
Choroby
chromosomowe
człowieka I
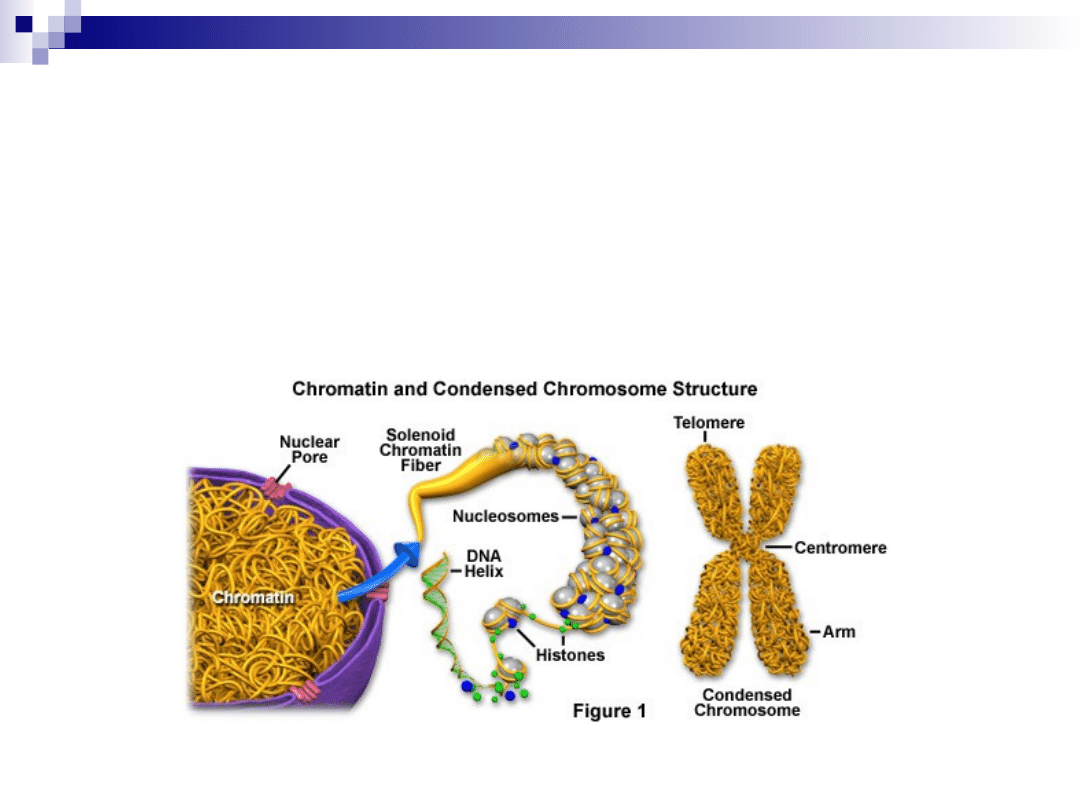
chromatyna
DNA + histony + białka niehistonowe

chromatyna
euchromatyna
mało skondensowana, zawiera aktywne
geny
lokalizacja: w całym jądrze
heterochromatyna
silnie skondensowana (kondensacja
uniemożliwia transkrypcję)
lokalizacja: pod otoczką jądrową i w
okolicach jąderka
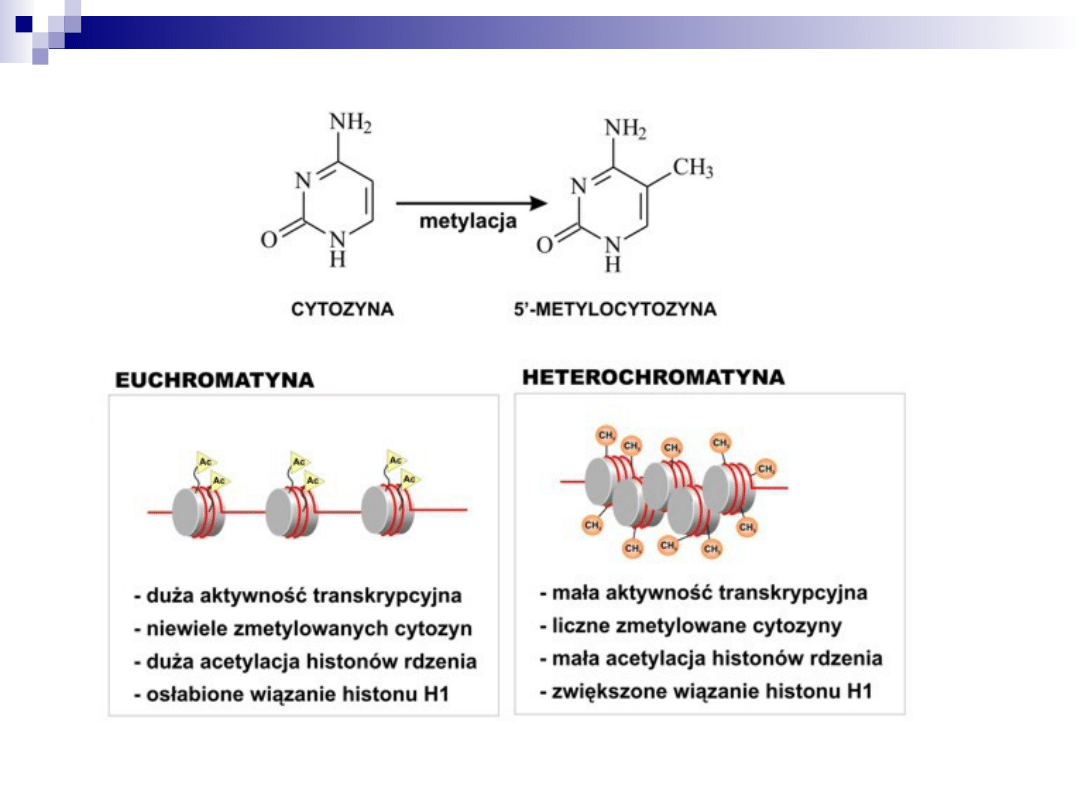

Chromosom
– forma upakowania materiału
genetycznego wewnątrz komórki
– struktura znajdująca się w jądrze,
zbudowana z liniowej cząsteczki
DNA i białek, wzdłuż której
ułożone są geny, widoczna pod
mikroskopem podczas podziałów
komórkowych
gr. Chromos – kolorowy,
Soma –ciało
nazwa pochodzi od zdolności
wybarwiania się

Chromosomy:
występują we wszystkich komórkach z jądrem
zawierają liniowo ułożone geny
liczba chromosomów jest cechą gatunkową w
komórkach somatycznych (diploidalnych) – 46
chromosomów (23 pary – jeden komplet
otrzymujemy od ojca, drugi od matki) w
komórkach rozrodczych - gametach
(haploidalny)
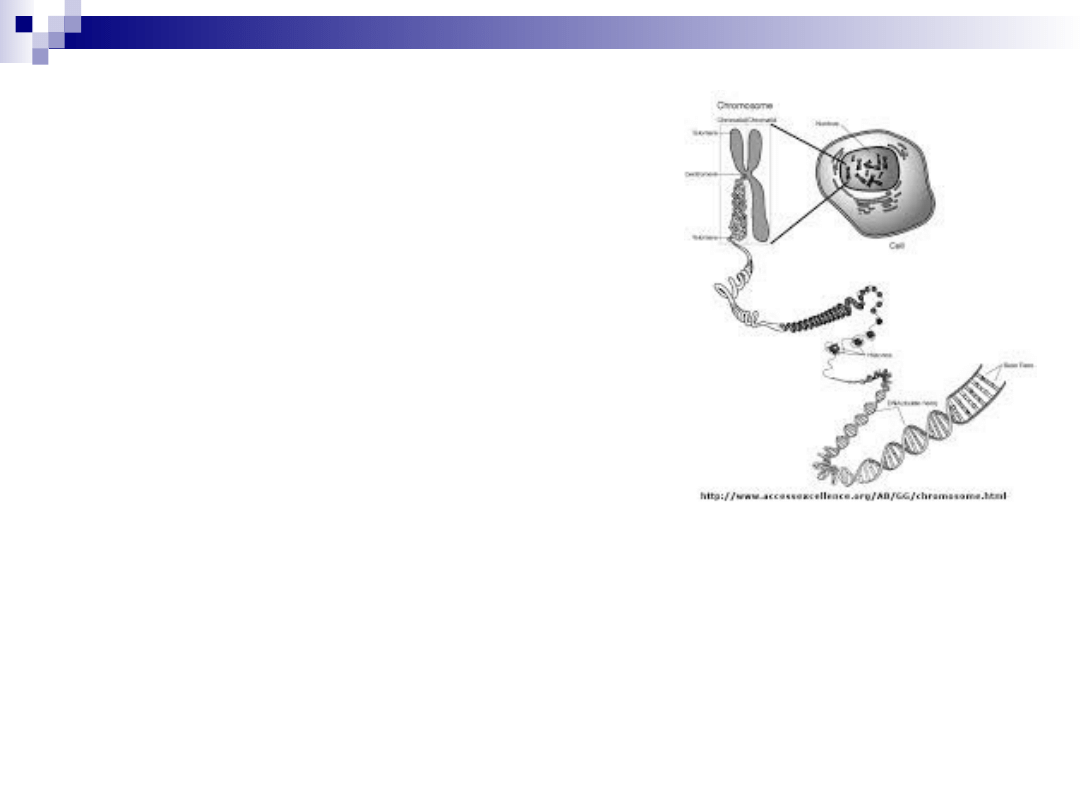
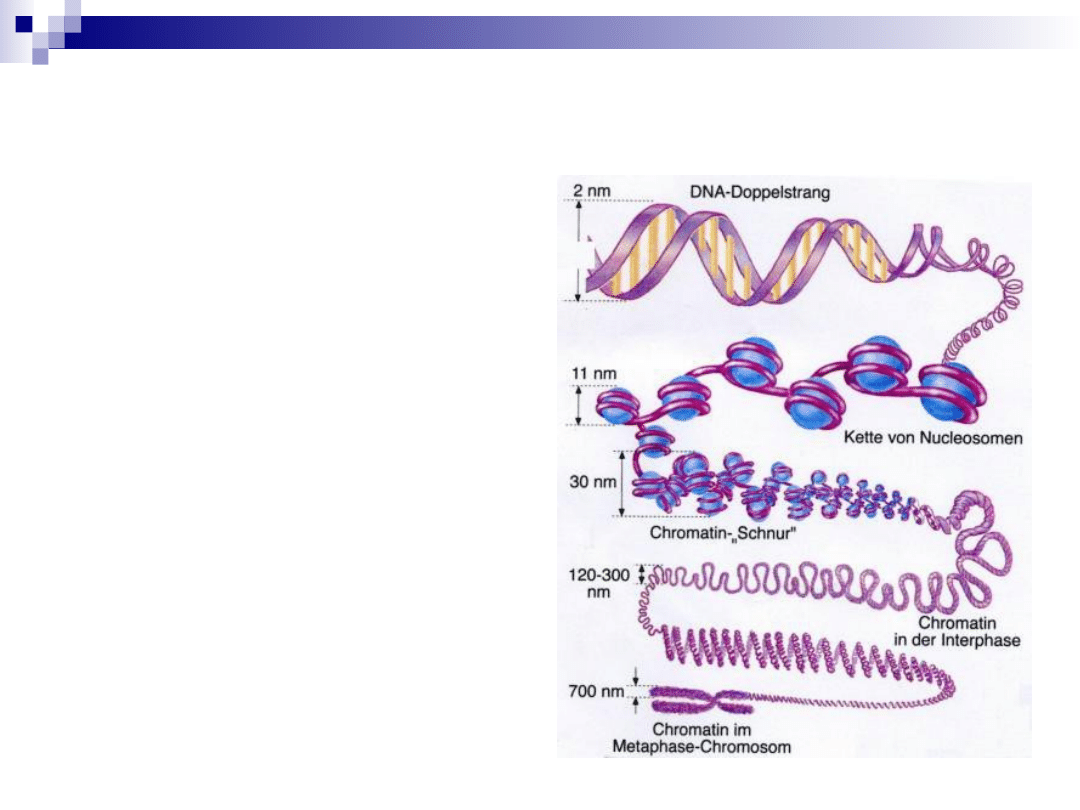
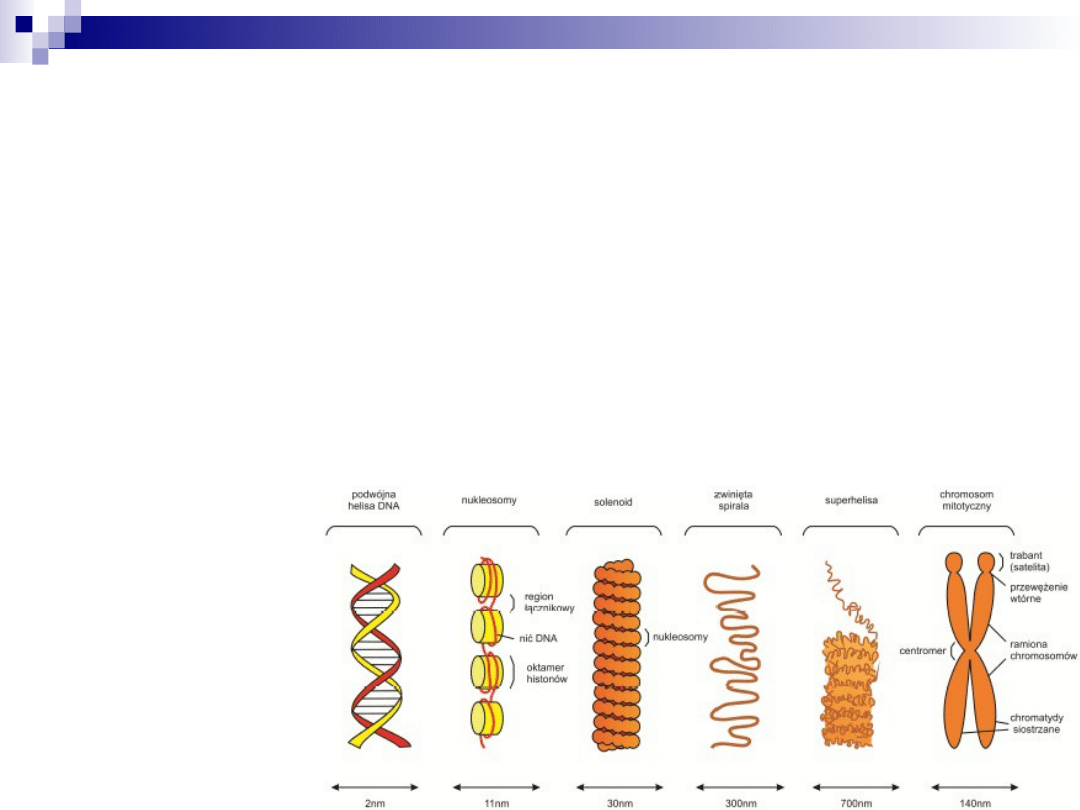
upakowanie materiału
genetycznego
1.
podwójna helisa DNA
2.
nukleosom
3.
fibryla chromatynowa
4.
solenoid
5.
chromatyda
6.
2 chromatydy = chromosom metafazowy
chromosom metafazowy- NAJWYŻSZY stopień
skondensowania chromatyny
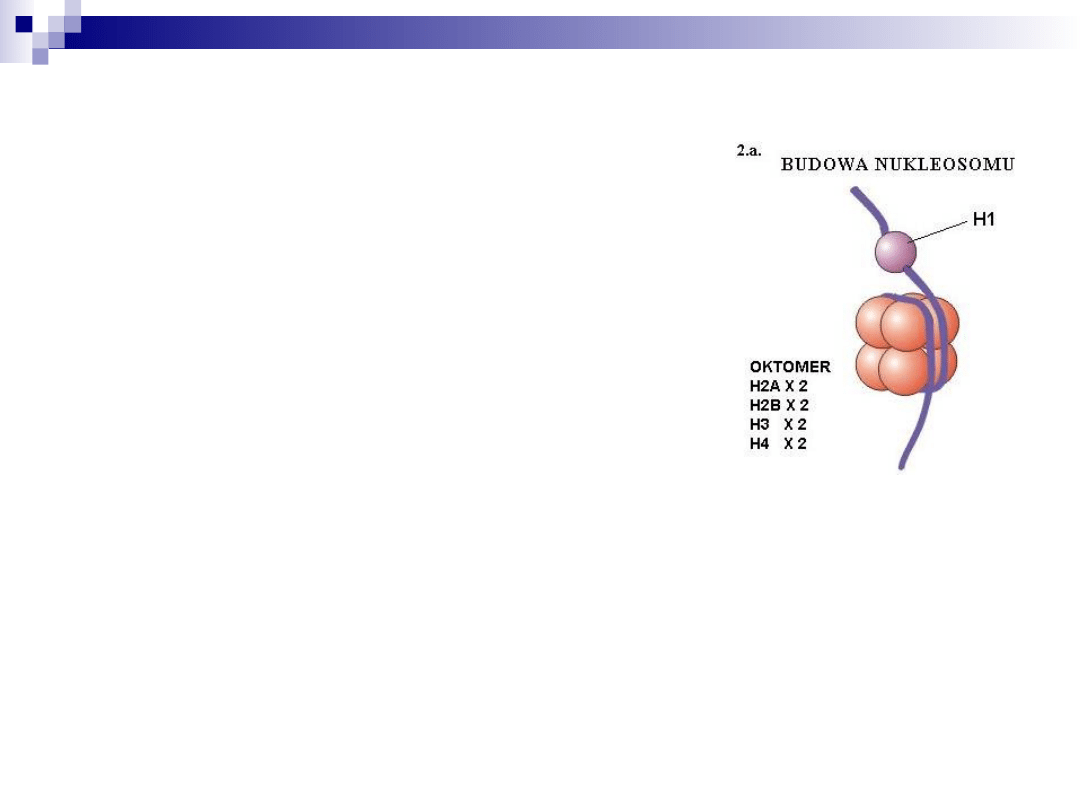
nukleosom
Podwójna helisa DNA + 8
cząsteczek histonów
(H2A, H2B, H3 i H4) x2
DNA o długości 146 par
zasad owija się wokół 8
cząsteczek histonów
tworzących rdzeń (1,75
zwoju). Na zewnątrz
znajduje się histon H1,
który łączy się z
łącznikowym DNA (60 par
zasad), oddziałuje z H2A i
dodatkowo chroni DNA.
fibryna chromatynowa
„sznur korali”
sąsiadujące
nukleosomy +
łącznikowy DNA
w mikroskopie
elektronowym
wyglądają jak
sznur koralików o
średnicy 10 nm
solenoid
dochodzi do zwijania we włókno chromatynowe o szerokości 30 nm
(skondensowane 40-krotnie)
…w wyniku dalszego upakowania dochodzi do 8000-krotnego
upakowania materiału genetycznego w chromosomie metafazowym
DNA w komórce to około 2 metrów!!!
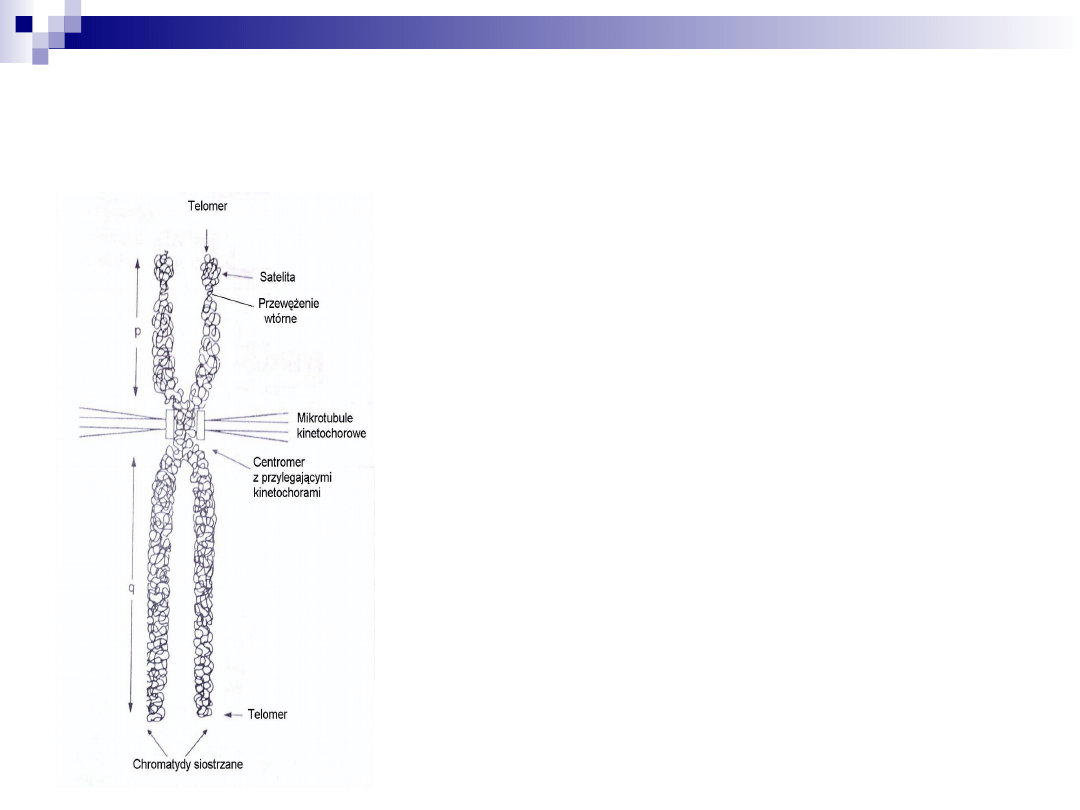
chromosom metafazowy
skład: 2 symetryczne chromatydy
(siostrzane) łączące się ze sobą
w obrębie centromeru
wielkość: 10 μm - 20 μm
rdzeń: z kwaśnych białek, do
których przyczepione są włókna
chromatyny bogate w sekwencje
AT
od rdzenia odchodzą promieniście:
pętle Laemliego, tworzące
chromatydę, które przyłączone są
przez multimery białkowe SAF-A
zakończone telomerami

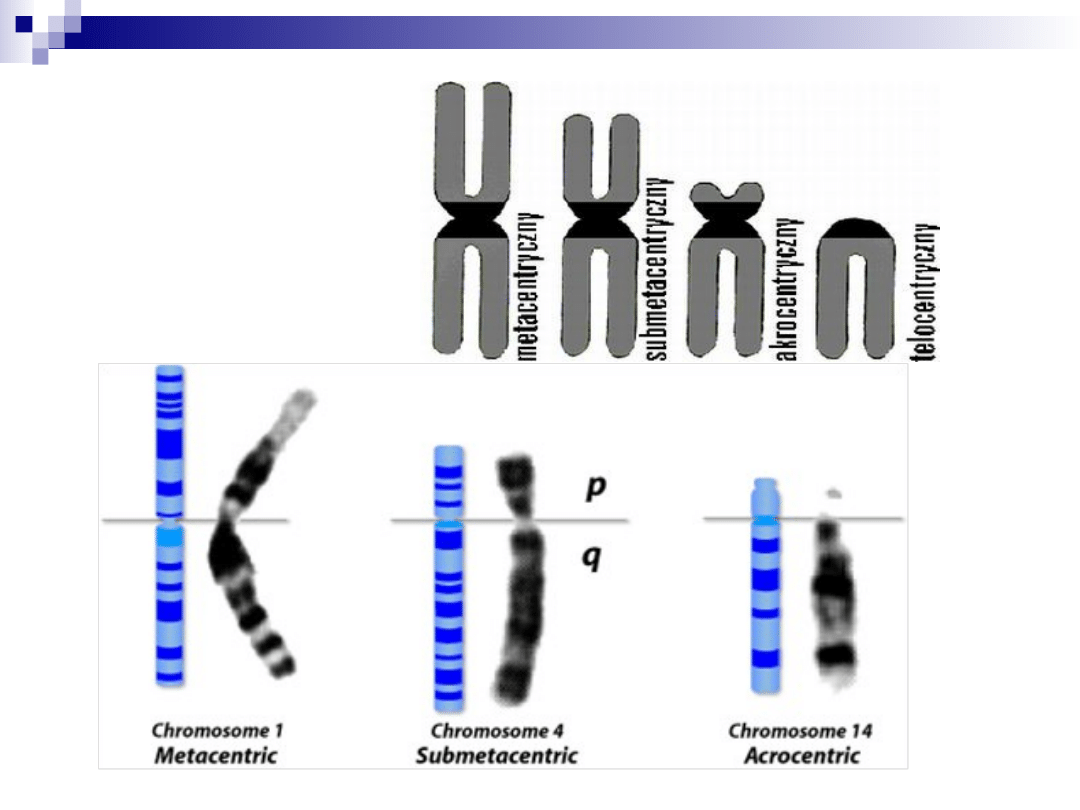
chromosomy
ze względu na położenie centromeru
(przewężenie pierwotne) chromosomy
dzielimy na:
1.
metacentryczny
2.
submetacentryczny
3.
akrocentryczny
4.
telocentryczny (brak u człowieka)

kariotyp
specyficzna liczba oraz charakterystyczny
układ chromosomów
44 autosomy (pary 1-22) + 2 chromosomy
płci, heterochromosomy (XX kobieta, XY
mężczyzna)

każdy prawidłowy chromosom ma jeden
centromer, jego położenie jest stałe dla
danego typu chromosomu i jest to miejsce
przyczepu włókien wrzeciona
podziałowego
każdy chromosom ma ramię krótkie p (z
franc. petit) oraz ramię długie q

kariogram (idiogram)
zestaw chromosomów przedstawionych
graficznie (wg zasad opracowanych na
konferencji cytogenetycznej w Paryżu
1971r.)
46 chromosomów ułożonych malejąco jako
22 homologiczne pary +chromosomy
płciowe

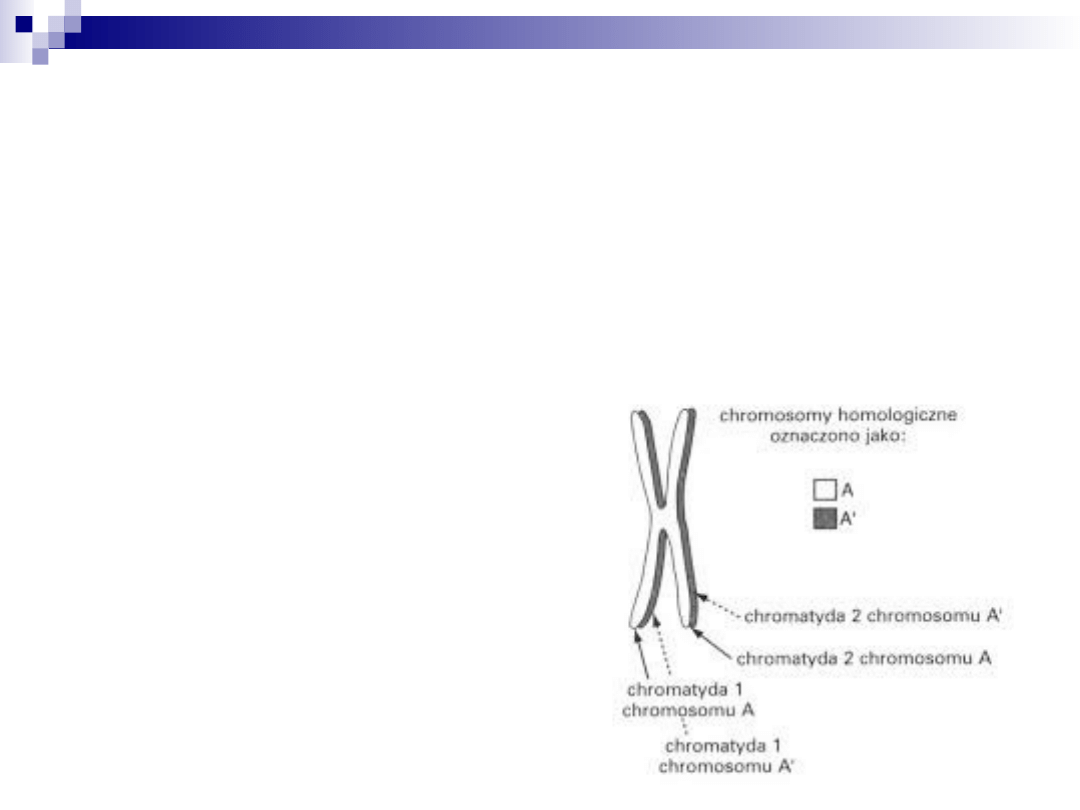
chromosomy homologiczne
zawierają te same geny, lecz różnią się
pochodzeniem (od ojca i od matki)

Aberracje
chromosomowe

aberracje chromosomowe
1.
liczbowe
2.
strukturalne

Aberracje chromosomalne liczbowe
najczęściej powstają w wyniku
nieprawidłowych rozdziałów
chromosomów w czasie podziału
poliploidie (euploidie)
aneuploidie

aneuploidie
zwiększenie/zmniejszenie diploidalnej
liczby chromosomów o pojedyncze
chromosomy
na skutek nondysjunkcji w mitozie/mejozie
czy utraty chromosomu w anafazie

aneuploidie
hipoaneuploidy
2n-1 monosomik
2n-1-1 podwójny monosomik
2n-2 nullisomik
hiperaneploidy
2n+1 trisomik
2n+1+1 podwójny trisomik
2n+2 tetrasomik
(trisomiki są bardziej żywotne niż monosomiki, w populacji
brak nullisomików i tetrasomików-letalne)

aneuploidie -przykłady
zespół Patau 47,XX+13 / 47,XY+13
zespół Downa 47,XX+21 / 47,XX+21
zespół Turnera 45,X
zespół Klinefeltera 47,XXX

euploidia
zwielokrotnienie całego haploidalnego
(1n=23) zestawu chromosomów
3n (69, XXX/ 69, XXY),
4n (92, XXXX/ 92 XXYY)
5n….
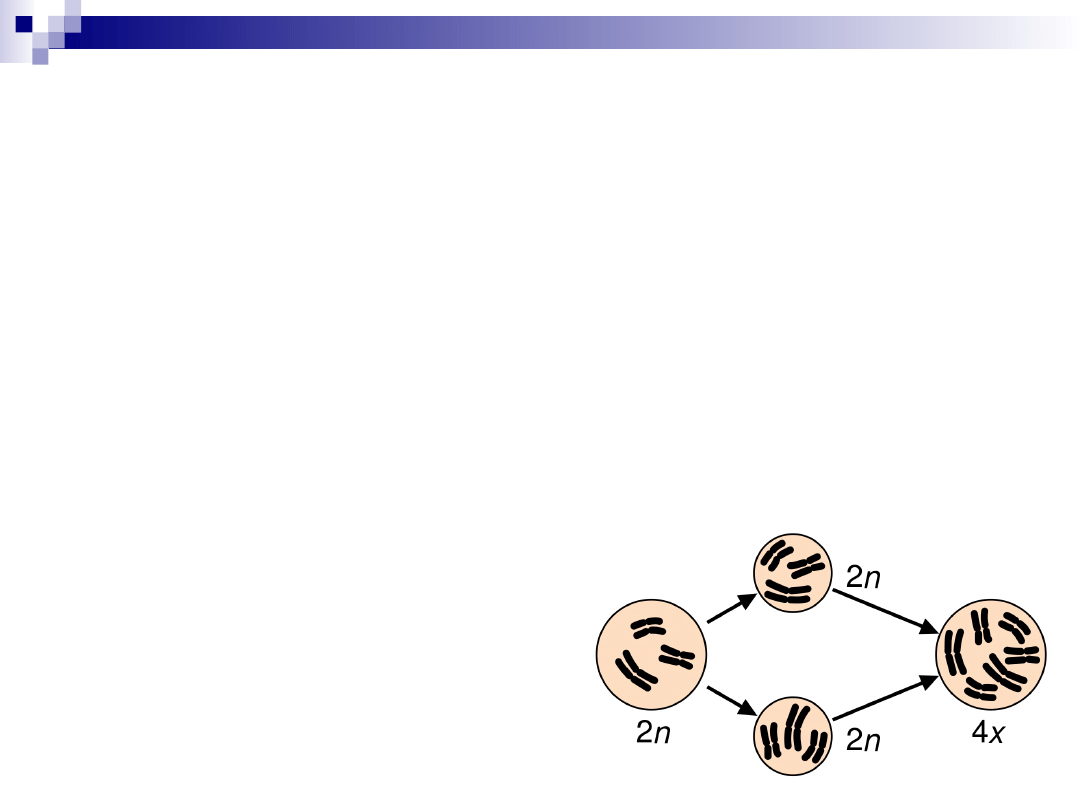
euploidia-przyczyny
1.
brak rozdziału chromosomów w mitozie –
powstaje jądro o podwójnej liczbie
chromosomów
2.
w mejozie powstają gamety o niezredukowanej
diploidalnej liczbie chromosomów
gameta 2n +gameta 2n= 4n
gameta 1n +gameta 2n= 3n

euploidia
autopoliploidy
zwielokrotniony ten sam zestaw chromosomów (ch.homologiczne)
u człowieka letalne
wykorzystywane w rolnictwie
allopoliploidy (amfiploidy)
dwa lub więcej zestawów niehomologicznych chromosomów (różne gatunki)
brak u człowieka, w rolnictwie pszen-żyto
przykład muł
bezpłodny, ponieważ niemożność koniugacji w I profazie mejozy

Aberracje chromosomalne
strukturalne
translokacje wzajemne i robertsonowskie
inwersje
insercje
delecje interstycjalne i dystalne
duplikacje i mikroduplikacje
izochromosomy
chromosomy dwucentromerowe
chromosomy pierścieniowe
chromosomy markerowe
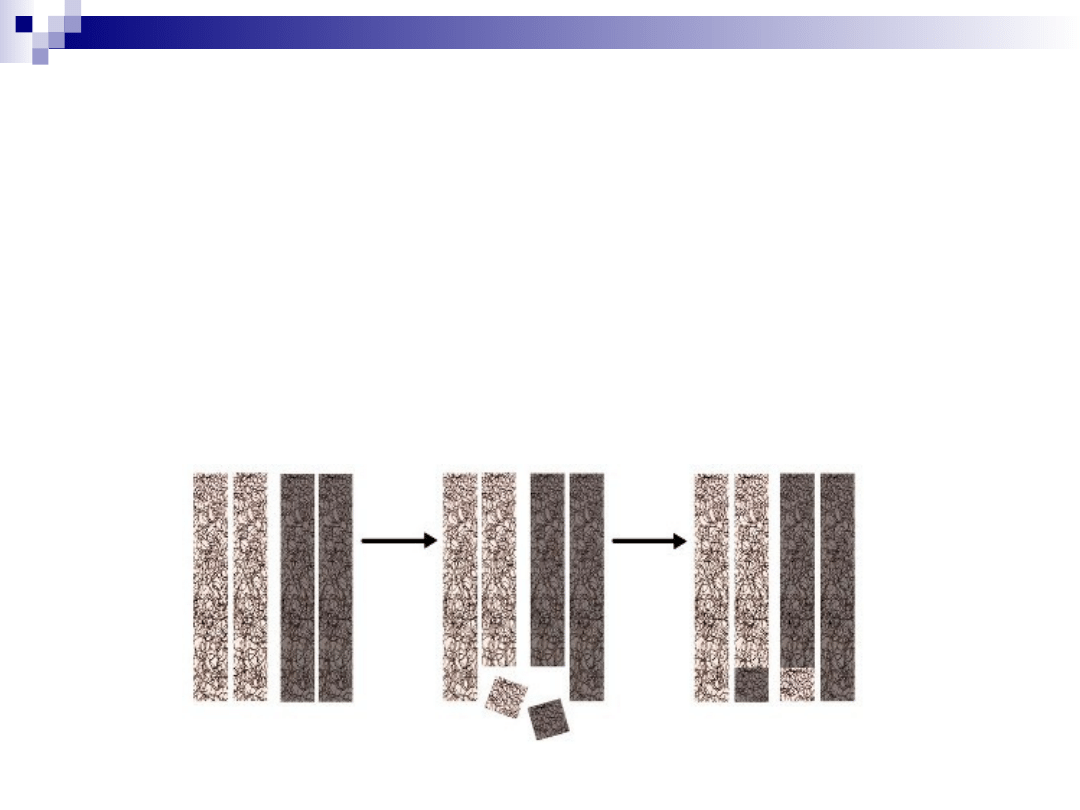
translokacja
przemieszczenie fragmentu chromosomu
w obrębie tego samego/innego
chromosomu

translokacja wzajemna
wzajemna wymiana odcinków między
chromosomami niehomologicznymi
całkowita liczba chromosomów:
niezmieniona
budowa: zmieniona

translokacja robertsonowska
(typu fuzji)
dochodzi do połączenia się całym lub prawie
całych ramion długich różnych chromosomów
miejscem połączenia jest centromer (połączenia
centryczne)
utrata nieistotnej pod względem funkcji części
materiału genetycznego
tylko chromosomy akrocentryczne

translokacja robertsonowska
zrównoważona
niezrównoważona

translokacja robertsonowska
zrównoważona
nie zmienia się ilość materiału
genetycznego
zmienia się lokalizacja materiału w
genomie
45 chromosomów metafazowych
brak objawów fenotypowych
możliwość przekazania potomstwu

translokacja robertsonowska
niezrównoważona
materiał genetyczny zostaje powiększony
o dodatkową kopię translokowanego
chromosomu
46 chromosomów metafazowych
zawsze objawy fenotypowe
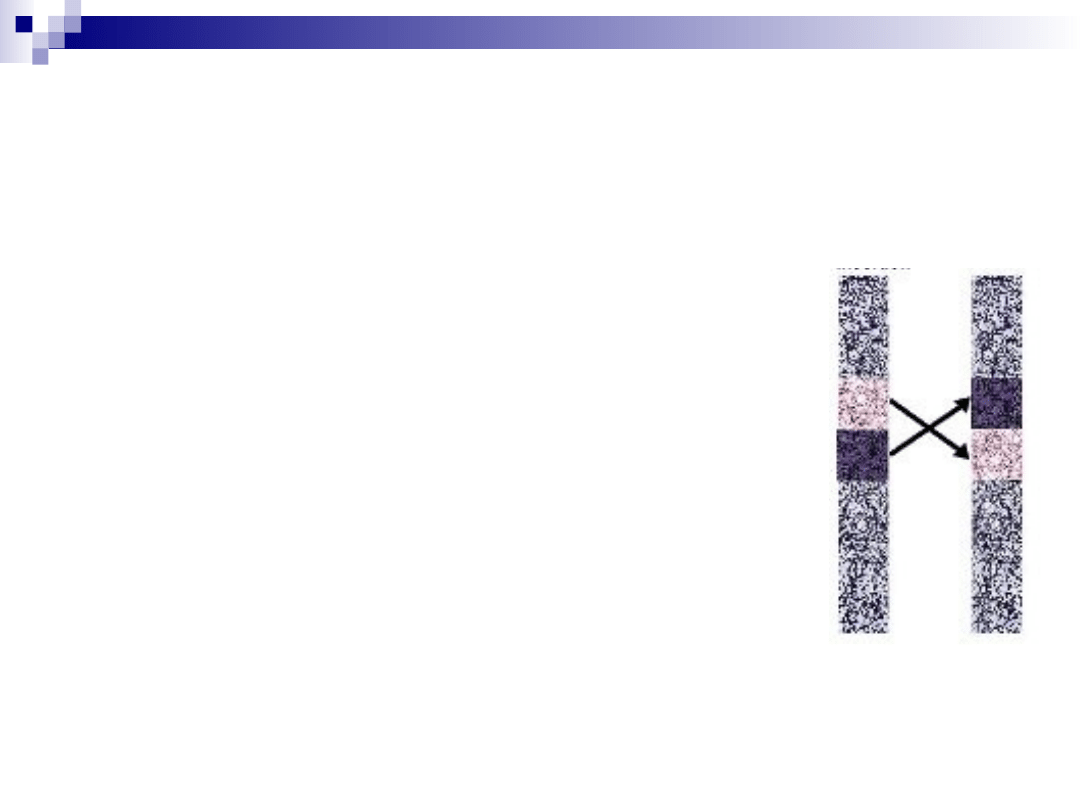
inwersja
chromosom ulega złamaniu w 2
miejscach, a fragment pomiędzy
złamaniami ulega odwróceniu o 180
inwersja pericentryczna
(eucentryczna) =obejmuje odcinek
chromosomu z centromerem
inwersja paracentryczna
(acentryczna) =nie obejmuje
centromeru

insercja
wstawienie materiału
genetycznego w nietypowe
miejsce w tym samym lub
innym chromosomie

delecja (deficjencja)
utrata odcinka chromosomu
delecja terminalna
utrata części dystalnej chromosomu
delecja interstycjalna
utrata fragmentu środkowego
kruche miejsce genomu: odcinki chromosomów o
szczególnej łamliwości

mikrodelecja
delecje o wielkości bliskiej granicy
rozdzielczości mikroskopii świetlnej
w ich identyfikacji pomocne są metody
techniki molekularnej

duplikacja
podwojenie tych samych
odcinków chromosomu
bezpośrednie powtórzenia
(proste powtórzenia
tandemowe)
odwrócone względem siebie
powtórzenia

mikroduplikacje
maleńskie duplikacje na poziomie
molekularnym (powtórzenia)
mogą mieć istotna rolę w powstawaniu
różnorodności genów w trakcie ewolucji

izochromosom
powstaje w wyniku nieprawidłowego,
poprzecznego podziału centromeru
chromosomu metafazowego
LUB poprzez złamanie izochromatydowe
i fuzję tuż nad centromerem
(dicentryczny)

izochromosom
skład: tylko z połączonych ramion krótkich LUB
tylko z połączonych ramion długich
ubytek genów (ramiona utracone)
podwojenie liczby genów (ramiona budujące
izochromosom)
dotyczy zarówno autosomów jak i chromosomu
X
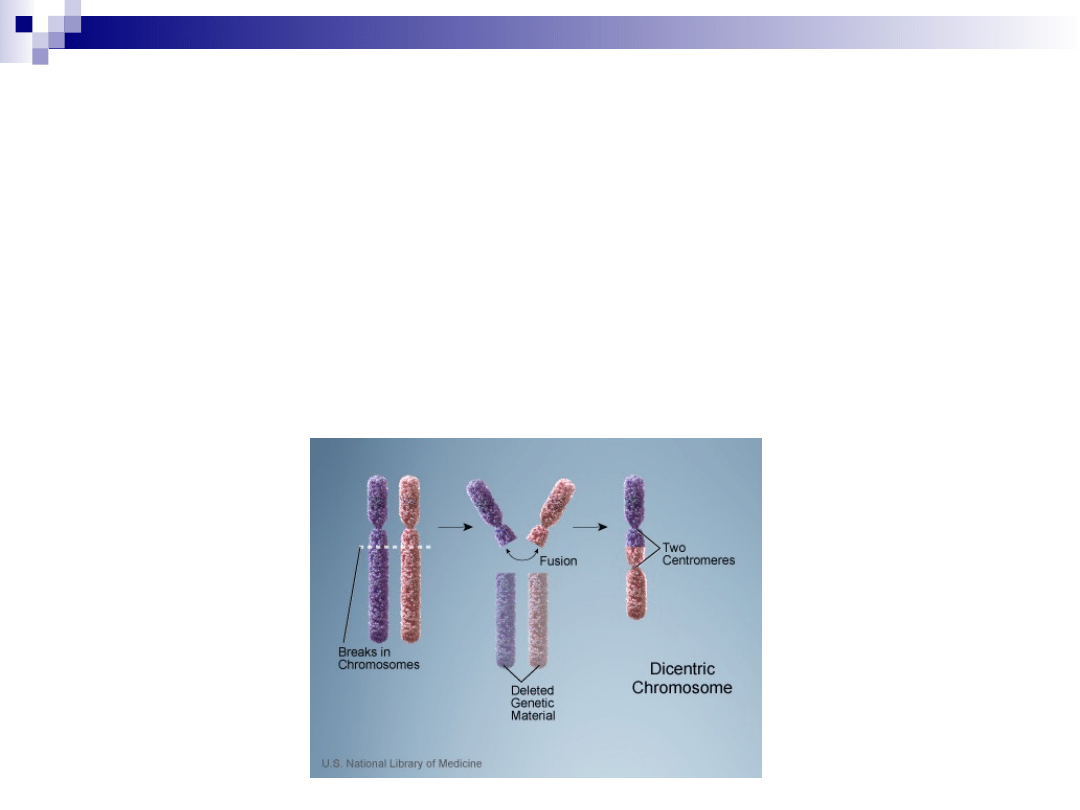
chromosomy dicentryczne
dwa centromery
jeden centromer jest nieczynny, dlatego
możliwa jest prawidłowa segregacja
chromosomów podczas podziału komórki

chromosom pierścieniowy
powstaje w wyniku złamań w
obydwu ramionach chromosomu
części terminalne ulegają utracie,
proksymalne lepkie końce łączą
się, tworząc pierścień
jeśli posiada centromer = może
uczestniczyć w podziałach
komórki
w kolejnych podziałach
komórkowych zwiększa
dwukrotnie swoją wielkość

chromosom markerowy
małe, dodatkowe, zazwyczaj metacentryczne
fragmenty chromosomów
mogą występować rodzinnie i być wynikiem
translokacji robertsonowskiej
brak skutków jeśli zawierają tylko sekwencje
powtórzone lub geny dla rRNA, lecz czasem
ulegają transkrypcji, stąd nieprawdiłowy fenotyp
zespół kociego oka

przyczyny powstawania aberracji
chromosomowych
przerwanie ciągłości chromatydy lub
chromosomu (obydwu chromatyd)

Dziękuję za uwagę!
Anna Puzio
III lek, gr. 45
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
- Slide 38
- Slide 39
- Slide 40
- Slide 41
- Slide 42
- Slide 43
- Slide 44
- Slide 45
- Slide 46
- Slide 47
Wyszukiwarka
Podobne podstrony:
2 Ch chromosomowe cz 1 Stec
3 Ch chromosomowe cz 2 Rabiej
5 Ch jednogenowe cz 2 Ch wieloczynnikowe Skrzypek
4 Ch jednogenowe cz 1 Ładosz
Prelekcja 10 - cz 2 - Mutacje chromosomowe człowieka, Genetyka
Ch zw krŕg cz 2 do wydruku
ch stawow o podl imm cz II
ch stawow o podl imm psow i kotow cz. II
Prelekcja 10 - cz 2 - Mutacje chromosomowe człowieka, Lekarski I rok ŚUM, biologia
Geometria w praktyce, cz 2 ?ch czterospadowy i kopertowy
Ch zw krŕg cz 1 do wydruku
więcej podobnych podstron