1.Teoria Hugo de Vriesa
Teoria Hugo de Vries (studia dzienne)
- gatunki powstają skokowo a poszczególne skoki są zjawiskiem, które można obserwować jak każdy inny proces fizjologiczny
skokowe zmiany nazywa mutacjami"zwykła" zmienność jest ciągła i zachodzi stale
cechy gatunkowe są kombinacją różnorodnych cech dziedzicznych Cechy dziedziczą się niezależnie
2. Teoria Darwina ( podać przykłady
Obserwacje:
- potencjalna liczba potomstwa danego gatunku jest nieskończona (teoretycznie może zasiedlić całą ziemię)
- rodzi się zawsze więcej potomstwa niż może przeżyć
- wzrost liczbowo danej populacji ograniczany jest przez kurczące się zasoby.
Wnioski:
- osobniki wykazujące cechy dziedziczne najlepiej przystosowujące je do środowiska, mają największą szansę przeżycia i przekazania tych korzystnych cech swojemu potomstwu.
- dziedziczone cechy, korzystne we współzawodnictwie z innymi osobnikami będą stawały się coraz częstsze w populacji
- wymusza to ciągłą walkę organizmów żywych o przeżycie i rozród (WALKA O BYT)
- cechy które są dziedziczne i pomagają przeżyć będą liczniejsze w kolejnych pokoleniach
- prowadzi to w konsekwencji do powstania nowych gatunków
Darwin zakładał:
- odmiany stanowią początek gatunków
- pomiędzy niewiele różniącymi się rasami, a w pełni rozdzielonymi gatunkami istnieje zmienność ciągła
- stopniowa przemiana to ewolucja
Teoria Darwina została opublikowana w dziele pt. "O pochodzeniu gatunków" w 1859 r. po tym jak Alfred Wallace niezależnie od niego doszedł do tych samych wniosków
Darwin wykazał, że problem zmienności i dziedziczności może być rozwiązany przez obserwację bezpośrednią. Dowiódł, że zmienność o charakterze ewolucyjnym rzeczywiście istnieje, a zmienność ta ma charakter ciągły. Określił różnice w zmienności środowiskowej a genetycznej
3. Prawa Mendla
Historyczne pojęcie genu:
- „gen” wprowadzono w 1910 roku na określenie abstrakcyjnej, niepodzielnej jednostki dziedziczenia, odpowiedzialnej za jakąś cechę danego gatunku. Prowadzona przez wiele pokoleń analiza statystyczna dziedziczenia się prostych cech w populacjach potwierdziła tę koncepcję genu.
Interpretacja praw Mendla:
- każdy organizm ma dwa allele dla każdej pojedynczej dziedziczonej cechy - po jednym od każdego rodzica.
- w każdym pokoleniu dwa allele jednego genu są rozdzielane (segregowane niezależnie) podczas powstawania nowych komórek rozrodczych.
* w każdej haploidalnej komórce rozrodczej jest tylko jeden element wyjściowej pary, jeden allel.
* w wyniku zapłodnienia powstaje nawa kombinacja alleli.
* dwa elementy z pary mogą być takie same - wtedy osobnik jest nazywany homozygotycznym (identyczne elementy) pod względem tej pary alleli. I odwrotnie, osobnik może otrzymać różne allele od obojga rodziców i wtedy jest heterozygotyczny (różnią się).
Pierwsze prawo Mendla zwane prawem segregacji lub prawem czystości gamet:
Każdy organizm posiada dwa czynniki (obecnie zwane genami allelicznymi), determinujące ekspresję danej cechy, które organizm otrzymuje od swoich rodziców, po jednym od każdego, natomiast w komórkach rozrodczych ( gametach) znajduje się zawsze tylko jeden taki czynnik.
II Prawo Mendla - prawo niezależnego dziedziczenia
Allele dwóch różnych genów są segregowane w gametogenezie niezależnie od siebie. Cechy dziedziczą się niezależnie, o ile geny determinujące te cechy nie są sprzężone, tzn. są zlokalizowane w różnych, niehomologicznych chromosomach.
Prawa dziedziczenia cech zostały powtórnie odkryte w 1900 niezależnie przez trzech uczonych C. Corrensa, E. Tschermaka i H. de Vriesa.
4. Omówić pojęcia: fenotyp, genotyp, mieszańce, cecha, dominowanie, allel (na rysunku), kodominacja, heterozygota, homozygota, modyfikatory, allele wielokrotne geny plejotropowe, locus, gameta, epistaza,itd.
Fenotyp:- wygląd osobnika, czyli zespół jego dostrzegalnych własności, będący efektem działania specyficznych genów lub ich kombinacji zapisanych w genotypie
Genotyp - zespół genów danego osobnika
Mieszańce - osobniki powstałe ze złączenia się gamet o różnym składzie genetycznym
Cecha - właściwość organizmu wynikająca z interakcji genotypowo-środowiskowej
Dominowanie - forma współdziałania allelicznego, polegająca na tym, iż w stanie heterozygotycznym jeden z alleli w większym lub mniejszym stopniu przeważa nad drugim
Pojęcie allel:
O ile gen jest strukturą DNA, odpowiedzialną za wykształcenie cechy, to allel jest jego realizacją. Jeden gen może występować w kilku formach; różne formy genów będą odpowiadać za wykształcenie różnych cech fenotypowych. Każda z tych form będzie allelem.
Allel
jeden z pary lub serii genów alternatywnych w procesie dziedziczenia (wskutek usytuowania w tym samym locus w homologicznych chromosomach), wykluczających się wzajemnie w konstytucji gamety
jedna z alternatywnych form genu występująca w tym samym miejscu genowym (locus) homologicznych chromosomów. Gdy allele są identyczne - osobnik jest homozygotą pod względem tej pary alleli, gdy różne - heterozygotą
Allele - różne formy tego samego genu. Chociaż zajmują to samo miejsce (locus) w chromosomach homologicznych, ale wywołują odmienne wykształcenie tej samej cechy. W komórce diploidalnej znajdują się dwa allele każdego genu (po jednym od każdego z rodziców). Jeden allel jest często dominujący w stosunku do drugiego nazywanego recesywnym, tzn. determinuje ujawnienie się określonej cechy organizmu.
Allele addytywne - kumultatywne w swoim działaniu
Allele kodominujące - obydwa przejawiają ekspresję u heterozygot
Allele letalne - wpływające na żywotność osobnika, często odpowiedzialne za procesy rozwojowe. (1 zmutowany allel - zmiany rozwojowe: 2 zmutowane allele - śmierć)
Allele subletalne - obniżające żywotność (szczątkowe skrzydła u muszki owocowej)
Kodominacja (dziedziczenie pośrednier)- heterozygota posiada cechy równocześnie obu alleli rodzicielskich.
Modyfikatory - geny wpływające na ekspresję genu nieallelicznego
Allele wielokrotne - trzy lub więcej alternatywnych alleli danego genu, reprezentujących to samo miejsce w danej parze chromosomów homologicznych
Przykład: gen barwy kwiatów u tulipanów
Geny plejotropowe - jeden gen wpływa równocześnie na kilka własności
Przykład: 1. u wyżlinu recesywny allel barwy białej kwiatu wpływa na zmniejszenie rozmiaru rośliny i zwiększa podatność na wybrane choroby grzybowe
Gameta - dojrzała komórka rozrodcza: plemnik i jajo
Gen i locus
Gen jest fragmentem DNA niosącym informację genetyczną i stanowiącym jego fizyczną oraz funkcjonalną jednostkę. Geny są umiejscowione w określonych regionach DNA określanych jako genetyczne loci (w liczbie pojedynczej: locus).
Pojęcie locus:
Locus
- pozycja na chromosomie genu lub innego markeru chromosomowego, jak również, DNA w tej pozycji. Pojęcie locus używa się czasami w odniesieniu do regionu DNA ulegającemu ekspresji.
Loci QTL - miejsca genów odpowiedzialnych za dziedziczenie poligeniczne określonych cech.
Epistaza jest to współdziałanie genów nieallelicznych lub interakcja pomiędzy produktami genów nieallelicznych
5. Co to jest gen (rozwiń temat)
Współczesna chemia genu ma swoje początki w 1869 roku, kiedy to Johann Friedrich Miecher odkrył, że materiał wyizolowany z jąder ludzkich komórek nie jest białkiem, ponieważ zawiera fosfor oraz jest odporny na działanie enzymu rozkładającego białka - pepsyny. Miescher nazwał odkrytą przez siebie substancję nukleiną, Dzisiaj nosi ona nazwę kwasu deoksyrybonukleinowego, czyli DNA.
Badacze zidentyfikowali podstawowe elementy budulcowe DNA: cztery odrębne małe cząsteczki nazwane nukleotydami. Podczas badań właściwości fizycznych okazało się, że DNA jest polimerem, to znaczy bardzo długą, podobną do łańcucha cząsteczką zbudowaną z nukleotydów. Ponadto odkryto, że większość komórkowego DNA znajduje się w chromosomach. dwaj młodzi naukowcy pracujący w Uniwersytecie w Cambridge w Anglii, James Watson i Francis Crick ustalili, że DNA ma strukturę podwójnej helisy. Wagę odkrycia struktury DNA dodatkowo podniosły wyniki wcześniejszych badań Oswalda Avery'ego i jego współpracowników z Instytutu Rockefellera w Nowym Jorku oraz Alfreda Hersheya i Margaret Chase z Uniwersytetu im. Waszyngtona w St. Louis w stanie Missouri, którzy dowiedli, że nośnikiem informacji genetycznej jest wyłącznie DNA. Główną rolę w dziedziczeniu, przypisywaną kiedyś chromosomom, przekazano teraz z całym zaufaniem chronosomowemu DNA.
Wnioskiem płynącym z tych badań była koncepcja genu jako informacji - takiej, która rządzi wzrostem i zachowaniem istot żywych oraz decyduje o ich cechach. Koncepcja ta zakłada zarazem, że wszystkie żywe istoty dysponują mechanizmem rozszyfrowującym albo „odczytującym” informację genetyczną, a geny przechowują stale tę informację, aby rodzice mogli przekazać ją swemu potomstwu.
Pojęcia genu:
jednostka informacji genetycznej zawierająca instrukcję dotyczącą syntezy polipeptydu lub cząsteczki strukturalnego RNA. W sensie fizycznym gen jest odcinkiem DNA o określonej sekwencji zasad. (Krótkie wykłady - Genetyka)
podstawowa fizyczna i funkcjonalna jednostka dziedziczenia. Gen jest odcinkiem DNA o określonej sekwencji zasad zlokalizowanym w określonej pozycji, na określonym chromosomie, kodującym specyficznie funkcjonalny produkt (białko lub cząsteczkę RNA)
Gen i locus
Gen jest fragmentem DNA niosącym informację genetyczną i stanowiącym jego fizyczną oraz funkcjonalną jednostkę. Geny są umiejscowione w określonych regionach DNA określanych jako genetyczne loci (w liczbie pojedynczej: locus).
6. Przedstawić i omówić na rysunku budowę chromosomu organizmów wyższych
Chromosomy - strukturalne jednostki jądra zawierające chromatynę i będące nośnikiem informacji genetycznej zawartej w ułożonych w porządku liniowym genach. Chromosomy są strukturami samoodtwarzającymi się, których ilość w komórce kształt oraz organizacja są charakterystycznymi cechami gatunkowymi.
Mejozę poprzedza interfaza, w czasie której replikuje się DNA, podwajają się białka chromosomowe i gromadzone są wszystkie czynniki potrzebne do przebiegu mejozy. Po tej fazie następują 2 podziały mejotyczne. W pierwszym redukcyjnym odbywa się koniugacja chromosomów homologicznych, zachodzi crossing-over, następuje redukcja liczby chromosomów i odbywa się synteza białek. Drugi podział mejotyczny przypomina podział mitotyczny .
7. Omówić przebieg mitozy i mejozy (wskazane na rysunkach)
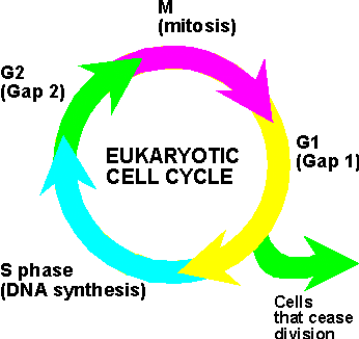
8. Omówić i narysować cykl życiowy komórki
Cykl komórkowy to okres między dwoma podziałami, od powstania komórki do podziału na dwie komórki potomne. Jest to proces cykliczny.
-Życie komórek można podzielić na wiele powtarzających się okresów. Każdy taki okres to jeden cykl komórkowy (inaczej: cykl życiowy komórki).
-Pojedynczy cykl komórkowy można podzielić na kilka głównych faz, które reguluje zegar biologiczny:
Faza G1 - wysoka aktywność metaboliczna, wzrost komórki wypełnianie przeznaczonych jej funkcji.
Faza S (synteza) - replikacja DNA;
Faza G2 - końcowe przygotowania do podziału. Po zakończeniu tej fazy rozpoczyna się stadium profazy
Faza M - mitoza - podział, w którym powstają dwie komórki potomne o tej samej liczbie chromosomów
Obie komórki powstające podczas mitozy wchodzą w fazę G1 i cykl komórkowy zaczyna się od nowa
|
|
U organizmów wielokomórkowych większość komórek nie ulega podziałom i pełni swoją funkcję życiową. U roślin najwięcej komórek w stadium podziału znajduje się w tkankach merystematycznych ( u zwierząt np. w szpiku kostnym).
Rozpoczęcie podziału komórek (faza G1) następuje po otrzymaniu odpowiedniego sygnału ze strony komórek sąsiadujących. Sygnał ten wywołują hormonalne czynniki wzrostu w procesie zwanym transdukcją sygnałów. Proces ten polega na „przekazaniu” sygnału jaki daje hormon wzrostu na nić DNA, powodując jej transkrypcję. Przechodzenie komórki przez poszczególne fazy cyklu jest napędzane prze z ewolucyjnie konserwatywny mechanizm regulujący, precyzyjnie kontrolowany. Ten mechanizm składa się z dwóch głównych rodzajów białek - cyklin i kinaz.
Cykliny są białkami regulatorowymi, których poziom w komórce zmienia się w różnych fazach cyklu komórkowego. W fazie G2 następuje stopniowe nagromadzenie się cyklin i przekroczenie ich progowej koncentracji. Po przekroczeniu progowej koncentracji cykliny te łączą się z enzymami należącymi do grupy kinaz. Połączenie się cyklin z kinazami powoduje, że enzymy te stają się aktywne. Podczas mitozy cykliny ulegają degradacji powodując, iż enzymy należące do grupy kinaz stają się nieaktywne umożliwiając komórce zakończenie mitozy.
Kierowanie cyklem zasadniczo polega na jego hamowaniu w specyficznych punktach kontrolnych, między fazą G1 i S, G2 i mitozą oraz w trakcie mitozy. Komórki zawierające błędy w zapisie DNA w prawidłowo funkcjonującej komórce nie wchodzą w fazę. Zabezpiecza to organizm przed powstawaniem mutacji. Komórki z niekompletnie zreplikowanym DNA nie mogą wejść w stadium mitozy. Zaburzenia w każdym etapie mitozy powodują przerwanie cyklu komórkowego.
Mutacje genów biorących udział w procesie transdukcji sygnałów mogą być przyczyną raka. Geny te (zmutowane) nazywamy onkogenami
Regulacja podziałów komórkowych
-Duże znaczenie w regulacji podziałów komórkowych odgrywają telomery. Telomery znajdują się na końcach chromosomów i chronią chromosomy przed sklejaniem podczas podziałów i innymi uszkodzeniami. Jednak w fazie S, czyli podczas replikacji DNA, telomery nieznacznie się skracają osiągając po kilkudziesięciu podziałach wartość progową . Stanowi to sygnał dla komórki iż należy przejść w etap starzenia się, a potem śmierci.
-Niektóre komórki wytwarzają enzym zwany telomerazą, który potrafi wydłużać telomery. Takie komórki mogą dzielić się dłużej niż inne. Telomerazę produkują m.in. komórki zarodkowe, komórki macierzyste szpiku kostnego, komórki jelit i inne komórki, które szybko się dzielą. Wiele komórek nowotworowych też wytwarza telomerazę; uważa się, że między innymi z tego powodu komórki raka mogą dzielić się tak szybko i w niekontrolowany sposób.
Niektóre komórki, na przykład te, w których doszło do poważnych uszkodzeń DNA, opuszczają cykl komórkowy i umierają na drodze programowanej śmierci, czyli apoptozy.
9. Apoptoza - omówić przebieg oraz wyjaśnić sens stwierdzenia „śmierć dla życia” (obowiązkowy rysunek)
"śmierć jest potrzebna dla życia"
Przykłady pozytywnej apoptozy w rozwoju płodu u człowieka: błony między palcami , owłosienie, komórki w mózgu
Błędy w apoptozie - choroby Alzheimera, Parkinsona, AIDS oraz autoimmunologiczne
Nowotwory - komórki powstałe w wyniku zaistnienia błędu w DNA.
Apoptoza daje "życie" dzięki:
eliminacji komórek rakowych
likwidacji nadmiaru komórek
usuwaniu komórek układu immulogicznego po wykonaniu zadania
usuwanie komórek wadliwych i niepełnowartościowych
Dzięki apoptozie komórki "nie zaśmiecają" organizmu.
Przebieg apoptozy:
Sygnał przychodzi od sąsiednich komórek poprzez tzw. receptory śmierci lub od własnego DNA. Sygnał przekazywany jest do mitochondriów. Toksyczna zawartość mitochondriów jest wylewana do komórki, odblokowane zostają enzymy restrykcyjne (tnące DNA i białka). Po zakończeniu fragmentacji DNA komórka gwałtownie rozpada się na wiele ciałek apoptycznych. Ciałka te są wchłaniane przez organizm.
Odkrywcy apoptozy otrzymali w 2002 r nagrodę Nobla
Sydnej Brenner - współodkrywca mRNA, wprowadził do badań jako organizm modelowy nicienia (Caenorhabditis elegans)
Robert Horvitz - stwierdza, że apoptoza stanowi integralną część rozwoju organizmu. Opisuje system przekazywania przez komórki "sąsiadki" sygnału do rozpoczęcia lub opóźnienia apoptozy
John Sulston - opisał po raz pierwszy cały genom nicienia. Stwierdza konserwatyzm genów odpowiedzialnych za apoptozę.
10. Teoria Morgana
- geny znajdujące się w chromosomie mają swoje locus, czyli miejsce występowania i ułożone są liniowo, w określonym porządku.
- dla każdej cechy istnieją dwa geny: 1 w chromosomie matczynym i 1 w chromosomie ojcowskim, są one dokładnie naprzeciw siebie i tworzą parę alleliczną
- Geny należące do jednej pary Aa nazwał Morgan - allelomorfami, gdyż wywołują dwie postacie tej samej cechy
- Między allelami istnieje współdziałanie.
Jeżeli osobnik ma jeden gen o cesze recesywnej i jeden gen o cesze dominującej to jest heterozygotą. aA (lub Aa). Heterozygota Aa (lub aA) ma mniej czerwonego barwnika (antocyjanu) niż homozygota AA o cechach dominujących AA. Gen A koduje więc enzymy zapewniające syntezę barwnika nadającego barwę kwiatu. U wyżlinu można wzrokowo zauważyć mniejszą zawartość barwnika czerwonego u heterozygoty - barwa różowa. Osobniki homozygotyczne AA mają więcej barwnika, bo są aż dwa geny kodujące syntezę barwnika, stąd intensywnie czerwona barwa. U grochu heterozygotycznego też można stwierdzić mniejszą zawartość barwnika, jednakże nie wzrokowo, lecz poprzez ilościowe metody analityczne.
11. Sprzężenia genów (zilustruj rysunkiem). Mapowanie genetyczne. Interferencja.
- Zjawisko sprzężenia to tendencja alleli do przechodzenia razem z pokolenia na pokolenie. --- Sprzężenia wykazują jedynie geny znajdujące się na tym samym chromosomie . Sprzężenie zostaje zerwane wówczas, gdy nastąpi wymiana odcinków (crossing over).
-geny leżące na różnych chromosomach nie są sprzężone i wykazują 50% rekombinacji
CROSSING OVER
- Rearanżacja genów leżących na tym samym chromosomie może nastąpić tylko w wyniku zajścia zjawiska crossing over
-stopień sprzężenia dwóch genów zależy od częstości crossing over między nimi podczas mejozy
- Im bliżej geny są zlokalizowane na chromosomie, tym mniej prawdopodobne jest wystąpienie między nimi crossing over.
- Grupy genów, które są sprzężone nazywamy grupami sprzężeń genów. Geny znajdujące się na chromosomach niehomologicznych z definicji nie wykazują sprzężenia.
Opierając się na dwóch zjawiskach: stałości w procencie wymiany między tymi samymi parami genów sprzężonych oraz występowaniu różnic w procencie wymiany między różnymi parami genów, Morgan wysunął hipotezę, że częstość wymiany jest proporcjonalna do odległości między genami znajdującymi się w jednym chromosomie
Mapowanie chromosomów
- mapowanie chromosomów polega nakreśleniu kolejności i odległości genów sprzężonych na odpowiednim chromosomie oraz identyfikacji chromosomu u którego przeprowadza mapowanie
-procent rekombinacji między dwoma genami położonymi na tym samym chromosomie informuje, jak daleko od siebie są one położone. Jeden procent rekombinacji równa się jednostce mapowej lub centymorganowi (cM). Obecnie badania sprzężeń genów przeprowadza się najczęściej z wykorzystaniem markerów molekularnych.
Mapowanie za pomocą krzyżówki dwucechowej (przykład z książki Genetyka -krótkie wyklady)
Krzyżowano pomidory: PPHH (purpurowe owoce i owłosiona łodyga)
X
Pphh (czerwone owoce, gładka łodyga)
F1 purpurowe owoce i owłosiona łodyga
Przeprowadzono krzyżowanie testowe (z homozygotą recesywną). Gdyby geny nie były sprzężone, winno się uzyskać stosunek rozszczepień między cechami 1:1:1:1
Uzyskano cztery typy potomstwa: 1 i 2 reprezentują typy form rodzicielskich (patrz tabela poniżej), klasy 3 i 4 są rekombinantami (prezentują nowe kombinacje alleli obu genów).
Fenotypy uzyskane w wyniku krzyżówki testowej między pokoleniem F1 pomidorów o purpurowych owocach i owłosionej łodydze (Pp, Hh), a podwójną recesywną homozygotą rodzicielską o czerwonych owocach i gładkiej łodydze (pp, hh)
Mapowanie na podstawie krzyżówki dwugenowej
-krzyżowanie dwugenowe polegają na skrzyżowaniu podwójnej heterozygoty z podwójną homozygotą(tzw. Krzyżówka testowa)
-proporcja gamet zrekombinowanych wyraża częstość rekombinacji
- procent rekombinacji między dwoma genami informuje jak daleko są od siebie położone
Mapowanie za pomocą krzyżowań "trójgenowych" (przykład z tego samego podręcznika)
- pozwala na ustalenie kolejności genów oraz ich odległości mapowej, przynajmniej na krótkich odcinkach chromosomu.
- pozwalają na sporządzenie map genetycznych
Interferencja
- Crossing over zachodzi między chromosomami homologicznymi w miejscach określanych jako hiazmy.
- interferencji pozytywnej pojawienie się jednej wymiany między chromosomami homologicznymi redukuje prawdopodobieństwo zajścia crossing over w sąsiedztwie,
- Interferencja ujemna negatywna - wystąpienie jednej wymiany między chromosomami homologicznymi zwiększa prawdopodobieństwo innej w sąsiedztwie pierwszej.
12. Poliploidy - wymień i opisz ich rodzaje, mechanizm powstawania, przykłady. Omów przebieg mejozy u tych form
Poliploidy - rośliny posiadające więcej niż dwa genomy
Poliploidy powstają wtedy, gdy chromosomy diploidalne ulegną podziałowi, lecz nie podzieli się komórka.
Wyróżnia się dwa procesy prowadzące do powstania poliploidów
Zaburzenia w mitozie, w komórkach merystematycznych
Cechą charakterystyczną jest pojawianie się chimer na roślinach
Zakłócenia w podziale mejotycznym a II podział odbywa się prawidłowo
Poliploidy stanowią około 30% roślin okrytozalążkowych. W wielu przypadkach są bujniejsze od form diploidalnych, nie jest to jednak reguła. Charakterystyczne są dla flor młodych
Róża - znane są gatunki diploidalne o 14 chromosomach, triploidalne o 21, tetraploidalne o 56 chromosomach.
Chryzantema - między innymi dekaploidalne o 90 chromosomach
Rodzaje poliploidów
Autopoliploidy - poliploidy, u których w komórkach somatycznych znajdują się te same genomy, każdy reprezentowany co najmniej dwukrotnie (np. AAAA). Powstają bezpośrednio z diploidów w wyniku zwielokrotnienia liczby chromosomów w komórkach somatycznych lub też zlania się gamet o niezredukowanej liczbie chromosomów. Poliploidy są często mniej płodne od form diploidalnych.
Przykłady:
płodnych tetraploidów : nagietki, wyżlin
z zaburzeniami: bieluń.
Formy ploidalne są często bujniejsze od diploidów np. narcyz 2x, 3x i 4x lub tytoń .
Dzięki ewolucji formy tetraploidalne występujące naturalnie w przyrodzie są w pełni płodne i lepiej o diploidów przystosowane do warunków środowiska.
Autotriploidy - powstają w wyniku krzyżowania tetraploida z diploidem (np. AAAA x aa daje gamety AA i a z których połączenia otrzymuje się triploidy AAa)
Przykład: triploidalne i beznasienne banany, melony oraz tulipany (charakteryzują się dłuższym okresem kwitnienia).
Autotetraploidy - powstają w wyniku zwielokrotnienia 2x liczby chromosomów do 4x .
W przypadku cechy kontrolowanej jedną parą genów Aaaa stosunek rozszczepień wynosi 1: 35
Przykłady: kawa, narcyz, hiacynty, tulipany. Autotetraploidy narcyz są zawsze niebieskie
14. Allopoliploidy, wymień i opisz ich rodzaje, omów na dowolnym przykładzie mechanizm ich powstawania
Allopoliploidy - poliploidy, u których w komórkach somatycznych znajdują się różne genomy, każdy reprezentowany dwukrotnie (np. AABBDD). Są organizmami, w których komórkach znajdują się podwojone genomy pochodzące z różnych gatunków w różnym stopniu ze sobą spokrewnionych.
Przykłady: śliwa domowa - powstała w czasach przedhistorycznych ze skrzyżowania formy 2x z 4x i podwojenia u triploida liczby chromosomów.
Truskawka - powstała ze skrzyżowania dwóch oktoploidalnych gatunków.
Pszenica o 2n = 6 x 42 chromosomy i składzie genomowym AABBDD. Dawcą genomu A jest Triticum boeoticum, B - Aegilops speltoides a C - Aegilops squarrosa.
Pszenżyto
Autoallopoliploidy - allopoliploidy , u których niektóre genomy są reprezentowane w liczbie większej niż dwa (np. AAABB)
Przykład: tymotka
Alloploidy segmentalne - poliploidy, u których w chromosomach należących do różnych genomów występują odcinki homologiczne
13. Aneuploidy, wymień i opisz ich rodzaje, mechanizm powstawania, przykłady. Omów przebieg mejozy u tych form
Aneuploidy - są to formy nie będące dokładną wielokrotnością całkowitej liczby genomowej.
2n-1 - nullisomik (brak pary chromosomów homologicznych)
2n-1 - monosomik (brak jednego chromosomu z danej pary chromosomowej)
Aneuploidami nazywamy również formy o nadmiarze chromosomów homologicznych w danej parze np. trisomik (2n+1 , tetrasomik (2n+2)
Przykład: maciejka. Dodatek od 1-2 chromosomów wpływa na zwężenie liści.
14. Aberracje chromosomowe, wymień i opisz ich rodzaje, mechanizm powstawania, przykłady. Omów przebieg mejozy u tych form
Aberracje chromosomowe
Aby mogło dojść do zmiany w strukturze chromosomów (aberracji) muszą nastąpić w chromosomie pęknięcia
Deficjencje - utrata przez chromosomy niektórych odcinków wraz z informacją genetyczną w nich zawartą
Np. chromosom normalny ABCD --- deficj
Duplikacja - zdwojenie niektórych odcinków w chromosomach
np. AABCD
Inwersja - odwrócenie niektórych odcinków w obrębie chromosomu
np. ADCB
Translokacja - wymiana niektórych odcinków między niehomologicznymi chromosomami
15. Omów systemy determinacji płci
Determinacja płci u organizmów wyższych
Płeć osobnika może być determinowana na kilku poziomach. Pierwszorzędowe cechy płciowe dotyczą formowania jajników (zalążni) lub jąder (pylników). Drugorzędowe dotyczą wyglądu fenotypowego osobnika.
Osobniki posiadające jednocześnie męskie i żeńskie organy rozrodcze nazywamy hermafrodytycznymi (obojnactwo). Zjawisko to jest powszechne u bezkręgowców i roślin
Systemy determinacji płci
dwa allele pojedynczego genu określają płeć np. drożdże
rozwój osobników męskich z niezapłodnionych jaj (np. pszczoły, mrówki, osy)
układy chromosomów płci
Układ XX-X0. Samice mają parę jednakowych chromosomów zwanych chromosomami X. Samce maja tylko jeden chromosom (np. koniki polne)
Układ XX-XY. Samice mają dwie kopie chromosomu X. Samce mają jeden chromosom X, a drugi Y (ssaki, niektóre owady). Samice są homogametyczne, to znaczy wytwarzają gamety zawierające wyłącznie chromosom X
Układ ZZ-ZW. Samice mają jeden chromosom Z a drugi W Samce mają dwie pary chromosomu ZZ (ptaki). Samice są heterogametyczne, to znaczy wytwarzają gamety z chromosomami Z i W (X i Y) z jednakową częstością.
Większość roślin okrytozalążkowych jest obupłciowa.
U roślin rozdzielnopłciowych (dwupiennych) kwiaty męskie wyrastają na innym osobniku, niż kwiaty żeńskie Przykłady: Melandrium, przestęp dwupienny, ogórek.
16. Chromosomy płciowe u Melandrium (0mówić szczegółowo)
Chromosomy płciowe u Melandrium
U tej rośliny osobniki żeńskie zawierają chromosomy XX, a męskie XY. Y znajdują się regiony hamujące rozwój cech żeńskich, a stymulujących męskich. Poza tym posiadają po 22 autosomy.
Chromosom Y u Melandrium często ulega defragmentacji.
Gdy chromosom Y utraci odcinek ze satelitą to rośliny są hermafrodytyczne (kwiaty dwupłciowe). Z tego wynika, iż w tym odcinku chromosomu są geny tłumiące rozwój żeńskich organów płciowych - rozwijają się tylko męskie
Utrata odcinka z telomerami powoduje powstawanie nieżywotnego pyłku (podziały redukcyjne następują jednak w sposób prawidłowy).
Dwupienność roślin jest wynikiem mutacji dwóch genów ściśle ze sobą sprzężonych (odkrył to Westergaard) : M i T .
M - gen lub grupa genów ściśle sprzężonych w obecności których tworzą się kwiaty obupłciowe
m - gen lub grupa genów ściśle sprzężonych w obecności których tworzą się kwiaty żeńskie
T - genu tłumiący żeńskość
t - jego allel recesywny.
wtedy:
MmTt - rośliny męskie
mmtt - rośliny żeńskie
MMtt - hermafrodytyczne (kwiaty obupłciowe)
17. Omów na dowolnym przykładzie dziedziczenie cech sprzężonych z płcią (daltonizm?, kolor oczu u owadow?)
Cechy sprzężone z płcią
Wszystkie cechy, które dziedziczą się wraz z chromosomami płci nazywamy cechami sprzężonymi z płcią, a sposób dziedziczenia określamy jako dziedziczeniem sprzężonym z płcią.
Allele recesywne położone na chromosomie X nie mogą się przejawiać fenotypowo u heterozygotycznych samic (u ssaków), lecz są widoczne u samców ponieważ mają tylko jeden chromosom
Przykład: daltonizm
18. Dziedziczenie cytoplazmatyczne (omów na przykładzie dziwaczka, nie wystarczy definicja, co wiesz na temat dziedziczenia plastydów)
Dziedziczenie cytoplazmatyczne
Zespół genów zlokalizowanych w chromosomach nazywamy genomem
Zespół czynników dziedzicznych znajdujących się w cytoplaźmie nazywamy plazmonem (plazmidem) a geny pozajądrowe - plazmogenami
Plazmogeny (plazmidy) są zlokalizowane w organoidach cytoplazmatycznych jak plastydy , mitochondria, centriole i inne
Dziedziczenie własności plastydów
Plastydy powstają z już istniejących struktur (rozmnażają się przez podział i są przekazywane z jednej komórki do drugiej), a niektóre cechy plastydów nie są przekazywane za pośrednictwem chromosomów.
Pstrolistność dziwaczków
Pyłek z pędów o typie ulistnienia |
Kwiaty zapylone na pędzie ulistnienia |
Potomstwo nasienne o typie ulistnienia |
Bladozielonym |
bladozielonym zielonym pstrym |
bladozielonym zielonym bladozielonym, zielonym, pstrym |
Zielonym |
bladozielonym zielonym pstrym |
bladozielonym zielonym bladozielonym, zielonym, pstrym |
Pstrym |
bladozielonym zielonym Pstrym |
bladozielonym zielonym pstrym |
Łatwo zauważyć, że rodzaj pyłku nie ma tu żadnego znaczenia. Czynnikiem decydującym jest gameta żeńska. Podane wyniki można tłumaczyć obecnością dwóch rodzajów plastydów (patrz tabelka poniżej)
Typ ulistnienia pędów |
Typ plastydów przekazywanych przez formę mateczną |
Bladozielone Zielone Pstre |
bladozielone zielone bladozielone i zielone |
19. Przedstawić na rysunku mechanizm cytoplazmatycznej męskiej sterylności (przykłady wykorzystania)
Geny cytoplazmatyczne mogą współdziałać z genami jądrowymi. Wykorzystuje się to zjawisko do otrzymywania roślin z cytoplazmatyczną sterylnością.
Cytoplazmatyczno-genetyczna męska niepłodność jest uwarunkowana współdziałaniem cytoplazmy (S) i recesywnych genów jądrowych (ms ms). Geny (ms ms) są odpowiedzialne za męską sterylność i nie funkcjonują w obecności cytoplazmy N. Rośliny z całkowicie męskoniepłodne posiadają więc genotyp (S ms ms). Do rozmnażania roślin męskoniepłodnych wykorzystuje się rośliny o genotypie (N ms ms). Są to tzw. linie dopełniające.
S ms ms x N ms ms ------ F1 S ms ms
męskosterylna płodna męskosterylna
Cytoplazmatyczną męską niepłodność wykorzystuje się w hodowli heterozyjnej.
Linię będącą zapylaczem w produkcji nasion heterozyjnych nazywamy linią restorerującą
20. Podstawowe definicje dotyczące genetyki populacji
Genetyka populacji zajmuje się badaniem jak rozkładają się geny pokolenia rodzicielskiego na osobniki pokolenia potomnego.
Populacja naturalna - wiele krzyżujących się zróżnicowanych genetycznie osobników, dzielących tą samą pulę genową.
Populacja lokalna obejmuje wszystkie osobniki, które mają udział w tej samej lokalnej puli genowej (np. dla owadów: drzewo, las).
Populacja mendlowska - zbiór osobników, które mają możliwość wzajemnego kojarzenia się.
Gatunek - wszystkie osobniki, które tworzą wspólnotę rozrodczą i których lokalne pule genowe pozostają w stałym kontakcie.
Częstość allelu określa się przez dodanie liczby kopii każdego allelu w populacji i przedstawienie jej jako częstości.
21. Omówić Prawo Hardy'ego-Weinberga
Prawo to przedstawia stan dynamicznej równowagi w populacji, na którą nie działają siły selekcyjne ani też żadne inne czynniki.
W dużej, losowo kojarzonej populacji, przy braku migracji, mutacji i selekcji, częstość genów i genotypów jest stała; populacja sama w sobie nie ma tendencji do zmiany swoich genetycznych właściwości z pokolenia na pokolenie.
Częstości genotypów z potomstwa, powstałego z losowego kojarzenia rodziców , są uwarunkowane jedynie częstością genów tych rodziców
populacja, znajdująca się w stanie równowagi Hardy- Weinberga, charakteryzuje się w każdym pokoleniu współzależnością między częstością genu i genotypów wyrażona równaniem:
p2 : 2pq : q2
gdzie:
p- częstość genotypu AA,
q - częstość genotypu aa,
pq -częstość genotypu Aa
częstość genotypów zgodna z prawem Hardy - Weinberga ustala się w populacji, po zastosowaniu losowego kojarzenia w jednym pokoleniu, niezależnie od genotypu rodziców
22. Wyprowadzenie wzoru na prawo Hardy'ego-Weinberga (tylko studia dzienne)
Częstość allelu M w populacji 1199/2000 = 0.5995,
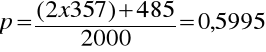
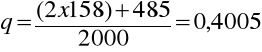
częstość allelu N w populacji 801/2000 = 0.4005.W populacji 1000 białych Amerykanów 357 z nich było MM, 485 MN i 158 NN, 357 osón MM posiadało 714 alleli M a 485 osób MN miało kolejne 485 alleli M, tak więc częstość allelu M (-p) wynosi
Podobnie częstość N wynosi
Na zmianę genetycznych właściwości populacji mogą wpływać następujące czynniki:
wielkość populacji (podczas gdy duża populacja jest stosunkowo niewrażliwa na losowe odchylenia składu genetycznego, to w populacji małej za sprawą przypadku mogą zajść nieodwracalne zmiany)
dobór (selekcja) - oddziaływuje na żywotność czy płodność osobników lub gamet poszczególnych genotypów. Usuwa np. wadliwe allele, a równowaga częstości wystąpi wtedy, kiedy tempo powstawania nowych alleli jest równe tempu powstawania nowych mutacji
różnice w płodności i żywotności (wśród rodziców , osobniki o różnych genotypach mogą wykazywać różną płodność)
dryf genetyczny - czynnik przypadkowości, proces losowy zmiany częstości genu, szczególnie ostro występujący w małych populacjach (pojedynczy allel nie występuje dokładnie w 50%)
migracja - przepływ genów między subpopulacjami
mutacja - zmiana w sekwencji zasad DNA, mogąca wywołać zmiany fenotypowe u organizmów żywych
Przykład faworyzowania w doborze naturalnym heterozygot:
Allison (1954r) wykrył, że w Afryce ludzie ze skazą sierpowatą (Ss) cechują się znaczną odpornością na malarię. Chorzy o genotypie SS umierają na anemię, natomiast heterozygoty maja przewagę w spadku umieralności na malarię około 25 % nad homozygotami (ss)
23.Podaj podstawowe różnice miedzy cechami ilościowymi jakościowymi
Genetyka ilościowa:
- genetyka ilościowa zajmuje się dziedziczeniem różnic występujących między osobnikami w stopniu natężenia cech, a nie w ich jakości.
- każdy organ lub funkcja jakiegokolwiek gatunku wykazuje indywidualne różnice, np. różnice w wysokości wśród ludzi.
- osobniki tworzą szereg ciągły, wzrastający od najniższej wartości do najwyższej a nie dzielą się na wyraźnie różniące się od siebie typy.
- różnice jakościowe dzielą natomiast osobniki na wyraźnie różniące się od siebie typy bez lub występowaniem (rzadziej) typów pośrednich.
- istnienie różnic ilościowych i jakościowych w stosunku do badanego zjawiska stwarza konieczność odróżnienia genetyki ilościowej od jakościowej.
- genetyka cech ilościowych w odróżnieniu do genetyki cech jakościowych bada różnice spowodowane działaniem większej ilości genów.
Dziedzicznie uwarunkowane różnice ilościowe zależą od różnic występujących w wielu loci miedzy genami, których efekty nie mogą być indywidualnie określone. W wyniku tego zjawiska, mendlowskie stosunki liczbowe nie uwidaczniają się w różnicach ilościowych, a metody genetyki mendlowskiej nie nadają się do badania tych różnic. Podstawowym założeniem genetyki ilościowej jest twierdzenie, że dziedziczenie różnic ilościowych zależy od genów podlegającym tym samym prawom co dziedziczenie cech jakościowych. Genetyka ilościowa jest więc dalszym rozwinięciem genetyki Mendla i opiera się na prawach Mendla.
24. Współczynniki odziedziczalności
Odziedziczalność ogólna (współczynnik odziedziczalności) w szerokim sensie służy do określenia jaką część zmienności całkowitej stanowi zmienność genetyczna, a więc jest to stosunek zmienności genetycznej do całkowitej
Odziedziczalność w wąskim sensie jest to stosunek wariancji addytywnej do zmienności całkowitej
25.Ocena efektów działania genów
Efekty addytywne: wynikają ze sumowania się efektów wywoływanych przez wiele loci w stanie homozygotycznym.
Efekty dominowania - występują na skutek działania genów w stanie heterozygotycznym.
Niealleliczna interakcja (EPISTAZA) - wiąże się z:
współdziałaniem genów w stanie heterozygotycznym
współdziałaniem genów w stanie homozygotycznym
współdziałaniem genów w stanie homozygotycznym i heterozygotycznym
Znajomość efektów działania genów może przyczynić się do zwiększenia efektywności hodowli roślin.
Badanie efektów działania genów wymaga przeprowadzania specjalnie w tym celu zakładanych doświadczeń.
Liczba czynników efektywnych - jest to liczba genów lub grup genów ściśle ze sobą sprzężonych, którymi różnią się badane formy rodzicielskie
Współczynnik rozkładu genów u form rodzicielskich
dyspersja genów występuje w przypadku zgrupowania u każdej z form rodzicielskich równej liczby genów zwiększających jak i zmniejszajacych wartość danej cechy
asocjacja genów występuje w przypadku kumulacji (+) alleli u jednej z form rodzicielskich i (-) alleli u drugiej z form rodzicielskich
26.Opisz zjawisko heterozji, teorie tłumaczące powstawanie tego zjawiska
Heterozja - wyższość genotypów heterozygotycznych nad wyjściowymi homozygotami w odniesieniu do jednej lub więcej cech, stanowiący fenotypowy efekt współdziałania genów.
Teorie dotyczące występowania efektów heterozji
Istnieje kilka teorii tłumaczących powstawanie zjawiska heterozji. Należą one do jednej z dwóch kategorii:
Przyczyną heterozji jest stan heterozygotyczny loci. Zakłada się, że geny A i a wytwarzają różne związki, które albo przez połączenie lub produktu ich współdziałania powodują większą bujność niż każdy z pojedynczych związków produkowanych przez każdy z alleli w stanie homozygotycznym
Przyczyną heterozji jest współdziałanie genów dominujących. Heterozja jest skutkiem dominowania genów sprzyjającym bujności nad odpowiednimi genami recesywnymi przeciwdziałającymi bujności.
3. Wystąpienie efektu heterozji zależy od genów dopełniających (podobnie jak chromogen i oksydaza są koniecznymi do powstania antocjanu). Współdziałanie genomu z plazmonem
27. Zjawisko transgresji genetycznej
Depresja wsobna - spadek żywotności (wigoru) organizmów wywołany zapłodnieniem formy matecznej przez blisko spokrewnioną formę ojcowską. Zjawisko to występuje zarówno u roślin samopylnych jak i u zwierząt
Transgresja - pojawianie się wśród segregującego pokolenia osobników o cechach przekraczających zakres zmienności form rodzicielskich. Efekt transgresji związany jest ze stanem homozygotycznym loci.
1.Teoria Hugo de Vriesa
Teoria Hugo de Vries (studia dzienne)
- gatunki powstają skokowo a poszczególne skoki są zjawiskiem, które można obserwować jak każdy inny proces fizjologiczny
- skokowe zmiany nazywa mutacjami"zwykła" zmienność jest ciągła i zachodzi stale
- cechy gatunkowe są kombinacją różnorodnych cech dziedzicznych Cechy dziedziczą się niezależnie
2. Teoria Darwina ( podać przykłady
Obserwacje:
- potencjalna liczba potomstwa danego gatunku jest nieskończona (teoretycznie może zasiedlić całą ziemię)
- rodzi się zawsze więcej potomstwa niż może przeżyć
- wzrost liczbowo danej populacji ograniczany jest przez kurczące się zasoby.
Wnioski:
- osobniki wykazujące cechy dziedziczne najlepiej przystosowujące je do środowiska, mają największą szansę przeżycia i przekazania tych korzystnych cech swojemu potomstwu.
- dziedziczone cechy, korzystne we współzawodnictwie z innymi osobnikami będą stawały się coraz częstsze w populacji
- wymusza to ciągłą walkę organizmów żywych o przeżycie i rozród (WALKA O BYT)
- cechy które są dziedziczne i pomagają przeżyć będą liczniejsze w kolejnych pokoleniach
- prowadzi to w konsekwencji do powstania nowych gatunków
Darwin zakładał:
- odmiany stanowią początek gatunków
- pomiędzy niewiele różniącymi się rasami, a w pełni rozdzielonymi gatunkami istnieje zmienność ciągła
- stopniowa przemiana to ewolucja
- Teoria Darwina została opublikowana w dziele pt. "O pochodzeniu gatunków" w 1859 r. po tym jak Alfred Wallace niezależnie od niego doszedł do tych samych wniosków
- Darwin wykazał, że problem zmienności i dziedziczności może być rozwiązany przez obserwację bezpośrednią. Dowiódł, że zmienność o charakterze ewolucyjnym rzeczywiście istnieje, a zmienność ta ma charakter ciągły. Określił różnice w zmienności środowiskowej a genetycznej
3. Prawa Mendla
Historyczne pojęcie genu:
- „gen” wprowadzono w 1910 roku na określenie abstrakcyjnej, niepodzielnej jednostki dziedziczenia, odpowiedzialnej za jakąś cechę danego gatunku. Prowadzona przez wiele pokoleń analiza statystyczna dziedziczenia się prostych cech w populacjach potwierdziła tę koncepcję genu.
Interpretacja praw Mendla:
- każdy organizm ma dwa allele dla każdej pojedynczej dziedziczonej cechy - po jednym od każdego rodzica.
- w każdym pokoleniu dwa allele jednego genu są rozdzielane (segregowane niezależnie) podczas powstawania nowych komórek rozrodczych.
* w każdej haploidalnej komórce rozrodczej jest tylko jeden element wyjściowej pary, jeden allel.
* w wyniku zapłodnienia powstaje nawa kombinacja alleli.
* dwa elementy z pary mogą być takie same - wtedy osobnik jest nazywany homozygotycznym (identyczne elementy) pod względem tej pary alleli. I odwrotnie, osobnik może otrzymać różne allele od obojga rodziców i wtedy jest heterozygotyczny (różnią się).
Pierwsze prawo Mendla zwane prawem segregacji lub prawem czystości gamet:
Każdy organizm posiada dwa czynniki (obecnie zwane genami allelicznymi), determinujące ekspresję danej cechy, które organizm otrzymuje od swoich rodziców, po jednym od każdego, natomiast w komórkach rozrodczych ( gametach) znajduje się zawsze tylko jeden taki czynnik.
II Prawo Mendla - prawo niezależnego dziedziczenia
Allele dwóch różnych genów są segregowane w gametogenezie niezależnie od siebie. Cechy dziedziczą się niezależnie, o ile geny determinujące te cechy nie są sprzężone, tzn. są zlokalizowane w różnych, niehomologicznych chromosomach.
Prawa dziedziczenia cech zostały powtórnie odkryte w 1900 niezależnie przez trzech uczonych C. Corrensa, E. Tschermaka i H. de Vriesa.
4. Omówić pojęcia: fenotyp, genotyp, mieszańce, cecha, dominowanie, allel (na rysunku), kodominacja, heterozygota, homozygota, modyfikatory, allele wielokrotne geny plejotropowe, locus, gameta, epistaza,itd.
Fenotyp:- wygląd osobnika, czyli zespół jego dostrzegalnych własności, będący efektem działania specyficznych genów lub ich kombinacji zapisanych w genotypie
Genotyp - zespół genów danego osobnika
Mieszańce - osobniki powstałe ze złączenia się gamet o różnym składzie genetycznym
Cecha - właściwość organizmu wynikająca z interakcji genotypowo-środowiskowej
Dominowanie - forma współdziałania allelicznego, polegająca na tym, iż w stanie heterozygotycznym jeden z alleli w większym lub mniejszym stopniu przeważa nad drugim
Pojęcie allel:
O ile gen jest strukturą DNA, odpowiedzialną za wykształcenie cechy, to allel jest jego realizacją. Jeden gen może występować w kilku formach; różne formy genów będą odpowiadać za wykształcenie różnych cech fenotypowych. Każda z tych form będzie allelem.
Allel
jeden z pary lub serii genów alternatywnych w procesie dziedziczenia (wskutek usytuowania w tym samym locus w homologicznych chromosomach), wykluczających się wzajemnie w konstytucji gamety
jedna z alternatywnych form genu występująca w tym samym miejscu genowym (locus) homologicznych chromosomów. Gdy allele są identyczne - osobnik jest homozygotą pod względem tej pary alleli, gdy różne - heterozygotą
Allele - różne formy tego samego genu. Chociaż zajmują to samo miejsce (locus) w chromosomach homologicznych, ale wywołują odmienne wykształcenie tej samej cechy. W komórce diploidalnej znajdują się dwa allele każdego genu (po jednym od każdego z rodziców). Jeden allel jest często dominujący w stosunku do drugiego nazywanego recesywnym, tzn. determinuje ujawnienie się określonej cechy organizmu.
Allele addytywne - kumultatywne w swoim działaniu
Allele kodominujące - obydwa przejawiają ekspresję u heterozygot
Allele letalne - wpływające na żywotność osobnika, często odpowiedzialne za procesy rozwojowe. (1 zmutowany allel - zmiany rozwojowe: 2 zmutowane allele - śmierć)
Allele subletalne - obniżające żywotność (szczątkowe skrzydła u muszki owocowej)
Kodominacja (dziedziczenie pośrednier)- heterozygota posiada cechy równocześnie obu alleli rodzicielskich.
Modyfikatory - geny wpływające na ekspresję genu nieallelicznego
Allele wielokrotne - trzy lub więcej alternatywnych alleli danego genu, reprezentujących to samo miejsce w danej parze chromosomów homologicznych
Przykład: gen barwy kwiatów u tulipanów
Geny plejotropowe - jeden gen wpływa równocześnie na kilka własności
Przykład: 1. u wyżlinu recesywny allel barwy białej kwiatu wpływa na zmniejszenie rozmiaru rośliny i zwiększa podatność na wybrane choroby grzybowe
Gameta - dojrzała komórka rozrodcza: plemnik i jajo
Gen i locus
Gen jest fragmentem DNA niosącym informację genetyczną i stanowiącym jego fizyczną oraz funkcjonalną jednostkę. Geny są umiejscowione w określonych regionach DNA określanych jako genetyczne loci (w liczbie pojedynczej: locus).
Pojęcie locus:
Locus
- pozycja na chromosomie genu lub innego markeru chromosomowego, jak również, DNA w tej pozycji. Pojęcie locus używa się czasami w odniesieniu do regionu DNA ulegającemu ekspresji.
Loci QTL - miejsca genów odpowiedzialnych za dziedziczenie poligeniczne określonych cech.
Epistaza jest to współdziałanie genów nieallelicznych lub interakcja pomiędzy produktami genów nieallelicznych
5. Co to jest gen (rozwiń temat)
Współczesna chemia genu ma swoje początki w 1869 roku, kiedy to Johann Friedrich Miecher odkrył, że materiał wyizolowany z jąder ludzkich komórek nie jest białkiem, ponieważ zawiera fosfor oraz jest odporny na działanie enzymu rozkładającego białka - pepsyny. Miescher nazwał odkrytą przez siebie substancję nukleiną, Dzisiaj nosi ona nazwę kwasu deoksyrybonukleinowego, czyli DNA.
Badacze zidentyfikowali podstawowe elementy budulcowe DNA: cztery odrębne małe cząsteczki nazwane nukleotydami. Podczas badań właściwości fizycznych okazało się, że DNA jest polimerem, to znaczy bardzo długą, podobną do łańcucha cząsteczką zbudowaną z nukleotydów. Ponadto odkryto, że większość komórkowego DNA znajduje się w chromosomach. dwaj młodzi naukowcy pracujący w Uniwersytecie w Cambridge w Anglii, James Watson i Francis Crick ustalili, że DNA ma strukturę podwójnej helisy. Wagę odkrycia struktury DNA dodatkowo podniosły wyniki wcześniejszych badań Oswalda Avery'ego i jego współpracowników z Instytutu Rockefellera w Nowym Jorku oraz Alfreda Hersheya i Margaret Chase z Uniwersytetu im. Waszyngtona w St. Louis w stanie Missouri, którzy dowiedli, że nośnikiem informacji genetycznej jest wyłącznie DNA. Główną rolę w dziedziczeniu, przypisywaną kiedyś chromosomom, przekazano teraz z całym zaufaniem chronosomowemu DNA.
Wnioskiem płynącym z tych badań była koncepcja genu jako informacji - takiej, która rządzi wzrostem i zachowaniem istot żywych oraz decyduje o ich cechach. Koncepcja ta zakłada zarazem, że wszystkie żywe istoty dysponują mechanizmem rozszyfrowującym albo „odczytującym” informację genetyczną, a geny przechowują stale tę informację, aby rodzice mogli przekazać ją swemu potomstwu.
Pojęcia genu:
jednostka informacji genetycznej zawierająca instrukcję dotyczącą syntezy polipeptydu lub cząsteczki strukturalnego RNA. W sensie fizycznym gen jest odcinkiem DNA o określonej sekwencji zasad. (Krótkie wykłady - Genetyka)
podstawowa fizyczna i funkcjonalna jednostka dziedziczenia. Gen jest odcinkiem DNA o określonej sekwencji zasad zlokalizowanym w określonej pozycji, na określonym chromosomie, kodującym specyficznie funkcjonalny produkt (białko lub cząsteczkę RNA)
Gen i locus
Gen jest fragmentem DNA niosącym informację genetyczną i stanowiącym jego fizyczną oraz funkcjonalną jednostkę. Geny są umiejscowione w określonych regionach DNA określanych jako genetyczne loci (w liczbie pojedynczej: locus).
6. Przedstawić i omówić na rysunku budowę chromosomu organizmów wyższych
Chromosomy - strukturalne jednostki jądra zawierające chromatynę i będące nośnikiem informacji genetycznej zawartej w ułożonych w porządku liniowym genach. Chromosomy są strukturami samoodtwarzającymi się, których ilość w komórce kształt oraz organizacja są charakterystycznymi cechami gatunkowymi.
Mejozę poprzedza interfaza, w czasie której replikuje się DNA, podwajają się białka chromosomowe i gromadzone są wszystkie czynniki potrzebne do przebiegu mejozy. Po tej fazie następują 2 podziały mejotyczne. W pierwszym redukcyjnym odbywa się koniugacja chromosomów homologicznych, zachodzi crossing-over, następuje redukcja liczby chromosomów i odbywa się synteza białek. Drugi podział mejotyczny przypomina podział mitotyczny .
7. Omówić przebieg mitozy i mejozy (wskazane na rysunkach)
8. Omówić i narysować cykl życiowy komórki
Cykl komórkowy to okres między dwoma podziałami, od powstania komórki do podziału na dwie komórki potomne. Jest to proces cykliczny.
-Życie komórek można podzielić na wiele powtarzających się okresów. Każdy taki okres to jeden cykl komórkowy (inaczej: cykl życiowy komórki).
-Pojedynczy cykl komórkowy można podzielić na kilka głównych faz, które reguluje zegar biologiczny:
Faza G1 - wysoka aktywność metaboliczna, wzrost komórki wypełnianie przeznaczonych jej funkcji.
Faza S (synteza) - replikacja DNA;
Faza G2 - końcowe przygotowania do podziału. Po zakończeniu tej fazy rozpoczyna się stadium profazy
Faza M - mitoza - podział, w którym powstają dwie komórki potomne o tej samej liczbie chromosomów
Obie komórki powstające podczas mitozy wchodzą w fazę G1 i cykl komórkowy zaczyna się od nowa
U organizmów wielokomórkowych większość komórek nie ulega podziałom i pełni swoją funkcję życiową. U roślin najwięcej komórek w stadium podziału znajduje się w tkankach merystematycznych ( u zwierząt np. w szpiku kostnym).
Rozpoczęcie podziału komórek (faza G1) następuje po otrzymaniu odpowiedniego sygnału ze strony komórek sąsiadujących. Sygnał ten wywołują hormonalne czynniki wzrostu w procesie zwanym transdukcją sygnałów. Proces ten polega na „przekazaniu” sygnału jaki daje hormon wzrostu na nić DNA, powodując jej transkrypcję. Przechodzenie komórki przez poszczególne fazy cyklu jest napędzane prze z ewolucyjnie konserwatywny mechanizm regulujący, precyzyjnie kontrolowany. Ten mechanizm składa się z dwóch głównych rodzajów białek - cyklin i kinaz.
Cykliny są białkami regulatorowymi, których poziom w komórce zmienia się w różnych fazach cyklu komórkowego. W fazie G2 następuje stopniowe nagromadzenie się cyklin i przekroczenie ich progowej koncentracji. Po przekroczeniu progowej koncentracji cykliny te łączą się z enzymami należącymi do grupy kinaz. Połączenie się cyklin z kinazami powoduje, że enzymy te stają się aktywne. Podczas mitozy cykliny ulegają degradacji powodując, iż enzymy należące do grupy kinaz stają się nieaktywne umożliwiając komórce zakończenie mitozy.
Kierowanie cyklem zasadniczo polega na jego hamowaniu w specyficznych punktach kontrolnych, między fazą G1 i S, G2 i mitozą oraz w trakcie mitozy. Komórki zawierające błędy w zapisie DNA w prawidłowo funkcjonującej komórce nie wchodzą w fazę. Zabezpiecza to organizm przed powstawaniem mutacji. Komórki z niekompletnie zreplikowanym DNA nie mogą wejść w stadium mitozy. Zaburzenia w każdym etapie mitozy powodują przerwanie cyklu komórkowego.
Mutacje genów biorących udział w procesie transdukcji sygnałów mogą być przyczyną raka. Geny te (zmutowane) nazywamy onkogenami
Regulacja podziałów komórkowych
-Duże znaczenie w regulacji podziałów komórkowych odgrywają telomery. Telomery znajdują się na końcach chromosomów i chronią chromosomy przed sklejaniem podczas podziałów i innymi uszkodzeniami. Jednak w fazie S, czyli podczas replikacji DNA, telomery nieznacznie się skracają osiągając po kilkudziesięciu podziałach wartość progową . Stanowi to sygnał dla komórki iż należy przejść w etap starzenia się, a potem śmierci.
-Niektóre komórki wytwarzają enzym zwany telomerazą, który potrafi wydłużać telomery. Takie komórki mogą dzielić się dłużej niż inne. Telomerazę produkują m.in. komórki zarodkowe, komórki macierzyste szpiku kostnego, komórki jelit i inne komórki, które szybko się dzielą. Wiele komórek nowotworowych też wytwarza telomerazę; uważa się, że między innymi z tego powodu komórki raka mogą dzielić się tak szybko i w niekontrolowany sposób.
Niektóre komórki, na przykład te, w których doszło do poważnych uszkodzeń DNA, opuszczają cykl komórkowy i umierają na drodze programowanej śmierci, czyli apoptozy.
9. Apoptoza - omówić przebieg oraz wyjaśnić sens stwierdzenia „śmierć dla życia” (obowiązkowy rysunek)
"śmierć jest potrzebna dla życia"
Przykłady pozytywnej apoptozy w rozwoju płodu u człowieka: błony między palcami , owłosienie, komórki w mózgu
Błędy w apoptozie - choroby Alzheimera, Parkinsona, AIDS oraz autoimmunologiczne
Nowotwory - komórki powstałe w wyniku zaistnienia błędu w DNA.
Apoptoza daje "życie" dzięki:
eliminacji komórek rakowych
likwidacji nadmiaru komórek
usuwaniu komórek układu immulogicznego po wykonaniu zadania
usuwanie komórek wadliwych i niepełnowartościowych
Dzięki apoptozie komórki "nie zaśmiecają" organizmu.
Przebieg apoptozy:
Sygnał przychodzi od sąsiednich komórek poprzez tzw. receptory śmierci lub od własnego DNA. Sygnał przekazywany jest do mitochondriów. Toksyczna zawartość mitochondriów jest wylewana do komórki, odblokowane zostają enzymy restrykcyjne (tnące DNA i białka). Po zakończeniu fragmentacji DNA komórka gwałtownie rozpada się na wiele ciałek apoptycznych. Ciałka te są wchłaniane przez organizm.
Odkrywcy apoptozy otrzymali w 2002 r nagrodę Nobla
Sydnej Brenner - współodkrywca mRNA, wprowadził do badań jako organizm modelowy nicienia (Caenorhabditis elegans)
Robert Horvitz - stwierdza, że apoptoza stanowi integralną część rozwoju organizmu. Opisuje system przekazywania przez komórki "sąsiadki" sygnału do rozpoczęcia lub opóźnienia apoptozy
John Sulston - opisał po raz pierwszy cały genom nicienia. Stwierdza konserwatyzm genów odpowiedzialnych za apoptozę.
10. Teoria Morgana
- geny znajdujące się w chromosomie mają swoje locus, czyli miejsce występowania i ułożone są liniowo, w określonym porządku.
- dla każdej cechy istnieją dwa geny: 1 w chromosomie matczynym i 1 w chromosomie ojcowskim, są one dokładnie naprzeciw siebie i tworzą parę alleliczną
- Geny należące do jednej pary Aa nazwał Morgan - allelomorfami, gdyż wywołują dwie postacie tej samej cechy
- Między allelami istnieje współdziałanie.
Jeżeli osobnik ma jeden gen o cesze recesywnej i jeden gen o cesze dominującej to jest heterozygotą. aA (lub Aa). Heterozygota Aa (lub aA) ma mniej czerwonego barwnika (antocyjanu) niż homozygota AA o cechach dominujących AA. Gen A koduje więc enzymy zapewniające syntezę barwnika nadającego barwę kwiatu. U wyżlinu można wzrokowo zauważyć mniejszą zawartość barwnika czerwonego u heterozygoty - barwa różowa. Osobniki homozygotyczne AA mają więcej barwnika, bo są aż dwa geny kodujące syntezę barwnika, stąd intensywnie czerwona barwa. U grochu heterozygotycznego też można stwierdzić mniejszą zawartość barwnika, jednakże nie wzrokowo, lecz poprzez ilościowe metody analityczne.
11. Sprzężenia genów (zilustruj rysunkiem). Mapowanie genetyczne. Interferencja.
- Zjawisko sprzężenia to tendencja alleli do przechodzenia razem z pokolenia na pokolenie. --- Sprzężenia wykazują jedynie geny znajdujące się na tym samym chromosomie . Sprzężenie zostaje zerwane wówczas, gdy nastąpi wymiana odcinków (crossing over).
-geny leżące na różnych chromosomach nie są sprzężone i wykazują 50% rekombinacji
CROSSING OVER
- Rearanżacja genów leżących na tym samym chromosomie może nastąpić tylko w wyniku zajścia zjawiska crossing over
-stopień sprzężenia dwóch genów zależy od częstości crossing over między nimi podczas mejozy
- Im bliżej geny są zlokalizowane na chromosomie, tym mniej prawdopodobne jest wystąpienie między nimi crossing over.
- Grupy genów, które są sprzężone nazywamy grupami sprzężeń genów. Geny znajdujące się na chromosomach niehomologicznych z definicji nie wykazują sprzężenia.
Opierając się na dwóch zjawiskach: stałości w procencie wymiany między tymi samymi parami genów sprzężonych oraz występowaniu różnic w procencie wymiany między różnymi parami genów, Morgan wysunął hipotezę, że częstość wymiany jest proporcjonalna do odległości między genami znajdującymi się w jednym chromosomie
Mapowanie chromosomów
- mapowanie chromosomów polega nakreśleniu kolejności i odległości genów sprzężonych na odpowiednim chromosomie oraz identyfikacji chromosomu u którego przeprowadza mapowanie
-procent rekombinacji między dwoma genami położonymi na tym samym chromosomie informuje, jak daleko od siebie są one położone. Jeden procent rekombinacji równa się jednostce mapowej lub centymorganowi (cM). Obecnie badania sprzężeń genów przeprowadza się najczęściej z wykorzystaniem markerów molekularnych.
Mapowanie za pomocą krzyżówki dwucechowej (przykład z książki Genetyka -krótkie wyklady)
Krzyżowano pomidory: PPHH (purpurowe owoce i owłosiona łodyga)
X
Pphh (czerwone owoce, gładka łodyga)
F1 purpurowe owoce i owłosiona łodyga
Przeprowadzono krzyżowanie testowe (z homozygotą recesywną). Gdyby geny nie były sprzężone, winno się uzyskać stosunek rozszczepień między cechami 1:1:1:1
Uzyskano cztery typy potomstwa: 1 i 2 reprezentują typy form rodzicielskich (patrz tabela poniżej), klasy 3 i 4 są rekombinantami (prezentują nowe kombinacje alleli obu genów).
Fenotypy uzyskane w wyniku krzyżówki testowej między pokoleniem F1 pomidorów o purpurowych owocach i owłosionej łodydze (Pp, Hh), a podwójną recesywną homozygotą rodzicielską o czerwonych owocach i gładkiej łodydze (pp, hh)
Mapowanie na podstawie krzyżówki dwugenowej
-krzyżowanie dwugenowe polegają na skrzyżowaniu podwójnej heterozygoty z podwójną homozygotą(tzw. Krzyżówka testowa)
-proporcja gamet zrekombinowanych wyraża częstość rekombinacji
- procent rekombinacji między dwoma genami informuje jak daleko są od siebie położone
Mapowanie za pomocą krzyżowań "trójgenowych" (przykład z tego samego podręcznika)
- pozwala na ustalenie kolejności genów oraz ich odległości mapowej, przynajmniej na krótkich odcinkach chromosomu.
- pozwalają na sporządzenie map genetycznych
Interferencja
- Crossing over zachodzi między chromosomami homologicznymi w miejscach określanych jako hiazmy.
- interferencji pozytywnej pojawienie się jednej wymiany między chromosomami homologicznymi redukuje prawdopodobieństwo zajścia crossing over w sąsiedztwie,
- Interferencja ujemna negatywna - wystąpienie jednej wymiany między chromosomami homologicznymi zwiększa prawdopodobieństwo innej w sąsiedztwie pierwszej.
12. Poliploidy - wymień i opisz ich rodzaje, mechanizm powstawania, przykłady. Omów przebieg mejozy u tych form
Poliploidy - rośliny posiadające więcej niż dwa genomy
Poliploidy powstają wtedy, gdy chromosomy diploidalne ulegną podziałowi, lecz nie podzieli się komórka.
Wyróżnia się dwa procesy prowadzące do powstania poliploidów
Zaburzenia w mitozie, w komórkach merystematycznych
Cechą charakterystyczną jest pojawianie się chimer na roślinach
Zakłócenia w podziale mejotycznym a II podział odbywa się prawidłowo
Poliploidy stanowią około 30% roślin okrytozalążkowych. W wielu przypadkach są bujniejsze od form diploidalnych, nie jest to jednak reguła. Charakterystyczne są dla flor młodych
Róża - znane są gatunki diploidalne o 14 chromosomach, triploidalne o 21, tetraploidalne o 56 chromosomach.
Chryzantema - między innymi dekaploidalne o 90 chromosomach
Rodzaje poliploidów
Autopoliploidy - poliploidy, u których w komórkach somatycznych znajdują się te same genomy, każdy reprezentowany co najmniej dwukrotnie (np. AAAA). Powstają bezpośrednio z diploidów w wyniku zwielokrotnienia liczby chromosomów w komórkach somatycznych lub też zlania się gamet o niezredukowanej liczbie chromosomów. Poliploidy są często mniej płodne od form diploidalnych.
Przykłady:
płodnych tetraploidów : nagietki, wyżlin
z zaburzeniami: bieluń.
Formy ploidalne są często bujniejsze od diploidów np. narcyz 2x, 3x i 4x lub tytoń .
Dzięki ewolucji formy tetraploidalne występujące naturalnie w przyrodzie są w pełni płodne i lepiej o diploidów przystosowane do warunków środowiska.
Autotriploidy - powstają w wyniku krzyżowania tetraploida z diploidem (np. AAAA x aa daje gamety AA i a z których połączenia otrzymuje się triploidy AAa)
Przykład: triploidalne i beznasienne banany, melony oraz tulipany (charakteryzują się dłuższym okresem kwitnienia).
Autotetraploidy - powstają w wyniku zwielokrotnienia 2x liczby chromosomów do 4x .
W przypadku cechy kontrolowanej jedną parą genów Aaaa stosunek rozszczepień wynosi 1: 35
Przykłady: kawa, narcyz, hiacynty, tulipany. Autotetraploidy narcyz są zawsze niebieskie
14. Allopoliploidy, wymień i opisz ich rodzaje, omów na dowolnym przykładzie mechanizm ich powstawania
Allopoliploidy - poliploidy, u których w komórkach somatycznych znajdują się różne genomy, każdy reprezentowany dwukrotnie (np. AABBDD). Są organizmami, w których komórkach znajdują się podwojone genomy pochodzące z różnych gatunków w różnym stopniu ze sobą spokrewnionych.
Przykłady: śliwa domowa - powstała w czasach przedhistorycznych ze skrzyżowania formy 2x z 4x i podwojenia u triploida liczby chromosomów.
Truskawka - powstała ze skrzyżowania dwóch oktoploidalnych gatunków.
Pszenica o 2n = 6 x 42 chromosomy i składzie genomowym AABBDD. Dawcą genomu A jest Triticum boeoticum, B - Aegilops speltoides a C - Aegilops squarrosa.
Pszenżyto
Autoallopoliploidy - allopoliploidy , u których niektóre genomy są reprezentowane w liczbie większej niż dwa (np. AAABB)
Przykład: tymotka
Alloploidy segmentalne - poliploidy, u których w chromosomach należących do różnych genomów występują odcinki homologiczne
13. Aneuploidy, wymień i opisz ich rodzaje, mechanizm powstawania, przykłady. Omów przebieg mejozy u tych form
Aneuploidy - są to formy nie będące dokładną wielokrotnością całkowitej liczby genomowej.
2n-1 - nullisomik (brak pary chromosomów homologicznych)
2n-1 - monosomik (brak jednego chromosomu z danej pary chromosomowej)
Aneuploidami nazywamy również formy o nadmiarze chromosomów homologicznych w danej parze np. trisomik (2n+1 , tetrasomik (2n+2)
Przykład: maciejka. Dodatek od 1-2 chromosomów wpływa na zwężenie liści.
14. Aberracje chromosomowe, wymień i opisz ich rodzaje, mechanizm powstawania, przykłady. Omów przebieg mejozy u tych form
Aberracje chromosomowe
Aby mogło dojść do zmiany w strukturze chromosomów (aberracji) muszą nastąpić w chromosomie pęknięcia
Deficjencje - utrata przez chromosomy niektórych odcinków wraz z informacją genetyczną w nich zawartą
Np. chromosom normalny ABCD --- deficj
Duplikacja - zdwojenie niektórych odcinków w chromosomach
np. AABCD
Inwersja - odwrócenie niektórych odcinków w obrębie chromosomu
np. ADCB
Translokacja - wymiana niektórych odcinków między niehomologicznymi chromosomami
15. Omów systemy determinacji płci
Determinacja płci u organizmów wyższych
Płeć osobnika może być determinowana na kilku poziomach. Pierwszorzędowe cechy płciowe dotyczą formowania jajników (zalążni) lub jąder (pylników). Drugorzędowe dotyczą wyglądu fenotypowego osobnika.
Osobniki posiadające jednocześnie męskie i żeńskie organy rozrodcze nazywamy hermafrodytycznymi (obojnactwo). Zjawisko to jest powszechne u bezkręgowców i roślin
Systemy determinacji płci
dwa allele pojedynczego genu określają płeć np. drożdże
rozwój osobników męskich z niezapłodnionych jaj (np. pszczoły, mrówki, osy)
układy chromosomów płci
Układ XX-X0. Samice mają parę jednakowych chromosomów zwanych chromosomami X. Samce maja tylko jeden chromosom (np. koniki polne)
Układ XX-XY. Samice mają dwie kopie chromosomu X. Samce mają jeden chromosom X, a drugi Y (ssaki, niektóre owady). Samice są homogametyczne, to znaczy wytwarzają gamety zawierające wyłącznie chromosom X
Układ ZZ-ZW. Samice mają jeden chromosom Z a drugi W Samce mają dwie pary chromosomu ZZ (ptaki). Samice są heterogametyczne, to znaczy wytwarzają gamety z chromosomami Z i W (X i Y) z jednakową częstością.
Większość roślin okrytozalążkowych jest obupłciowa.
U roślin rozdzielnopłciowych (dwupiennych) kwiaty męskie wyrastają na innym osobniku, niż kwiaty żeńskie Przykłady: Melandrium, przestęp dwupienny, ogórek.
16. Chromosomy płciowe u Melandrium (0mówić szczegółowo)
Chromosomy płciowe u Melandrium
U tej rośliny osobniki żeńskie zawierają chromosomy XX, a męskie XY. Y znajdują się regiony hamujące rozwój cech żeńskich, a stymulujących męskich. Poza tym posiadają po 22 autosomy.
Chromosom Y u Melandrium często ulega defragmentacji.
Gdy chromosom Y utraci odcinek ze satelitą to rośliny są hermafrodytyczne (kwiaty dwupłciowe). Z tego wynika, iż w tym odcinku chromosomu są geny tłumiące rozwój żeńskich organów płciowych - rozwijają się tylko męskie
Utrata odcinka z telomerami powoduje powstawanie nieżywotnego pyłku (podziały redukcyjne następują jednak w sposób prawidłowy).
Dwupienność roślin jest wynikiem mutacji dwóch genów ściśle ze sobą sprzężonych (odkrył to Westergaard) : M i T .
M - gen lub grupa genów ściśle sprzężonych w obecności których tworzą się kwiaty obupłciowe
m - gen lub grupa genów ściśle sprzężonych w obecności których tworzą się kwiaty żeńskie
T - genu tłumiący żeńskość
t - jego allel recesywny.
wtedy:
MmTt - rośliny męskie
mmtt - rośliny żeńskie
MMtt - hermafrodytyczne (kwiaty obupłciowe)
17. Omów na dowolnym przykładzie dziedziczenie cech sprzężonych z płcią (daltonizm?, kolor oczu u owadow?)
Cechy sprzężone z płcią
Wszystkie cechy, które dziedziczą się wraz z chromosomami płci nazywamy cechami sprzężonymi z płcią, a sposób dziedziczenia określamy jako dziedziczeniem sprzężonym z płcią.
Allele recesywne położone na chromosomie X nie mogą się przejawiać fenotypowo u heterozygotycznych samic (u ssaków), lecz są widoczne u samców ponieważ mają tylko jeden chromosom
Przykład: daltonizm
18. Dziedziczenie cytoplazmatyczne (omów na przykładzie dziwaczka, nie wystarczy definicja, co wiesz na temat dziedziczenia plastydów)
Dziedziczenie cytoplazmatyczne
Zespół genów zlokalizowanych w chromosomach nazywamy genomem
Zespół czynników dziedzicznych znajdujących się w cytoplaźmie nazywamy plazmonem (plazmidem) a geny pozajądrowe - plazmogenami
Plazmogeny (plazmidy) są zlokalizowane w organoidach cytoplazmatycznych jak plastydy , mitochondria, centriole i inne
Dziedziczenie własności plastydów
Plastydy powstają z już istniejących struktur (rozmnażają się przez podział i są przekazywane z jednej komórki do drugiej), a niektóre cechy plastydów nie są przekazywane za pośrednictwem chromosomów.
Pstrolistność dziwaczków
Łatwo zauważyć, że rodzaj pyłku nie ma tu żadnego znaczenia. Czynnikiem decydującym jest gameta żeńska. Podane wyniki można tłumaczyć obecnością dwóch rodzajów plastydów (patrz tabelka poniżej)
19. Przedstawić na rysunku mechanizm cytoplazmatycznej męskiej sterylności (przykłady wykorzystania)
Geny cytoplazmatyczne mogą współdziałać z genami jądrowymi. Wykorzystuje się to zjawisko do otrzymywania roślin z cytoplazmatyczną sterylnością.
Cytoplazmatyczno-genetyczna męska niepłodność jest uwarunkowana współdziałaniem cytoplazmy (S) i recesywnych genów jądrowych (ms ms). Geny (ms ms) są odpowiedzialne za męską sterylność i nie funkcjonują w obecności cytoplazmy N. Rośliny z całkowicie męskoniepłodne posiadają więc genotyp (S ms ms). Do rozmnażania roślin męskoniepłodnych wykorzystuje się rośliny o genotypie (N ms ms). Są to tzw. linie dopełniające.
S ms ms x N ms ms ------ F1 S ms ms
męskosterylna płodna męskosterylna
Cytoplazmatyczną męską niepłodność wykorzystuje się w hodowli heterozyjnej.
Linię będącą zapylaczem w produkcji nasion heterozyjnych nazywamy linią restorerującą
20. Podstawowe definicje dotyczące genetyki populacji
Genetyka populacji zajmuje się badaniem jak rozkładają się geny pokolenia rodzicielskiego na osobniki pokolenia potomnego.
Populacja naturalna - wiele krzyżujących się zróżnicowanych genetycznie osobników, dzielących tą samą pulę genową.
Populacja lokalna obejmuje wszystkie osobniki, które mają udział w tej samej lokalnej puli genowej (np. dla owadów: drzewo, las).
Populacja mendlowska - zbiór osobników, które mają możliwość wzajemnego kojarzenia się.
Gatunek - wszystkie osobniki, które tworzą wspólnotę rozrodczą i których lokalne pule genowe pozostają w stałym kontakcie.
Częstość allelu określa się przez dodanie liczby kopii każdego allelu w populacji i przedstawienie jej jako częstości.
21. Omówić Prawo Hardy'ego-Weinberga
Prawo to przedstawia stan dynamicznej równowagi w populacji, na którą nie działają siły selekcyjne ani też żadne inne czynniki.
W dużej, losowo kojarzonej populacji, przy braku migracji, mutacji i selekcji, częstość genów i genotypów jest stała; populacja sama w sobie nie ma tendencji do zmiany swoich genetycznych właściwości z pokolenia na pokolenie.
Częstości genotypów z potomstwa, powstałego z losowego kojarzenia rodziców , są uwarunkowane jedynie częstością genów tych rodziców
populacja, znajdująca się w stanie równowagi Hardy- Weinberga, charakteryzuje się w każdym pokoleniu współzależnością między częstością genu i genotypów wyrażona równaniem:
p2 : 2pq : q2
gdzie:
p- częstość genotypu AA,
q - częstość genotypu aa,
pq -częstość genotypu Aa
częstość genotypów zgodna z prawem Hardy - Weinberga ustala się w populacji, po zastosowaniu losowego kojarzenia w jednym pokoleniu, niezależnie od genotypu rodziców
22. Wyprowadzenie wzoru na prawo Hardy'ego-Weinberga (tylko studia dzienne)
Częstość allelu M w populacji 1199/2000 = 0.5995,
częstość allelu N w populacji 801/2000 = 0.4005.W populacji 1000 białych Amerykanów 357 z nich było MM, 485 MN i 158 NN, 357 osón MM posiadało 714 alleli M a 485 osób MN miało kolejne 485 alleli M, tak więc częstość allelu M (-p) wynosi
Podobnie częstość N wynosi
Na zmianę genetycznych właściwości populacji mogą wpływać następujące czynniki:
wielkość populacji (podczas gdy duża populacja jest stosunkowo niewrażliwa na losowe odchylenia składu genetycznego, to w populacji małej za sprawą przypadku mogą zajść nieodwracalne zmiany)
dobór (selekcja) - oddziaływuje na żywotność czy płodność osobników lub gamet poszczególnych genotypów. Usuwa np. wadliwe allele, a równowaga częstości wystąpi wtedy, kiedy tempo powstawania nowych alleli jest równe tempu powstawania nowych mutacji
różnice w płodności i żywotności (wśród rodziców , osobniki o różnych genotypach mogą wykazywać różną płodność)
dryf genetyczny - czynnik przypadkowości, proces losowy zmiany częstości genu, szczególnie ostro występujący w małych populacjach (pojedynczy allel nie występuje dokładnie w 50%)
migracja - przepływ genów między subpopulacjami
mutacja - zmiana w sekwencji zasad DNA, mogąca wywołać zmiany fenotypowe u organizmów żywych
Przykład faworyzowania w doborze naturalnym heterozygot:
Allison (1954r) wykrył, że w Afryce ludzie ze skazą sierpowatą (Ss) cechują się znaczną odpornością na malarię. Chorzy o genotypie SS umierają na anemię, natomiast heterozygoty maja przewagę w spadku umieralności na malarię około 25 % nad homozygotami (ss)
23.Podaj podstawowe różnice miedzy cechami ilościowymi jakościowymi
Genetyka ilościowa:
- genetyka ilościowa zajmuje się dziedziczeniem różnic występujących między osobnikami w stopniu natężenia cech, a nie w ich jakości.
- każdy organ lub funkcja jakiegokolwiek gatunku wykazuje indywidualne różnice, np. różnice w wysokości wśród ludzi.
- osobniki tworzą szereg ciągły, wzrastający od najniższej wartości do najwyższej a nie dzielą się na wyraźnie różniące się od siebie typy.
- różnice jakościowe dzielą natomiast osobniki na wyraźnie różniące się od siebie typy bez lub występowaniem (rzadziej) typów pośrednich.
- istnienie różnic ilościowych i jakościowych w stosunku do badanego zjawiska stwarza konieczność odróżnienia genetyki ilościowej od jakościowej.
- genetyka cech ilościowych w odróżnieniu do genetyki cech jakościowych bada różnice spowodowane działaniem większej ilości genów.
Dziedzicznie uwarunkowane różnice ilościowe zależą od różnic występujących w wielu loci miedzy genami, których efekty nie mogą być indywidualnie określone. W wyniku tego zjawiska, mendlowskie stosunki liczbowe nie uwidaczniają się w różnicach ilościowych, a metody genetyki mendlowskiej nie nadają się do badania tych różnic. Podstawowym założeniem genetyki ilościowej jest twierdzenie, że dziedziczenie różnic ilościowych zależy od genów podlegającym tym samym prawom co dziedziczenie cech jakościowych. Genetyka ilościowa jest więc dalszym rozwinięciem genetyki Mendla i opiera się na prawach Mendla.
24. Współczynniki odziedziczalności
Odziedziczalność ogólna (współczynnik odziedziczalności) w szerokim sensie służy do określenia jaką część zmienności całkowitej stanowi zmienność genetyczna, a więc jest to stosunek zmienności genetycznej do całkowitej
Odziedziczalność w wąskim sensie jest to stosunek wariancji addytywnej do zmienności całkowitej
25.Ocena efektów działania genów
Efekty addytywne: wynikają ze sumowania się efektów wywoływanych przez wiele loci w stanie homozygotycznym.
Efekty dominowania - występują na skutek działania genów w stanie heterozygotycznym.
Niealleliczna interakcja (EPISTAZA) - wiąże się z:
współdziałaniem genów w stanie heterozygotycznym
współdziałaniem genów w stanie homozygotycznym
współdziałaniem genów w stanie homozygotycznym i heterozygotycznym
Znajomość efektów działania genów może przyczynić się do zwiększenia efektywności hodowli roślin.
Badanie efektów działania genów wymaga przeprowadzania specjalnie w tym celu zakładanych doświadczeń.
Liczba czynników efektywnych - jest to liczba genów lub grup genów ściśle ze sobą sprzężonych, którymi różnią się badane formy rodzicielskie
Współczynnik rozkładu genów u form rodzicielskich
dyspersja genów występuje w przypadku zgrupowania u każdej z form rodzicielskich równej liczby genów zwiększających jak i zmniejszajacych wartość danej cechy
asocjacja genów występuje w przypadku kumulacji (+) alleli u jednej z form rodzicielskich i (-) alleli u drugiej z form rodzicielskich
26.Opisz zjawisko heterozji, teorie tłumaczące powstawanie tego zjawiska
Heterozja - wyższość genotypów heterozygotycznych nad wyjściowymi homozygotami w odniesieniu do jednej lub więcej cech, stanowiący fenotypowy efekt współdziałania genów.
Teorie dotyczące występowania efektów heterozji
Istnieje kilka teorii tłumaczących powstawanie zjawiska heterozji. Należą one do jednej z dwóch kategorii:
Przyczyną heterozji jest stan heterozygotyczny loci. Zakłada się, że geny A i a wytwarzają różne związki, które albo przez połączenie lub produktu ich współdziałania powodują większą bujność niż każdy z pojedynczych związków produkowanych przez każdy z alleli w stanie homozygotycznym
Przyczyną heterozji jest współdziałanie genów dominujących. Heterozja jest skutkiem dominowania genów sprzyjającym bujności nad odpowiednimi genami recesywnymi przeciwdziałającymi bujności.
3. Wystąpienie efektu heterozji zależy od genów dopełniających (podobnie jak chromogen i oksydaza są koniecznymi do powstania antocjanu). Współdziałanie genomu z plazmonem
27. Zjawisko transgresji genetycznej
Depresja wsobna - spadek żywotności (wigoru) organizmów wywołany zapłodnieniem formy matecznej przez blisko spokrewnioną formę ojcowską. Zjawisko to występuje zarówno u roślin samopylnych jak i u zwierząt
Transgresja - pojawianie się wśród segregującego pokolenia osobników o cechach przekraczających zakres zmienności form rodzicielskich. Efekt transgresji związany jest ze stanem homozygotycznym loci.
1
15
(synteza DNA)
Faza
G2
Faza S
Komórki
bez dalszych podziałów
Cykl komórkowy
Faza
G1
M
(mitoza)
Wyszukiwarka
Podobne podstrony:
Ochrona środ pytania na koło sciaga
pytania na kolo z inzynierii genetycznej
pytania na kolo sciaga
Pedagogka res. Kijak (pytania na koło) - ściąga, Resocjalizacja, Pedagogika resocjalizacyjna
Materiały ściąga, Studia budownictwo pierwszy rok, Materiały budowlaneII, Pytania na koło 1
PYTANIA NA KOLO ściąga
pytania na kolo sciaga2
pytania na koło 8 14
Etyka pytania na kolo
Pytania na I koło
pytania na I kolo, Prywatne, Anatomia od Olgi
pytania na koło z rysunku wykład
Pytania na kolo
Ochrona środ pytania na koło
Biofizyka wszystkie pytania na kolo
pytania na kolo z ZZL
Opracowane pytania na koło 3 7 11 15, Budownictwo UTP, III rok, DUL stare roczniki, GEODEZJA, geodez
pytania na kolo (2)
więcej podobnych podstron