybko więdną po zerwaniu (paprocie)organizmy roślinne, wymagające w swoim środowisku dużej wilgoci., wilgociolubne. Charakteryzują się m.in. dużymi liśćmi o cienkich, miękkich blaszkach (niekiedy szpiczasto zakończonych dla ułatwienia spływu wody), oraz płasko rozłożonym systemie korzeniowym, wynikającym z wysokiego stanu wód gruntowych (np. namorzyny). Przeciwieństwem higrofitów są kserofity (rośliny sucholubne).
Kserofity, rośliny sucholubne, kserofilne - rośliny przystosowane (fizjologicznie i anatomicznie) do życia w suchych miejscach takich jak step, pustynia, półpustynia, wydmy, piaski. Niektóre z nich potrafią magazynować wodę, np. kaktusy. Przy niewielkim udziale wody mogą budować swoje ciała. Dzieli się je na:
kserofity (gr. kserós `suchy' + phytón `roślina') bot. rośliny występujące w b. suchym środowisku, w warunkach niskiej wilgotności, posiadające cechy przystosowawcze do takich warunków; rośliny sucholubne, suchorośla.
Sukulenty, rośliny gruboszowate - grupa kserofitów, czyli roślin, które przystosowały się do życia w warunkach bardzo gorących dzięki możliwości magazynowania wody w różnych tkankach. Rośliny gruboszowate odznaczają się mocno rozwiniętą tkanką magazynującą wodę w śluzowatym soku komórkowym, redukcją liści, małą transpiracją, ciśnieniem osmotycznym (2-3 atmosfer) oraz wolną produkcją materii organicznej. Przystosowane są do bezpośredniego pobierania wody deszczowej.
Ze względu na miejsce magazynowania wody dzieli się sukulenty na 3 grupy:
Cechy anatomiczne przystosowujące sukulenty do środowisk, w których żyją, to:
zmniejszone przestrzenie międzykomórkowe,
zamieranie zakończeń korzeni w czasie suszy.
Sklerofity, rośliny należące do grupy kserofitów, występujące w środowiskach suchych, przystosowane do okresów niedoboru wody przez silne ograniczenie parowania, natomiast nie magazynujące jej.
Sklerofity mają wieloletnie, drobne, twarde, często błyszczące liście o budowie łuskowato-igiełkowatej, z silnie rozbudowaną tkanką mechaniczną, pokryte kutykulą lub gęstymi włoskami, zaopatrzone w liczne, lecz głęboko osadzone, aparaty szparkowe. Częste jest występowanie cierni pochodzenia liściowego lub pędowego (pęd roślinny). Niekiedy blaszka liściowa ulega redukcji, a jej funkcje przejmuje spłaszczony ogonek liściowy (liściaki) lub liściokształtna łodyga (gałęziaki).
W okresie dostatku wody sklerofity silnie transpirują, dzięki dobrze rozwiniętemu systemowi korzeniowemu i dużej liczbie szparek.
Epifit (porośle, aerofit) - roślina rosnąca na innej roślinie, ale nie prowadząca pasożytniczego trybu życia. Korzysta z innego gatunku jako podpory, a odżywia się samodzielnie. Wyrasta w miejscach, gdzie gromadzi się martwa materia organiczna (np. w kącie gałęzi/pędu) lub osiedla się na pędach innych roślin, a składniki odżywcze i wodę pobiera z powietrza lub opadów.
Epifity nie pozostają zupełnie obojętne dla roślin służących im za podłoże. Zatykają bowiem ich przetchlinki utrudniając wymianę gazów. Przyczyniają się także do butwienia kory, poprzez akumulację cząstek mineralnych i organicznych w swych darniach. Tworzą też tym samym warunki do rozwoju pasożytów: grzybów i owadów.
W naszych warunkach klimatycznych epifitami są głównie rośliny zarodnikowe: mszaki, porosty, glony. Rzadziej (jako semiepifity) na innych roślinach wyrastają gatunki zielne, lub nawet krzewy i drzewa. Ma to miejsce w sytuacjach szczególnie sprzyjających kolonizacji np. w przypadku starych, próchniejących wierzb, zwłaszcza głowiastych. Między ich konarami gromadzi się gruba warstwa próchnicy umożliwiająca rozwój np. bzu czarnego lub jarzębinie.
35. Klasyfikacja roślin Raunkiera
System Raunkiaera jest to powszechnie stosowany system klasyfikacji form życiowych roślin, stworzony przez duńskiego botanika C. Raunkiaera w 1905 roku. Jest to przykład klasyfikacji niesystematycznej. Raunkiaer dzieli rośliny według sposobu, w jaki przeżywają niekorzystną porę roku, szczególną rolę przypisując rozmieszczeniu i ochronie pąków spoczynkowych. Wyróżniono w oparciu o te kryteria takie typy form życiowych:
Kryptofity (skrytopączkowe) - są to rośliny mające pąki odnawiające osadzone na ukrytych w glebie, błocie lub wodzie organach spichrzowych, np. bulwach, kłączach, rozłogach i cebulach. Rośliny wykształcające podziemne kłącza, rozłogi, bulwy lub cebule (geofity) mogą zakwitać bardzo wcześnie, przed wytworzeniem pełnego ocienienia przez drzewa. Rośliny wodne, które są całkowicie zanurzone w wodzie, reprezentują grupę hydrofitów. Rośliny wieloletnie wyrastające ponad wodę zalicza się do helofitów.
Klasyfikacja ta jest przydatna do określenia z jakiej szerokości geograficznej pochodzi dana roślina oraz wskazuje na warunki bytowanie owych roślin.
36. Genotyp a fenotyp (ekspresja genu)
Genotyp - zespół genów danego osobnika warunkujących jego właściwości dziedziczne. Jest to sparowany układ alleli (często myli się go z genomem, czyli składem genetycznym danego organizmu). Można go wyrazić symbolicznie za pomocą oznaczeń aa, AA lub Aa, gdzie aa i AA oznaczają homozygotę pod względem tego genu, a Aa oznacza heterozygotę.
Przykład: świnki morskie o genotypach BB i Bb są podobne fenotypowo, to znaczy obie mają czarną sierść. Po skrzyżowaniu świnki czarnej BB ze świnką brązową (homozygotą bb) otrzymuje się osobniki wyłącznie czarne (heterozygoty Bb). Jednakże, po skrzyżowaniu heterozygotycznej świnki czarnej Bb z homozygotą recesywną bb otrzymuje się osobniki dwóch kolorów: czarne (Bb) i brązowe (bb).
Metoda krzyżowania z homozygotą recesywną jest najprostszą metodą badania genotypu. Jest to tak zwane krzyżowanie testowe, stosowane od dawna w hodowli zwierząt i roślin użytkowych.
Fenotyp (gr. phainomai - przejawiać; typos - wzór, norma) to najogólniej mówiąc zespół cech organizmu, włączając w to nie tylko morfologię, lecz również np. właściwości fizjologiczne, płodność, zachowanie się, ekologię, cykl życiowy, zmiany biologiczne, wpływ środowiska na organizm. Fenotyp jest ściśle związany z genotypem, bowiem to właśnie oddziaływanie między genotypem a środowiskiem daje fenotyp. Dlatego ten sam genotyp może dać różne fenotypy w różnych środowiskach (tzw. plastyczność fenotypowa), lub odwrotnie - mimo odmiennych genotypów uzyskać ten sam fenotyp.
Gdy mamy do czynienia z organizmem dobrze poznanym pod względem genetycznym, można stwierdzić jakie zmiany na poziomie genomu objawiają się w fenotypie. Zapoznanie się z genomem populacji naturalnej jest niezwykle trudne, dlatego zazwyczaj przyjmuje się po prostu, że fenotyp jest obrazem genomu w środowisku i pyta się, jaka część zmienności obserwowanej w naturze ma podłoże genetyczne .
Ekspresja genu (ang. gene expression) - proces, w którym informacja genetyczna zawarta w genie zostaje odczytana i przepisana na jego produkty, które są białkami lub różnymi formami RNA.
Przebieg tego procesu różni się nieco pomiędzy bakteriami i eukariotami. U bakterii geny są zwykle zorganizowane w grupy genów zwane operonami (np. operon laktozowy), które są regulowane jako grupa i przepisywane na zawierający kilka genów mRNA.
U eukariotów regulacja oraz przepisywanie na mRNA odnosi się do pojedynczego genu. Proces ten zachodzi w kilku etapach (Eukariota, w uproszczeniu, dla genu kodującego białko):
degradacja mRNA
przemieszczenie białka do właściwej pozycji (np. błony komórkowej, mitochondrium, etc.) (może poprzedzać poprzedni proces)
degradacja białka
Ekspresja RNA jest w etapach podobna do białkowej. Nie występuje translacja. Nie dla każdego rodzaju nieinformatycznego-RNA obejmuje wszystkie etapy. Występuje inicjalizacja, transport, splicing, modyfikacje, samoporządkowanie cząsteczki, funkcjonowanie i degradacja.
Regulacja ekspresji genu to złożony wieloczynnikowy proces. Na każdym z etapów ekspresja genu może być regulowana za pomocą różnych mechanizmów. Ekspresja genu zależy od rodzaju komórki, fazy rozwoju organizmu, metabolicznego/fizjologicznego stanu komórki.
37. Z czego powstaje owoc. Jego budowa i rola.
Owoc (łac. fructus) - w znaczeniu botanicznym występujący u okrytozalążkowych organ powstający z zalążni słupka i ewentualnie dna kwiatowego zawierający w swym wnętrzu nasiona, osłaniający je i ułatwiający rozsiewanie.
Kluczowym składnikiem owoców ze względu na ich funkcję są nasiona, będące właściwymi organami rozmnażania i rozprzestrzeniania roślin. Budowa owoców służy usprawnieniu różnych metod rozsiewania. Owoce mięsiste roznoszone są przez zwierzęta, dla których stanowią pokarm, owoce suche rozprzestrzeniane są przez wiatr, wodę lub zwierzęta.
Gdy w jednym kwiecie występuje większa liczba słupków wolnych, wtedy powstają z nich samodzielne owocki, które tworzą owoc zbiorowy na wspólnym dnie kwiatowym (malina, truskawka). Są one często mylone z owocostanami, które powstają z kilku kwiatów zebranych w skupiony kwiatostan (morwa, kłębek buraka, figowiec, ananas).
W znaczeniu kulinarnym owoc to jadalna część rośliny będąca owocem w znaczeniu botanicznym ale rosnąca tylko na drzewach lub krzewach. Owoce roślin zielnych uważane są za warzywa (np. pomidor, ogórek, groch, dynia). Wyjątkowo wśród roślin zielnych owoce w znaczeniu kulinarnym dają truskawki, poziomki i ananas.
W terminologii rolniczo-sadowniczej jakość owoców określa się za pomocą kształtu owocu i urody owocu. Terminy te są wyznacznikiem wartości owocu na podstawie oceny wizualnej.
38. Związek między budową owocu i nasienia a sposobem rozprzestrzeniania się rośliny
Owoc składa się z owocni i nasion. Nasiona powstają z zalążków, a owocnie z zalążni słupka (w tzw. owocach właściwych), czasem w ich wykształceniu uczestniczą też inne części kwiatu, np. dno kwiatowe, okwiat, liście przykwiatowe lub oś kwiatostanu (powstają wtedy tzw. Owoc rzekomy, owoc szupinkowy, w którego wykształceniu, oprócz zalążni, uczestniczą inne składniki kwiatu, tworzące tzw. szupinkę. Przykładem owocu rzekomego jest jabłko, którego soczysta część powstaje z dna kwiatowego, a owocolistki tworzą łuski otaczające nasiona.
Owocnia (perykarp) składa się z trzech warstw: zewnętrzna jednowarstwowa skórka (egzokarp), często zabarwiona, pełni rolę powabni dla zwierząt (w owocach mięsistych), lub (w owocach suchych) zaopatrzona jest w urządzenia czepne (szczecinki, haczyki), lotne (skrzydlate wyrostki, włoski) albo śluzowacieje, co ułatwia przyczepianie się owoców do ciał zwierząt lub rozsiewanie ich przez wiatr. Warstwa środkowa (mezokarp) może być różnej grubości, mięsista lub sucha. Warstwa wewnętrzna (endokarp) jest na ogół jednowarstwowa, może być błoniasta i cienka (jak np. w strąkach grochu) lub gruba i zdrewniała, tworząca pestkę (np. u śliwy).
39. Budowa nasienia
Nasiono, nasienie (semen, diaspora) - zarodek otoczony tkanką zapasową i osłonięty łupinami, który powstaje z zapłodnionego zalążka roślin nasiennych. Funkcją nasiona jest przede wszystkim ochrona zarodka przed niesprzyjającymi warunkami zewnętrznymi.
Nasiona roślin okrytozalążkowych powstają z zalążka, po zapłodnieniu komórki jajowej przez komórkę plemnikową. Zawierają trzy najważniejsze składniki: zarodek, łupina nasienna (testa) i tkanka odżywcza w postaci bielma lub obielma.
U roślin nagozalążkowych materiał zapasowy nasiona stanowi bielmo pierwotne. Nasiona u okrytozalążkowych mogą występować jako obielmowe, bielmowe i bezbielmowe.
2)Budowa nasienia
Nasienie otoczone jest upiną nasienna zawiera zarodek oraz materiały zapasowe np. skrobię w nasionach zbóż albo tłuszcze w nasionach słonecznika lub rzepaku. Materiały te zn. się w specjalnej tkance - bielnie lub liścieniach. Rośliny okrytonasienne w zależności od liczby liścieni w nasieniu dzielimy na jedno i dwu liścienne.
Typowe nasienie składa się z :
zarodka
bielma
łupiny nasiennej
Nasiona okrytozalążkowych powstają w zalążni w wyniku rozwoju zalążków, co następuje po zapyleniu i podwójnym zapłodnieniu
40. Rozmnażanie generatywne i jego rola
Rozmnażanie płciowe, generatywne - rozmnażanie za pomocą haploidalnych komórek rozrodczych (gamet) - męskiej i żeńskiej, powstających zazwyczaj w wyspecjalizowanych narządach płciowych (u zwierząt) lub organach generatywnych (u roślin). Po połączeniu się dwóch komórek rozrodczych powstaje diploidalna zygota, z której rozwija się diploidalny zarodek, a następnie nowy organizm.Występują trzy sposoby rozmnażania płciowego: izogamia, anizogamia, oogamia, przy czym u roślin wyższych występuje tylko oogamia. Nietypowym sposobem rozmnażania jest tzw. koniugacja, która u roślin polega na kopulowaniu dwóch komórek stykających się ze sobą a pochodzących od dwóch organizmów poprzez przepłynięcie protoplastu męskiego do żeńskiego i zlanie się w zygotę.Koniugację, obserwuje się również u bakterii, ale polega ona jedynie na przekazywaniu sobie przez bakterie fragmentów materiału genetycznego. Nie towarzyszy temu podział komórki, nie jest to więc forma rozmnażania. O zachowaniu bakterii w koniugacji decyduje obecność w nich tzw. czynnika płci (czynnika F).
41. Rozmnażanie wegetatywne (sposoby) i jego rola
Rozmnażanie bezpłciowe, wegetatywne, agamiczne - typ rozmnażania organizmów, który nie jest związany z wytwarzaniem komórek generatywnych czyli gamet. Organizm potomny powstaje z części organizmu rodzicielskiego. U bakterii może dojść do prostego przewężenia i powstania dwóch komórek potomnych z jednej rodzicielskiej, organizm rozgwiazdy może zostać odtworzony tylko z jednego ramienia oderwanego od organizmu rodzicielskiego, rośliny mogą wytwarzać bulwy, cebule, rozłogi, porosty urwistki itd. (patrz typy rozmnażania bezpłciowego poniżej)
W rozmnażaniu bezpłciowym organizm potomny otrzymuje zestaw genów identyczny z organizmem rodzicielskim. Ewentualne zmiany genotypu odbywają się zazwyczaj na drodze mutacji. Organizmy rozmnażające się bezpłciowo o genotypach mało podatnych na mutacje są gorzej przystosowane do zmieniających się warunków środowiska (w związku z brakiem rekombinacji). Ten typ rozmnażania umożliwia im jednak szybkie zasiedlenie wolnej niszy ekologicznej pojawiającej się efemerycznie. Dobrym przykładem jest tu mszyca, która w okresie wegetacji roślin rozmnaża się bezpłciowo (partenogenetycznie) a jesienią płciowo.
Rodzaje rozmnażania bezpłciowego
42. Rośliny samożywne, pasożyty, półpasożyty, owadożerne
Samożywność, autotrofizm, autotrofia (gr. autos-sam, trofe-pożywienie, pokarm) - sposób odżywiania się charakterystyczny dla roślin, części bakterii i protistów, polegający na syntezie związków organicznych z prostych związków mineralnych. Energia konieczna do tego procesu pochodzi z energii słonecznej (fotosynteza) lub z prostych związków organicznych, nieorganicznych lub pierwiastków np. z metanu, siarkowodoru, jonów żelaza (chemosynteza). W związku z tym wyróżnia się organizmy fotoautotroficzne i chemoautotroficzne.
Pasożyt - organizm cudzożywny, który wykorzystuje stale lub okresowo organizm żywiciela jako źródło pożywienia i środowisko życia. Pasożytnictwo to oddziaływanie antagonistyczne między organizmami, która przynosi korzyść jedynie pasożytowi; żywicielowi, zwanemu czasem gospodarzem, związek ten przynosi wyłącznie szkody (straty substancji odżywczych, destrukcja tkanek, zatrucie produktami przemiany materii pasożyta itp.). Pasożyt może doprowadzić organizm żywiciela do wyniszczenia a nawet śmierci.Ze względu na stopień zależności od żywiciela pasożyty dzieli się na:
fakultatywne, okolicznościowe, względne - mogące rozwijać się zarówno na żywych organizmach, jak również na materii nieożywionej. Forma odżywiania zależy od predyspozycji genetycznych, dostępu do rośliny żywicielskiej, jej podatności na zakażenie oraz warunków pogody
obligatoryjne, ścisłe, bezwzględne i względne - mogące rozwijać się w fazie wegetatywnej tylko w lub na organizmach żywych. Poza organizmem żywiciela pasożyty te mogą przetrwać tylko jako niektóre formy rozwojowe organizmu - przetrwalniki.
Ze względu na środowisko życia pasożyty dzieli się na:
PÓŁPASOŻYT
organizmy roślinne, które wytwarzają w procesie fotosyntezy substancje odżywcze, a od żywiciela pobierają wodę i sole mineralne, np. jemioła. Do pobierania tych związków z naczyń gospodarza jemioła wytwarza korzenie ssawki, tzw. haustoria.
roślina czerpiąca z innych roślin wodę i sole mineralne, a jednocześnie zdolna do asymilacji dwutlenku węgla
Roślina mięsożerna (roślina owadożerna) - roślina zdolna do chwytania i pobierania z ciał ofiar potrzebnych jej składników. Ofiarami są drobne zwierzęta, głównie owady i skorupiaki.Są to rośliny przeprowadzające fotosyntezę, ale rosnące na glebach ubogich w azot i inne pierwiastki potrzebne do prawidłowego rozwoju, to głównie te związki zdobywają poprzez trawienie ciał zwierząt.Do chwytania i trawienia służą im specjalnie przekształcone liście. Mogą one przybrać postać między innymi dzbanków, pułapek zatrzaskowych, lepów.
43. Jesienne przebarwianie liści
Warstwa odcinająca przerywa też odpływ cukrów, które jeszcze w liściach powstają, choć w coraz mniejszych ilościach. Z tych cukrów niektóre gatunki drzew produkują nowe pigmenty - antocyjany. To one powodują, że żółknące korony drzewa stają się czerwone. Te ciekawe barwniki zmieniają kolor od fioletu do intensywnej purpury, w zależności od pH soku komórkowego, w którym są rozpuszczone. Poza liśćmi występują także w kwiatach i owocach (na przykład winogronach i jabłkach). Do ich powstania potrzebne jest światło. Dlatego jabłka są rumiane tylko z jednej - słonecznej strony, a ich zacieniona część pozostaje zielona.
Tyle na temat jesiennych kolorów mówi fizjologia roślin.
44. Przyrost łodygi na grubość u roślin drzewiastych
Przyrost wtórny, przyrost łodygi i korzenia na grubość dzięki odkładaniu drewna wtórnego (ku środkowi) i łyka wtórnego (ku obwodowi) przez miazgę twórczą (kambium) oraz przyrost wtórnej tkanki okrywającej (korek) dzięki działalności miazgi korkotwórczej (fellogen).
W wyniku przyrostu wtórnego powstają jednolite pierścienie drewna i łyka. Przyrost wtórny występuje u większości roślin dwuliściennych, przy czym u roślin zielnych jest ograniczony do jednego sezonu wegetacyjnego (okres wegetacyjny), a u roślin drzewiastych trwa przez wiele okresów wegetacyjnych, prowadząc do znacznego przyrostu grubości pni, gałęzi i korzeni.
Na podstawie przyrostu wtórnego drewna można określać wiek drzew. U roślin jednoliściennych przyrost wtórny występuje tylko w nielicznych przypadkach (np. u draceny, juki), a mechanizm jego jest inny.
45. Przyrost łodygi na grubość u roślin jednoliściennych
Jak wiadomo rośliny jednoliścienne nie przyrastaja na grubość gdyż nie posiadaja kambium ani fellogenu. Są jednak wyjątki takie jak draceny, juki, i palmy.
W jaki sposób udaje im się zwiększać średnice łodygi
Wiązki przewodzące u roślin jednoliściennych są zamknięte i nie układają się w regularny cylinder (są rozrzucone w miękiszu). Jeśli kambium pojawia się (tak jak U rodzajów Dracaena i Yucca) to w postaci cylindra będącego kontynuacją merystemu podwierzchołkowego łodygi. Po wewnętrznej stronie (do środka) odkładane są komórki które różnicują się w kolejne zamknięte wiązki kolateralne. Na zewnątrz odkłądane są jedynie komórki miękiszowe, w niewielkiej ilości.
. Jednoliścienne rośliny nie posiadają możliwości przyrostu na grubość. W ten sposób w tej grupie są zazwyczaj rośliny roczne lub dwuletnie. Są to trawy i zioła.
Charakterystyka roślin jednoliściennych: Kiełkujący zarodek posiada tylko jeden liścień. Wiązki przewodzące rozrzucone nieregularnie na całym przekroju łodygi mają charakter wiązek zamkniętych, nie mają więc możliwości wytwarzania drewna wtórnego. Brak jest przyrostu na grubość (przyrost wtórny), typowego dla roślin dwuliściennych. Pnie (kłodziny) drzewiastych przedstawicieli jednoliściennych, zbudowane są ze zbitej masy wiązek przewodzących. Liście przeważnie równowąskie, bez wyraźnego ogonka liściowego, nerwacja równoległa. Kwiaty zbudowane są z 3-elementowych okółków, okwiat z dwu okółków po 3 działki, 3-6 pręcików, słupek 3-komorowy z trzema znamionami.
Do jednoliściennych należą takie rodziny jak: trawy, turzyce, palmy, liliowate, storczykowate
46. Czynniki wpływające na roślinę
Czynniki środowiskowe wpływające na rozwój roślin nasiennych
- Do czynników ekologicznych - środowiskowych zaliczamy: temperaturę, wilgotność, światło oraz zaopatrzenie w substancje odżywcze - CO2 i sole mineralne.
- Każdy z tych czynników wpływa w określony sposób na dany gatunek rośliny, to znaczy działa w innym zakresie wartości, wyznaczonym przez wartość minimalną i maksymalną. To zjawisko nazywamy zakresem tolerancji organizmu na dany czynnik.
- Czynniki środowiskowe działają jednocześnie i kompleksowo, ale stopień wzrostu i rozwoju każdego organizmu jest określony przez czynnik występujący w najmniejszej ilości (prawo minimum Liebiega).
- Światło ma na roślinę ogromny wpływ pośredni lub bezpośredni. Pośredni wpływ na roślinę ma światło inicjujące fotosyntezę, dla której jest źródłem energii. Światło pośrednio reguluje także intensywność transpiracji, stopień otwierania aparatów szparkowych.
Bezpośrednio światło wpływa na kiełkowanie nasion wrażliwych na światło, indukcję kwitnienia, otwieranie i zamykanie się kwiatów, różnicowanie się płci u roślin rozdzielnopłciowych, wytwarzanie bulw, rozłogów i cebul, wydłużanie międzywęźli u traw. U roślin wykazano obecność barwnika fitochromu, który absorbując fale świetlne o określonej długości, stymuluje wzrost i rozwój rośliny. Są to reakcje fotomorfogenetyczne.
- Temperatura wpływa na ogół reakcji biochemicznych, ponieważ decyduje o aktywności enzymów. Optymalna temperatura dla większości roślin naszego klimatu to 20-30 0 C.
- Tlen jest niezbędnym substratem oddychania komórkowego, natomiast dwutlenek węgla substratem fotosyntezy. Roślina musi mieć nieprzerwany dostęp do tych gazów.
- Znaczenie wody i soli mineralnych jest nie do przecenienia i zostało omówione w temacie dotyczącym biologicznego znaczenia wody.
Czynniki wewnętrzne wpływające na rozwój roślin nasiennych
Fitohormony są specyficznymi związkami organicznymi wytwarzanymi w tkankach roślinnych (najczęściej merystematycznych) w bardzo niskich stężeniach, regulującymi (stymulującymi lub hamującymi) przebieg pewnych procesów fizjologicznych. Nie mają one żadnego znaczenia odżywczego dla rośliny, są jedynie sygnałami chemicznymi wysyłanymi do mniej lub bardziej oddalonych części rośliny, w których ujawnia się ich fizjologiczna aktywność (od tej reguły są wyjątki). W przeciwieństwie do hormonów zwierzęcych charakteryzują się małą specyficznością działania, gdyż uczestniczą w regulacji wielu procesów wzrostowych i rozwojowych. Oznacza to, że jeden rodzaj hormonu roślinnego wywołuje różnorodne efekty. Związki hormonalne powodujące przyspieszenie procesów wzrostowych nazywamy stymulatorami lub aktywatorami, a hamujące te procesy - inhibitorami wzrostu.
Wyszukiwarka
Podobne podstrony:
Zagadnienia na kolokwium opracowanie
opracowanie zagadnień na egzamin, opracowanie pytań egzaminacyjnych
Zagadnienia na filozofię i opracowanie, Filozofia
PG zagadnienia na kolokwium opracowanie, FiR UMK Toruń 2010-2013, III FIR, Prognozowanie gospodarcze
Politka zagadnienia na egz opracowany
Zagadnienia na Pająka!!! opracowane i poukladane po kolei
Zagadnienia na zagrożenia (opracowanie), Ochrona Środowiska studia, 3 rok (2008-2009), Semestr VI (R
zagadnienia na egzamin opracowane technologia roślinna
Opracowanie Zagadnień na egzamin Mikroprocki
Opracowane zagadnienia na koło z podstaw turystyki, Notatki na koła
Opracowanie zagadnień na prawo handlowe
opracowane zagadnienia na egazamin
Opracowanie zagadnień na egzamin z MO
Jasiorski, chemia ogólna, Opracowane zagadninia na kolowium
więcej podobnych podstron