spermatogeneza�4
się mostki cytoplazmatyczne (ryc. 1.24, 1.26), pozwalające na zsynchronizowanie; podziałów komórkowych, a także umożliwiające transport hormonów i innych niezbęi nych substancji przekazywanych z komórek podporowych (Sertolego).
Nie wszystkie spermatocyty, które przechodzą podziały mejotyczne, przekształcają! się ostatecznie w plemniki. Jak obliczono, w przypadku szczurów z pojedynczego spep matogonium macierzystego może powstać teoretycznie 4096 plemników. Jednakże" w praktyce liczba ta jest znacznie mniejsza i wynosi ok. 25-50%, w wyniku tzw. śmierci,; programowanej (apoptozy) komórek płciowych podczas spermatogenezy. Śmierć pro-gramowana jest procesem prawidłowym, stanowiącym m.in. rodzaj mechanizmu ściek cyjnego, który uniemożliwia kontynuowanie rozwoju komórkom nieprawidłowy^ (zwłaszcza z aberracjami chromosomowymi).
•■'•I
SPERMIOGENEZA
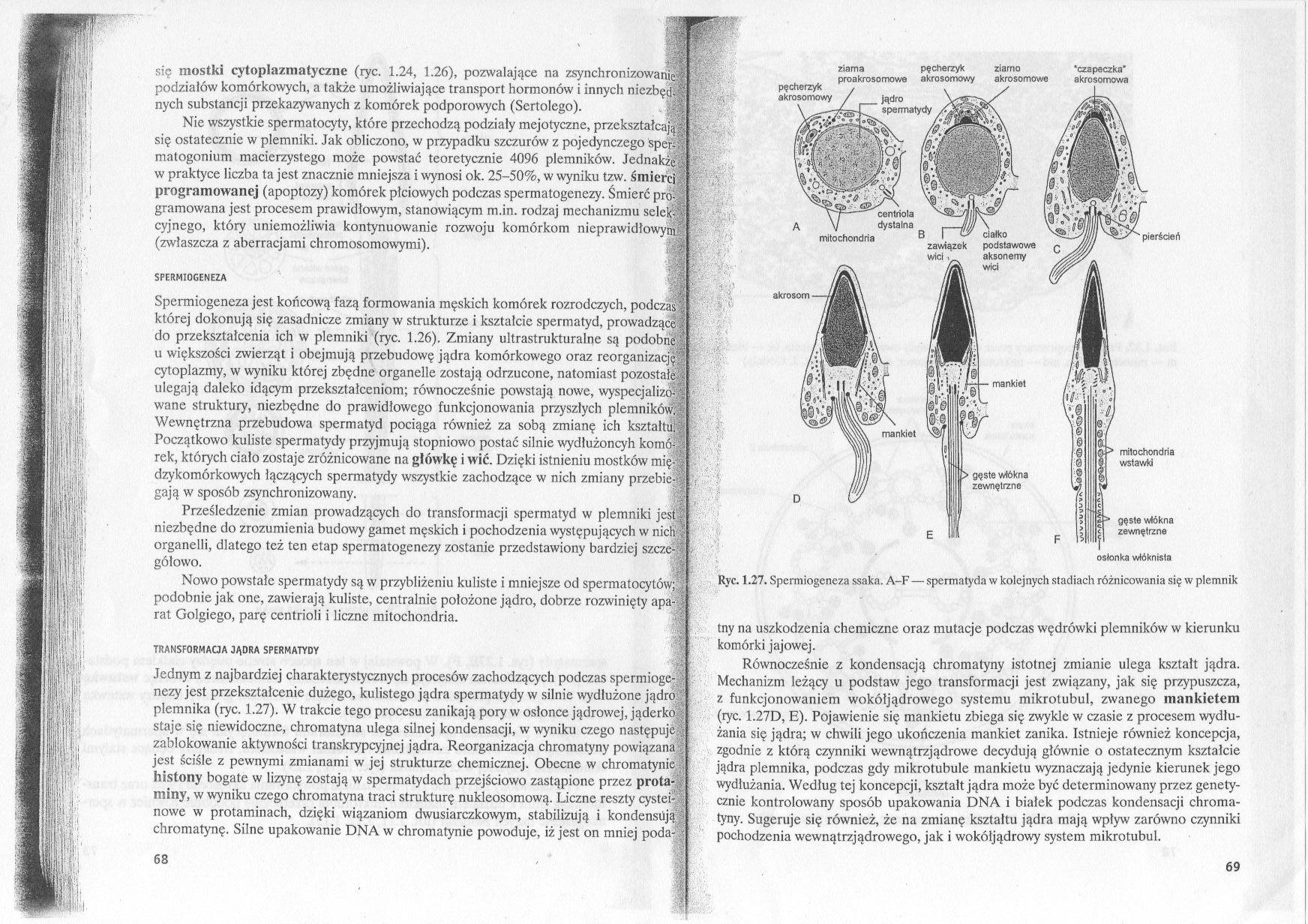
Spermiogeneza jest końcową fazą formowania męskich komórek rozrodczych, podczas której dokonują się zasadnicze zmiany w strukturze i kształcie spermatyd, prowadzące do przekształcenia ich w plemniki (ryc. 1.26). Zmiany ultrastrukturalne są podobne u większości zwierząt i obejmują przebudowę jądra komórkowego oraz reorganizację cytoplazmy, w wyniku której zbędne organelle zostają odrzucone, natomiast pozostałe; ulegają daleko idącym przekształceniom; równocześnie powstają nowe, wyspecjalizof wane struktury, niezbędne do prawidłowego funkcjonowania przyszłych plemników) Wewnętrzna przebudowa spermatyd pociąga również za sobą zmianę ich kształtu! Początkowo kuliste spermatydy przyjmują stopniowo postać silnie wydłużoncyh komo! rek, których ciało zostaje zróżnicowane na główkę i wić. Dzięki istnieniu mostków mię dzykomórkowych łączących spermatydy wszystkie zachodzące w nich zmiany przebić-' gają w sposób zsynchronizowany.
Prześledzenie zmian prowadzących do transformacji spermatyd w plemniki jest; niezbędne do zrozumienia budowy gamet męskich i pochodzenia występujących w nich . organelli, dlatego też ten etap spermatogenezy zostanie przedstawiony bardziej szcze-t gółowo.
Nowo powstałe spermatydy są w przybliżeniu kuliste i mniejsze od spermatocytów;; podobnie jak one, zawierają kuliste, centralnie położone jądro, dobrze rozwinięty aparS rat Golgiego, parę centrioli i liczne mitochondria.
■i*
TRANSFORMACJA JĄDRA SPERMATYDY
Jednym z najbardziej charakterystycznych procesów zachodzących podczas spermiogeę • nezy jest przekształcenie dużego, kulistego jądra spermatydy w silnie wydłużone jądro plemnika (ryc. 1.27). W trakcie tego procesu zanikają pory w osłonce jądrowej, jąderko staje się niewidoczne, chromatyna ulega silnej kondensacji, w wyniku czego następuje zablokowanie aktywności transkrypcyjnej jądra. Reorganizacja chromatyny powiązana ? jest ściśle z pewnymi zmianami w jej strukturze chemicznej. Obecne w ehromatynie . histony bogate w lizynę zostają w spermatydach przejściowo zastąpione przez prota- i miny, w wyniku czego chromatyna traci strukturę nuklcosomową. Liczne reszty cystei-nowe w protaminach, dzięki wiązaniom dwusiarczkowym, stabilizują i kondensiiją; chromatynę. Silne upakowanie DNA w ehromatynie powoduje, iż jest on mniej poda-,
pęcherzyk
akrosomowy
ziarna pęcherzyk ziarno
proakrosomowe akrosomowy akrosomowe
mitochondria ® \ ^ cia?°
zawiązek podstawowe wici aksonemy
wid
centriola dystalna
mankiet \&J]
• mankiet
gęste włókna zewnętrzne
pierścień
gęste włókna zewnętrzne
osłonka włóknista
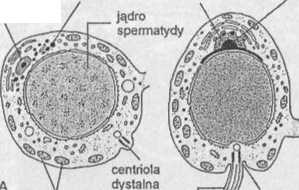
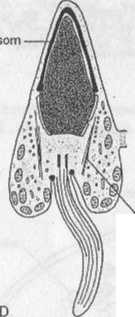
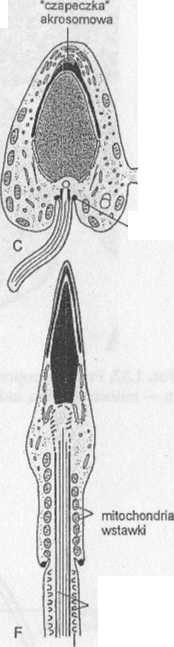
Ryc. 1.27. Spermiogeneza ssaka. A-F — spermatyda w kolejnych stadiach różnicowania się w plemnik
tny na uszkodzenia chemiczne oraz mutacje podczas wędrówki plemników w kierunku komórki jajowej.
Równocześnie z kondensacją chromatyny istotnej zmianie ulega kształt jądra. Mechanizm leżący u podstaw jego transformacji jest związany, jak się przypuszcza, z funkcjonowaniem wokóljądrowego systemu mikrotubul, zwanego mankietem (ryc. 1.27D, E). Pojawienie się mankietu zbiega się zwykle w czasie z procesem wydłużania się jądra; w chwili jego ukończenia mankiet zanika. Istnieje również koncepcja, zgodnie z którą czynniki wewnątrzjądrowe decydują głównie o ostatecznym kształcie jądra plemnika, podczas gdy mikrotubule mankietu wyznaczają jedynie kierunek jego wydłużania. Według tej koncepcji, kształt jądra może być determinowany przez genetycznie kontrolowany sposób upakowania DNA i białek podczas kondensacji chromatyny. Sugeruje się również, że na zmianę kształtu jądra mają wpływ zarówno czynniki pochodzenia wewnątrzjądrowego, jak i wokóljądrowy system mikrotubul.
69
Wyszukiwarka
Podobne podstrony:
spermatogeneza�4 Bi się mostki cytopłazmatyczne (ryc. 1.24, 1.26), pozwalające na zsynchronizowanie
Zdjęcie552 Dalszy rczu ój wirusologii wiązał się z rozwojem technik laboratoryjm ch i metod pozwalaj
0000068 (3) zaznacza się obecność struktur helikalnych. Metoda ORD pozwala na określenie, jaka liczb
62 wtóre. będziemy starali sie określić, czy wyeliminowanie części założeń pozwala na uznanie teorii
zaznacza się obecność struktur helikalnych. Metoda ORD pozwala na określenie, jaka liczba reszt
0000068 (3) zaznacza się obecność struktur helikalnych. Metoda ORD pozwala na określenie, jaka liczb
CCF20100503�001 12 Charakterystyka fizjograficzna Wielkopolski Ryc. 1. Kraina Wielkopolsko-Kujawska
0000014 3 UWAGA: Przy nieco zmienionych parametrach metoda wirowania pozwala na frakcjonowanie skład
Obraz 012 r rsdn* Z odpo^edzi mi* j*sf pf«wid
Warzywnictwo105 Ryc. 26. Roszponka Ryc. 24. Główki cykorii sałatowej tworzące się w trakcie pędzenia
img013 (60) a1L_ „ Ryc. 5*24. Schemat przebiegu endo- i egzocytozy; a-c adsorbcja materiału i wpukle
Hoglądy/ Zdarzenia W dniach 24 26 listopada 2006 roku odbyła się w Kazimierzu Dolnym IX Międzynarodo
41101 IMG)24 (3) 600 Podstawy chirurgii i Ryc. 20.26. Płat TRAM z dodatkowym unaczynieniem
44787 img013 (60) a1L_ „ Ryc. 5*24. Schemat przebiegu endo- i egzocytozy; a-c adsorbcja materiału i
Aby otrzymać rekonstrukcję objętościową (ryc. 24.21) do komputera wprowadza się wartości liczb CT -
img013 (60) a1L_ „ Ryc. 5*24. Schemat przebiegu endo- i egzocytozy; a-c adsorbcja materiału i wpukle
więcej podobnych podstron