
Narząd Słuchu
Dariusz Nowak
Zakład Fizjologii
Klinicznej, Łódź
Mazowiecka 6/8

Słuch
• Odbieranie fal głosowych przez ucho
• Rozróżnianie częstotliwości i głośności
• Przewodzenie do OUN i dekodowanie
• Błona bębenkowa i układ kosteczek
• Młoteczek (malleus)
• Kowadełko (incus)
• Strzemiączko (stapes)
• Okienko owalne
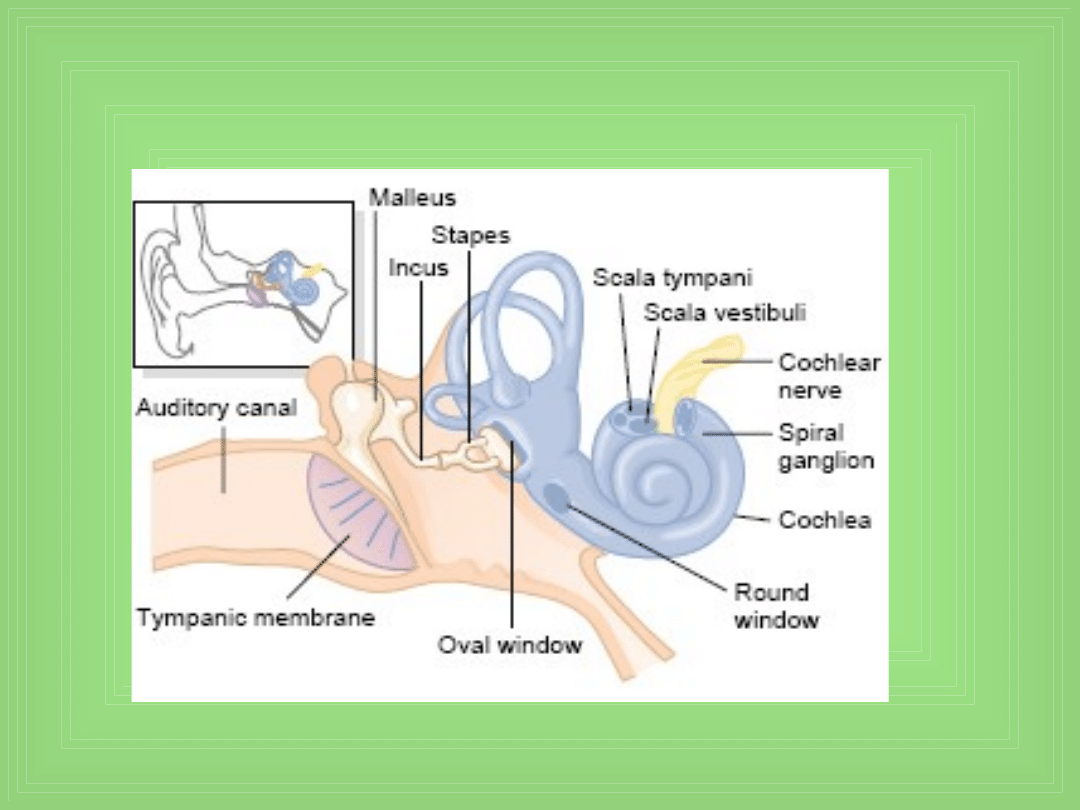
Tympanic membrane, ossicular system of the
middle ear, and
inner ear.
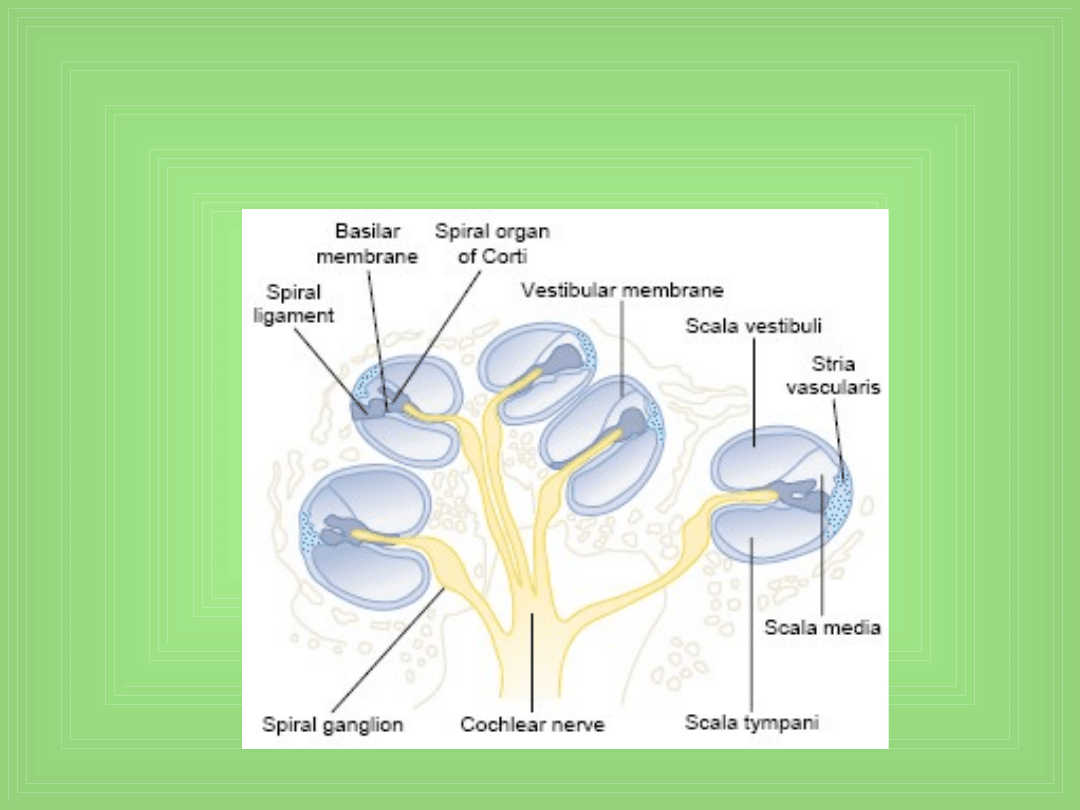
Cochlea

Układ kosteczek
• Punkt błony bębenkowej do którego jest
przymocowany koniec rękojeści młoteczka jest
stale pociągany przez „tensor tympani muscle” →
utrzymuje napięcie błony bębenkowej → pozwala to
przewodzić drgania akustyczne do kosteczek przez
błonę
• Więzadła pomiędzy kosteczkami →powodują że
młoteczek i kowadełko działają jak jedna dźwignia,
punkt podparcia jest na granicy błony bębenkowej.
• Połączenie przegubowe pomiędzy kowadełkiem a
strzemiączkiem popycha okienko owalne i płyn
ślimaka gdy rusza się błona bębenkowa

Dopasowanie impedancji (oporności akustycznej)
przez układ kosteczek
• Amplituda ruchu strzemiączka powodowana
drganiami akustycznymi stanowi jedynie ¾
amplitudy rękojeści młoteczka ( a wydawać by się
mogło że dźwignia zwiększa amplitudę !?)
• W rzeczywistości układ kosteczek redukuje zakres
ruchu ale zwiększa około 1.3-raza siłę tego ruchu
• Dodatkowo S błony bębenkowej = 55 mm
2
, S
strzemiączka = 3.2 mm
2
( iloraz = 17)
• 17 x 1.3 = 22 jest to wzmocnienie siły
wzbudzającej płyn w ślimaku , jest to potrzebne bo
ma on dużo większą bezwładność niż powietrze

Dopasowanie impedancji (oporności
akustycznej) przez układ kosteczek
• Układ kosteczek dopasowuje oporność akustyczną
i siły pomiędzy falami dźwiękowymi w powietrzu a
drganiami w płynie ślimaka
• Dopasowanie osiąga 50-75% doskonałości dla
częstotliwości f od 300 do 3000 Hz → powoduje to
wykorzystanie większości energii wpadających do
ucha fal dźwiękowych
• Brak błony bębenkowej i kosteczek → fale
akustyczne idą powietrzem i mogą wejść do
ślimaka przez okienko owalne, jednakże spada
czułość ucha o około 15 – 20 dbeli (zaledwie
słyszymy)

Osłabienie dźwięku
•
Realizowane poprzez skurcz mięśnia tensor tympani i
strzemiączkowego
•
Gdy ogłuszający dźwięk jest przewodzony przez układ
kosteczek a później do OUN → po ok. 40-80 ms
występuje odruch powodujący skurcz mięśnia
strzemiączkowego i w mniejszym stopniu napinacza bł.
bębenkowej.
•
Siły obu mięsni ciągną układ kosteczek w przeciwne
strony → rośnie sztywność układu kosteczek → redukcja
przewodzenia dźwięków zwłaszcza o f< 1000 Hz
•
Odruch ten zmniejsza przewodzenie o 30-40 dbeli
•
Ogłuszający dźwięk zamienia się w szept

Osłabienie dźwięku
• Znaczenie tego odruchu
• 1. Ochrona ślimaka przed nadmiernymi wibracjami
• 2. Wytłumienie dźwięków o niższej częstotliwości
w hałaśliwym otoczeniu wygasza tło i pozwala się
skupić na dźwiękach o f > 1000 Hz (one niosą więcej
istotnych dla człowieka informacji)
• 3. Zmniejszenie czułości słuchu na własną mowę
w momencie gdy mówimy ten odruch jest
aktywowany przez oboczne sygnały z kory mózgowej
przewodzone do tych mięsni w momencie gdy mózg
aktywuje mechanizmy emisji głosu

Przewodzenie kostne dźwięków
• Ślimak jest otoczony jamą kostną (w kości
skroniowej) – labirynt kostny. → drgania całej
czaszki powodują wibracje płynu w ślimaku
• Wyniosłość czaszki, wyrostek sutkowaty
przystawić drgający kamerton – słychać dźwięk
• W normalnych warunkach to przewodzenie nie
jest w stanie zastąpić przewodzenia przez układ
kosteczek słuchowych
• Wzmacniacze przewodnictwa kostnego

Ślimak (Cochlea)
• Trzy spiralne kanały
Scala vestibuli
Scala media
Scala tympani
• Vestibular membrane (Reissner’s membrane)– oddziela Sv.
od Sm.
• Basilar membrane – oddziela Sm. od St. – na jej powierzchni
jest organ Corti’ego – zawiera serie elektromechanicznie
czułych komórek – Hair cells
• Hair cells są to odbiorcze (końcowe) organy, generują
impulsy nerwowe w odpowiedzi na drgania akustyczne

Ślimak (Cochlea)
• Vestibular membrane jest bardzo cienka i nie
przeszkadza wcale w rozchodzeniu się wibracji
akustycznych
• Przy rozważaniu przewodzenia dźwięku można
uznać że jej nie ma i Sm i Sv nie są oddzielone ,
są jedna komorą.
• Sama błona ma znaczenie dla utrzymania
specjalnego składu płynu w Sm – niezbędnego
dla funkcji hair cells
• Drgania akustyczne→ strzemiączko→ okienko
owalne→ ruch płynu (działa w obie strony)
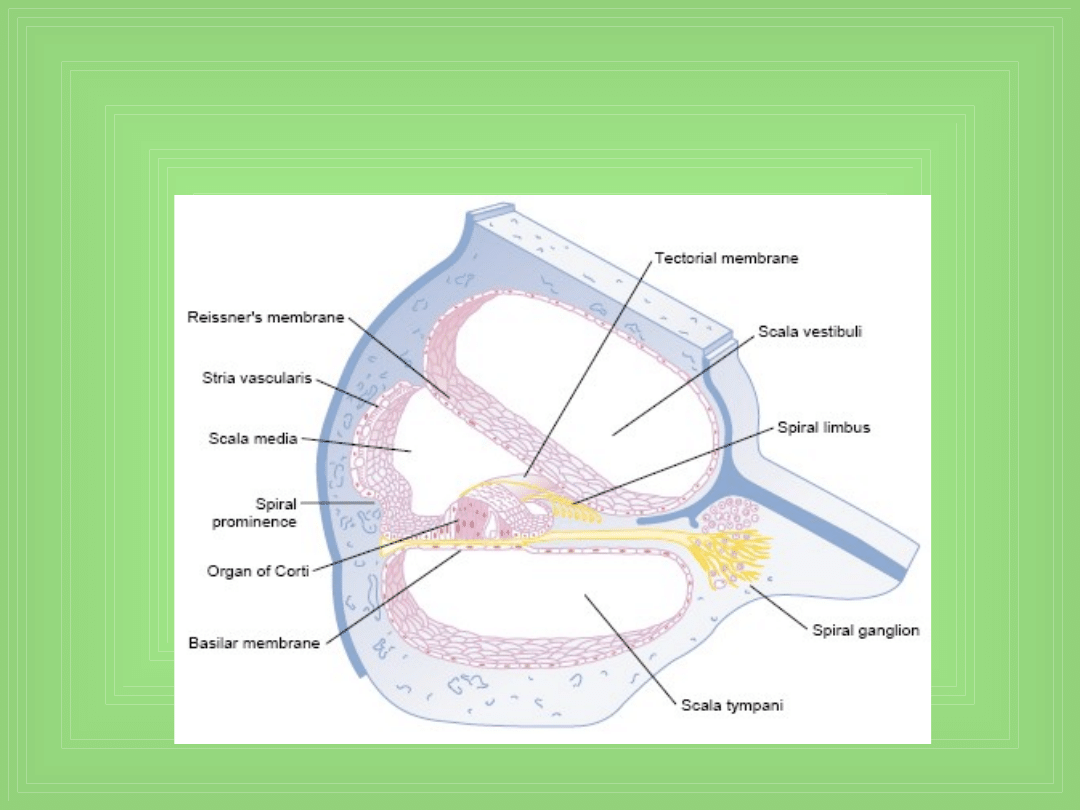
Section through one of the turns
of the cochlea.
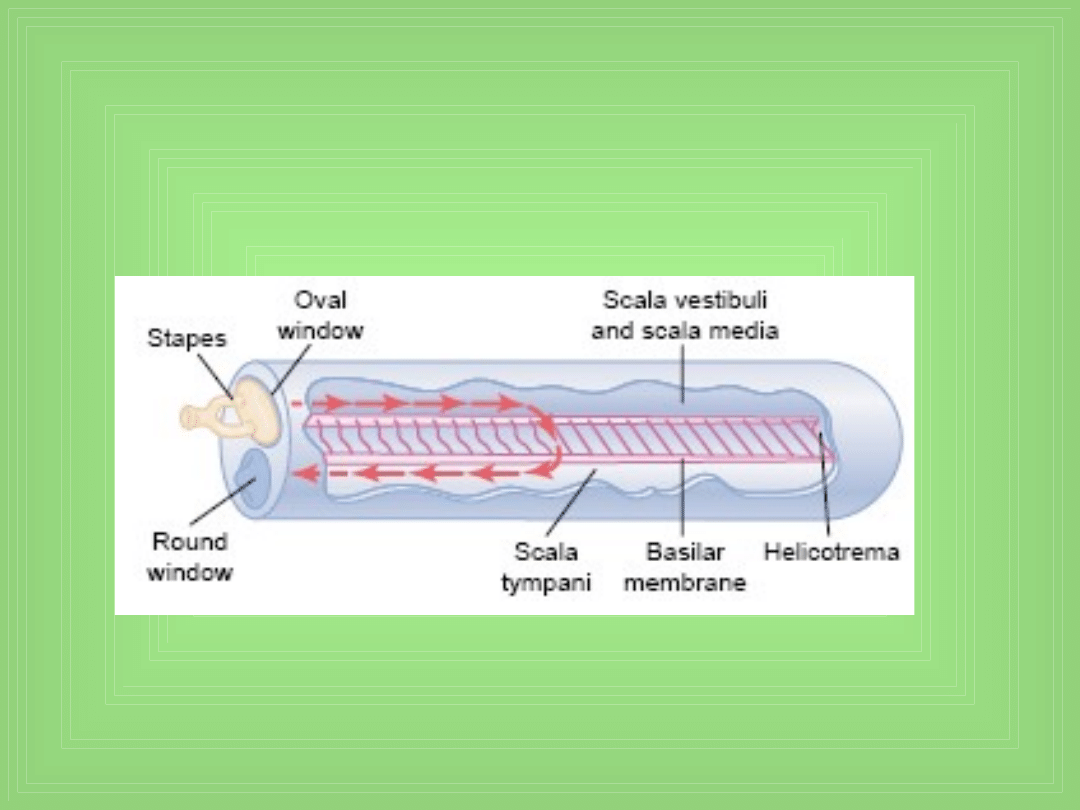
Movement of fluid in the cochlea after
forward thrust of the stapes

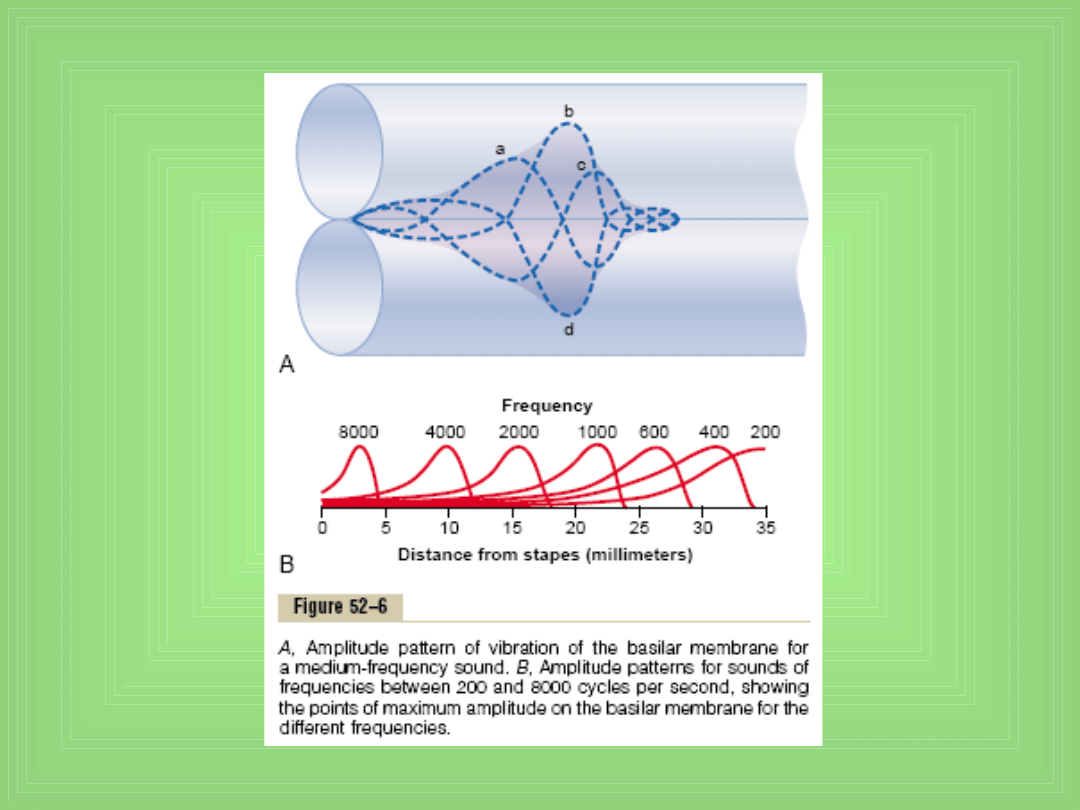
Basilar membrane i rezonans w ślimaku
• Basilar membrane jest włóknistą błoną która oddziela
Sm od St
• Jest w niej 20 000 do 30 000 włókien podstawnych, są
one sztywne, elastyczne i „sprężyste jak sucha trzcina”
• Są umocowane w centrum kostnym a drugi koniec
wolny – mogą drgać jak stroik w harmonijce ustnej
• Długość tych włókien rośnie progresywnie od okienka
owalnego do szczytu ślimaka : od 0.04 mm do 0.5 mm
(12 razy) a ich średnica maleje od okienka do szczytu
ok. 100 razy
• Przy okienku dobrze wibrują przy bardzo wysokich f
(rezonans wysokich częstotliwości) a przy szczycie
przy b. małych f ( rezonans niskich częstotliwości)

Transmisja fal dźwiękowych w ślimaku
• Rola „round window”
• Wędrująca fala
• Wzór wibracji basilar membrane dla różnych f dźwięku
• Każda fala jest słaba na początku , ale staje się silna
gdy osiągnie część błony o tej częstotliwości
rezonansowej co dany dźwięk (który tę falę
spowodował) – i w tym miejscu energia fali jest
rozpraszana i dalej ona nie idzie ( w tym miejscu ginie,
kończy się)
• Czyli dźwięk o wysokim f – krótka droga, trafia na
miejsce rezonansu i ginie
• Niska f – dalej idzie – rezonans i ginie !!!

Transmisja fal dźwiękowych w
ślimaku
• Zmienia się również szybkość wędrówki fali po
błonie bo zmienia się elastyczność błony (spada w
kierunku do szczytu ślimaka)
• Fala na początku membrany dużo szybciej się
przemieszcza – znaczenie – pozwala rozróżnić
kilka dźwięków o wysokiej f
• Rezonans daje podstawę do rozróżnienia
częstotliwości dźwięków- miejsce wystąpienia
rezonansu jest jednocześnie miejscem
maksymalnej stymulacji włókien nerwowych
narządu Corti’ego który leży na basilar membrane
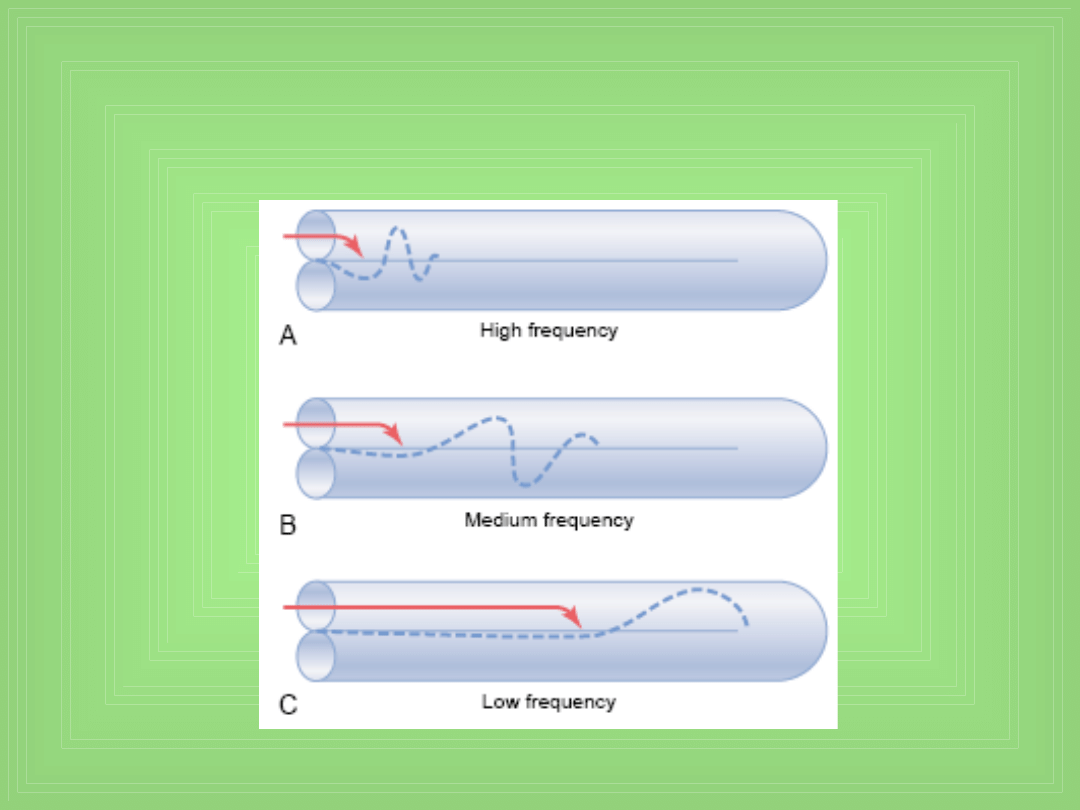
“Traveling waves” along the basilar
membrane for high-,medium-, and low-
frequency sounds.

Funkcja narządu Corti’ego
• Receptorowy narząd który generuje impulsy nerwowe
w odpowiedzi na wibracje basilar membrane, leży na
jej powierzchni i na jej włóknach
• Właściwym receptorem są 2 typy wyspecjalizowanych
komórek nerwowych - hair cells: pojedynczy rząd
internal (inner) komórek – n= 3500 (Ф 12 µm) oraz 3
lub 4 rzędy external (outer) hair cells – n= 12000 (Ф 8
µm)
• Tworzą synapsy z siecią zakończeń nerwu ślimaka, 90-
95% tych zakończeń idzie do inner cells
• Pobudzenie idzie do zwoju spiralnego narządu
Corti’ego który leży w centrum ślimaka, komórki
zwoju ślą aksony (ok. 30 000) do nerwu ślimaka →
OUN
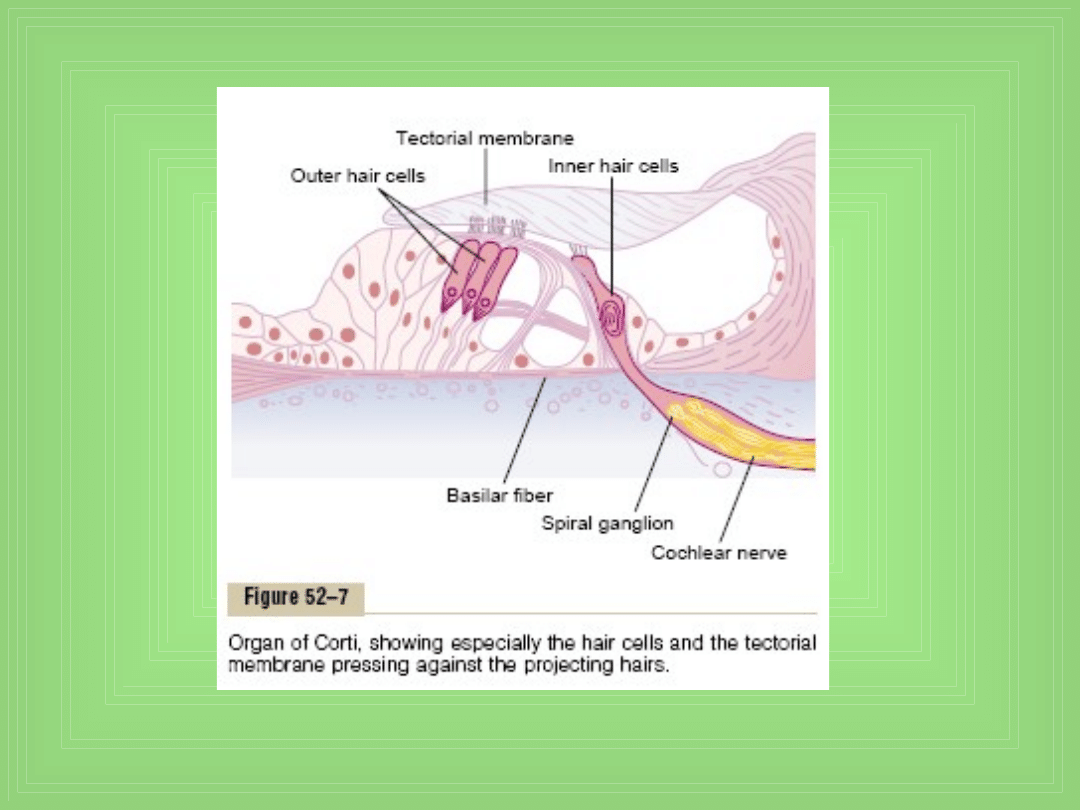
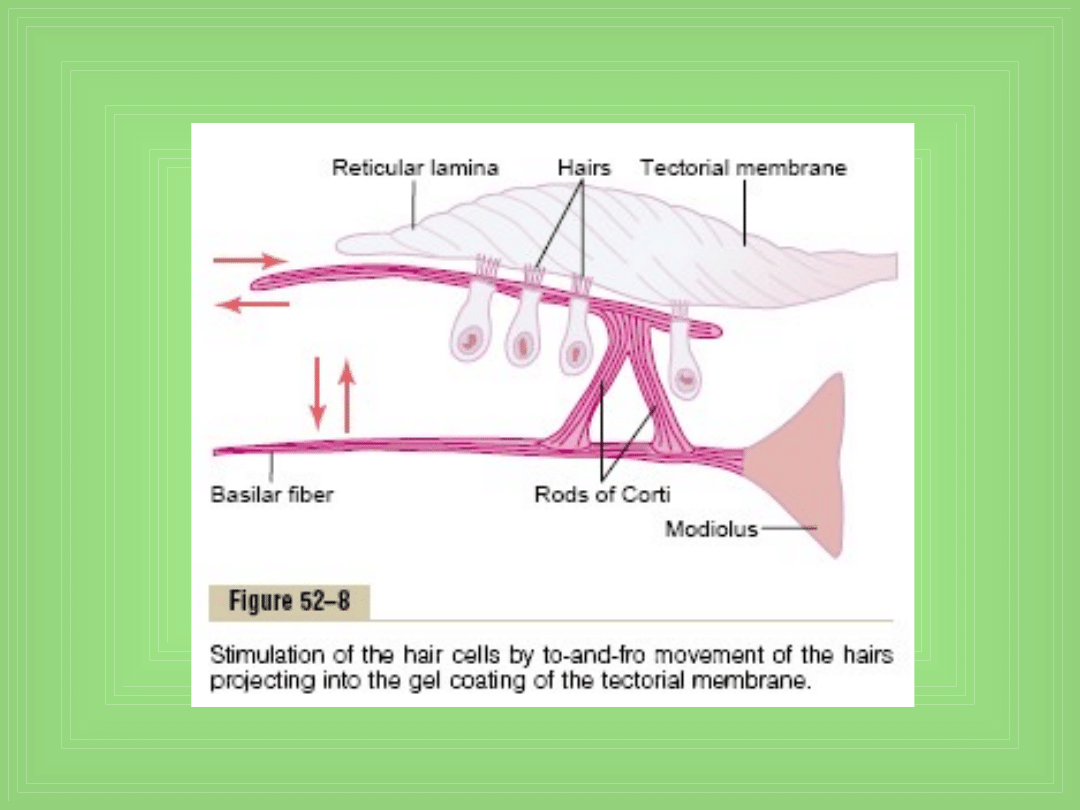

Pobudzenie hair cells
• Małe włoski (stereocilia) wystają z komórek i dotykają lub
są otoczone przez powierzchnię żelu pokrywającego
„tectorial membrane”, która leży nad stereociliami w S.
media
• Zakręcenie włosków w jedna stronę – depolaryzacja, a w
drugą stronę ich hiperpolaryzacja → pobudzenie włókien
nerwu słuchowego które łączą się synapsą z komórkami
• W jaki sposób wibracje basilar membrane pobudzają
włoski ? – zewnętrzne końce komórek są w sztywnej
„reticular lamina”, podpartej trójkątnymi pręcikami
Corti’ego, które są przymocowane do basilar fibers →
wszystkie te struktury ruszają się jak jedna sztywna
jednostka

Pobudzenie hair cells
• Ruch do przodu włókien podstawnych (b. fibers)
kołysze reticular lamina w kierunku do środka
ślimaka (i odwrotnie) → to powoduje że włoski trą o
tectorial membrane i komórki się pobudzają
• Sygnały słuchowe są głównie generowane przez
inner hair cells , jest ich mniej a pobudzają ok. 90%
zakończeń nerwu słuchowego
• Ale jeśli outer cells są zniszczone a inner pracują
dobrze to mimo wszystko jest b. duża utrata
słuchu !!!
• Outer cells regulują czułość inner cells przy różnych
pozycjach dźwięku (są specjalne gałązki nerwowe) –
jest to tuning – strojenie układu receptorowego

hair cells
• Na jednej hair cell jest 100 stereocilii – ich ułożenie ,
długość jest uporządkowane – zapewnia odpowiednie
pobudzenie
• Sygnał mechaniczny → otwarcie kanałów , napływ K+
→ depolaryzacja → synapsa (jest neurotransmiter,
prawdopodobnie glutamina)
• S.m. – jest endolimfa (dużo K+ a mało Na+)
• S.t. i S.v. jest w nich perilimfa (mało K+, dużo Na+)
• Dlatego jest stała różnica potencjałów między endo- a
perilimfą = + 80 mV – Potencjał wewnątrzślimakowy.
• Włoski są w endolimfie – to uczula dodatkowo
komórki, zwiększa czułość a słabe dźwięki

Rozróżnianie częstotliwości dźwięków
• „ place principle” – wykrywamy miejsce na basilar
membrane które jest najbardziej stymulowane
• Ale jak rozróżnić f= 200 Hz od f=20 Hz skoro
powstają na końcu prawie w tym samym
miejscu ?
• „Volley- frequency principle” – dźwięki o f od 20
do 2000 Hz powodują salwy impulsów nerwowych
o tej samej częstotliwości co wywołujący je
dźwięk

Jak rozróżniamy głośność ?
• Przynajmniej 3 sposobami
• 1. większa głośność → większa energia → większe
wibracje→ większe pobudzenie hair cells
→szybsze pobudzenie zakończeń nerwowych
• 2. większe wibracja →więcej w tym miejscu
pobudzonych hair cells (sumowanie
przestrzenne) → więcej zakończeń nerwowych
pobudzonych
• 3. silna wibracja pobudza także outer cells i to
daje sygnał że dźwięk jest głośny

Głośność
• Jaki przedział głośności rozróżnia ucho ?
• Najsłabszy szept do - najgłośniejszy dźwięk
• Między nimi różnice w natężeniu 1 bilion razy
• Różnice w amplitudzie drgań basilar membrane –
1 milion razy
• Ale ucho (wrażenia odczuwane) spłaszcza te
różnice do 10 000 razy

decybele
• Wartości wyrażane w decybelach odnoszą się do stosunku
dwóch wielkości: P do wielkości P
o
odniesienia
• p
dB
= 10 log
10
(P/P
o
)
• p
dB
- wielkość P w decybelach, log
10
- logarytm dziesiętny,
P
o
- wielkość odniesienia
• P
o
= 1
• P
1
= 10 p
1
= 10dB
• P
2
= 100 p
2
= 20dB
• P
3
= 1000 p
3
= 30 dB
• P
4
= 10000 p
4
= 40 dB
• Jednostka natężenia dźwięku . Ucho zaledwie rozróżnia
zmianę o 1 dB ( zmiana energii o 1.26 raza)
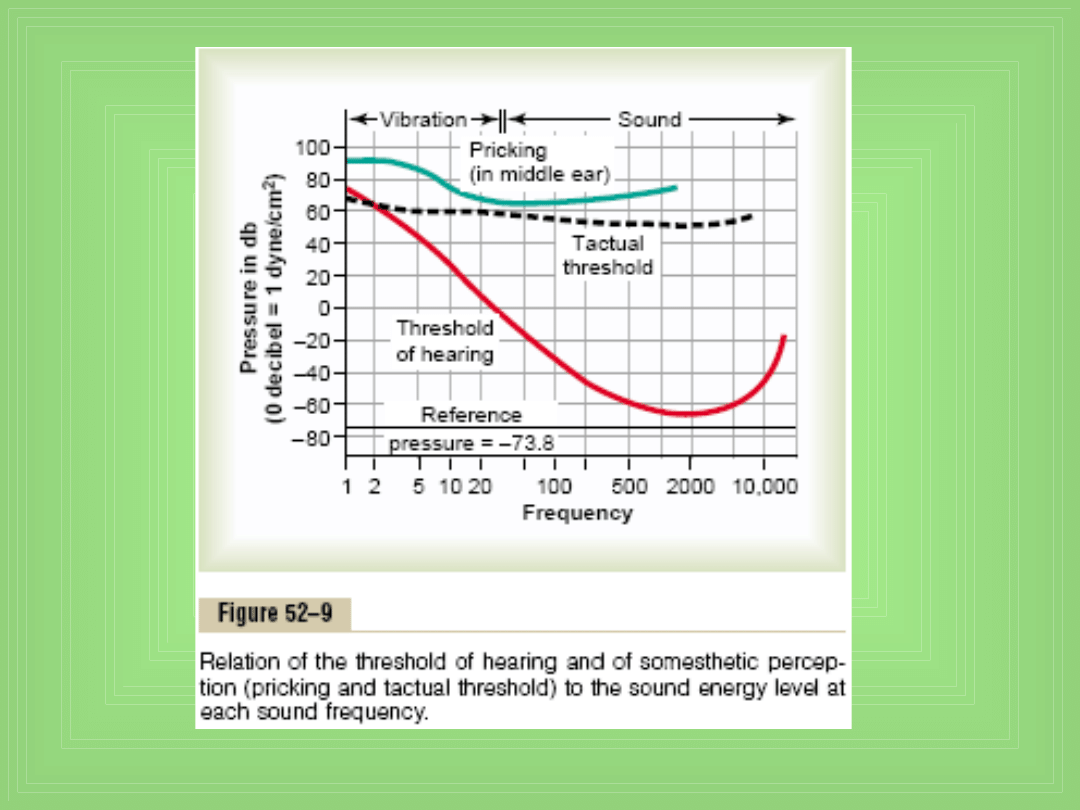

Centralny mechanizm słuchu
• Spiral ganglion of Corti→ brzuszne i grzbietowe
jądro ślimaka (górna część rdzenia) (synapsy) →
drugo-rzędowe neurony (przechodzą głównie na
drugą stronę pnia mózgu (brain stem) ale też idą
po tej samej stronie → superior olivary nucleus
…….→ promienistość słuchowa → kora słuchowa
(górny zakręt płata skroniowego)
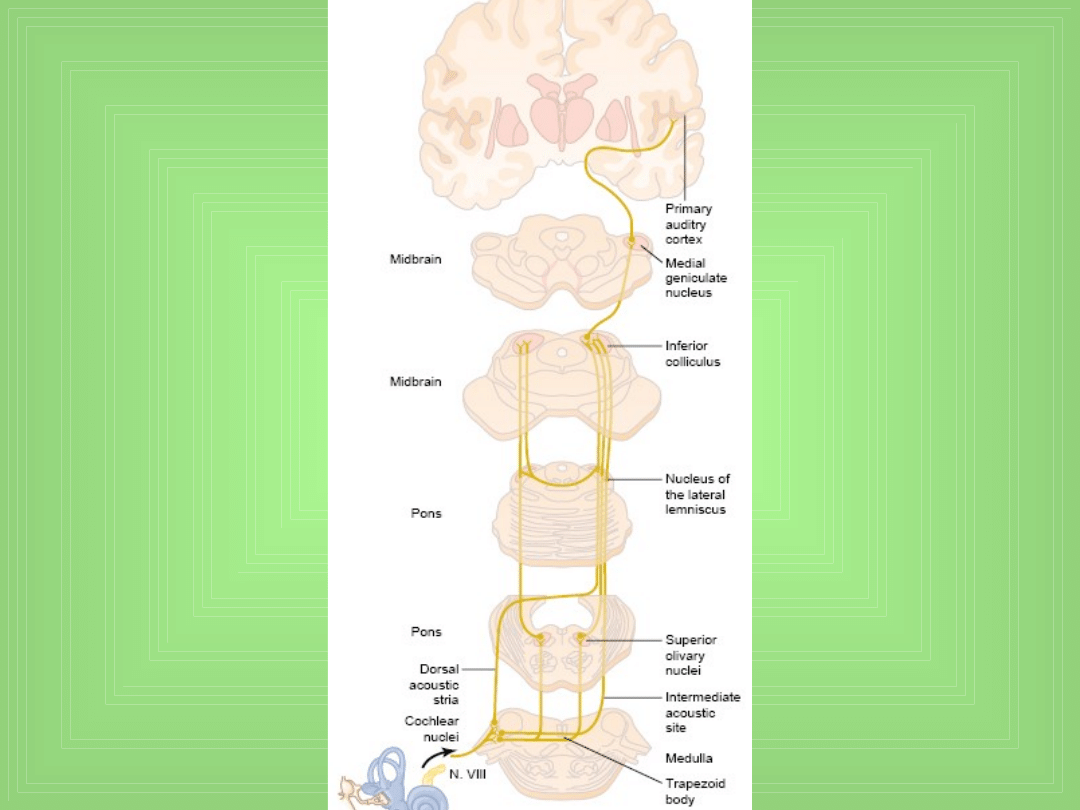

Przewodzenie do OUN
• 1. Z obu uszu impulsy idą po obu stronach z przewagą
po stronie przeciwnej
• 2. Są 3 miejsca skrzyżowania dróg w pniu mózgu
• 3. Wiele obocznych włókien z dróg słuchowych idzie
bezpośrednio do części pobudzającej tworu
siatkowatego pnia mózgu – z tego systemu idą sygnały
w górę do pnia i w dół do rdzenia kręgowego i
aktywują cały układ nerwowy w odpowiedzi na b. głośny
dźwięk.
• Częstotliwości wyładowań w drogach słuchowych
zmieniają się niezależnie od dźwięku – dla tego samego
dźwięku f wyładowań zmieniają się na kolejnych
etapach przewodzenia
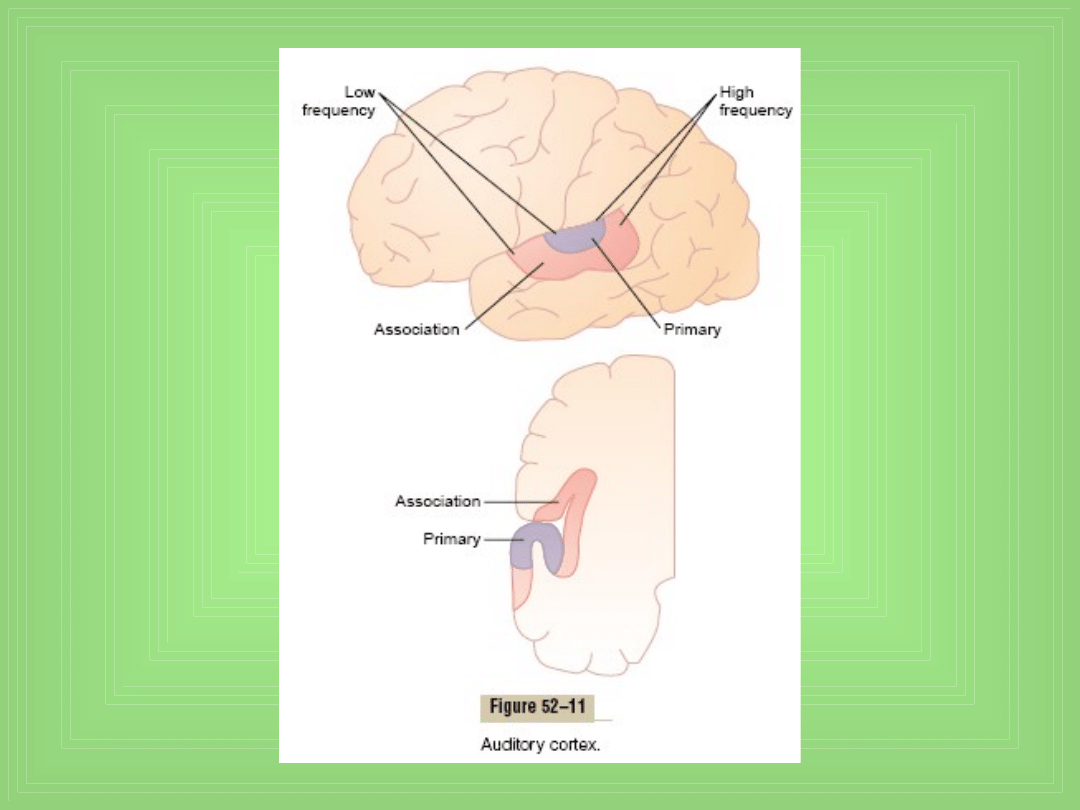

Kora słuchowa
• Mapy tonowe w korze słuchowej – dla rozróżniania
różnych dźwięków (f)
• Obustronne uszkodzenie pierwszorzędowych pól
korowych słuchowych – bardzo redukuje wrażliwość na
dźwięk (słuch)
• Uszkodzenie po 1 stronie – słabo redukuje słuch po
przeciwnej stronie – dlaczego ? , ale upośledza
zdolności lokalizacji źródła dźwięku
• Uszkodzenie kojarzeniowych pól słuchowych nie
upośledza zdolności słuchu, rozróżniania tonów i
interpretowania nawet prostych wzorów dźwiękowych .
Ale nie mogą interpretować znaczenia słyszanych
dźwięków (słyszy słowa, może je powtórzyć, ale ich nie
rozumie)

Jak rozróżniamy kierunek z którego dochodzi dźwięk
?
• 2 mechanizmy (poziomy kierunek)
• 1. rozróżnienie różnic w czasie dojścia dźwięku do 1 ucha i
do ucha po przeciwnej stronie
• Najlepiej działa dla dźwięków o F < 3000 Hz
• 2. Różnica w intensywności między dźwiękami
dochodzącymi do uszu
• Lepiej działa dla f > 3000 Hz - głowa jest lepszą barierą dla
tych dźwięków
• Mechanizm 1 jest bardziej precyzyjny od 2
• Ale bez małżowiny usznej to nie działa !!!

Małżowina uszna
• Jest to lejek zbiera dźwięk – wzmocnienie i
skierowanie do przewodu słuchowego
• Przeprowadza spektralną transformację co
pozwala na lokalizację dźwięku
• Odbicia od struktur małżowiny – proces filtracji
dźwięku, modulacja dźwięku zależna od
amplitudy – zapewnia rozróżnianie kierunku
• Ludzka małżowina robi to najlepiej dla dźwięków
o f zbliżonych dla f naszej mowy
• U zwierząt jest także promiennikiem ciepła i
sposobem sygnalizacji

Lis pustynny Fenek

Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
- Slide 38
- Slide 39
Wyszukiwarka
Podobne podstrony:
2. Anatomia narządu słuchu, Anatomia i fizjologia, anatomia
dziecko z dysfunkcją narządu słuchu
Narzad sluchu
Narząd słuchu
LIST 2 Anatomia, fizjologia i patologia narządu słuchu
Wykład 13 narządy słuchu, Lekarski, I, PIERWSZY ROK MEDYCYNA MATERIAŁY, HISTOLOGIA
NARZĄD SŁUCHU, JĘZYK MIGOWY(1)
Algorytm - Ocena narządu słuchu, PIELĘGNIARSTWO 1 sem, Podstawy Pielęgniarstwa, laborka
NARZĄD SŁUCHU I RÓWNOWAGI, weterynaria, anatomia
Biofizyka narządu słuchu, Fizjoterapia, Biofizyka
Biofizyka narządu słuchu i wzroku
FIZYCZNE MECHANIZMY DZIAŁANIA NARZĄDU SŁUCHU
Budowa i dzialanie narzadu sluchu
Budowa narządu słuchu
fizyka, Hałas, Każda fala dźwiękowa może być wykryta przez ucho, w narządzie słuchu przetworzone na
Narząd słuchu i równowagi klasa 2 karta pracy
13 NARZAD SLUCHU I ROWNOWAGIid Nieznany (2)
NARZĄD SŁUCHU
dziecko z dysfunkcją narządu słuchu
więcej podobnych podstron