Farm1967
podjednostcc a, natomiast dla pikrotoksyny - wewnątrz kanału chlorkowego (ryc 4.1). Funkcjonowanie kanału chlorkowego związane jest z aktywacją poszczegóL nych miejsc wiązań w obrębie kompleksu receptora GABA-A. Rola GABą w czynności OUN została poznana tylko w niewielkim stopniu i stosunkowo najlepiej zbadana w takich strukturach, jak substancja czarna, prążkowic oraz móżdżek. Stosunkowo dobrze udokumentowana jest rola GABA w Jęku oraz padaczce (p rozdz. 52). Leki anksjolityczne. do których należą przede wszystkim pochodne benzodiazepiny. nasilają powinowactwo GABA do receptora GABA-A, co w konsekwencji prowadzi do zwiększenia częstotliwości otwierania kanału chlorkowego i mechanizm ten miałby być odpowiedzialny za przeciwlękowe działanie tej grupy leków. W przeciwieństwie do działania leków anksjolitycznych. związi^ anksjogenne z grupy p-karbolin zmniejszają powinowactwo GABA do receptora GABA-A i częstotliwość otwarcia kanału chlorkowego, natomiast związej-drgawkotwórczy pikrotoksyna po związaniu się z miejscem receptorowym wewnątrz kanału chlorkowego utrzymuje go w stanie zamkniętym, a więc jego działanie związane jest z hamowaniem przewodnictwa chlorkowego.
Kanał wapniowy związany z receptorem NMDA
Pierwsze doniesienia o działaniu aminokwasów pobudzających na ośrodkowy układ nerwowy ukazały się w latach 50. XX w., gdy wykazano, że kwas glutaminowy i asparaginowy wywołują drgawki u zwierząt laboratoryjnych. Jednakże dopiero w' latach 80. XX w. zaczęto pow szechnie uznawać neuroprzekaźnikowe funkcje aminokwasów pobudzających. Układ glutaminergiczny nie tworzy zwartych szlaków, tak jak układ dopaminergiczny lub noradrenergiczny, lecz jest rozproszony w ca-k tym mózgu, podobnie jak neurony GABA-ergiczne. Obecnie dominuje pogląd, że I każda komórka nerwowa ma receptory' w-rażliw-e na aminokwasy pobudzające, co ■ może świadczyć o istotnej roli aminokwasów pobudzających w- funkcji OUN.
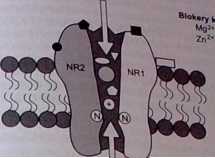
Kwas glutaminowy, a ściślej glutaminian, jest głównym pobudzającym neuro-f przekaźnikiem w mózgu oraz endogennym ligandem receptora wiążącego kwas N-metylo-D-asparaginowy (NMDA ). Podobnie jak w przypadku receptora GABA--A. receptor NMDA jest także złożonym kompleksem o wielu miejscach wiązania różnych endogennych ligandów biorących udział w regulacji częstotliwości otwarcia kanału wapniowego. I tak rozróżnia się miejsca wiązania specyficznych endogennych agonistówkwasu glutaminow ego oraz NMDA, endogennych związków nasilający ch funkcję receptora NMDA (miejsce glicynowe, miejsce poliamin) oraz miejsce wiązani a Mg-’* oraz Zn2* w obrębie kanału wapniowego i hamujące funkcję receptora (ryc. 4.2). Magnez osiąga w' mózgu stężenie ok. 2 mmol/1 i w tych warunkach, przy potencjale spoczynkowym - 70 m V, większość receptorów NMDA jest zablokow ana, ponieważ Mg2* jako jon naładowany dodatnio wciągany jest do wnętrza kanału wapniowego, gdzie łączy się ze swoistym miejscem, zamykając drogę dia innych jonów. Przy depolaryzacji neuronu i spadku różnicy potencjałów blokujące działanie magnezu znika.
Badania ostatnich łat wykazały, że fencyklidyna, środek znieczulający ogólnie, wprowadzony do lecznictwa na początku lat 60. XX w. i wycofany z powodu wyraźnych działań psychozomimetycznych, ma silne powinowactwo do receptora NMDA i blokuje kanał jonowy. Fizjologiczna rola tego miejsca jest nieznana, jak-
-"■-■ASfiKap
podjednostka NR 2B
miejsce wiązania.
- agonistów (glutaminian. NMDaj . antagonistów (AP,. łfonprodyi)
- modulatorów (poliamin) '
Ca?*/Na*
K*
4 2 Schemat receptora NMDA uwzględniający ml«8«__.
^~«u,ujWNRUNM_^^
i .....1 tńnv hnrlar7i>
Sei Kra N,',DA
kol" "A niektórzy badacze sugerują istnienie endoge„„vch ,io ^
np peptyd a-cndopsychozyna. Perspektywy tera^,1 8 "S® ™=j-
,;,rem NMDA są duże Szczególnie istou,awVajeS tw"WflnCzre“P-w „.ektórych ostrych stanach niedotlenienia, a także w nrzrwi^ 8'u,ami"°«go , odegeneracyjnych (p. rozdz. 56). Sugeruje się rolę aminXS *"•
J ...tających głownie poprzez receptor NMDA w etiologii takkh s,^“l^!),C1'ch u.ch, jak: padaczka, uszkodzenie mózgu (stany neut^enmSl „ u Uliku niedokrwienia, choroba Alzheimera, schizofrenia^^?? 1 kmsona. W związku z wyraźnymi dztalaniami psychozom,metyczńy4"S, blokujących kanał wapniowy receptora NMDA (typu fencyklidyny tabtolni m nadzieje związane z możliwością bezpiecznego hamowania komnlekś^-eń' lora NMDA przez tego typu związki wydają się nikle, jakkolwiek pewne naddek budzi memantyna (p. rozdz. 56). Zarnteresowania badaczy skupione są obecnie na miejscu glicynowym. gdyż. jak wynika z ostatnich badań, zastosowanie antagoni Stów glicyny (pochodne kwasu kinureninowego) prowadzi do hamowania aktywności kanału jonowego, co mogłoby mieć bardzo istotne znaczenie terapeutyczne
Kanały jonowe zależne od napięcia (potencjału)
Kanały jonowe zależne od napięcia regulują szybkie, zależne od potencjału zmiany \\ przepuszczalności jonów w komórkach pobudzających i neuronach. Rozróżnia się 3 typy zależnych od napięcia kanałów jonowych: potasowy, sodowy i wap-niou \ Najmniej skomplikowany pod względem budowy molekularnej i najbardziej pow szechny zarówno w świecie roślin, jak i zwierząt jest kanał potasowy; znacznie bardziej skomplikowany, zbudowany aż z 4 domen, jest zależny od na-
97
Wyszukiwarka
Podobne podstrony:
natomiast dla kanału wypełnionego częściowo (lys. 8 lb) promień hydrauliczny jest równy: a b rh “ 2
skanuj0106 (26) (5.9) 0 H-h S = —— - tg fi natomiast dla klina dwustronnego symetrycznego podaje się
skanowanie0052 (14) i uprzemysłowionych, wysokie natomiast dla województw rolniczych. Szczególnie du
instrukcja 9#7 sowania), natomiast dla dużych prądów wzbudzenia (tzn. dla dużych nasyceń obwodu magn
Zintegrowanatechnologia litowo-jonowa usną Przeznaczone zarówno dla prac wewnątrz, jaki na zewnątrz,
20587 P1010319 (2) Prostowniki natomiast dla przebiegu ciągłego obowiązujący jest wzór: H = Uck, cos
IMAG0414 (2) Cln)©IR©®Y WĄYROiY SPECYFICZNE DLA CIĄŻY © Wewnątrzwątrobowa cholestaza ciężarnych ® Os
poruszający sie -> 2 — , nieruchomy -» c 1 sek. dla obserwatora wewnątrz poruszającego się 1 r
CCF20110311�011 11 elektrostatyczne Fd są skierowane zgodnie z silami grawitacyjnymi F , natomiast d
natomiast dla stanu granicznego użytkowalności dla tzw.: - kombinacji rzadkiej, -
Jeżeli natomiast dla dużych wartości E b.kQ otrzymamy:A.a.kQ 0 (E) = -5- exp b OTA.a.kQ podstawiając
P1090433 122 w trakcie ciągnienia następuje pogrubienie ścianki ruty, natomiast dla rur cienkościenn
Działanie takie jest zakazane w rozmiarze w jakim może być szkodliwe dla handlu wewnątrz wspólnoty.
więcej podobnych podstron