

Wyróżnia się trzy typy
błonowych receptorów
histaminowych, sprzężonych z
białkiem regulatorowym G:
postsynaptyczne H
1
postsynaptyczne H
2
presynaptyczne H
3

W budowie receptorów H
1
obserwuje się różnice
gatunkowe. W zależności od gatunku receptory
H
1
zbudowane są z różnej liczby aminokwasów
np. u człowieka z 487 aminokwasów, u małpy –
488, wołu – 491 aminokwasów.
Receptory H
1
mają szeroką trzecią
wewnątrzkomórkową pętlę i krótki łańcuch C –
terminalny. Jest to charakterystyczne dla
receptorów mobilizujących jony Ca
2+
i
receptorów hamujących cyklazę adenylową tj.
np. receptory M
2
– muskarynowe. Potwierdza to
fakt, że antagoniści receptorów H
1
wykazują
często aktywność antymuskarynową, ale nie
działają antagonistycznie w stosunku do
receptorów H
2
.
Wiązanie dwusiarczkowe stabilizujące układ
tworzą cząsteczki cysteiny 101 i 181, łączące
1 i 2 pętlę zewnątrzkomórkową.
POSTSYNAPTYCZNY
RECEPTOR
H
1

Homologia pośród różnych receptorów H
1
,
ogólnie przekracza 60%.
Między receptorami H
1
i H
2
homologia jest
rzędu 40%. Pomimo tego przypuszczalnym
miejscem wiązania histaminy przez oba
receptory jest kwas asparaginowy trzeciej
domeny. Ponadto w wiązaniu z histaminą
uczestniczą treonina i asparagina piątej
domeny.
W przypadku receptora H
1
w wiazaniu
histaminy bierze udział:
kwas asparaginowy 108 trzeciej
domeny
kwas asparaginowy 199 piątej domeny
treonina 195 i 204 piątej domeny

Związanie agonisty z receptorami H
1
,
zmienia konformację białka
receptorowego i w konsekwencji
aktywuje białko G(sprzężone z
receptorem w obszarze 2 i 3 pętli
wewnątrzkomórkowej). Białko G
przekazuje sygnał z receptora na układ
enzymatyczny, którym jest fosfolipaza
C (PLC), aktywując ją. Aktywna postać
PLC katalizuje hydrolizę PhIP
2
do
inozynotrifosforanu (IP
3
) i
diacyloglicerolu (DG).
W konsekwencji wzrost stężenia IP
3
wywołuje:
skurcz mięśni gładkich
wzmożona przepuszczalność
naczyń; obrzęki
wzmożone wydzielanie śluzu w
drogach oddechowych
spadek ciśnienia krwi

IP
3
reaguje ze sprzężonymi receptorami (IP
3r
)
w błonie wewnątrzkomórkowych magazynów
wapnia powodując uwolnienie jonów Ca
2+
do
cytozolu. Uwolnione jony Ca
2+
reagują z
kalmoduliną tworząc razem kompleks, który
pośrednio prowdzi do utworzenia cGMP
(wtórny przenośnik wewnątrzkomórkowych
zmian) zGTP.
Z drugiej strony jony Ca
2+
wywołują
odpowiedź glikogeniczną, która może być
wyjaśniona aktywacją fosforylazy.
Decydującą rolę w przekazywaniu sygnału
ma proteinokanaza C (PKC), która
reguluje różne funkcje i rozwój komórki.

Diacyloglicerol (DG) poprzez ujemne
sprzężenie zwrotne hamuje
fosfolipazę C i prowadzi do
zmniejszenia produkcji IP
3
oraz
redukcji mobilizacji Ca
2+
.
Ponadto ze stymulacją receptora H
1
związany jest napływ
zewnątrzkomórkowych jonów Ca
2+
w wyniku otwarcia kanałów
wapniowych, zależnych od
potencjału i kanałów wapniowych
modulowanych receptorem.

Receptor H jest polipeptydem zbudowanym
z 359 aminokwasów. Jedną transbłonową
hydrofobową -helisę (łańcuch N –
terminalny) tworzy 22 – 25 aminokwasów.
Wiązanie dwusiarczkowe stabilizujące
układ tworzą cząsteczki cysteiny 91 i 174,
łącząc 1 i 2 pętlę zewnątrzkomórkową.
W wiązaniu z histaminą uczestniczy:
kwas asparaginowy 98 trzeciej
domeny
kwas asparaginowy 186 piątej
domeny
treonina 190 piątej domeny
POSTSYNAPTYCZNY
RECEPTOR H
2

Związanie histaminy z receptorem H
2
–
histaminowym powoduje kolejno: zmianę
konformacji białka receptorowego, wskutek
czego zwiększa się jego powinowactwo do
białka G
s
, aktywację cyklazy adenylowej i w
konsekwencji wzrost stężenia cAMP,
pełniącego rolę przekaźnika II rzędu.
cAMP jest odpowiedzialny za aktywację H
+
/K
+
-
ATPazy w cytozolu komórek okładzinowych
żołądka.
W efekcie wzrostu stężenia cAMP
występuje:
zwiększenie wydzielania kwaśnego soku
żołądkowego
zwiększenie sekrecji śluzu w drogach
oddechowych i w żołądku
wzrost częstotliwości i kurczliwości
serca
Selektywnym agonistą receptora H
1
jest
2-(2-pirydylo)etyloamina,
natomiast receptora H
2
dimaprit (2-[3-
(dimetyloamino)propylo]-2-
tiopseudomocznik).
Histamina pobudza zarówno receptory H
1
jak i H
2
, co tłumaczy się możliwością
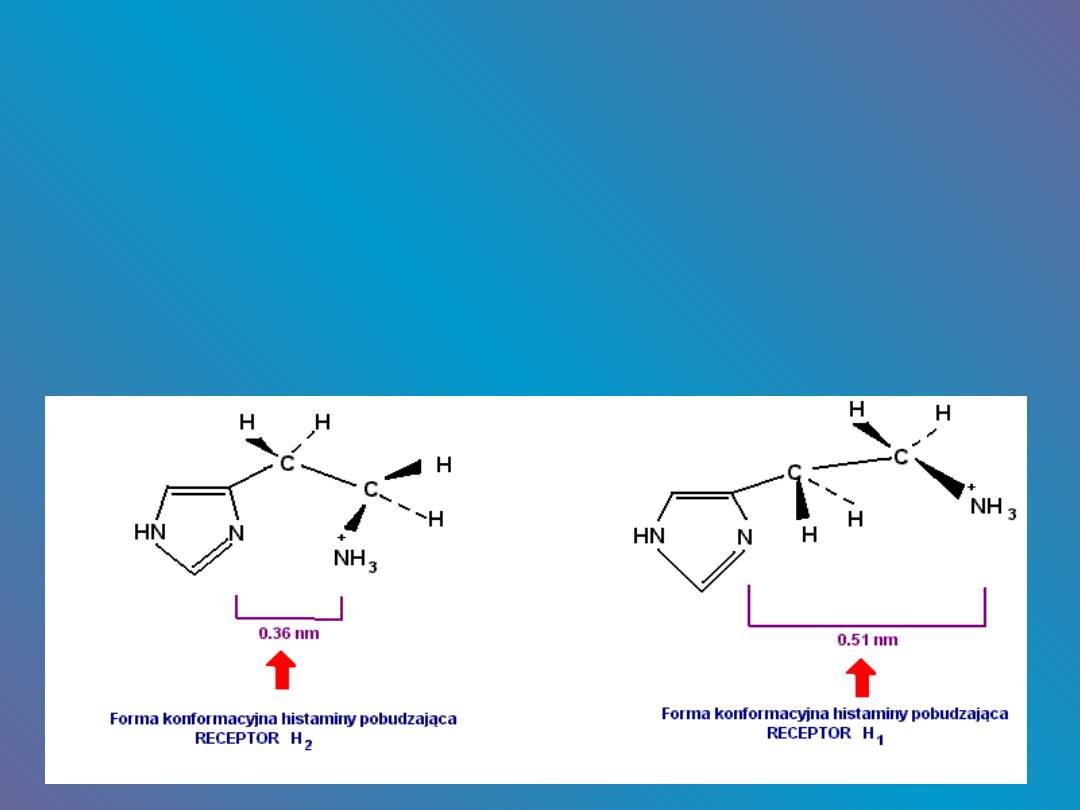
przybierania przez histaminę różnych form
konformacyjnych.

Presynaptyczne receptory H
zlokalizowane są w zakończeniach
neuronów histaminergicznych
(H-autoreceptory) oraz w
zakończeniach innych neuronów (H-
heteroreceptorów). H-autoreceptory i
H-heteroreceptory obecne są w
licznych ośrodkowych i obwodowych
tkankach.
PRESYNAPTYCZNY
RECEPTOR H
3

Stymulacja presynaptycznych receptorów H
3
w neuronach histaminergicznych prowadzi,
poprzez mechanizm ujemnego sprzężenia
zwrotnego do zmniejszenia syntezy
histaminy z L-histidyny, katalizowanej przez
specyficzną dekarboksylazę histydyny oraz
do hamowania uwalniania histaminy.
Stymulacje presynaptycznych receptorów
innych neuroprzekaźników w neuronach
histaminergicznych – M
1
-muskarynowych,
2
-adrenoreceptorów i receptorów
opiatowych , hamuje uwalnianie histaminy.
W tkankach obwodowych receptory H
3
działające jako heteroreceptory w innych
neuronach, wpływają na neurotransmisję
cholinergiczną, adrenergiczną,
dopaminergiczną, serotoniczną i
peptydoergiczną.

Histamina stymuluje wszystkie podtypy
receptorów histaminowych. Przez
metylowanie histaminy możliwe jest w
poszczególnych przypadkach zwiększenie
selektywności do jednego z trzech podtypów
receptora.
Metylowanie łańcucha w pozycji i prowadzi w
zasadzie do wzrostu H
3
-aktywności związków o
odpowiedniej konfiguracji stereochemicznej.
Metylowanie atomu N
zwiększa H
3
i H
2
-aktywność, przy
czym H
3
> H
2
>H
1
Związki z dużym podstawnikiem przy atomie N
są H
3
-
antagonistami.
N
-metylohistamina (główny metabolit histaminy) oraz
N
-metylohistamina są nieaktywne
Metylowanie atomu C2 pierścienia imidazolu prowadzi
do H
1
selektywnych agonistów, jednak o słabszym
działaniu od histaminy
Metylowanie atomu C5 pierścienia prowadzi do H
2
-
selektywnych agonistów
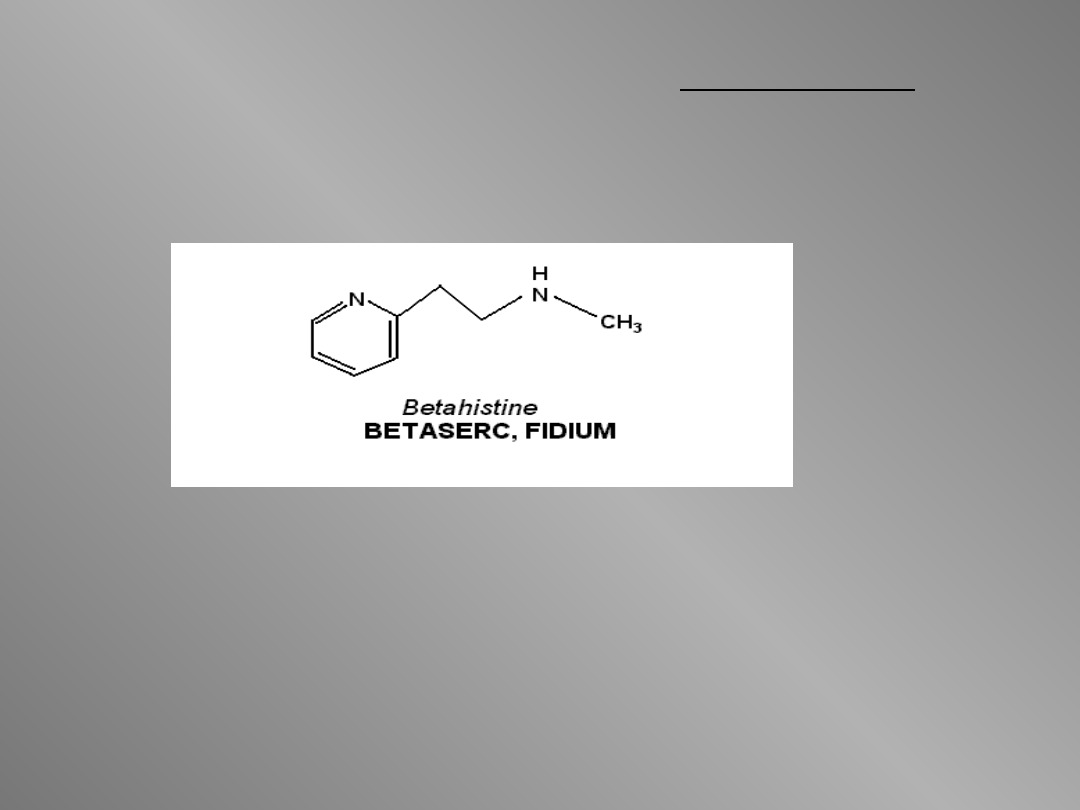
W lecznictwie stosowana jest betahistyna
(2-[2-(metyloamino)etylo]pirydyna).
Pokonuje ona barierę krew-mózg i
wykazuje aktywność H
3
-antagonistyczną.
Stosowana jest w zespole Menier’a.
Potencjalnymi wskazaniami dla ligandów
receptorów H
3
są: bezsenność, zaburzenia
psychiczne, stres migrena, skurcz oskrzeli i
zaburzenia wydzielania kwasu żołądkowego.
Potenjalnymi agonistami receptorów H są (R )-
-metylohistamina,(R,S)-,-
dimetylohistamina, imetit i immipep.
Selektywnymi antagonistami
receptorów H
3
są tioperamid,
clobenpropit.
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
Wyszukiwarka
Podobne podstrony:
Antagoniści receptora histaminowego H1
Antagonisci receptorow histaminowych
Antagoniści receptora histaminowego H1
Receptory histaminowepart prezentacja 5
Histamina i receptory1
prezentacja ścieżki sygnalizacyjne z udziałem receptora błonowego
Receptura zbiorczy GWSH 2
Agoni Ťci receptor w alfa i beta adrenergicznych
p histaminowymi wysylka
C Receptury Wydanie II cshre2
Zasady wyceny leków recepturowych i gotowych, Farmacja, farmakologia
Zaliczenie z receptury-2, materiały ŚUM, IV rok, Farmakologia, III rok, 7 - Receptura (TheMordor), Z
więcej podobnych podstron