
prof. Wiesław Przybylski
prof. Wiesław Przybylski
KATEDRA TECHNIKI I
KATEDRA TECHNIKI I
TECHNOLOGII GASTRONOMICZNEJ
TECHNOLOGII GASTRONOMICZNEJ

Zarys historii genetyki
Zarys historii genetyki

Genetyka stanowi wyodrębnioną dyscyplinę nauk
biologicznych, której przedmiotem badań jest zmienność i
dziedziczenie cech organizmów żywych. Powstanie tej
dyscypliny stało się możliwe dzięki intensywnemu rozwojowi
biologii w wieku XIX. Postęp ten znaczony był z jednej strony
poznawaniem struktury i funkcji organizmów, tkanek, komórek
i ich organelli, a z drugiej formułowaniem przełomowych
hipotez i teorii, wśród których część miała epokowe znaczenie
dla rozwoju wiedzy o świecie ożywionym. Opis struktury i
funkcji organizmów na poziomie komórkowym lub
wewnątrzkomórkowym jest zależny od dostępnych urządzeń i
metod badawczych. Tym samym rozwój nauk biologicznych
opiera się na osiągnięciach fizyków, chemików, konstruktorów,
a ostatnio również informatyków.

W wieku XIX sformułowano trzy teorie o ogromnym
znaczeniu dla dalszego rozwoju biologii. Autorami
pierwszej z nich – teorii komórkowej budowy
organizmów, ogłoszonej w 1838 roku, byli Schwann i
Schleiden. Główną tezą tej teorii było twierdzenie, że
wszystkie rośliny i zwierzęta zbudowane są z komórek,
które mogą powstać jedynie przez podział innych
komórek. Okazało się to niezwykle stymulujące dla
rozwoju cytologii, czyli nauki o budowie i funkcji
komórek. Dwadzieścia lat później – w 1859 roku –
Darwin opublikował dzieło pt. O powstawaniu gatunków
drogą doboru naturalnego, które stało się początkiem
nauki o ewolucji. Zgodnie z teorią Darwina, podstawą
ewolucji organizmów żywych jest dobór naturalny, który
faworyzuje osobniki najlepiej przystosowane do
środowiska bytowania, umożliwiając im nie tylko
przeżycie, ale także przekazanie tych korzystnych
właściwości potomstwu. W ten sposób powstają
organizmy coraz lepiej przystosowane do danego
środowiska.

Kilka lat później – w 1866 roku – Mendel opublikował
skromną pracę pt. Badania nad mieszańcami roślin,
stanowiącą zaczątek genetyki. Była ona syntezą
kilkuletnich obserwacji dziedziczenia siedmiu cech
grochu (Pisum sativum), rośliny samopylnej. Każda z
analizowanych cech miała dwie łatwo odróżnialne
formy: kwiaty czerwone lub białe, nasiona gładkie lub
pomarszczone, nasiona żółte lub zielone itd.
Dokonując krzyżowania między ustalonymi formami
przeciwstawnymi, Mendel uzyskał pierwsze pokolenie
mieszańców (F1), które rozmnażając się w sposób
naturalny (samopylnie) dawało drugie pokolenie
mieszańców (F2). Bardzo skrupulatna analiza cech w
obu pokoleniach dała podstawy do sformułowania
wniosków, że dziedziczenie cech przebiega na drodze
przekazywania zawiązków dziedzicznych (genów) od
rodziców do potomstwa za pośrednictwem gamet,
przy czym jedna gameta zawiera tylko jeden taki
element. Spostrzeżenia te ostatecznie pozwoliły na
sformułowanie praw dziedziczenia. Niestety
doniosłość tego odkrycia nie została doceniona przez
ówcześnie żyjących biologów.
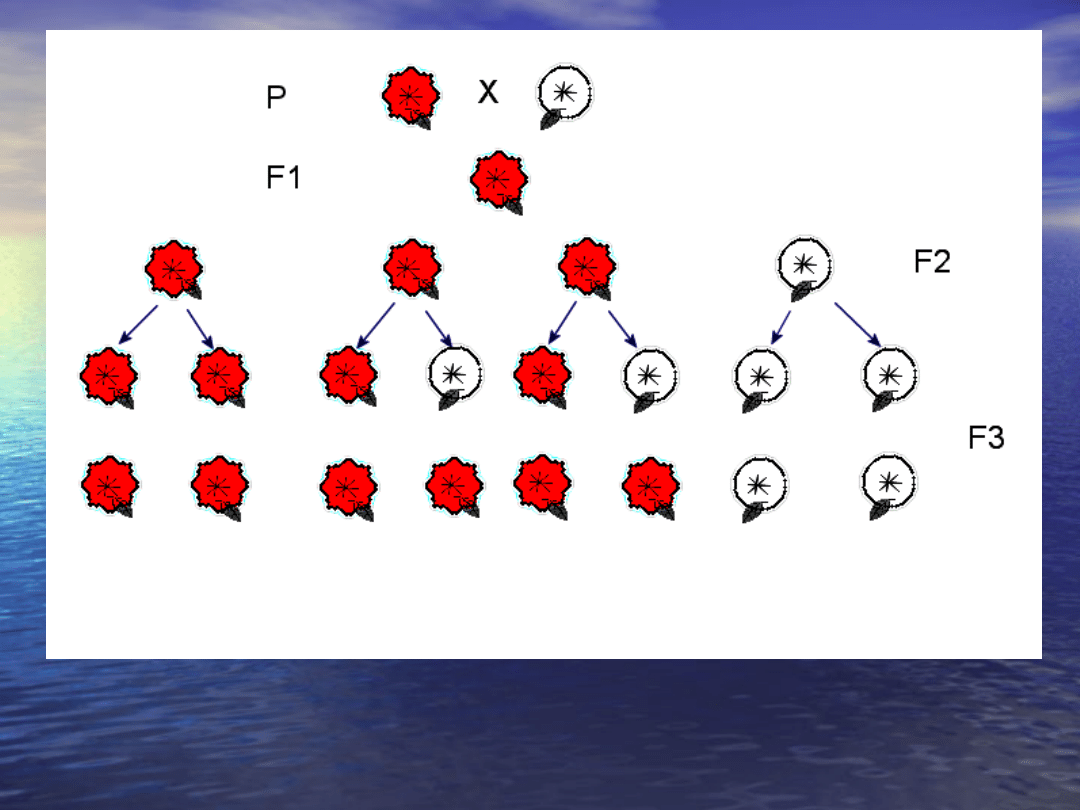
Dziedziczenie barwy kwiatów u
Pisum sativum: P - pokolenie
rodzicielskie,
F1 - pierwsze pokolenie potomne, F2 - drugie pokolenie potomne, F3 - trzecie
pokolenie potomne

Wiek XIX przyniósł także ważne odkrycia dotyczące
budowy komórki, w tym struktur związanych z
dziedziczeniem. Jądro komórkowe zostało po raz pierwszy
opisane przez Browna już w 1831 roku. Pierwsze doniesienia
o „pałeczkowatych” strukturach, nazwanych w 1888 roku
przez Waldeyera chromosomami, zostały podane przez
polskiego botanika Strasburgera w 1880 roku. Podziały jądra
komórkowego zostały opisane pod koniec XIX wieku. Opis
mitozy przedstawił w 1882 roku Fleming, a opis mejozy
powstał w 1883 roku i zawdzięczmy go van Bendenowi i
Boveriemu.
Na początku XX wieku podjęto wiele badań, których
celem byo poznanie praw rządzących dziedziczeniem cech.
Dopiero wówczas zrozumiano genialność odkryć Mendla.
Stało się to za sprawą trzech badaczy: Corrensa, de Vriesa i
Tschermacka, którzy niezależnie od siebie doszli do
wniosków, które kilkadziesiąt lat wcześniej podał Mendel.
Następne lata to lawinowy rozwój nauki nazwanej przez
Batesona w 1906 roku genetyką. W ślad za tym terminem w
1909 roku Johannsen zaproponował określenie „gen” dla
czynnika dziedzicznego warunkującego pojawienie się cechy.
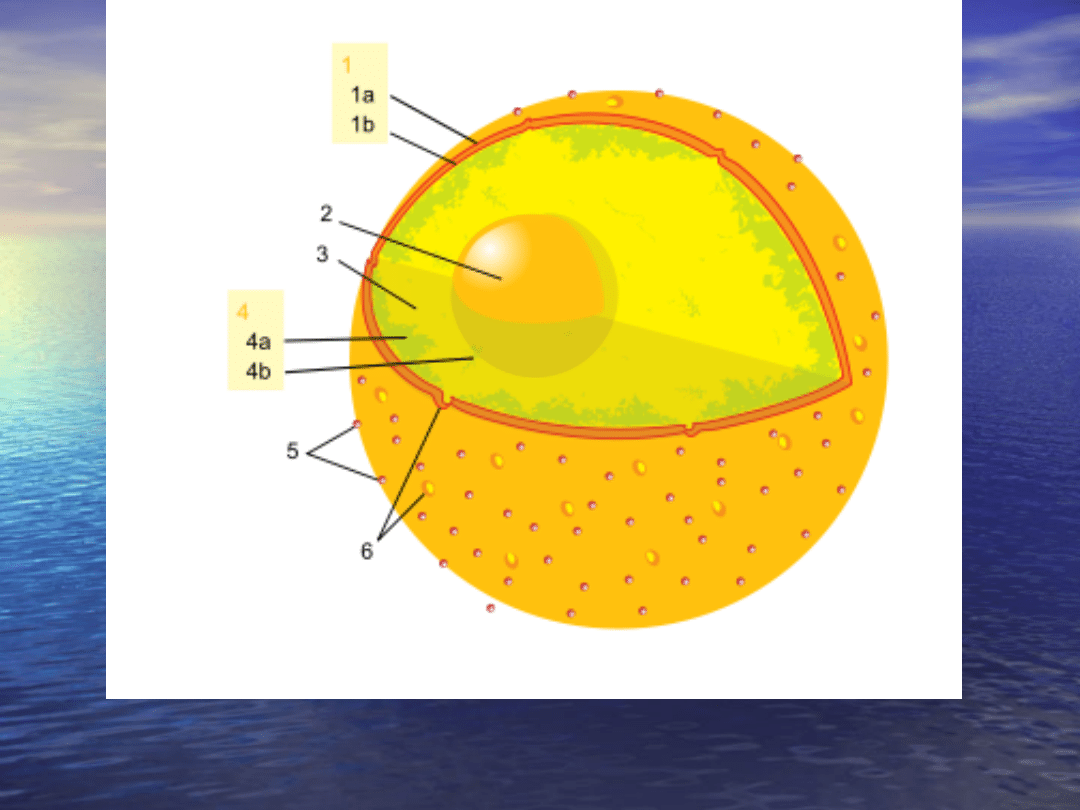
Schemat budowy jądra komórkowego
1 -
, 2 -
, 3 - nukleoplazma, 4a - heterochromatyna,
4b - euchromatyna, 5 - rybosomy, 6 - pory jądrowe
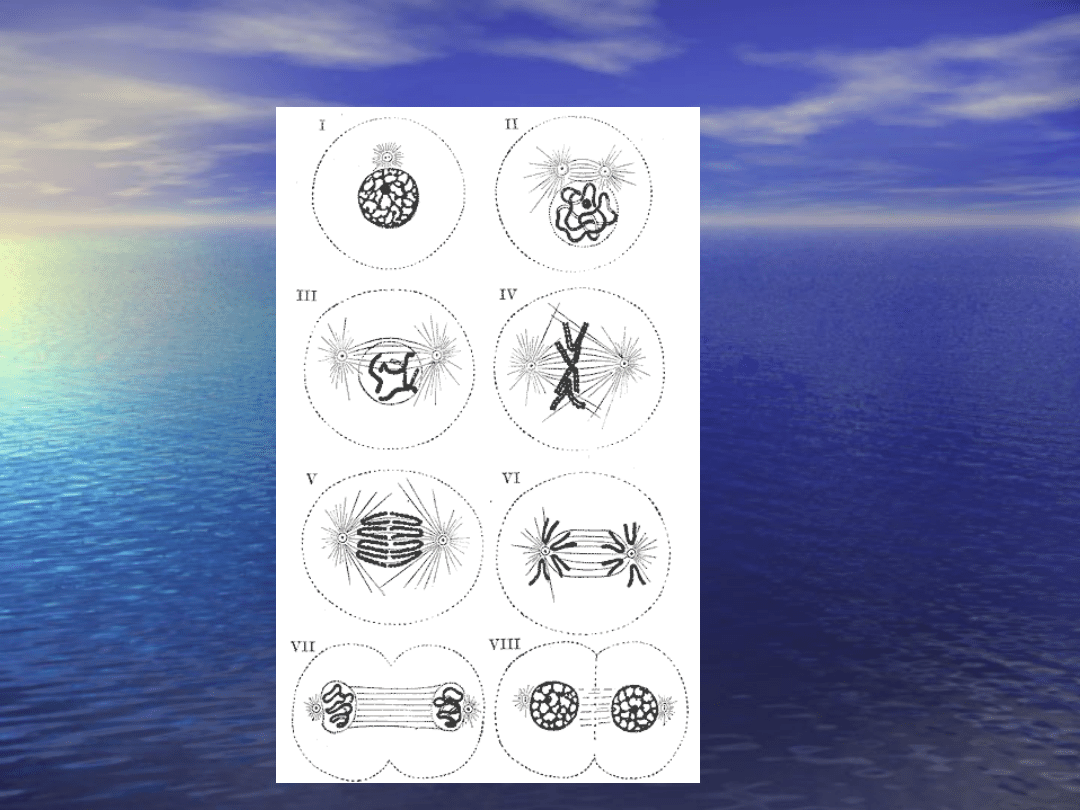
Mitoza I:
II:
III:
IV:
VIII:
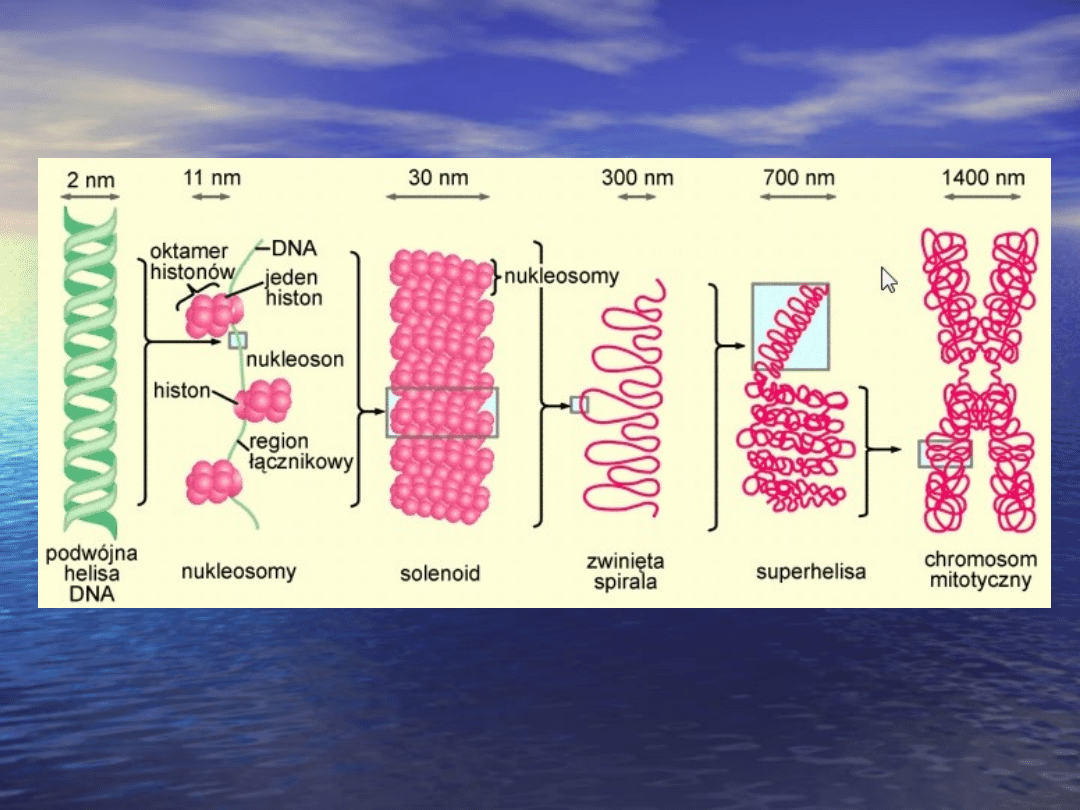
Budowa chromosomu

W 1909 roku Hardy i Weinberg stworzyli podstawy genetyki
populacji. Badacze ci niezależnie od siebie sformułowali
prawo równowagi genetycznej. Rok później Morgan ogłosił
chromosomową teorię dziedziczenia, którą można uważać
za początek cytogenetyki. W latach 20. Fischer, Wright i
Haldane wprowadzili do genetyki metody statystyczne,
które okazały się bardzo istotnymi narzędziami
badawczymi, za pomocą których możliwe stało się
poznawanie mechanizmów odpowiedzialnych za zmienność
i dziedziczenie tzw. cech ilościowych. Niektórzy autorzy
uważają, że zagadnienia genetyki populacji i genetyki cech
ilościowych są na tyle sobie bliskie, że można je określać
jednym terminem – genetyka populacji.
Pomimo ogromnego zaangażowania się wielu
naukowców w badania genetyczne, do końca pierwszej
połowy XX wieku niewiele można było powiedzieć o
chemicznej naturze informacji genetycznej. Przez wiele lat
przypuszczano, ze nośnikiem informacji genetycznej są
białka, o których różnorodności wiedziano już wówczas dość
dużo.

Przełom nastąpił w 1944 roku, kiedy to trzej
badacze: Avery, MacLeod i McCarty wykazali, że
nośnikiem informacji genetycznej jest kwas
dezoksyrybonukleinowy (DNA). Do wniosku tego
doszli na podstawie wyników eksperymentu, w
którym do niechorobotwórczych szczepów bakterii
na drodze transformacji wprowadzili DNA
pochodzący z zabitych bakterii wywołujących
zapalenie płuc u myszy. W wyniku tego bakterie
niechorobotwórcze nabyły cechy zjadliwości. W
doświadczeniu tym można upatrywać początki
genetyki molekularnej. Odkrycie to nie zostało od
razu zaakceptowane przez środowisko biologów.
Początek lat 50. to czas usilnych badań, których
celem było poznanie struktury chemicznej DNA.
Model cząsteczki DNA przedstawili w 1953 roku
Watson i Crick. Po tym odkryciu przyszedł czas
żmudnych prac zmierzających do rozszyfrowania
kodu genetycznego.
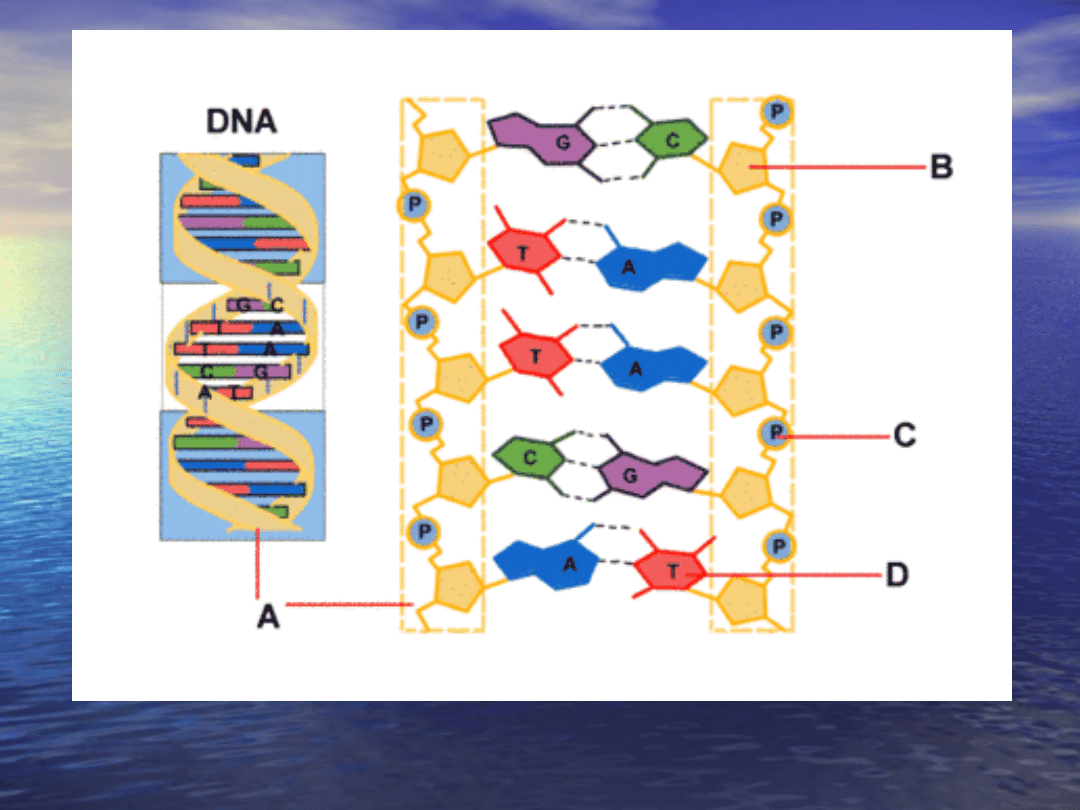
Cząsteczka DNA: A) fosforanowo-węglowodanowy łańcuch główny,
B) węglowodan, C) reszta fosforanowa, D) zasada azotowa

Zostały one zakończone w pierwszej połowie lat 60., a
wiodącą rolę w tych badaniach odegrali między innymi
Khorana, Nirenberg i Ochoa. W tym czasie podjęto również
molekularne badania nad regulacją ekspresji genów.
Doniosłym osiągnięciem stało się ogłoszenie w 1960 roku
przez Jacoba i Monoda tzw. teorii operonu, która
wyjaśniała mechanizm ekspresji genów u bakterii.
Rozwój genetyki molekularnej i związanych z nią
wyrafinowanych metod badawczych umożliwił przejście od
opisu materiału dziedzicznego do prowadzenia na nim
manipulacji i w ten sposób wyodrębniła się nowa
dyscyplina – inżynieria genetyczna. Odkrycie enzymów
restrykcyjnych (1962 r.) oraz enzymu odwrotnej
transkryptazy (1970 r.), opracowanie techniki rekombinacji
i klonowania DNA (1972 r.), sekwencjonowania DNA (1975
r.) czy amplifikacji fragmentów DNA za pomocą
łańcuchowej reakcji polimerazy – PCR (1983 r.) to tylko
najważniejsze przykłady współczesnych narzędzi i metod
badawczych wykorzystywanych obecnie powszechnie w
genetyce molekularnej i inżynierii genetycznej.

Warto podkreślić, że molekularne poznanie genomów
człowieka, zwierząt, roślin, czy mikroorganizmów ma
ogromne znaczenie aplikacyjne związane z diagnostyką i
terapią chorób genetycznych, hodowlą zwierząt i roślin,
wykrywaniem i zwalczaniem patogenów, przetwórstwem
żywności, ochroną środowiska itp. Dążenie do jak
najgłębszego poznania informacji genetycznej zwierząt
gospodarskich doprowadziło w ostatnich latach do
uruchomienia interdyscyplinarnych programów mapowania
genomów takich gatunków, jak świnia, bydło, koń, owca,
kura czy pies. celem tych programów jest wskazanie
położenia genów w chromosomach oraz ustalenia
odległości między genami umiejscowionymi w tym samym
chromosomie. W przypadku człowieka jest nie tylko
mapowanie genomu, ale również jego sekwencjonowanie,
tzn. ustalenie kolejności par nukleotydów. Program ten
został uruchomiony w 1987 roku, a w lutym 2001 roku
ogłoszono na łamach dwóch renomowanych czasopism:
Nature oraz Science zakończenie podstawowego etapu tych
badań.

Warto zaznaczyć, że przedsięwzięcie to przez kilka ostatnich
lat było prowadzone przez dwa niezależne zespoły:
1) międzynarodowe konsorcjum HUGO (ang. Human Genome
Organisation Projekt) oraz
2) prywatną firmę CELERA.
W grudniu 2002 roku ustalono sekwencję genomu myszy, a we
wrześniu 2003 roku ogłoszono wstępną sekwencję genomu
psa. Przewiduje się, że do 2005 roku zostaną
zsekwencjonowane genomy bydła, świni i kury.
Osiągnięcia z zakresu genetyki stały się motorem postępu
nauk biologicznych i ich aplikacji, czego jednym z licznych
przykładów jest hodowla zwierząt. Głównymi metodami
hodowlanymi są selekcja i krzyżowanie, dla których podstawą
teoretyczną jest wiedza z zakresu genetyki cech ilościowych
oraz genetyki populacji. Istotnym elementem pracy
hodowlanej jest kontrola pochodzenia, którą prowadzi się z
wykorzystaniem wiedzy dotyczącej grup krwi i polimorfizmu
białek, a ostatnio także polimorfizmu DNA.

Identyfikacja i eliminacja z populacji hodowlanych nosicieli
mutacji chromosomowych lub genomowych wymaga
zastosowania metod cytogenetycznych, a w przypadku
mutacji genowych – metod genetyki molekularnej.
Wytwarzanie zwierząt mających wprowadzony sztucznie
obcy gen (zwierzęta transgeniczne) związane jest z
zastosowaniem technik inżynierii genetycznej. Ostatnio
duże nadzieje są pokładane w wykorzystaniu map
genomowych do molekularnej identyfikacji genów
mających znaczący wpływ na kształtowanie się cech
użytkowych zwierząt. wykrycie takich genów stworzy nowe
warunki do prowadzenia selekcji, bowiem zamiast oceny
genotypu za pomocą metod statystycznych można będzie
ustalić genotyp osobnika na podstawie badań
molekularnych. Pierwsze osiągnięcia zostały już
opublikowane. Ustalono, jakie mutacje są odpowiedzialne
za hipertrofię mięśniową bydła, wysoką plenność owiec
rasy boorola i inverdale, wysoką mięsność świń czy
wydajność tłuszczu w mleku krów.

Obecny poziom wiedzy genetycznej i możliwości jej
wykorzystania wywołują czasami również niepokój.
Wydaje się, że za postępującym w oszałamiającym
tempie rozwojem genetyki nie nadąża niestety
świadomość niektórych twórców i wielu odbiorców tych
osiągnięć. Możliwość ingerowania metodami inżynierii
genetycznej i komórkowej w informację genetyczną
drobnoustrojów, roślin, zwierząt, a także człowieka
powinna wywołać głęboki zastanowienie się nad tym,
gdzie jest granica, której dla wspólnego dobra
wszystkich organizmów zamieszkujących Ziemię nie
można przekroczyć.
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
Wyszukiwarka
Podobne podstrony:
II. Zarys historycznego kszta towania sie Chin wspo czesnych, współczesne Chiny - Artur Wysocki
Zarys historii
Zarys historii i źródeł teorii krytycznej, Metodologia
zarys historii wychowania
Zarys historii protestantyzmu
Zarys historii procesu beatyfikacyjnego ks, zchomikowane, 35 000 edukacyjnych plików z każdej b
zarys historii psychologii (11 str), ☆♥☆Coś co mnie kręci psychologia
Zarys historii filozofii
1 Zarys historyczny
zarys histori ruchu światło życie - 1, Ruch Światło i Życie
Schuman, LIC I rok, I semestr, Zarys historii integracji europejskiej
Konsp 09 integracja(fin), LIC I rok, I semestr, Zarys historii integracji europejskiej
Rozdział II lastawski- notatki, LIC I rok, I semestr, Zarys historii integracji europejskiej
Cichocka H, Lichański J Zarys historii retoryki Volkmann R Wprowadzenie do retoryki Greków i Rzymi
ZARYS HISTORII PRZEMIAN TURYSTYKI W POLSCE AŻ DO DNIA DZISIEJSZEGO
więcej podobnych podstron