
Układ wydzielania wewnętrznego (
wer.robocza
)
Hormon
– substancja regulacyjna wytwarzana przez wyspecjalizowane narządy i komórki gruczołowe, wydzielana
przez nie do krwi, płynu zewnątrzkomórkowego, chłonki i transportowana do komórek docelowych, które
posiadają na swojej powierzchni, w cytoplazmie lub w jądrze swoiste receptory wiążące dany hormon. Hormony są
chemicznymi przekaźnikami ustroju, a do ich zadań należy:
●
regulowanie syntezy i wydzielania innych hormonów oraz feromonów
●
regulowanie syntezy i wydzielanie enzymów, kwasu solnego, żółci
●
wpływanie na procesy metaboliczne komórek
●
kontrola procesów reprodukcji
●
kontrola równowagi jonowej organizmu
Gruczołami wydzielania wewnętrznego są:
●
przysadka mózgowa
●
tarczyca
●
przytarczyce
●
nadnercza
●
wyspy Langerhansa trzustki
●
gonady
Czynność wydzielniczą wykazuja ponadto:
●
podwzgórze
●
komórki układu APUD występujące m.in. w obrębie błony śluzowej układu pokarmowego
●
nerka
●
tkanka tłuszczowa
●
skóra
Hormony ze
względu na ich budowę chemiczną
dzielimy na trzy główne grupy:
a)
hormony peptydowe i białkowe
●
ich synteza rozpoczyna się od transkrypcji genu zawartego w DNA komórkowym. Powstały mRNA koduje
powstawanie w rybosomach siateczki śródplazmatycznej szorstkiej komórki substancji peptydowej
lub białkowej stanowiącej następnie hormon. Cząsteczka jest przekazywana do aparatu Golgiego gdzie
zachodzi jego ostateczna modyfikacja, stamtąd gotowe hormony wędrują do pęcherzyków komórki gdzie
będą magazynowane do czasu pojawienia się sygnału indukującego ich uwolnienia poza komórkę.
b)
hormony steroidowe
●
wspólnym prekursorem wszystkich tych hormonów jest cholesterol, który zostaje w komórce
syntetyzującej hormon przeniesiony do mitochondrium gdzie ulega przemianie do pregnenolonu.
Związek ten podlega modyfikacjom prowadzonym przez siateczką śródplazmatyczną gładką, a efektem
ich działania jest powstawanie różnych hormonów z tych samych prekursorów. Ważne jest iż hormony
steroidowe nie gromadzą się w komórce, ale są z niej tworzone w miarę bieżących potrzeb organizmu.
1

Układ wydzielania wewnętrznego (
wer.robocza
)
c)
hormony pochodne aminokwasów głównie tyrozyny
●
w zależności od rodzaju hormonu są one wydzielane i magazynowane przez:
✓ komórki chromochłonne rdzenia nadnerczy – aminy katecholowe: adrenalina, noradrenalina i
dopamina
✓ komórki pęcherzykowate gruczoły tarczowego – tyroksyna i trójjodotyronina
Hormony mogą wykazywać działanie:
●
autokrynne – wówczas hormon oddziałuje na komórkę, przez którą został wydzielony
●
parakrynne – hormon oddziałuje na komórki sąsiednie do komórki wydzielającej
●
endokrynne – hormon działana komórki odległe, do których przenoszony jest przez krew
Transport hormonów we krwi
Hormony są transportowane przez
krew
w stanie wolnym lub związane z białkami osocza, ważne jest ponadto, że
biologiczne efekty są w stanie wywrzeć tylko hormony w stanie wolnym
.
●
w stanie wolnym transportowane są: hormony peptydowe i białkowe, wyjątkami są:
GH, IGF-I, wazopresyna,
oksytocyna, hormony tarczycy
●
związane z białkami osocza są transportowane: hormony steroidowe oraz hormony tarczycy
Mechanizmy działania hormonów na komórki docelowe
Błona komórkowa
jest przepuszczalna dla cząsteczek lipidowych (hydrofobowych) a nieprzepuszczalna m.in. dla
białek i peptydów (hydrofilnych). W związku z tym różne hormony zaliczane do dwóch w/w grup będą miały inaczej
rozmieszczone swoiste receptory komórkowe, przez które będą wywoływały swoje działanie na komórki docelowe.
a)
hormony białkowe, polipeptydowe oraz pochodne tyrozyny
(poza hormonami tarczycy) – nie są w stanie
przenikać przez błonę komórkową, ich receptory znajdują się na powierzchni zewnętrznej tej błony. Receptor
wiążący dany hormon będzie wywoływał kaskadę reakcji doprowadzających do określonyc zmian w obrębie
komórki, wyróżniamy dwie grupy receptorów błonowych:
●
receptory związane z białkiem G – białka G są białkami trójdomenowymi, składają się z podjednostek
α,β i γ związanych w spoczynku przez GDP. Aktywacja jakiej doznaje to białko po przyłączeniu swoistego
hormonu do receptora powoduje zastąpienie GDP przez GTP i uwolnienie podjednostki α związanej tym
substancją (de facto jest to kompleks podj. α – GTP). W tym miejscu działanie kompleksu α – GTP staje się
dwukierunkowe zależne od rodzaju przyłączonej substancji:
✓ dochodzi do aktywacji enzymu cyklazy adenylanowej, która prowadzi syntezę wtórnego przekaźnika
komórkowego – cyklicznego-3’5’-adenozynomonofosforanu (cAMP)
✓ aktywowana jest inna domena białkowa – fosfolipaza C, która hydrolizuje błonowy
fofatydyloinozytolo-4,5-bisfosfoforan (PIP
2
), rozpad tego związku powoduje powstanie
diacyloglicerolu (DAG) i inozytolotrisfosforanu (IP
3
) będących podobnie jak cAMP wtórnymi
przekaźnikami komórkowymi.
Wydzielone w ten sposób wtórne przekaźniki: cAMP, IP
3
oraz DAG uruchamiają kaskadę
wewnątrzkomórkowych reakcji, które prowadzą do regulacji obecnych w komórce szlaków metabolicznych.
W ten sposób działają następujące receptory: TRH, GnRH, TSH, LH, FSH, HCG, ACTH, PTH, glukagonu
●
receptory związane z pojedynczą domeną transbłonową (receptory katalityczne) – przyłączenie do tego
typu receptorów hormonów nadaje mu aktywność kinazy białkowej, która powoduje fosforylację samego
2
Układ wydzielania wewnętrznego (
wer.robocza
)
receptora oraz niektórych białek komórkowych. Fosforylacja białka, szczególnie enzymatycznego, w
zasadniczy sposób zmienia jego funkcj biologiczne.
W ten sposób działają między innymi: receptory insuliny, EGF, insulinopodobnego czynnika wzrostu I i II
(IGF-I, IGF-II), GH, PRL, niektóre cytokiny
b)
hormony steroidowe oraz hormony tarczycy
– dzięki swojemu hydrofobowemu charakterowi są w stanie
przejść przez błonę komórkową, ich swoiste receptory znajdują się wewnątrz komórki. Najczęściej są to
receptory jądrowe, rzadziej cytosolowe
●
receptory jądrowe składają się z dwóch domen: jednej wiążącej DNA i drugiej wiążącej hormon. Domena
pierwsza jest aktywna dopiero po przyłączeniu do domeny drugiej hormonu
●
receptory te wiążąc DNA pobudzają ekspresję ściśle określonych genów, które kodują określone białka
powodujące daną odpowiedź metaboliczną komórki
●
efekt działania hormonu steroidowego nie ustaje natychmiast po odłączeniu go od swoistego dla niego
receptora oraz nie pojawia się natychmiast po zadziałaniu hormonu na receptor. Zależy to od czasu
półtrwania mRNA i białka będącego produktem genu pobudzanego przez hormon
Rytmy wydzielania hormonów
Większość hormonów nie jest wydzielana w sposób ciągły. Wiele z nich m.in. hormony podwzgórza, przysadki
czy insulina wydzielane są
pulsacyjnie
– a więc co pewien czas, różny dla różnych hormonów następuje wzrost
ich wydzielania. Wydzielanie hormonów jest cechą zależną od wieku, dotyczy to np. GH – hormonu wzrostu, lub
hormonów wydzielanych przez gonady.
Wydzielanie wielu hormonów ma ponadto
charakter okołodobowy
, dobitnym przykładem tego typy rytmu
wydzielania jest szyszynkowa melatonina, której wydzielanie jest hamowane pod wpływem światła.
Dłuższe rytmy wydzielania to tzw.
rytmy infradobowe
, których przykładem jest 28-dniowy cykl wydzielania
hormonów płciowych kobiet.
Mechanizmy regulacji wydzielania hormonów
Regulacja wydzielania hormonów a także ich aktywności w komókach docelowych odbywa się na trzech drogach, z
których pierwsza jest wysoce dominująca w organizmie ludzkim
●
Regulacja przez sprzężenie zwrotne ujemne
●
Regulacja przez sprzężenie zwrotne dodatnie
●
Regulacja przez przekształcanie hormonu nieaktywnego do formy aktywnej w komórkach
1) Regulacja przez sprzężenie zwrotne ujemne
Najczęściej jest ona omawiany na przykładzie podwzgórza, przysadki i gruczołu dokrewnego
●
hormon pobudzający (liberyna) podwzgórza
stymuluje przysadkę
do uwalniania określonego hormonu
tropowego
●
hormon tropowy przysadki działa na gruczoł docelowy powodując
wzrost wydzielania hormonu
przez ten
określony gruczoł
3

Układ wydzielania wewnętrznego (
wer.robocza
)
✓ jednocześnie
hormon tropowy przysadki hamuje uwalnianie przez podwzgórze liberyn
pobudzających
uwalnianie danego hormonu tropowego przez przysadkę (pętla krótka sprzężenia zwrotnego)
●
wydzielany hormon gruczołowy działa
hamująco na wydzielanie hormonu tropowego przez przysadkę
(pętla bezpośrednia)
✓ ponadto
hormon gruczołowy hamuje wydzielanie liberyny przez podwzgórze
(pętla pośrednia/długa
pętla sprzężenia zwrotnego)
2) Regulacja przez sprzężenie zwrotne dodatnie
Wzrost wydzielania hormonu tropowego przysadki powoduje zwiększone wydzielanie hormonu przez gruczoł
docelowy.
Hormon ten zwiększa wydzielanie hormonu przysadkowego
.
Przykładem sprzężenia zwrotnego dodatniego może być wydzielanie oksytocyny , której stężenie gwałtownie
wzrasta w ostaniej fazie porodu w trakcie przesuwania się płodu w dół i poszerzania kanału rodnego. Zwiększone
stężenie oksytocyny w tym okresie stymuluje przysadkę do uwalniania jej jeszcze większych ilości aby nasilić
skurcze macicy.
3) Regulacja przez przekształcanie hormonu nieaktywnego do formy aktywnej w komórkach
Mechanizm ten polega na tym, że w tkankach dochodzi do przekształcania nieaktywnej formy hormonu do jego
postaci czynnej biologicznie, zdolnej do wywoływanie efektów w komórce docelowej. Przykładami tego sposobu
regulacji mogą być:
●
testosteron przekształcany w tkankach do dihydrotestosteronu
●
tyroksyna przekształcana docelowo w czynną trójjodotyroninę
Hormony trzustkowe
Trzustka jak wiadomo pełni funkcję zarówno wewnątrz- jak i zewnątrzwydzielnczą. W kontekście hormonów
ważniejsza jest oczywiście jest funkcja wewnątrzwydzielnicza. Sprawują ją stanowiące do 2% masy trzustki wyspy
Langerhansa znajdujące się w przewadze w ogonie i głowie trzustki. Wyspy te są utworzone przez cztery typy komórek
wydzielających różne hormony:
●
komórki α – wydzielają glukagon (60-70% wszystkich komórek)
●
komórki β – wydzielają insulinę (20% wszystkich komórek)
●
komórki D (δ) – wydzielają somatostatynę
●
komórki F (PP) – wydzielają polipeptyd trzustkowy
a) budowa
● zbudowany z dwóch łańcuchów aminokwasowych:
-A (21 reszt aminokwasowych)
-B (30 reszt aminokwasowych)
● łańcuchy połączone są dwoma mostkami disiarczkowymi, ponadto w
łańcuchu A występuje wewnątrzłańcuchowy mostek disiarczkowy
4

Układ wydzielania wewnętrznego (
wer.robocza
)
● dzięki krótkiemu okresowi półtrwania – około 6 minut – stężenie insuliny może być szybko zmieniane,
zależnie od sytuacji metabolicznej
b) biosynteza
●
prekursorem insuliny jest preproinsulina, która po wycięciu jednej części sekwencji aminokwasów tworzy
jednołańcuchową proinsulinę połączona z peptydem C (peptydem łączącym)
●
proinsulina w aparacie Golgiego ulega ponownemu skróceniu (o peptyd C) i związaniu mostkami
disiarczkowymi tworząc dwułańcuchową aktywną metabolicznie insulinę
●
insulina jest następnie pakowana do pęcherzyków cytoplazmatycznych i przenoszona w okolice błony
komórkowej
c) regulacja wydzielania
●
najważniejszym czynnikiem kontrolującym wydzielanie insuliny jest stężenie glukozy we krwi
✓
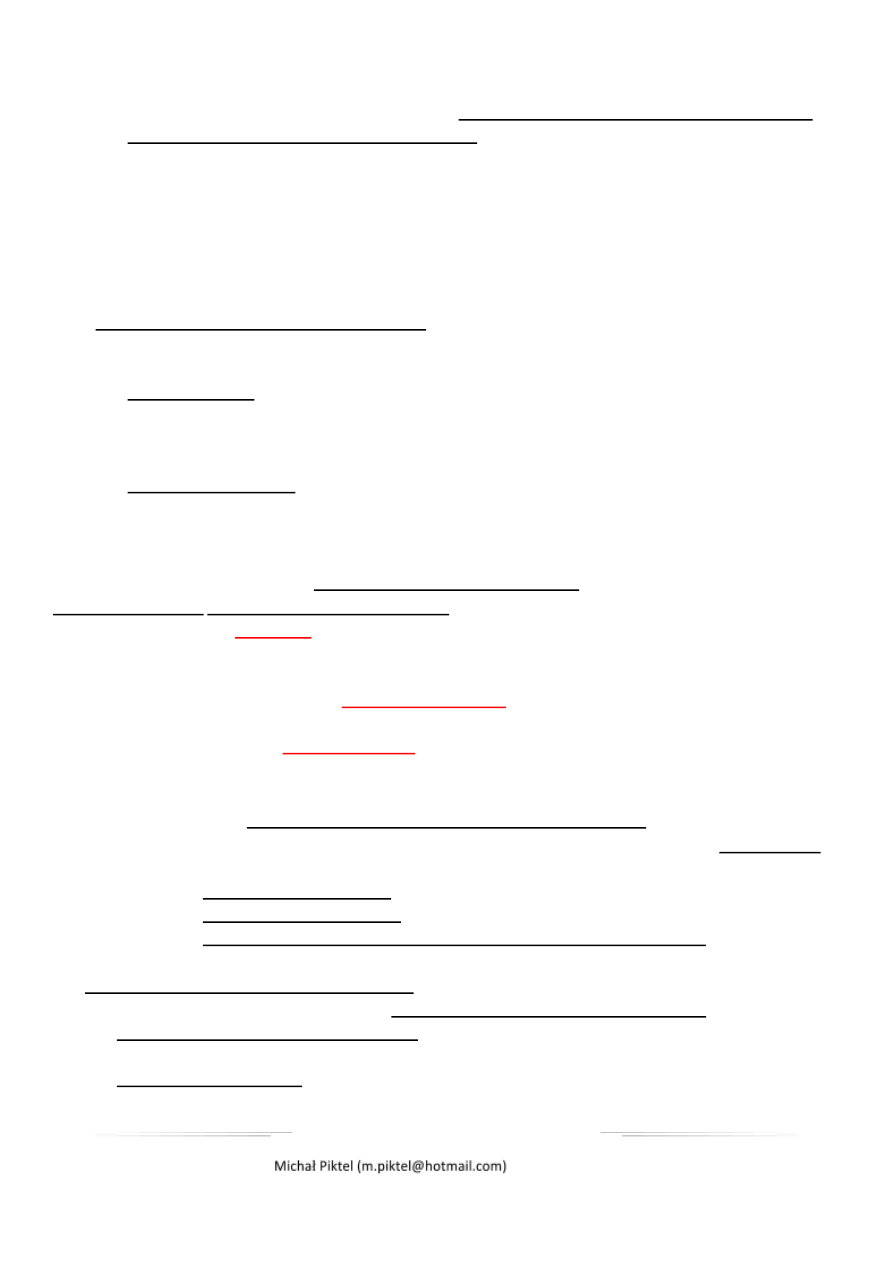
przenośnik glukozy przenosi ją do komórki β trzustki gdzie ulega ona cyklom przemian metabolicznych
prowadzących do wzrostu stężenia ATP w komórkach
✓
podwyższony poziom ATP w cytoplazmie zamyka ATP-zależna kanały potasowe i prowadzi do zmiany
potencjału błonowego – dochodzi do depolaryzacji
✓
depolaryzacja błony komórkowej otwiera bramkowane napięciem kanały Ca
2+
, które napływają do
komórki
✓
podwyższony poziom jonów Ca
2+
umożliwia egzocytozę ziarnistości zawierających insulinę
●
podobny jak glukoza wpływ na wydzielanie insuliny posiadają:
cholecystokinina, glukagon, GLP-1,
acetylocholina, aminokwasy: arginina, leucyna
●
hamujący wpływ ma natomiast
somatostatyna
, oraz
hipotermia
,
hipoksja
,
stres
i
wysiłek fizyczny
Wyżej omówione wydzielanie insuliny jest reakcją dwufazową:
●
faza szybka – polega na gwałtownym wyrzucie insuliny zgromadzonej w ziasrnistościach komórek β
●
faza wolna – polega na syntezie ziarnistości pod wpływem działania czynników stymulujących
d) receptory insulinowe
Receptor insuliny ze względu na jej charakter białkowy znajduje się na powierzchni komórki, jednak w odróżnieniu
do typowych receptrów błonowych insulina działa bez udziału wtórnego przekaźnika
●
jest to białko tetrameryczne (α
2
β
2
) składające się z
✓ dwóch podjednostek α – położonych na zewnątrz błony komórkowej, są domenami wiążącymi insulinę
✓ dwóch podjednostek β – składających się z domen wewnątrz- i zewnątrzkomórkowej, są białkami o
aktywności enzymatycznej kinazy tyrozynowej
●
przyłączenie do podjednostki α insuliny powoduje autofosforylację grup tyrozynowych
wewnątrzkomórkowej domeny podjednostki β
●
zaktywowana kinaza tyrozynowa powoduje fosforylację szeregu białek w komórce doprowadzając do
określonych zmian aktywności metabolicznej komórek docelowych
e) komórkowe efekty działania insuliny
5
Układ wydzielania wewnętrznego (
wer.robocza
)
●
przyłączona insulina zwiększa liczbę przenośników glukozy w błonach co pobudza jej transport do wnętrza
komórek, niektóre komórki np.erytrocyty, komórki nerwowe mają system transportu glukozy niezależny od
insuliny
●
najszybszą odpowiedzią, pojawiającą się już po kilku sekundach od związania insuliny z receptorem, jest
wzrost transportu glukozy do komórek, po kilku minutach lub godzinach wzrasta aktywność enzymatyczna,
natomiast efekty indukcji produkcji białek enzymatycznych pojawiają się nawet po kilku dniach
f) Narządowe efekty działania insuliny
Wątroba
●
spadek poziomu glukozy poprzez:
↓ glukoneogenezy
↓ glikogenolizy
↑
glikogenogenezy
●
↓ ketogenezy
●
↑ syntezy białek
●
↑ syntezy lipidów
6

Układ wydzielania wewnętrznego (
wer.robocza
)
Mięśnie
↑ transportu glukozy do miocytów
↑ glikogenogenezy
↑ wychwytu wolnych aminokwasów
↑ syntezy białek
↑ wychwytu jonów K+
Tkanka tłuszczowa
↑ transportu glukozy do adipocytów
↑ syntezy kwasów tłuszczowych
↑ syntezy
glicerolo-3-
fosforanu
↓ lipazy hormonowrażliwej
↑ lipazy lipoproteinowej
↑ wychwytu K
+
G
LUKAGON
a) budowa i biosynteza
●
budowa polipeptydowa, zbudowany z jednego łańcucha o 29 aminokwasach
●
sekwencja aminokwasowa jednakowa u wszystkich ssaków
●
syntetyzowany w postaci prekursora - preglukagonu ulegającego
odpowiednim obróbkom do czynnego glukagonu
●
należy, razem z adrenaliną i kortyzolem do grupy hormonów przeciwregulacyjnych działających
antagonistycznie do insuliny
●
okres półtrwania 5-10 minut
b) komórkowe efekty działania glukagonu
●
receptory glukagonu są typowymi receptorami błonowymi działającymi poprzez wtórny przekaźnik
●
podjednostka α-GTP białka G odłącza się od niego pod wpływem przyłączenia glukagonu i aktywuje enzym
cyklazy adenylanowej
●
cyklaza adenylanowa przekształca cytosolowy ATP w 3’5’-AMP (cAMP)
●
cAMP aktywuje kinazę białkową A, która fosforyluje białka enzymatyczne, modyfikując ich aktywność
katalityczną, może fosforyzować histony i białka rybosomowe, pobudzając procesy transkrypcji i translacji
c)
metaboliczne i narządowe efekty działania glukagonu
(glukagon nie działa na mięśnie szkieletowe !)
●
metabolicznie glukagon powoduje:
↑ glukoneogenezy
↑ glikogenolizy
↑lipolizy (aktywacja lipazy hormonowrażliwej)
7
Układ wydzielania wewnętrznego (
wer.robocza
)
↑ ketogeneza
↓ syntezy białek
↓ lipogenezy
Serce
●
działanie inotropowe dodatnie (wzmagają kurczliwość
mięśnia sercowego)
●
działania chronotropowe dodatnie (wzmagają
częstotliwość skurczów mięśnia sercowego)
Układ pokarmowy
↑ wydzielanie żółci
↑ wydzielanie jelitowe
↓ motoryka żołądka
↓ wydzielanie żołądkowe
↓ wydzielanie trzustkowe
Inne:
↑ wydzielanie insuliny i somatostatyny
d) regulacja wydzielania glukagonu
●
wydzielanie glukagonu jest stymulowane obniżeniem stężenia glukozy we krwi obwodowej, np. w ciągu nocy
lub podczas długich przerw między posiłkami
●
podwyższone stężenie glukagonu zapobiega więc hipoglikemii
●
aminokwasy pochodzące z pożywienia, głównie alanina, seryna, glicyna, cysteina i treonina, pobudzają
wydzielanie glukagonu jak i insuliny
●
podobnie pobudzająco działają
podwyższone stężenia adrenaliny i noradrenaliny
, które działają niezależnie
od stężenia cukru we krwi – stężenie glukagonu jest podwyższane w oczekiwaniu na skutki zwiększonego
zużycia glukozy
●
ponadto pobudzająco na wydzielanie glukagonu działają:
CCK, stres i wysiłek fizyczny
●
hamująco działają natomiast:
wzrost stężenia glukozy, insulina i wzrost stężenia WKT
S
OMATOSTATYNA
●
jest to 14-aminokwasowy peptyd, którego głównym zadaniem jest
hamowanie wydzielania insuliny i glukagonu na
zasadzie działania parakrynnego
●
ponadto hormon ten może opóźniać opróżnianie żołądka i pęcherzyka żółciowego, oraz:
✓ ↓sekrecję gastryny
✓ ↓ wydzielanie enzymów trawiennych przez trzustkę
8

Układ wydzielania wewnętrznego (
wer.robocza
)
P
OLIPEPTYD TRZUSTKOWY (
PP
)
●
jest to hormon peptydowy, składający się z 36-aminokwasów, którego największy wzrost następuje po posiłku
bogatym w białka
●
stanowi
silny inhibitor enzymów trzustkowych
, oraz
hamuje obkurczanie pęcherzyka żółciowego
, a jego
podwyższone stężenie we krwi utrzymuje się stosunkowo długo
●
zmniejszenie wrażliwości tkankowej na działanie tego hormonu lub niedostateczny poziom insuliny we krwi
prowadzi do objawów cukrzycy
Tarczyca
Jest to gruczoł dokrewny położony z przodu i bocznie od krtani, dzieli się na dwa płaty połączone ze sobą cieśnią.
Podstawową jednostką budulcową tarczycy jest
pęcherzyk
, który składa się z błony podstawnej na której spoczywają
komórki pęcherzykowe
●
wnętrze pęcherzyka wypełnia koloid, a sam pęcherzyk otoczony jest niewielką ilością tkanki łącznej i bardzo gęstą siecią
naczyń krwionośnych
●
nieaktywne pęcherzyki są duże, bogate w koloid natomiast pęcherzyki aktywne są małe i ubogie w koloid
a) powstawanie hormonów tarczycy
●
komórki pęcherzykowe wydzielają do wnętrza koloidu tyreoglobulinę (TG) z zawartą w niej tyrozyną
●
następnie z krwi aktywnie wychwytywane są jodki (120 µg/dobę), odbywa się to dzięki symportowi sodowo-
jodowemu, a inhibitorami tego procesu są: tioycyjanina i nadchloran
●
do koloidu wydzielany jest enzym tyreoperoksydaza
●
tyreoperoksydaza pozwala jodkom na przejście w jod atomowy co powoduje:
-
jodowanie tyrozyny zawartej w tyreoglobulinie tworząc
MIT
(
monojodotyrozynę
) oraz
DIT
(
dijodotyrozynę
)
-
kondensację jodowanych tyrozyn
MIT + DIT = T
3
oraz
DIT + DIT= T
4
●
w ten sposób jodowana tyreoglobulina zostaje zmagazynowana w postaci koloidu w świetle pęcherzyków
tarczycy
●
pod wpływem tyreotropiny (TSH) komórki pęcherzykowe pobierają i rozkładają jodowaną tyreoglobulinę
tworząc aktywne hormony wędrujące wraz z krwią do komórek docelowych
-
90% hormonów uwalnianych stanowi T
4
, który jest właściwie prohormonem dla T
3
(w tkankach
obwodowych T
4
ulega przekształceniu do T
3
●
fakt iż w tarczycy zgromadzony jest zapas T
3
i T
4
związanych z tyreoglobuliną zabezpiecza prawidłowe
stężenie tych hormonów we krwi przez co najmniej 2 miesiące
b)
transport T
3
i T
4
we krwi odbywa się poprzez:
●
TGP – globulinę wiążącą tyroksynę
●
TBPA – prealbuminę wiążącą tyroksynę
●
TBA – albuminę wiążącą tyroksynę
●
niewielki procent przenoszony jest w postaci wolnej
●
w krążaniu znajduje się również odwrotna T
3
(rT
3
), która jest nieaktywna biologicznie
c) regulacja wydzielania hormonów tarczycy
●
TRH (tyreoliberyna) wydzielana jest przez podwzgórze w odpowiedzi na zimno, sen itd.
9

Układ wydzielania wewnętrznego (
wer.robocza
)
●
liberyna podwzgórzowa działa indukująco na przysadkę mózgową stymulując ją do wydzielania tyreotropiny
(TSH)
●
TSH wywołuj opisany wyżej efekt, a uwolnione w ten sposób T
3
i T
4
działają hamująco zarówno na
wydzielanie TRH w podwzgórzu jak i na TSH w przysadce
Działanie tyroksyny (T
4
) i trójjodotyroniny (T
3
)
●
hormony te są w stanie
zwiększyć podstawową przemianę materii (PPM) nawet o 60-100 %
●
T
3
ma duzo wyższe powinowactwo do receptorów niż T
4
●
są to enzymy wyjątkowe – pomimo swojej peptydowej budowy ich receptory znajdują się w jądrze komórkowym,
a więc ich działanie będzie opierało się na indukowaniu transkrypcji odpowiednich genów, następie wytworzone w
ten sposób mRNA w cytosolu będzie kodowało odpowiednie sekwencje aminokwasowe tworzonych białek.
-
z tego powodu efekt działnia hormonów tarczycy na komórki docelowe jest widoczny dopiero po pewnym
okresie czasu (dla T
3
jest to kilka godzin) okres ten nazywamy
„okresem działania utajonego”
Sumarycznie efekty działania T
3
i T
4
polegają na:
●
nasileniu termogenezy
(głównie przez wzrost aktywności enzymu Na
+
/K
+
-ATP-azy w błonach komórkowych)
●
zwiększeniu zużycia tlenu
(z wyjątkiem mózgowia, gonad męskich, macicę, śledzionę, tkankę limfatyczną oraz
przedni płat przysadki)
●
stymulacja do syntezy białek
(fizjologiczne stężenie hormonów stymulują syntezę białek, natomiast ich nadmiar
prowadzi do ich zwiększonego rozpadu)
●
zwiększenie stężenia glukozy
we krwi poprzez:
↑ glikogenolizy
↑ glukoneogenezy
↑ wchłaniania glukozy z przewodu pokarmowego
●
wpływ na układ krążenia (
zwiększenie siły skurczów i ich częstości
, co sumarycznie daje
wzrost ciśnienia tętniczego
)
●
zwiększenie wrażliwości tkanek na katecholaminy
oraz współdziałanie z nimi (działanie synergistyczne)
●
układ nerwowy
– hormony tarczycy są niezbędne do prawidłowego rozwoju układu nerwowego
-
niedoczynność tarczycy powoduje różnego stopnia niedorozwój umysłowy, łącznie z kretynizmem, zwolnienie
procesów myślowych
-
nadczynność tarczycy powoduje wzmożoną pobudliwość osobnika, przyspieszenie procesów myślowych
Hormony regulujące metabolizm wapnia
W organizmie znajduje się około 1100 g wapnia (25 moli) związanego głównie z tkanka kostną. Najważniejszą
rolę w procesach metabolicznych odgrywa wapń zjonizowany w postaci dwuwartościowego kationu Ca
2+
.
10

Układ wydzielania wewnętrznego (
wer.robocza
)
o
fizjologiczne stężenie Ca
2+
w osoczu wynosi 2,5 mmol/L
o
wapń krąży związany z albuminami, cytrynianem lub jako Ca
2+
o
w organizmie tworzy dwie pule:
●
łatwowymienialną (ok. 100 µmoli) – związaną z płynami ustrojowymi, służy do regulacji stężenia
Ca
2+
●
trudnowymienialną (ok. 25 000 µmoli) – związaną z hydroksyapatytami, odpowiadającą za
remodelację kości
Rola wapnia w organizmie:
o
bierze udział w krzepnięciu krwi
o
odpowiada za kurczliwość mięśni
o
odpowiada za pobudliwość nerwowo – mięśniową
o
mineralizuje kości
o
stanowi jeden z wtórnych przekaźników nerwowych
Przeciętnie spożywamy około 1 gram wapnia w ciągu doby, z czego do krwi wchłania się około 50-60%.
Nadmiar wapnia w organizmie powoduje rozwój zespołu chorobowego jakim jest tężyczka, której objawy
kliniczne to nadmierna pobudliwość nerwów i mimowolne skurcze mięśniowe.
Znaczny nadmiar wapnia w organizmie prowadzi do śpiączki hiperkalcemicznej.
Metabolizm wpnia jest ściśle regulowany przez trzy hormony:
o
parathormon (PTH)
o
kalcytoninę
o
witaminę D3
Parathormon (PTH)
Jest to 84-aminokwasowy hormon polipeptydowy wydzielany przez przytarczyce leżące na tylnej powierzchni
gruczołu tarczowego. Jest gromadzony w pęcherzykach komórek przytarczyc, skąd uwalniany jest w
odpowiedzi na spadek stężenia Ca2+ w osoczu
Mechanizm oddziaływania PTH na komórki
Podobnie jak inne peptydowe hormony PTH posiada swój swoisty receptor w błonie komórkowej komórek
docelowych.
●
wtórnym przekaźnikiem PTH jest cAMP powstająca pod wpływem aktywowanej cyklazy adenylanowej
Efekty wywoływane działaniem PTH
●
obniżenie stężenia Ca2+ zawartego w osoczu aktywuje pęcherzyki przytarczyc do wydzielania z ich
ziarnistości PTH
●
trójmiejscowe działanie PTH ma za zadanie przede wszystkim podwyższyć obniżony poziom jonów
Ca2+, dzieje się to poprzez:
o
wzrost uwalniania (resorpcji) Ca2+ z kości
o
wchłanianie zwrotne Ca2+ w kanalikach nerkowych
o
wzrost wchłaniania Ca2+ z pożywienia
●
dokładniej w poszczególnych narządach wywołuje on następujące efekty:
o
w układzie kostnym może powodować:
▪
normokalcemię – remodeling kości – wzrost resorpcji i kości otworzenia
11

Układ wydzielania wewnętrznego (
wer.robocza
)
▪
hiperkalcemię – powodującą patologiczną demineralizację kości (współdziałając z
witaminą D3)
o
w nerkach
▪
spadek zwrotnej resorpcji fosforanów co powoduje wzrost ich stężenia w moczu
▪
wzrost wytwarzania aktywnej dormy witaminy D (kalcytriolu)
o
w układzie pokarmowym
▪
zwiększona ilość witaminy D3 powoduje wzrost wchłaniania Ca2+ z pożywienia
Kalcytonina
Jest to 32-aminokwasowy hormon peptydowy regulujący stężenie wapnia w organizmie, tworzony przez
komórki parafolikularne (komórki C) tarczyc. Hormon ten jest odpowiedzialny za obniżanie po posiłkowego
poziomu Ca2+.
●
głównym czynnikiem pobudzającym komórki C do wydzielania kalcytoniny jest wzrost stężenia Ca2+ w
osoczu krwi
Efekty działania:
●
ogólnie kalcytonina obniża poziom wapnia poprzez:
o
spadek uwalniania Ca2+ w kościach (osiągany poprzez supresję, czyli hamowanie aktywnych
osteoklastów)
o
zwiększenie zwrotnego wchłaniania Ca2+ w nerkach co zwiększa jego wydalanie z moczem
●
innymi efektami działani kalcytoniny jest:
o
spadek wydzielania żołądkowego i trzustkowego
o
spadek łaknienia
o
tzw. działanie analgetyczne – polega na wzroście uwalniania endogennych opiatów oraz
na wzroście opatowych receptorów w podwzgórzu. Sumarycznie powoduje to mniejszą
pobudliwość na bodźce bólowe (zwiększony próg pobudliwości)
Witamina D
3
(cholekalcyferol)
✓ zaliczana jest do grupy witamin mających charakter egzogenny, lecz skóra człowieka pod
wpływem promieni słonecznych zdolna jest do syntezy witaminy D w ilościach wystarczających
organizmowi
✓ substratem syntezy witaminy D jest 7-dehydrocholesterol występujący w małych ilościach we
wszystkich tkankach, przekształcanie drogą fotochemiczną tego związku daje cholekalcyferol
(witamina D
3
)
✓ cholekalcyferol powstały w skórze jest transportowany poprzez krew do wątroby gdzie ulega
przekształceniu do 25-hydroksycholekalcyferolu (kalcydiol)
✓ kalcydiol transportowany jest do nerek, gdzie pod wpływem PTH jest rozkładany do aktywnej
formy – kalcytriolu oraz do nieaktywnego metabolitu – 24,25-dihydocholekalcyferolu
✓ kalcytriol stanowi główną, fizjologicznie czynną formę witaminy D, jest hormonem steriodowym,
która przenika przez błonę komórkową (jako witamina rozpuszczalna w tłuszczach) i oddziałuje
na receptor jądrowy, powodując sumarycznie:
-
podwyższenie stężenie Ca
2+
w osoczu, przez:
o ułatwienie resorpcji Ca
2+
i fosforanów z
kości
o pobudzenie resporpcji Ca
2+
w nerkach
12

Układ wydzielania wewnętrznego (
wer.robocza
)
o
pobudzenie absorpcji Ca
2+
w jelicie cienkim
Hormony kory nadnerczy
Nadnercza są to parzyste gruczoły leżące na górnych biegunach nerek, na przekroju wyróżniamy w nich dwie odmienne
części wydzielające zupełnie różne hormony:
●
część korowa
●
część zewnętrzna
Hormony kory nadnerczy zaliczamy do grupy hormonów steroidowych, których wspólnym prekursorem jest cholesterol.
Histologicznie w korze nadnerczy wyróżniamy trzy warstwy, z których każda wydziela inny hormon, idąc od zewnątrz są
to:
●
warstwa kłębkowata – wydziela Mineralokortykoidy, głównie aldosteron
●
pasmowata – wydziela glikokortykosteroidy, głównie kortyzol
●
warstwa siateczkowata –wydziela hormony płciowe androgeny: dehydroepiandrosteron (DHEA) i androstendion
Kontrolę hormonalną nad wydzielaniem wszystkich hormonów warstwy korowe nadnerczy sprawuje
przysadkowy hormon adrenokortykotropowy (ACTH) powstający pod wpływem kortykoliberyny (CRH)
wydzielanej przed podwzgórze w wyniku strachu, stresu oraz w czasie cyklu sen/czuwanie.
Aldosteron
Jest to hormon składający się z 21 atomów węgla, we krwi transportowany najczęściej w stanie wolnym. W
komórkach docelowych wiąże się z receptorem wewnątrzkomórkowym, który aktywowany poprzez połączenie
z aldosteronem indukuje w jądrze komórki syntezę mRNA kodującego syntezę białek transportujących Na+,
głównie kanałów Na+, K+ i ATP-azy.
●
głównym miejscem działania aldosteronu są kanaliki dalsze i cewki zbiorcze nerek
●
aldosteron zwiększa tam resorpcję zwrotną jonów Na+ a zwiększa wydalanie jonów K+ i H+
●
zwiększona ilość Na+ powoduje wzrost objętości osocza, oraz zwiększa wrażliwość tętniczek na
działanie substancji je zwężających, sumarycznie powodując wzrost ciśnienia tętniczego
Regulacja wydzielania aldosteronu
●
zasadniczym czynnikiem regulującym wydzielanie aldosteronu jest peptyd angiotensyna II, powstający
w wyniku działania enzymu reniny, dlatego mówimy o układzie renina-angiotensyna-aldosteron
13

Układ wydzielania wewnętrznego (
wer.robocza
)
o
renina tworzona jest w nerkach (aparat przykłebuszkowy) w zwiększonych ilościach
w odpowiedzi na obniżenie ciśnienia tętniczego krwi lub obniżenia objętości płynu
zewnątrzkomórkowego oraz w wyniku wzrostu aktywności układu androgenicznego
o
renina oddziaływuje na angiotensynogen, który ulega konwersji do angiotensyny bedącej
formą nieaktywną
o
angiotensyna I jest konwertowana pod wpływem enzymu konwertującego do aktywnej
angiotensyny II
o
angiotensyna II stymuluje warstwę kłębkowatą do uwalniania aldosteronu oraz obkurcza
tętniczki obwodowe
●
innym czynnikiem regulującym, lecz w znacznie słabszym stopniu, wydzielania aldosteronu jest
przysadkowy ACTH, który stymuluje wydzielanie tego hormonu jedynie w reakcjach stresowych
●
czynnikiem zwiększającym wydzielanie aldosteronu jest ponadto podwyższone stężenie jonów K+ w
osoczu, obniżenie stężenia Na+ osoczowe wywiera niewielki efekt stymulujący
Kortyzol
Hormon składający się z 21 atomów węgla, zaliczany do grupy glikokortykosteroidów (glukokortykoidów),
wydzielany przez warstwę pasmowatą kory nadnerczy. Kortyzol transportowany jest we krwi przez swoiste
białko transportujące – transkortynę (globulinę wiążącą steroidy).
●
uwolniony od transportera kortyzol łączy się ze swoimi swoistymi receptorami wewnątrzkomórkowymi
w komórkach docelowych
●
działanie kortyzolu w komórce polega na wzbudzeniu ekspresji odcinka DNA odpowiedzialnego za
kodowanie odpowiednich enzymów odpowiadających wspólnie za:
o
wzmożony katabolizm białek, szczególnie w mięśniach szkieletowych i kościach (nadmiar
kortyzolu przedstawia się jako ujemny bilans azotowy)
o
stymulację hepatocytów do glukoneogenezy i glikogenogenezy
o
hamowanie wpływu insuliny na komórki, co powoduje wzrost ilości glukozy we krwi
o
aktywację lipolizy przez aminy katecholowe (wpływ przyzwalający)
o
zwiększenie liczby erytrocytów, neutrofili i trombocytów i spadek liczby limfocytów, eozynofili
i bazofili
o
hamowanie reakcji immunologicznych, alergicznych i zapalnych
o
wzrost wrażliwości naczyń na noradrenalinę → wzrost RR
Regulacja wydzielania
●
wydzielanie kortyzolu pozostaje pod wyłączonym wpływem pobudzającego działania ACTH, którego
wzrost stężenia we krwi powoduje szybki wzrost uwalniania kortyzolu
●
kortyzol jako jedyny z hormonów kory nadnerczy tworzy petlę sprzężenia zwrotnego z ACTH i CRH –
hamuje ich uwalnianie przez przysadkę i podwzgórze
●
stężenie kortyzolu podlega wahaniom dobowym – najwyższe stężenie wystepuje około godziny 6-8
rano, najniższe zaś o północy
Dehydroepiandrosteron (DHEA) i androstendion
●
adrenarche – gwałtowny wzrost stężenia DHEA przed okresem pokwitania, powodujący przypieszenie
tempa wzrostu, charakteryzuje się:
o
wzrostem masy mięśniowej
14

Układ wydzielania wewnętrznego (
wer.robocza
)
o
odkładaniem wapnia w kościach
o
rozwojem prącia i łechtaczki
o
pojawieniem się owłosienia łonowego, zarostu na twarzy i owłosienia pachowego
o
powiększeniem chrząstek krtani
Podsumowanie
●
często do chodzi do nadczynności warstw kory nadnerczy w odpowiedzi na patologicznie wysokie
stężenie ACTH lub przez guzy autonomiczne kory nadnerczy
o
nadmierne wydzielanie aldosteronu prowadzi do podwyższonego ciśnienia tętniczego,
obniżonego stężenia jonów K+ i podwyższonego stężenia Na+ we krwi (zespół Conna)
o
nadmierne wydzielanie kortyzolu powoduje powstanie zespołu Cushinga, który objawia się
nadmiernym pobudzeniem katabolizmu (spadek grubości skóry, atrofia), odkładaniem tkanki
tłuszczowej w obrębie brzucha i karku, osteoporozą i wzrostem poziomu cukru we krwi.
●
niedobór hormonów kory nadnerczy występuje jako skutek uszkodzenia układu
autoimmunologicznego lub powikłanie pod chirurgicznym usunięciu nadnerczy
o
skutkiem jest obniżone ciśnienie tetnicze, obniżone stężnie Na+ i podwyższone stężenie K+ we
krwi, a także obniżone stężenie w niej glukozy.
o
nieleczona niedoczynność nadnerczy prowadzi do zapaści i śmierci organizmu
Hormony podwzgórza
Zaliczane jest do międzymózgowia, posiada dwa odrębne zespoły komórek:
a)
pole hipofizjotropowe (wyniosłość pośrodkowa) – utworzone przez jądra drobnokomórkowe
wydzielające substancje pobudzające (liberyny) i hamujące (statyny) wydzielanie hormonów przedniego
płata przysadki
●
liberyny:
●
Gonadoliberyna, działająca na uwalnianie FSH i LH
●
Kortykoliberyna, działająca na uwalnianie ACTH
●
Tyreoliberyna, działająca na uwalnianie TSH
●
Somatoliberyna, działająca na uwalnianie GH
●
statyny:
●
Somatostatyna, działająca na hamowanie uwalniania GH
●
Prolaktostatyna, działająca na hamowanie uwalniania PRL
b)
jądra wielkokomórkowe, czyli jądra nadwzrokowe i przykomorowe, które syntetyzują dwa
hormony transportowane następnie przez aksony tyh neuronów do tylnego płata przysadki
mózgowej gdzie są gromadzone.
Do neurohormonów podwzgórza zaliczamy:
●
hormon antydiuretyczny (ADH, wazopresyna)
●
oksytocynę
Hormony przysadki mózgowej
Przysadka mózgowa leżyy w siodle tureckim kości klinowej, Ze względu na różne pochodzenie oraz odmienne
struktury i funkcje wyróżniamy: przysadkę gruczołową i przysadkę nerwową.
1)
Przysadka gruczołowa, rozwijająca się z
a)
jej zasadnicze części to:
15

Układ wydzielania wewnętrznego (
wer.robocza
)
●
część dalsza (płat przedni)
●
część pośrednia
●
część guzowa
b)
Hormony płata przedniego przysadki gruczołowej
●
adrenokortykotropina (ACTH) – działająca na korę nadnerczy. Łączy się ona tam ze swoistymi receptorami
w błonie komórkowej i stymuluje syntezę i wydzielanie hormonów steroidowych, zwłaszcza kortyzolu i w
mniejszym stopniu kortyzolu.
-
wydzielanie ACTH podlega wahaniom dobowym, najwyższe stężenie osiągane jest w godzinach rannych
-
ponadto ACTH może zostać uwolnione w wyniku reakcji organizmu na sytuacje stresowe (hipoglikemia, ból,
strach, gorączka)
-
nadmierne wydzielanie ACTH obserowane w przebiegu gruczolaków przysadki powoduje przerost
nadnerczy i ich nadczynność, wywołując zespół hipokortyzolemii
-
niedobór ACTH może być powikłaniem uszkodzenia podwzgórza lub przysadki nerwowej, prowadząc do
wtórnej niewydolności nadnerczy
●
tyreotropina (TSH) – jest regulatorem funkcji tarczycy, pobudzając ją do uwalniania T3 i T4
-
wydzielanie TSH jest pobudzane przez podwzgórzową tyreotropinę
-
wydzielanie TSH jest hamowane przez podwzórzową somatostatynę oraz na zasadzie sprzężenia zwrotnego
przez hormony tarczycy
●
hormon folikulotropowy (FSH) i hormon lutenizujący (LH)
-
FSH – pobudza u kobiet dojrzewanie pęcherzyków Graafa w jajniku i wydzielanie przez nie estradiolu; u
mężczyzn natomiast stymuluje spermatogenezę
-
LH – pobudza u kobier syntezę progesteronu w pęcherzyku Graafa i podtrzymuje funkcję wydzielniczą ciałka
żółtego; u mężczyzn natomiast stymuluje wytwarzanie i wydzielanie testosteronu przez komórki Leydiga
jąder
●
hormon wzrostu (GH) – hormon ten wywiera na organizm wpływ bezpośredni i pośredni
-
pośredni – polega na stymulacji organizmu do tworzenia insulinopodobnych czynników wzrostu (IGF-I i IGF-
II) powstających w wątrobie oraz innych tkankach, głównym czynnikiem oddziałującym na tkanki jest IGF-I,
który zwiększa synteze białka w mięśniach prowadzać do wzrostu ich masy oraz zwiększa wzrost kości
-
bezpośredni – polega na:
16

Układ wydzielania wewnętrznego (
wer.robocza
)
✓ zwiększeniu lipolizy w tkance tłuszczowej
✓ zmniejszeniu transportu glukozy do miocytów mięśni szkieletowych
✓ zwiększeniu różnicowania chondrocytów w kosciach (wzrost chrząstek nasadowych)
✓ oraz zwiększenie produkcji glukozy przez wątrobę
-
gigantyzm – nadmierne wydzielanie GH u dzieci i młodzieży spowodowane najczęściej gruczolakami
przysadki prowadzące do nadmiernego wzrostu organizmu
-
akromegalia – nadmierne wydzielanie GH u dorosłych powodujące powiększenie dystalnych części ciała
(stopy, dłonie) oraz twarzy i narządów gruczołowych
-
karłowatość przysadkowa – niedobór GH prowadzacy do zahamowania wzrostu u dzieci
-
niedobór GH u dorosłych jest najcześciej następstem operacji, radioterapii, a charakteryzuje się
zmniejszeniem masy mięśniowej, zwiększeniem ilości tkanki tłuszczowej trzewnej, zmniejszeniem gęstości
kości i zaburzeniami gospodarki węglowodanowej i lipidowej
●
prolaktyna (PRL) – mimo że hormon te wystepuje u obu płci, jego działanie wykazano jedynie w organizmach
kobiet gdzie stymuluje on wytwarzanie mleka w okresie poporodowym, oraz bierze udział w rozwoju gruczołów
piersiowych w okresie wzrostu
2)
Przysadka nerwowa, magazynuje hormony wydzielone przez jądra wielkokomórkowe podwzórza, a
więc:
●
hormon antydiuretyczny (ADH, wazopresyna) – oddziałująca na cewki zbiorcze w nerkach,
zwiększając wchłanianie zwrotne wody, a w wysokich dawkach podnosi ciśnienie krwi
●
oksytocyna – wywołuje skurcze porodowe mięśni gładkich macicy
17
Wyszukiwarka
Podobne podstrony:
Hormony (14 12)
MPLP 360;361 02.12.;14.12. 2012
14 12 08
14 12 2012
Finanse publiczne 14 12 13
14 12 2006
DERMATOLOGIA 14.12, dermatologia
Podstawy turystyki 14.12.2008, Turystyka I Rekrecja, podstawy turystyki
11. 14-12-2010 Kontury i kolory
14 2 12
14 12 86
14 12 2012r Stal wykład
14 12 2015 W 9 harmonogram konsult zima 2015 16 popraw
Wykład 11 [14.12.05], Biologia UWr, II rok, Zoologia Kręgowców
Hormony 10.12.2009
9 Uskoki (14 12 2010)
W9 14 12 121 rynek
więcej podobnych podstron