DSC00757

swych właściwości, podczas gdy w komórkach nowotworowych przebudowa błony zachodzi bardzo powoli, dopiero w fazie S cyklu raitotycznego. Znaczenie przebudowy błony komórek prawidłowych w fazie Gi staje się widoczne dopiero przy gęstościach komórek wystarczająco dużych, aby mógł zaznaczyć się wpływ wzajemnych oddziaływali komórek na siebie. Błona komórkowa pełni wówczas funkcje regulujące w kontrolowaniu wzrostu komórek, ale nie należy zapominać, że właściwości błony określane są przez jej skład chemiczny zależny od genomu komórki. Za zmianę w budowie i właściwościach określonych białek kompleksu powierzchniowego komórki odpowiedzialne są poszczególne geny.
Mimo że w kontrolnych warunkach aktywność podziałowa komórek jest skorelowana z szybkością ich migracji, procesy te mogą zostać rozdzielone (rys. 29.4). W kontaktowo zahamowanych, stacjonarnych komórkach, można indukować syntezę DNA w rozmaity sposób, np. działając na błonę komórkową nadmiarem wielowartościowych kationów lub prądem elektrycznym, zwiększając stężenie surowicy, działając na błonę enzymami proteolitycznymi lub niejonowymi detergentami, zwiększając niespecyficznie przepuszczalność błony nadmiarem pozakomórkowe-go ATP, względnie bardziej specyficznie, działając na komórki np. insuliną lub swoistymi, peptydowymi czynnikami wzrostowymi. Także zahamowanie syntezy DNA za pomocą swoistych inhibitorów nie powoduje zahamowania migracji komórek. Podobnie korelację między kształtem komórek prawidłowych a ich aktywnością podziałową można zmienić rozmaitymi czynnikami, a zazwyczaj nie występuje ona w przypadku komórek nowotworowych.
W latach osiemdziesiątych wykazano, że zahamowanie syntezy DNA w gęstych hodowlach wynika z bezpośrednich, często bardzo swoistych, oddziaływań między składnikami kontaktujących się ze sobą komórek. Glikoproteiny obecne w błonie jednej komórki, najczęściej jako białka integralne, mogą swoiście oddziaływać z receptorami występującymi w błonie komórki sąsiedniej. W oddziaływania takie zaangażowane są białka integralne błon określane jako swoiste tkankowo białka adhezji międzykomórkowej CAM (z ang. — celi adhesion molecules) oraz tzw. kadheryny — białka odpowiedzialne za adhezję międzykomórkową, wrażliwe na jony wapnia. Są to białka zbudowane z jednego łańcucha polipeptydowego o długości 723-748 reszt aminokwasowych, raz przebijające dwuwarstwę lipidową, z miejscem receptorowym przy końcu wydłużonego, glikozylowanego fragmentu łańcucha po zewnętrznej stronie błony.

*

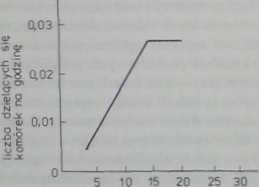
prędkość migrocji l,um/h)
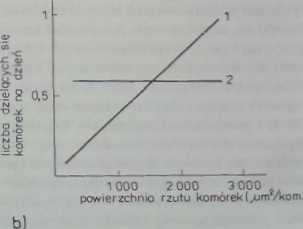

Rys. 29.4. W naturalnych warunkach hodowli komórek prawidłowych obserwuje się:
a — zależność aktywności mitotycznej od prędkości migracji komórek (nachylenie krzywej zależy od stężenia surowicy w pożywce): b — liniową zależność od stopnia ich rozpłaszczenia (krzywa BI V. aktywność podziałowa komćrck nowotworowych nie zależy od stopnia ich rozpłaszczenia (krzywa B2) (wg Castor 1970)

492
Wyszukiwarka
Podobne podstrony:
0000039 (12) rozmnażania komórek, podczas którego komórkom nowotworzonym zostaje przez komórki macie
DSCF0048 SVEN ERIK ASTRÓM głównie w dobrach szlacheckich, podczas gdy owies odgrywał ważną rolę u za
CCF20090321�014 podczas gdy udział właściciela domu gry wynosi zawsze ponad lft/o wkładów *, co przy
DSC00728 CYSTERNA SER SRyŁ 28.10. Aktywacja cyklazy guanylanowej podczas odpowiedzi komórkowej. Rece
DSC00756 aglutynin) niemal wyłącznie komórki nowotworowe. Chociaż w hodowlach in vitro aglutyniny wy
DSC00760 — w komórkach nowotworowych obserwuje się zmniejszenie ilości fi lamentów
10780 Pok tafil1544 wydzielają Insulinę. Stanowią one główną masę komórkową wysp i układają się w ic
FizykaII389�01 385 słoneeznem okazują, właściwy innobarwny połysk, jeśli spoglądamy pa nie z góry, p
skanuj0015 Biologia nowotworzenia — Ćwiczenie 3 Zaobserwuj: przy pow. 200x komórki nowotworowe tworz
Image321 układ, na wejścia którego są podawane jednocześnie wszystkie bity porównywanych liczb, podc
więcej podobnych podstron