większość pytań się powtórzyło się z tego, co było na gmailu. Z pytań "niespodziewanych"
pojawiło się:
- 3 reaktory do narysowania (a nie tylko do podpisania)
- jakie są rodzaje filtrów i filtracji do izolowania biomasy. Od czego zależy szybkość izolacji
biomasy.
- jedno zadanie brzmiało mniej więcej tak: "Zawiesina o objętości 75m3 zawierała biomasę o
stężeniu 12 kg suchej masy. Po od wirowaniu z odpłukaniem, stężenie biomasy wyniosło 36
kg suchej masy. Do jakiej objętości zagęszczono zawiesinę i ile wody odpłukano?"
- jedno zadanie jak na ostatnim kole u Wojczukowej (to ze stała Henry'ego)
- też zadanie z ostatniego koła: zadanie na policzenie liczby obrotów mieszadła dla przebiegu
burzliwego, przy podanej gęstości, siły lepkości i średnicy mieszadła
WYKŁAD 1, 16.02.2012
Wykład organizacyjny
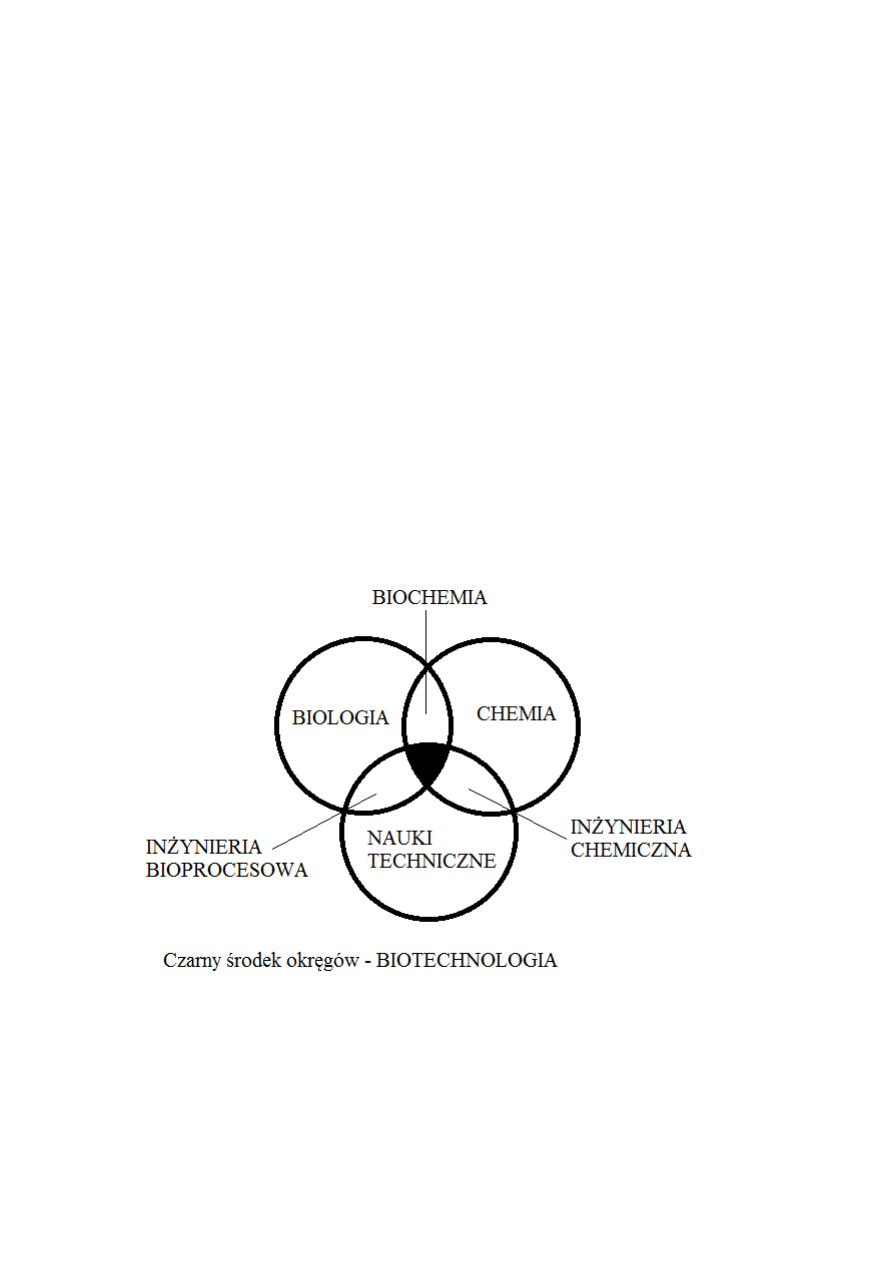
Biotechnologia jest interdyscyplinarną dziedziną nauki, obejmującą różne kierunki
technicznego wykorzystania materiałów i procesów biologicznych.
Zgodnie z definicją UE, biotechnologia jest dziedziną integrującą biochemię, mikrobiologię,
biologię molekularną, genetykę i chemię w celu praktycznego, użytkowego wykorzystania
kultur mikroorganizmów, tkanek lub elementów ich struktury.

IUPAC określa biotechnologię jako zastosowanie biochemii, biologii molekularnej,
mikrobiologii i inżynierii chemicznej w procesach przemysłowych i ochronie środowiska.
Wspólną cechą tych definicji jest:
interdyscyplinarny charakter biotechnologii
przemysłowe zastosowanie mikroorganizmów, enzymów lub kultur tkankowych
Interdyscyplinarny charakter biotechnologii powoduje, że może ona być opisywana z różnych
punktów widzenia – najczęściej z punktu widzenia biochemika lub mikrobiologa. W takich
przypadkach główny nacisk położony jest na wyjaśnienie przemian biochemicznych
zachodzących w komórkach i fizjologicznych uwarunkowań procesów technologicznych.
Termin „technologia” ma dwojakie znaczenie:
oznacza metodę wytwarzania określonego produktu
oznacza naukę o metodach wytwarzania określonego produktu
EKOLOGIA, BIOLOGIA KOMÓRKI, MIKROBIOLOGIA, GENETYKA, BIOCHEMIA,
BIOLOGIA MOLEKULARNA, CHEMIA, INŻYNIERIA, MATEMATYKA,
INFORMATYKA, FIZYKA, EKONOMIA
↓
BIOTECHNOLOGIA
↓
OCHRONA ZDROWIA, OCHRONA ŚRODOWISKA, ROLNICTWO, PRZEMYSŁ
SPOŻYWCZY, PRZEMYSŁ CHEMICZNY, SUROWCE, NOŚNIKI ENERGII,
ANALITYKA, INNE ZASTOSOWANA
Biotechnologia zielona → rolnictwo, produkcja żywności
Biotechnologia czerwona → medycyna, farmacja
Biotechnologia szara → ochrona środowiska, bioremediacja
Biotechnologia biała → zastosowania przemysłowe, wykorzystanie enzymów
Biotechnologia niebieska → technologie wodne, wykorzystanie organizmów wodnych
Biotechnologia fioletowa → ustawodawstwo związanie z biotechnologią
Główne zalety biotechnologii:
przetwarzanie surowców odnawialnych – cukrowce (skrobia, sacharoza, celuloza)
stanowią około 1% biomasy (10
14
-10
15
kg); obecnie cukrowce i ich pochodne (np.
melasa) są głównymi surowcami w procesach biotechnologicznych; szacuje się, że w

niedalekiej przyszłości można liczyć na przynajmniej 10-krotne zwiększenie
biotechnologicznego
wykorzystania
surowców roślinnych, pod warunkiem
opracowania efektywnej metody przetwarzania celulozy, lignin i hemiceluloz
(biopolimery ulegają bardzo powolnej biodegradacji)
duża różnorodność bioprocesów i otrzymywanych bioproduktów
selektywne otrzymywanie enancjomerów biologicznie czynnych
łagodne warunki przebiegu bioprocesów
niska energochłonność bioprocesów
duży stopień bezpieczeństwa
mniejsze niż w przypadku procesów chemicznych zanieczyszczenia środowiska;
łatwiejsze do neutralizacji ewentualne skażenia
Biotechnologia dzieli się na:
biotechnologię tradycyjną – wykorzystuje do procesów chemicznych naturalne
enzymy, szczepy drobnoustrojów i komórki organizmów wyższych nie zawierające
obcego materiału genetycznego
biotechnologię nowoczesną – stosowane są szczepy drobnoustrojów lub linie
komórkowe skonstruowane metodami inżynierii genetycznej, względnie enzymy
zmodyfikowane technikami inżynierii białka; nowoczesne metody i rozwiązania
biotechnologiczne obejmują również:
o rekombinacje genetyczne in vitro i klonowanie genów
o
fuzje komórek (protoplastów)
o
inżynierię białka
o techniki hodowli in vitro komórek organizmów wyższych
o
technologie immobilizacji enzymów
o
biokatalizę w układach niewodnych
o komputerowe modelowanie i sterowanie bioprocesami
o
technikę ciągłych procesów biotechnologicznych
o
nowoczesne techniki izolacji i oczyszczania bioproduktów
Najważniejsze wydarzenia w historii biotechnologii:
1. Wyjaśnienie przez Ludwika Pasteura (1857) roli drożdży w fermentacji alkoholowej.

2. Wprowadzenie do praktyk przemysłowych i medycznych czystych kultur
bakteryjnych, grzybów mikroskopijnych (pleśni) i drożdży.
3. Uruchomienie produkcji drożdży piekarniczych w warunkach napowietrzonych
hodowli wgłębnych (1880), wprowadzenie czystych kultur drożdży do piwowarstwa
(1883).
4. Wykorzystanie wyciągu z pleśni Aspergillu oryzae do scukrzania zacierów
skrobiowych.
5. Uruchomienie przemysłowej produkcji amylazy pleśniowej TAKA-DIASTAZA w
USA (1894).
6. Opracowanie techniki złoża zraszanego z mikroflorą degradującą składniki wód
ściekowych.
7. Wykorzystanie bakterii do oczyszczania ścieków w Manchesterze (1914).
8. Opracowanie mikrobiologicznych metod syntezy butanolu i acetonu na drodze
fermentacji acetonowo-butanolowej przez Clostridium (1918).
9. Opracowanie metod beztlenowej degradacji zanieczyszczeń ściekowych.
10. Uruchomienie przemysłowej produkcji etanolu z hydrolizatów drewna.
11. Uruchomienie wielkoprzemysłowej produkcji penicyliny pod koniec II Wojny
Światowej, co przyczyniło się do opracowania w następnych latach wielu procesów
produkcji innych antybiotyków.
12. Opracowanie metod biotransformacji mikrobiologicznej różnych steroidów.
13. Wprowadzenie do praktyki przemysłowej technologii immobilizowanych enzymów.
14. Konstruowanie nowych genotypów metodami inżynierii genetycznej.
15. Dalszy postęp w konstruowaniu bioreaktorów i prowadzeniu bioprocesów.
WYKŁAD 2, 23/02/2012
Prawie każdy proces biotechnologiczny można przedstawić w ogólnym zarysie jako
kombinację kolejnych operacji:
wybór mikroorganizmów do danego procesu
przygotowanie pożywki
hodowla mikroorganizmów, wytwarzanie produktu
wydzielanie i oczyszczanie produktu

Realizacja każdego procesu biotechnologicznego wymaga harmonijnego zgrania wszystkich
tych elementów składowych – należy przy tym pamiętać, że głównym kryterium efektywnego
prowadzenia procesu biotechnologicznego jest ekonomika produkcji. Oznacza to, że nie
wystarczy otrzymać wymagany produkt, lecz że należy wytwarzać go w taki sposób, aby
produkcja przynosiła zysk.
Udział różnych dyscyplin w rozwoju procesów biotechnologicznych
Operacje
Dyscypliny
Wybór mikroorganizmów
Systematyka
Genetyka
Fizjologia
Biochemia
Przygotowanie podłoża
Fizjologia
Chemia
Inżynieria bioprocesowa
Hodowla mikroorganizmów
Fizjologia
Inżynieria bioprocesowa
Wydzielanie produktów
Chemia
Inżynieria bioprocesowa
Praktyka przemysłowa pokazuje, że wiedza z zakresu chemii, biochemii i mikrobiologii jest
niewystarczająca do rozwiązania problemów przemysłu fermentacyjnego, spożywczego i
farmaceutycznego. Reakcja biochemiczna stanowi zasadniczy, ale tylko jeden etap procesu
produkcyjnego. Znacznie więcej problemów stwarza przygotowanie surowców, prowadzenie
procesu oraz izolacja produktu.
Generalnie, przemysł biotechnologiczny uszeregowany jest w sposób branżowy, np.
produkcja etanolu
produkcja kwasu cytrynowego
produkcja lizyny lub metioniny
produkcja penicylin (i ogólnie antybiotyków)
produkcja drożdży paszowych i piekarniczych
W różnych nitkach technologicznych występują takie same operacje jednostkowe oraz
procesy podstawowe, np.
fermentacja tlenowa lub beztlenowa
sedymentacja, wirowanie

odparowywanie, zatężanie
filtracja, krystalizacja, suszenie
Pomimo, że liczba bioprocesów i otrzymywanych produktów jest bardzo duża, liczba
procesów podstawowych jest niewielka.
Celem inżynierii bioprocesowej jest stworzenie matematycznego opisu procesów
podstawowych i operacji jednostkowych:
aby można było przewidzieć ich przebieg
w dowolnej skali
bez konieczności wykonywania doświadczeń pomocniczych
Umożliwia to budowę instalacji przemysłowej bez konieczności prowadzenia kosztownych
prób w skali półtechnicznej i technicznej.
Obszar tematyczny inżynierii bioprocesowej:
przenoszenie pędu – operacje dynamiczne, zachodzące pod działaniem siły
przenoszenie masy – operacje dyfuzyjne, zachodzące pod wpływem różnicy stężeń
jako siły napędowej
inżynieria reakcji chemicznych i biochemicznych
bioreaktory
Oczekiwania biotechnologii wobec inżynierii bioprocesowej:
tworzenie modeli matematycznych oddziaływań biologicznych
procesy rozdzielania (izolacji) złożonych i wrażliwych produktów
projektowanie bioreaktorów
Ogólny schemat pełnego procesu biotechnologicznego:
PRZYGOTOWANIE SUROWCÓW
(SUBSTRATÓW)
up-stream processing
↓
BIOPRZEMIANA
bioreaktor
↓
IZOLACJA ORAZ OCZYSZCZANIE
PRODUKTU
down-stream processing

1929r – Fleming uzyskał penicylinę G w postaci rozcieńczonego, zanieczyszczonego,
niestabilnego preparatu.
1940-1943r – produkcja i oczyszczanie penicyliny metodą wymrażania pary wodnej ze
stratą >60% produktu w kolejnych etapach
1943r – firma SHELL opracowała technologię zbliżoną do obecnej – zwiększona wydajność
do 85%

Podstawy logistyczne procesu biotechnologicznego:
DOSTAWY
↓
dostarczenie i magazynowanie surowców
UP-STREAM PROCESSING
↓
dejonizacja, pasteryzacja, mieszanie
FERMENTACJA
↓
DOWN-STREAM PROCESSING
↓
wytrącanie, dejonizacja, chromatografia,
odparowywanie, filtracja, krystalizacja,
suszenie
pakowanie, magazynowanie, dostawa
KLIENT
Surowce
Materiały używane w procesach fermentacyjnych:
woda, powietrze
cukry (jako główne źródło węgla)
źródła azotu (głównie amoniak)
sole mineralne
czynniki wzrostowe (witaminy, aminokwasy)
Źródła węgla i energii stosowane w procesach fermentacji mogą być sklasyfikowane jako:
nieoczyszczone (surowe)
częściowo oczyszczone
oczyszczone (rafinowane)
Surowe substraty są tańsze niż substraty oczyszczone lub częściowo oczyszczone, lecz
zawierają różne zanieczyszczenia, które będą musiały być usunięte na dalszych etapach
obróbki (down-stream lub up-stream). Występujące zanieczyszczenia mogą nawet
powodować zahamowanie procesu fermentacji. Surówka może także charakteryzować się
zmienną jakością, co może wpływać na jakość końcowego produktu.

Przykłady stosowanych substratów:
Surówka
Olej palmowy
Mączka rybna, mączka sojowa
Melasa
Namok kukurydziany
Woda odpadowa
Substraty częściowo oczyszczone
Syropy glukozowe, syropy fruktozowe
Oleje roślinne
Skrobia
Substraty oczyszczone (rafinowane)
Media do hodowli komórek
Hydrolizaty białkowe
Glukoza (proszek)
Sacharoza (krystaliczna)
Pepton – hydrolizat białkowy powstały przez trawienie białka pepsyną.
Trypton – hydrolizat białkowy powstały przez trawienie białka trypsyną.
Niektóre różnice pomiędzy procesem chemicznym i biochemicznym (różnice odnoszą się do
procesu biochemicznego):
bardziej złożone mieszaniny reagujące (skład, fazy)
przyrost stężenia biomasy reagenta jako wynik reakcji biochemicznej
zdolność mikroorganizmów do syntezy katalizatorów (enzymów) swoich własnych
reakcji
łagodne warunki temperaturowe reakcji
zasadniczo reakcje przebiegają w fazie wodnej
stosowane jest niskie stężenie substratu i produktu w mieszaninie reakcyjnej
Kluczowe składniki w projektowaniu bioreakcji:
termodynamika – zmiany równowagi reakcji, ciepło dostarczane lub wyzwalane,
równowaga fazowa
kinetyka – szybkości reakcji biochemicznych, kinetyka wymiany masy
stechiometria – ilościowe przekształcenia biochemiczne
równowaga – bilans masowy i energetyczny
parametry fizyczne – przepływ materiału, mieszanie w reaktorze

Skala laboratoryjna
Eksperymenty na małą skalę
Badanie kinetyki i wydajności
Hodowle wytrząsane lub małe bioreaktory
Wstępny szacunek kosztów
Skala półtechniczna (pilotowa)
Bioreaktor 100-1000L
Badanie kinetyki i wymiany mas
Ocena kosztów i zysków (ocena ekonomiczna)
Badania nad zwiększeniem skali
Badania nad izolacją produktu
Skala przemysłowa
Produkcja komercyjna
Bioreaktor 1000-1000000L
Rozwiązywanie problemów technicznych
Wprowadzanie ulepszeń
Optymalizacja procesu
Zapewnienie jakości
WYKŁAD 3, 1/03/2012
Główne typy komórek stosowanych w procesach fermentacyjnych
Typy komórek
Przykład
Komórki zwierzęce
CHO – komórki jajnika chomika chińskiego
komórki HYBRIDOMA (do produkcji
przeciwciał monoklonalnych
komórki owadzie
Grzyby
Penicillum sp.
Aspergillus sp.
Drożdże
Saccharomyces sp.
Saccharomycopsis sp.
Kluyveromyces sp.
Pichia sp.
Candida sp.
Bakterie
Escherichia coli
Bacillus sp.
Pseudomonas sp.
Streptomyces sp.
Porównanie niektórych właściwości organizmów/komórek stosowanych w procesie
biokatalizy
BAKTERIE
DROŻDŻE
GRZYBY
KOMÓRKI
ZWIERZĘCE
szybkość
wzrostu
+++++
+++
++
+
łatwość
manipulacji
genetycznych
+++++
+++
++
++
fałdowanie
białek
-
++
++
++++
glikozylacja
białek
-
?
?
++++
odporność
mechaniczna
++++
++++
++++
+
Chociaż przedstawione porównanie jest bardzo dużym uogólnieniem, to z tabeli tej wynika,
że komórki bakteryjne pomimo szybkiego wzrostu i łatwości manipulacji genetycznej, nie
posiadają dwóch bardzo ważnych cech: właściwego fałdowania obcych białek
(rekombinowanych) oraz ich glikozylacji (generalnie zdolności do modyfikacji
potranslacyjnych). Z tych powodów szereg firm farmaceutycznych coraz powszechniej
wykorzystuje komórki zwierzęce do produkcji białek zwierzęcych (ludzkich) dla celów
terapeutycznych (w tym np. enzymów do leczenia niektórych genetycznie uwarunkowanych
schorzeń genetycznych).
Główne produkty procesu biochemicznego
TYP PRODUKTU
TYPOWY STOSOWANY
MIKROORGANIZM
WIELKOŚĆ RYNKU
ŚWIATOWEGO
(ton/rok)
związki organiczne
Saccharomyces cerevisiae
2 x 10
7
biomasa
drożdże
5 x 10
5
kwasy organiczne
Aspergillus niger
3 x 10
5
aminokwasy
Brevibacterium flavum
3 x 10
5
transformacje
mikrobiologiczne
Rhizopus arrhizus
4 x 10
4
antybiotyki
Penicillum chrysogenum
4 x 10
4
enzymy
Aspergillus niger
1,4 x 10
3
Transformacje mikrobiologiczne – np. transformacje steroidów (zmiana podstawników,
stereoizomeria).

Rodzaje procesów przemysłowych
PROCES
CZYNNIK KATALITYCZNY
PRZYKŁADY
BIOSYNTEZY
drobnoustroje
biosynteza aminokwasów,
antybiotyków, polisacharydów
produkcja preparatów paszowych
komórki zwierzęce
produkcja szczepionek i przeciwciał
monoklonalnych
komórki roślinne
biosynteza terpenów, estrów i
alkaloidów
enzymy
biosynteza makrolidów, dekstranu,
akrylamidu
BIOTRANS-
FORMACJE
drobnoustroje
biotransformacje steroidów
otrzymywanie kwasu octowego i
innych kwasów, sorbozy
enzymy
konwersja glukozy do fruktozy,
fumaranu do jabłczanu
transformacje steroidów
HYDROLIZA
enzymy
hydroliza białek, oligosacharydów i
polisacharydów
hydroliza enancjomerów
Bilansowanie wzrostu mikroorganizmów
Ilościowe ujęcie procesu technologicznego jest podstawowym narzędziem w każdej
dyscyplinie inżynierskiej. Modele matematyczne są szeroko stosowane w projektowaniu,
optymalizacji i sterowaniu procesami inżynierii chemicznej. Opisy ilościowe obejmują
bilanse masowe i energetyczne oraz opisy kinetyki danego procesu. Zestawienie bilansów
masowego i energetycznego jest podstawową czynnością przy projektowaniu procesu
technologicznego. Zasady bilansowania procesów (przemian) chemicznych są znane
(stechiometria reakcji) i zasadniczo nie sprawiają trudności.
Przemiany biochemiczne różnią się od przemian chemicznych tym, że główny składnik
przemian biochemicznych – mikroorganizmy – cechuje zmienność typowa dla organizmów
żywych, zaś reakcje przebiegające wewnątrz komórek są bardziej złożone. Z tych powodów
bezpośrednie zastosowanie reguł stechiometrycznych do przemian biochemicznych nie jest
możliwe.
Trudności, jakie stwarza ilościowe ujęcie przemian biochemicznych ilustruje podany niżej
przykład.
Reakcję utleniania węgla można opisać prostym równaniem:
C + O
2
→ CO
2
+ Q (Δ
h
0
= -397,7kJ/mol)
Zgodnie z powyższym równaniem stechiometrycznym, podczas spalania 1kg węgla
powinniśmy otrzymać 3,67kg CO
2
oraz 32,8MJ energii.

W rzeczywistości bezpośrednie wykorzystanie tego równania nie jest możliwe, gdyż węgiel
kamienny, obok pierwiastka węgla (około 60%), zawiera również inne składniki, zarówno
palne (siarka), jak i niepalne. Skład węgla kamiennego jest zmienny. Spalanie węgla
kamiennego można przedstawić w sposób ogólny na schemacie:
WĘGIEL
KAMIENNY
+
POWIETRZE
→
GAZY
SPALINOWE
+
ŻUŻEL
+
Q
Ilość wydzielanego ciepła zależy od rodzaju spalanego węgla. Wynosi przeciętnie
16-22MJ/kg węgla. Ilość i skład gazów spalinowych zależą również od składu spalanego
węgla.
Zatem do sporządzenia bilansu spalania węgla kamiennego niezbędne są dane o jego składzie
i stopniu przekształcenia składników węgla w gazy spalinowe. Bilans taki będzie prawdziwy
dla konkretnego surowca oraz warunków spalania. Z praktycznego punktu widzenia możliwe
jest sporządzenie takiego bilansu dla pewnych przeciętnych danych dotyczących składu węgla
i stopnia konwersji. Przy takim podejściu nie uwzględnia się wszystkich przemian
zachodzących podczas spalania, nie przeprowadza się również bilansowania wszystkich
pierwiastków występujących w węglu kamiennym.
W przypadku bilansowaniu procesów biochemicznych stopień napotykanych trudności jest
znacznie większy, ponieważ:
skład elementarny mikroorganizmów jest zmienny
zmienny jest stopień przetworzenia substratów w produkty
Wzrost mikroorganizmów jest zjawiskiem bardzo złożonym. Komórki do wzrostu potrzebują
szeregu substancji. Wewnątrz komórki zachodzi szereg przemian biochemicznych, w wyniku
których powstają produkty i wytwarzana jest niezbędna energia.
Typowe substraty w układach biologicznych (źródła pierwiastków):
źródła węgla
glukoza, aminokwasy
źródła azotu
NH
4
Cl, (NH
4
)
2
SO
4
, aminokwasy, białka
źródła tlenu
powietrze
źródła wodoru
związki organiczne
źródła fosforu
fosforan nieorganiczny PO
4
3-
Typowe produkty w systemach biologicznych to biomasa, produkty reakcji i metabolity.

Ustalenie struktury chemicznej i podanie wzoru produktu i metabolitu jest proste. Lecz jaki
wzór chemiczny przypisać można komórce (biomasie)? Z uwagi na złożoność i różnorodność
procesów biochemicznych zachodzących w komórce, nie jest możliwe uwzględnienie
wszystkich relacji stechiometrycznych i kinetycznych tych przemian.
Generalnie, punktem wyjścia jest oparcie się o znane szlaki metaboliczne. Można zauważyć,
że makroskopowe przedstawienie procesu nie uwzględnia wszystkich elementów danej
przemiany. Dobrym przykładem może być produkcja etanolu przez drożdże S. cerevisiae oraz
X. mobilis.
W przypadku drożdży, makroskopowo równanie wygląda w ten sposób:
C
6
H
12
O
6
→ 2C
2
H
5
OH + 2CO
2
Biochemicznie, reakcja netto:
C
6
H
12
O
6
+ 2ADP + 2P
i
→ 2C
2
H
5
OH + 2CO
2
+ 2ATP + 4H
2
O +Q
W przypadku X. mobilis zapis makroskopowy jest identyczny, jak u drożdży, ale zapis
biochemiczny netto jest inny:
C
6
H
12
O
6
+ ADP + P
i
→ 2C
2
H
5
OH + 2CO
2
+ ATP + 2H
2
O +Q
Z uwagi na wspomnianą złożoność procesu wzrostu mikroorganizmów, wszelkie opisy
ilościowe muszą ograniczać się do wybrania istotnych zależności występujących podczas
rozwoju populacji mikroorganizmów. Modelowanie procesów biologicznych jest zatem
trudniejsze od modelowania procesów chemicznych. Podstawowa metoda ilościowego ujęcia
procesu wzrostu mikroorganizmów polega na poszukiwaniu takich wielkości
charakteryzujących mikroorganizmy oraz procesy ich namnażania, których wartości są
względnie stałe. Umożliwia to konstruowanie modeli uogólniających. Zależności takie,
średnie wartości parametrów, służyć mogą do obliczeń szacunkowych procesu wzrostu
mikroorganizmów.
Skład biomasy mikroorganizmów
Podanie zawartości wszystkich pierwiastków tworzących biomasę jest trudne – w praktyce
ogranicza się do czterech podstawowych pierwiastków: C, H, O, N. Ponieważ stanowią one
około 95% całości suchej masy, zatem sporządzenie bilansu dla tych pierwiastków jest
wystarczająco dobrym przybliżeniem. Jedynie w wyjątkowych przypadkach uwzględnia się
siarkę i fosfor.

Dla celów bilansu elementarnego, skład biomasy mikroorganizmów można przedstawić w
postaci analogicznej do wzoru stechiometrycznego związku chemicznego:
C
δ
H
a
O
b
N
c
δ = 0 lub 1 (węgla nie ma w związku lub w nim jest)
Dla E. coli:
C: 50% suchej masy
H: 8% suchej masy
O: 20% suchej masy
N: 14% suchej masy
Na tej podstawie wyliczono średni wzór dla biomasy:
CH
1,8
O
0,5
N
0,2
Ilość substancji odpowiadająca temu wzorowi nazywana jest węglomolem (C-mol), a zatem
węglomol to taka ilość substancji, która zawiera 1 mol węgla.
Zgodnie z tą definicją wzory i masy węglomola podanych substancji są następujące:
etanol → C
2
H
5
OH → CH
3
O
0,5
→ 23g/C-mol
glukoza → C
6
H
12
O
6
→ CH
2
O → 30g/C-mol
Dla związków nie zawierających węgla (δ=0) wzór nie ulegnie zmianie, a węglomol jest
równy molowi tego związku.
W przypadku biomasy należy ustalić strukturę węglomola w oparciu o dane analizy
elementarnej. Doświadczalnie wykazano, że współczynniki stechiometryczne zmieniają się w
podanych niżej przedziałach:
H: 1,5-2,0
O: 0,43-0,56
N: 0,10-0,25
Jako średnią wartość przyjmujemy wzór:
CH
1,8
O
0,5
N
0,2
Skład elementarny wybranych mikroorganizmów
MIKROORGANIZM
WĘGLOMOL
Saccharomyces cerevisiae
CH
1,83
O
0,54
N
0,1
Candida utilis
CH
1,83
O
0,50
N
0,17
Klebsiella aerogenes
CH
1,87
O
0,56
N
0,20
Aerobacter aerogenes
CH
1,83
O
0,55
N
0,25

Znając wzór węglomola biomasy można wyliczyć zapotrzebowanie na substraty podczas jej
produkcji.
Stechiometria pewnej reakcji:
C
6
H
12
O
6
+ aO
2
+ bNH
3
→ cCH
1,66
O
0,5
N
0,18
+ dCO
2
+ eH
2
O
Bilans pierwiastkowy:
węgiel
6 = c + d
wodór
12 + 3b = 1,66c + 2e
tlen
6 + 2a = 0,5c + 2d + e
azot
b = 0,18c
Mamy układ czterech równań z pięcioma niewiadomymi. Konieczna jest znajomość
parametru, który pozwoli na wyeliminowanie jednej niewiadomej (aby rozwiązać układ
równań). Często do tego celu używa się tzw. iloraz oddechowy RQ, określający stosunek
uwalnianego CO
2
do ilości wykorzystywanego O
2
.
W omawianym przypadku:
RQ =
d
/
a
= 1,02
RQ pozwala na wyeliminowanie jednej wartości (d lub a) i rozwiązanie układu równań
opisującego bilans pierwiastków. Znając współczynniki stechiometryczne i masę węglomola
biomasy można łatwo obliczyć wydajność tworzenia biomasy w przeliczeniu na zużycie
substratu i tlenu.
WYKŁAD 4, 8/03/2012
Oznaczanie zawartości biomasy jest jedną z podstawowych czynności analitycznych podczas
bioprocesu. Znajomość zmian stężeń biomasy jest niezbędne do wyznaczenia kinetyki
wzrostu mikroorganizmów i przyrostu produktu podczas hodowli i obliczania wydajności.
Biomasa – masa mikroorganizmów rosnących na pożywce; może odnosić się do dwóch pojęć:
mokra biomasa – masa mikroorganizmów oddzielonych od pożywki poprzez
wirowanie lub filtrację; jest to zatem łączna masa mikroorganizmów i wody zawartej
w przestrzeni pomiędzy komórkami
sucha biomasa – masa mikroorganizmów oddzielonych od pożywki i wysuszonych w
temperaturze 105ºC do stałej wagi
105ºC – temperatura, w której pozbywa się wody krystalizacyjnej. Wyższa temperatura
mogłaby zniszczyć związki organiczne.

Metody oznaczania biomasy mikroorganizmów hodowanych w ciekłych podłożach:
bezpośrednie, polegające na wagowym oznaczeniu masy drobnoustrojów
oddzielonych od pożywki – dające wartość mokrej biomasy
pośrednie:
o metoda turbidymetryczna – polega na pomiarze zmętnienia zawartości
komórek w pożywce; metoda ta może być wykorzystywana tylko w
określonym zakresie stężeń; dokładność pomiaru zależy również od
właściwego przygotowania próby (konieczne jest wyeliminowanie składników
pożywki, które wpływają na pomiar)
o metody chemiczne – polegają na oznaczeniu stężenia określonego składnika
komórki; przyjmując, że jego zawartość jest wartością stałą; najczęściej
oznacza się zawartość ATP (np. metody luminescencyjne z lucyferazą)
W ciągu doby organizm ludzki jest w stanie wyprodukować około 45kg ATP.
Technika filtracji membranowej obejmuje:
zawiesina fermentacyjna jest filtrowana przez wysuszony, wytarowany filtr
membranowy (standardowo sączek z octanu celulozy)
filtr jest przemywany celem usunięcia rozpuszczalnych składników medium
filtr jest suszony i ważony
sucha masa (g/dm
3
) =
(masa membrany po filtracji – masa membrany przed filtracją)
/
objętość próby
Technika wirowania obejmuje:
wirowanie zawiesiny fermentacyjnej w wysuszonych, zważonych probówkach przy
najwyższych obrotach
usunięcie supernatantu, przemycie osadu i ponowne wirowanie
wysuszenie osadu w probówce i jej zważenie
sucha masa (g/dm
3
) =
(masa probówki po filtracji – masa probówki przed filtracją)
/
objętość próby
Wzrost mikroorganizmów jest wysoce złożoną przemianą, w której z surowców podłoża
hodowlanego wytwarzana jest biomasa oraz produkty metabolizmu.
Bilans ogranicza się w zasadzie do czterech podstawowych pierwiastków (C, H, O, N). W
rzeczywistości w pożywce może występować kilka substancji pełniących identyczne funkcje
(np. stanowiących źródło węgla). Do celów bilansowych można je zastąpić jednym

substratem o odpowiednio dobranym składzie (w sensie chemicznym). W takim ujęciu ogólny
schemat rozpatrywanego procesu można przedstawić następująco:
S
S
+ S
N
+ O
2
→ X + P + CO
2
+ H
2
O + Q
S
S
– źródło węgla
S
N
– źródło azotu
X – biomasa
P – produktu metabolizmu
Q – ciepło
ν
S
CH
aS
O
bS
N
cS
+ ν
N
C
δN
H
aN
O
bN
N
cN
+ ν
O
O
2
→ ν
X
CH
aX
O
bX
N
cX
+ ν
P
CH
aP
O
bP
N
cP
+ ν
C
CO
2
+ ν
w
H
2
O
C:
ν
S
+ ν
N
δ
N
= ν
X
+ ν
P
+ ν
C
H:
ν
S
a
S
+ ν
N
a
N
= ν
X
a
X
+ ν
P
a
P
+ 2ν
W
O:
ν
S
b
S
+ ν
N
b
N
+ 2ν
O
= ν
X
b
X
+ ν
P
b
P
+ 2ν
C
+ ν
W
N:
ν
S
c
S
+ ν
N
c
N
= ν
X
c
X
+ ν
P
c
P
W tym układzie równań występuje 7 niewiadomych współczynników bilansowych, układ nie
ma zatem rozwiązania. Niezbędne więc są 3 równania dodatkowe, wiążące ze sobą
współczynniki bilansowe.
Przemianę ogólną można opisać równaniem:
CH
i
O
j
+ a NH
3
+ b O
2
→ c CH
k
O
l
N
m
+ d CH
n
O
o
N
p
+ e H
2
O + f CO
2
+ Q
c, d, f – ułamki ilości węgla przekształconego w biomasę, produkty metabolizmu i CO
2
Bilans pierwiastków prowadzi do następującego układu równań:
C:
1 = c + d + f
H:
i + 3a = c
k
+ d
n
+ 2e
O:
j + 2b = c
l
+ d
o
+ e + 2f
N:
a = c
m
+ d
p
Otrzymano 4 równania i znacznie więcej niewiadomych, co uniemożliwia rozwiązanie
równania. Problem jest znacznie prostszy, jeżeli znamy wartości poszczególnych
współczynników.
Rozważmy wzrost drożdży Saccharomyces cerevisiae. Przyjmując, że skład biomasy drożdży
jest następujący C
6
H
10
O
3
N i rosną one na glukozie, wykorzystując ją jako źródło węgla,
można napisać następujące równanie:
C
6
H
12
O
6
+ b O
2
+ c
NH
3
→ d C
6
H
10
O
3
N + e H
2
O + f CO
2
C
6
H
12
O
6
→ 2 C
2
H
5
OH + 2 CO
2

Uwzględniając tu fakt, że glukoza jest również przekształcana w etanol. Jeżeli oznaczymy
przez ‘E’ ułamek ilości glukozy przekształconej w etanol, to w biomasę przekształcone będzie
‘1-E’ glukozy. Jeśli zatem pierwsze równanie pomnożymy przez ‘1-E’, a drugie przez ‘E’ i
dodamy je stronami, otrzymamy równanie:
C
6
H
12
O
6
+ (1-E) b O
2
+ (1-E) c NH
3
→ (1-E) d C
6
H
10
O
3
N + [(1-E) + 2E] CO
2
+ (1-E) e H
2
O + 2E C
2
H
6
O
C:
6 = 6(1-E)d + (1-E)f + 2E + 4E
H:
12 + 3(1-E)c = 10(1-E)d + 2(1-E)e + 12E
O:
6 + 2(1-E)b = 3(1-E)d + 2(1-E)f + 4E + (1-E)e + 2E
N:
(1-E)c = (1-E)d
Doświadczalnie można wyznaczyć kilka wartości i pozbyć się współczynników b, f i c lub d.
Współczynniki wydajności określają ilość powstającej biomasy lub produktu z określonej
ilości substratu.
1. wyrażone w węglomolach
współczynnik wydajności biomasy względem substratu (c-mol/c-mol)
y
X/S
= ν
X
/ν
S
współczynnik wydajności produktu względem substratu (c-mol/c-mol)
y
P/S
= ν
P
/ν
S
2. współczynniki masowe (g
X
/g
S
, g
P
/g
S
)
Y
X/S
= Δm
X
/-Δm
S
= y
X/S
∙ (M
C, X
/M
C, S
)
Współczynnik wydajności wzrostu drożdży na glukozie
Wydajność masowa: Y
X/S
= 0,5g/g
c-mol glukozy: 30g/c-mol
c-mol biomasy: 25,9g/c-mol
y
X/S
= Y
X/S
∙ (M
C,S
/M
C,X
) = 0,5 ∙ 30/25,9 = 0,58 c-mol/c-mol
W przedstawionej metodzie bilansowania wzrostu podstawową rolę odgrywa założenie, że
skład biomasy mikroorganizmów jest stały w warunkach wzrostu ustalonego.
Można zatem przyjąć, że relacje pomiędzy ilością zużywanego substratu i ilością powstającej
biomasy są również stałe. Relacje te opisuje współczynnik wydajności biomasy względem
substratu. W praktyce stosuje się masowy współczynnik wydajności:
Y
X/S
= y
X/S ∙
(σ
S
/σ
X
)
gdzie σ oznacza zawartość węgla w danym związku.
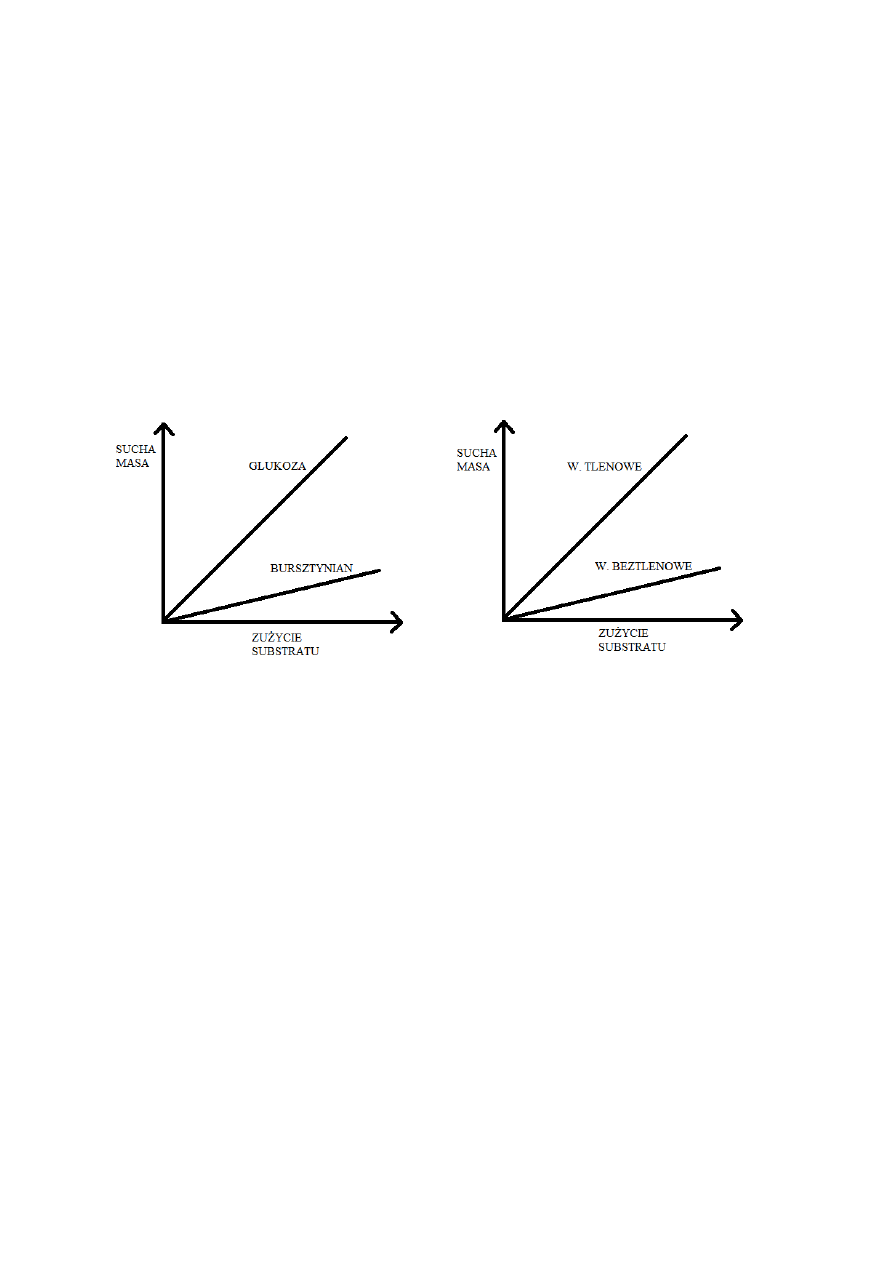
σ
glukoza
= 6C/C
6
H
12
O
6
= 72/180 = 0,40
σ
biomasa
= 12/24,6 = 0,49
WYKŁAD 5, 15/03/2012
Współczynnik wydajności biomasy jest zależny od:
rodzaju substratu
rodzaju medium (pożywki) i warunków wzrostu (wydajność jest wyższa w
optymalnych warunkach pH, składu pożywki i natlenienia)
Zasada Finka
Podczas wzrostu drożdży około 1/3 węgla z substratu wydzielana jest w postaci CO
2
,
natomiast 2/3 jest wbudowywane w biomasę. Zgodnie z tą zasadą wartość współczynnika
y
X/S
= 0,67. Wartość ta jest zbliżona do średnich wartości tego współczynnika wydajności w
przypadku wzrostu na węglowodanach. Nie jest to jednak reguła ogólna i obserwuje się
różnice w wartości tego współczynnika w odniesieniu do różnych substratów
metabolizowanych przez ten sam mikroorganizm.
Współczynnik wydajności biomasy:
wyznaczony eksperymentalnie
wielkość tego współczynnika różni się w zależności od organizmu, warunków wzrostu
lub substratu

Teoretyczna wydajność produktu
Znajomość teoretycznej maksymalnej wydajności, z jaką surowiec jest przekształcany w
produkt jest ważnym parametrem w projektowaniu bioprocesu i jego doskonaleniu. Zwrot
zainwestowanych środków w udoskonalenie produkcji jest wyższy, jeśli wyjściowa
wydajność była znacząco niższa od teoretycznego maksimum niż wtedy, gdy wydajność jest
bliska maksimum.
Przykład
Obliczyć teoretyczną maksymalną wydajność produkcji fenyloalaniny z glukozy dla E. coli
rosnącej na pożywce z glukozą i amoniakiem.
Przebieg szlaku metabolicznego:
GLUKOZA → → → → → FENYLOALANINA
W uproszczeniu przebieg reakcji jest następujący:
1 GLUKOZA → 2 PEP (fosfoenolopirogronian)
1 GLUKOZA → 1 E4P (erytrozo-4-fosforan)
1 PEP + 1 E4P → „INTERMEDIAT A“
INTERMEDIAT A + 1 PEP → FENYLOALANINA
Sumarycznie:
2 GLUKOZA → FENYLOALANINA
2 ∙ 180g
165g
Y
P/S
= 165/360 = 0,458g Phe/ g glukozy
Można wyróżnić następujące sytuacje:
1. Produkty są wytwarzane bezpośrednio z substratów w sposób energetycznie
uprzywilejowany, tzn. następuje wytworzenie energii chemicznej netto.
1 mol glukozy → 2 mole etanolu + 2 mole ATP
Teoretyczna maksymalna wydajność wynosi:
Y
P/S
= 96/180 = 0,51 kg etanolu/kg glukozy
Rzeczywista wydajność jest niższa, gdyż część glukozy jest przekształcana w biomasę i na
pokrycie energetycznego zapotrzebowania komórek. Podstawą obliczeń jest sumaryczne
równanie glikolizy.
2. Produkcja glutaminianu z glukozy przez Corynebacterium glutamicum przebiegające w
następującym szlaku metabolicznym:

GLUKOZA + 2 ADP + 2 P
i
→ 2 PIROGRONIAN + 2 ATP
PIROGRONIAN + NAD
+
→ ACETYLO-CoA + CO
2
+ NADH/H
+
PIROGRONIAN + CO
2
+ ATP + H
2
O → SZCZAWIOOCTAN + ADP + P
i
ACETYLO-CoA + SZCZAWIOOCTAN + NAD
+
→ α-KETOGLUTARAN + CO
2
+ NADH/H
+
α-KETOGLUTARAN + NADPH/H
+
+ NH
3
→ GLUTAMINIAN + NADP
+
+ H
2
O
Sumarycznie:
GLUKOZA + NH
3
+ ADP + P
i
+ NADPH/H
+
+ 2 NAD
+
→
→ GLUTAMINIAN + CO
2
+ ATP + 2 NADH/H
+
+ NADP
+
Zatem reakcja jest energetycznie możliwa (powstaje 1 mol ATP i 2 mole NADH/H
+
);
maksymalna teoretyczna konwersja (w odniesieniu do węgla) wynosi:
5 c-moli Glu/6 c-moli glukozy
Y
P/S
= 143/180 = 0,79g Glu/g glukozy
3. Wytwarzanie produktu wymaga nakładu energii (pochodzącej z katabolizmu substratu) i
można proces ten opisać równaniem ogólnym:
a C
α
H
β
O
γ
N
δ
+ b C
α
H
β
O
γ
N
δ
+ c ATP → C
α
H
β
O
γ
N
δ
+ c ADP + c P
i
Energia jest generowana w wyniku katabolizmu substratu.
C
α
H
β
O
γ
N
δ
+ b O
2
+ d ADP + d P
i
→ c CO
2
+ d ATP
Maksymalna wydajność jest kombinacją dwóch reakcji, w których produkcja/zużycie ATP
wynosi 0. W obliczeniach, dla uproszczenia, przyjmuje się, że NADH/H
+
jest
równoważnikiem 3 ATP. We wszystkich obliczeniach zakłada się również, że ΔG
0
dla każdej
reakcji jest ujemna.
Przykład obliczania wydajności biomasy
Komórki zarodka sosny Douglasa hodowano w bioreaktorze. Po okresie wstępnego wzrostu
komórki potraktowano regulatorami wzrostu, powodując powstanie zarodków (identycznych
klonów), które tworzyły następnie „syntetyczne nasiona”.
Medium wyjściowe zawierało 200kg glukozy i 25kg amoniaku na 1m
3
.
Do 0,95m
3
medium dodano 0,05m
3
komórek, inicjując hodowlę. Po dwóch tygodniach
nastąpiło całkowite zużycie glukozy.
Otrzymano 0,35m
3
biomasy.
Stężenie amoniaku obniżyło się do 0,001kg/m
3
.
Specyficzna gęstość mokrej biomasy była równa 1020kg/m
3
, a zawartość H
2
O wynosiła 90%.

Obliczyć Y
X/glukoza
i Y
X/amoniak
. Wyrazić biomasę jako suchą masę.
1. Obliczyć suchą masę biomasy.
X
wstępna
: 0,05m
3
∙ 1020kg/m
3
∙ 10% = 5,1kg
X
końcowa
: 0,35m
3
∙ 1020kg/m
3
∙ 10% = 35,7kg
Przyrost biomasy: 35,7kg – 5,1kg = 32,6kg
2. Obliczyć wydajność biomasy.
Y
X/glukoza
=
[przyrost biomasy]
/
[ilość przekształconej glukozy]
= 32,6/(0,95∙200) = 0,16kg biomasy/kg glukozy
Y
X/amoniak
= 32,6/(0,95∙25) = 1,29kg biomasy/kg amoniaku
Masę końcową amoniaku można pominąć, ze względu na bardzo niskie stężenie.
Bilanse pierwiastków można uogólniać, wprowadzając pojęcie stopnia redukcji. Jest to
liczba moli wolnych elektronów w przeliczeniu na 1 mol węgla. Elektrony te mogą być
przekazywane na tlen w procesie utlenienia.
Liczba elektronów obliczana jest addytywnie.
C – dawca 4 elektronów (+4)
H – dawca 1 elektronu (+1)
O – akceptor 2 elektronów (-2)
N – akceptor 3 elektronów (-3)
Bilans tlenu
W przypadku bilansu tlenu w procesie wzrostu mikroorganizmów korzystamy ze stopnia
redukcji.
Rozważmy całkowite utlenienie substratu o wzorze ogólnym:
C
δ
H
a
O
b
N
c
Jak obliczyć ilość tlenu niezbędną do utlenienia tego substratu?
Równanie reakcji jest następujące:
C
δ
H
a
O
b
N
c
+ n O
2
→ δ CO
2
+ ½ a H
2
O + ½ c N
2
Bilans elementarny dla tlenu jest następujący:
b + 2n = 2δ + ½ a
n = ¼ (4δ + a – 2b)
Γ = 4δ + a – 2b

Γ (bezwzględny stopień redukcji) – liczba elektronów przeniesionych na tlen przy jego
całkowitej redukcji.
γ
i
= Γ
i
– c
i
/c
N
∙ Γ
N
γ
i
(względny stopień redukcji) – zależny od związku, który jest źródłem azotu; dla soli
amonowych Γ
N
= 3, zatem:
γ
i
= 4 + a
i
– 2b
i
– 3c
i
Przyjmując za punkt wyjścia równanie stechiometryczne reakcji:
ν
S
CH
aS
O
bS
N
cS
+ ν
N
C
δN
H
aN
O
bN
N
cN
+ ν
O
O
2
→ ν
X
CH
aX
O
bX
N
cX
+ ν
P
CH
aP
O
bP
N
cP
+ ν
C
CO
2
+ ν
w
H
2
O
Można w oparciu o współczynnik bilansowy rozwinąć równanie bilansowe względem ν
O
:
ν
O
= ¼ (ν
S
Γ
S
+ ν
N
Γ
N
– ν
X
Γ
X
– ν
P
Γ
P
)
Znajomość współczynników wydajnośći biomasy z substratu i współczynnika wydajności
produktu umożliwia wyznaczenie ilości zużywanego azotu:
ν
N
/ν
X
= 1/c
N
∙ (c
X
– c
S
/y
X/S
– c
P
∙ y
P/X
)
Pozwala to wyeliminować z równania ν
N
. Równanie przyjmuje postać:
ν
O
= ¼ (ν
S
γ
S
– ν
X
γ
X
– ν
P
γ
P
)
Wprowadzenie pojęcia względnego stopnia redukcji oznacza przyjęcie za stan odniesienia
stopień redukcji związku stanowiącego źródło azotu. Jeżeli uwzględnimy definicję względnej
wydajności biomasy y
X/S
, to można wyznaczyć współczynnik zużycia tlenu.
Dla glutaminianu C
5
H
9
O
4
N:
Γ
X
= 4 + 1,8 – 2 ∙ 0,5 – 3 ∙ 0,2 = 4,2
Γ
N
= 4 ∙ 5 + 9 – 2 ∙ 4 – 3 = 18
γ
X
= Γ
X
– c
X
/c
N
∙ Γ
N
= 4,2 – 1/5 ∙ 18 = 0,6
y
O/X
= ν
O
/ν
X
= ¼ (γ
S
/y
X/S
– γ
X
– y
P/S
γ
P
)
Jeżeli ν
P
= 0 (czyli nie powstaje produkt metabolizmu):
y
O/X
= ¼ (γ
S
/y
X/S
– γ
X
)
W przypadku wzrostu drożdży na glukozie:
y
O/X
= ¼ (4/0,52 – 4,2) = 0,87 mola tlenu/g suchej masy = 1,07g tlenu/g s.m.
Wartość współczynnika y
O/X
jest wartością nieujemną, zatem:
y
X/S
< γ
S
/γ
X
– y
P/S
∙ γ
P
/γ
X
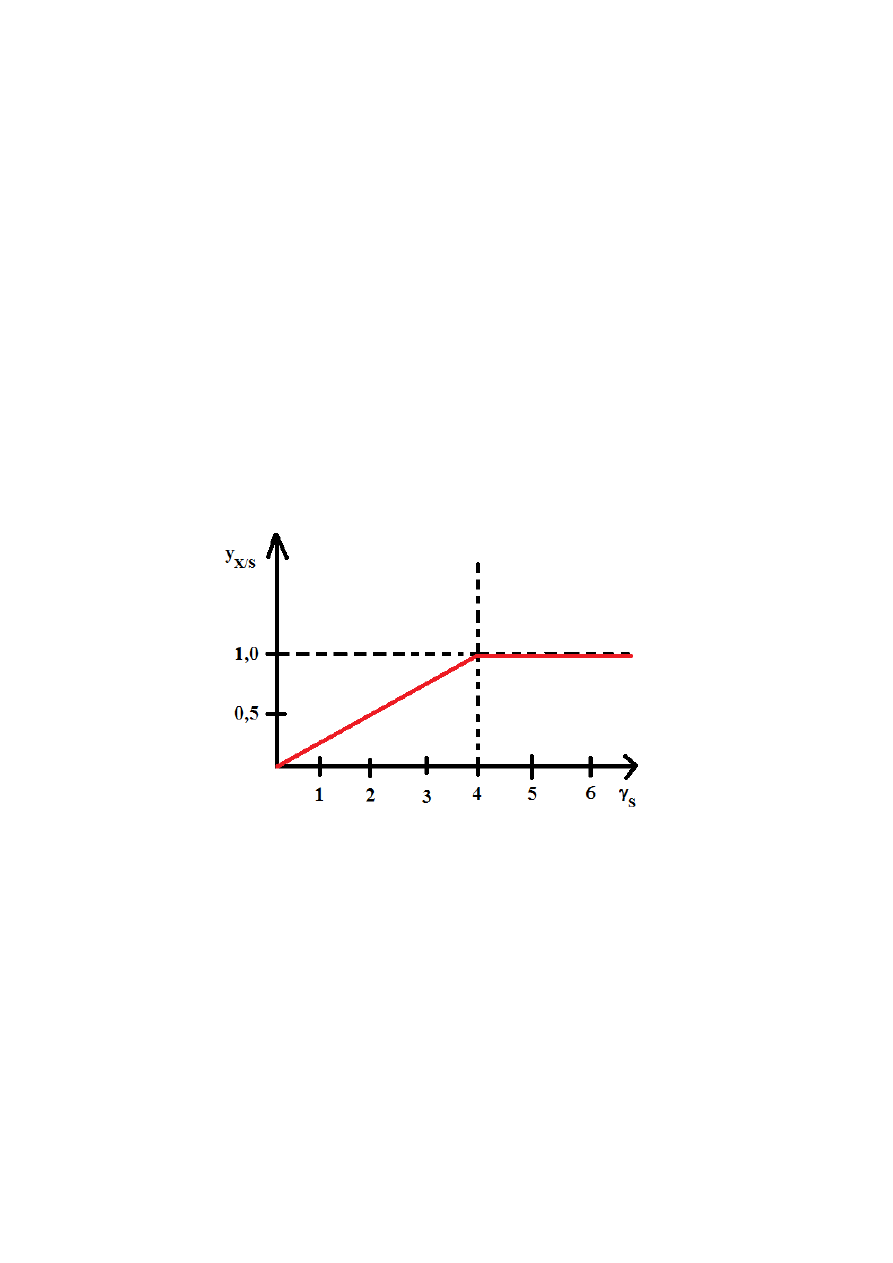
To równanie podaje ograniczenie bilansowe, wynikające z bilansu tlenu. Z bilansu węgla
wynika kolejne ograniczenie:
y
X/S
< 1 – y
P/S
Zatem dla prostego wzrostu (y
P/S
= 0) będą występowały dwa zakresy limitowania
współczynnika wydajności biomasy z substratu:
dla γ
S
< γ
X
ograniczenie wynika z bilansu tlenu
dla γ
S
> γ
X
ograniczenie wynika z bilansu węgla
WYKŁAD 6, 22/03/2012
Zależności między współczynnikiem wydajności biomasy z substratu y
X/S
i stopniem redukcji
substratu γ
S
wyznaczono doświadczalnie, otrzymując wartości:
γ
S
< 4,67 → y
X/S
= 0,13γ
S
γ
S
> 4,67 → y
X/S
= 0,60
Wykres zależności teoretycznej, przy założeniu, że z substratu powstaje wyłącznie biomasa.
Wartości te umożliwiają przybliżone obliczenie asymilacji tlenu. Na podstawie tych wartości,
dla przeciętnego składu biomasy:
dla γ
S
≤ 4,67
y
O/X
= ν
O
/ν
X
= 0,87 mola tlenu/c-mol biomasy
dla γ
S
≥ 4,67
Y
O/X
= 0,417γ
S
- 1,05 mola tlenu/kg biomasy
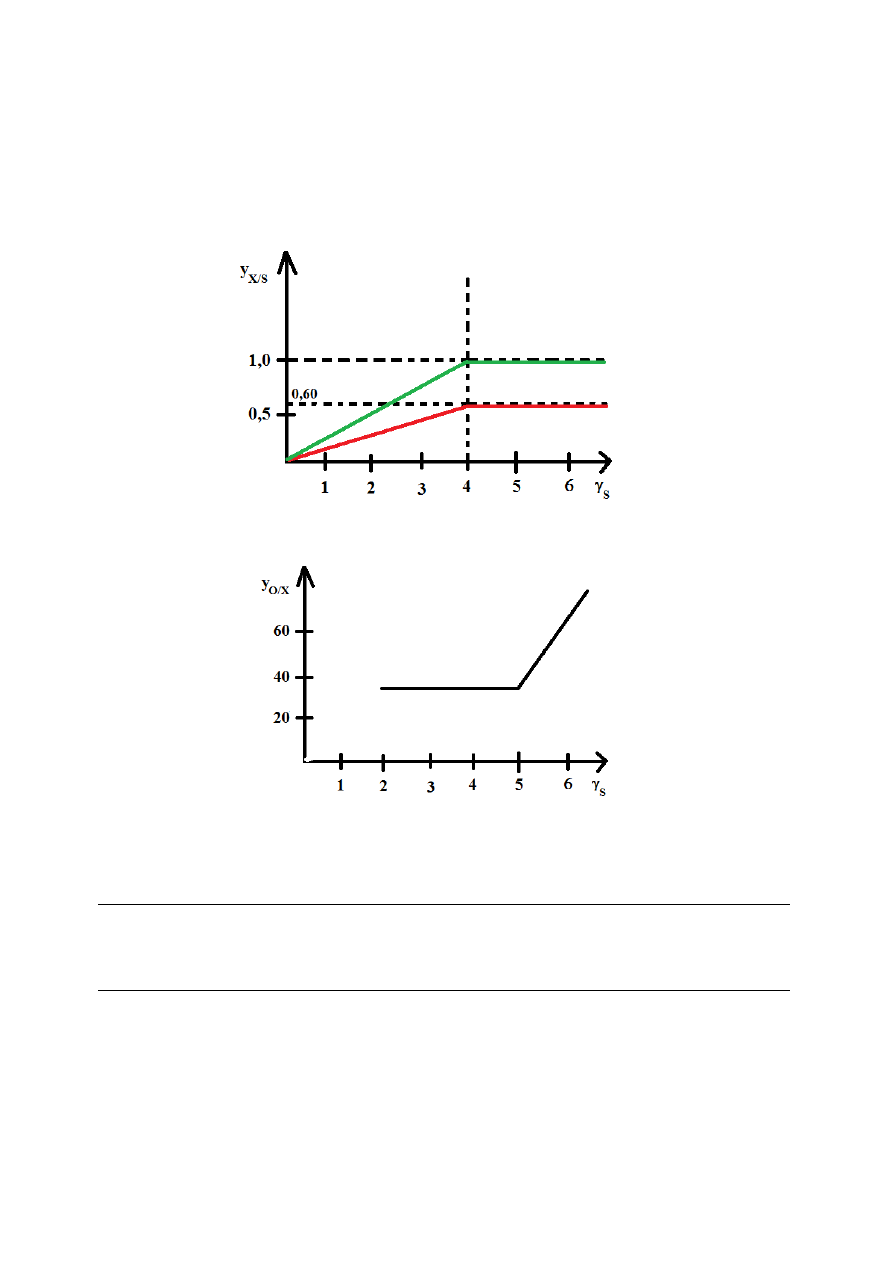
Należy pamiętać, że współczynniki wydajności dla tego samego substratu mogą być różne dla
różnych mikroorganizmów. Znajomość ograniczeń pozwala na optymalny dobór składu
pożywki.
Zależność współczynnika wydajności biomasy od stopnia redukcji substratu:
Zależność współczynnika zużycia tlenu od stopnia redukcji substratu:
Przeciętne
wartości
współczynnika
wydajności
(uśrednione
dla
wszystkich
mikroorganizmów):
y
X/S
(c-mol/c-mol)
Y
O/X
(mol/kg)
Y
O/X
(MJ/kg)
GLUKOZA
0,54
32,0
14,7
GLICEROL
0,61
34,5
15,9
METAN
0,59
93,6
43,1
ETANOL
0,57
63,3
29,1
Współczynniki wydajności wzrostu różnych mikroorganizmów na glukozie w warunkach
tlenowych:

y
X/S
(c-mol/c-mol)
Y
O/X
(mol/kg)
Y
O/X
(MJ/kg)
Candida utilis
0,60-0,66
18,2-23,8
8,7-11,4
Saccharomyces
cerevisiae
0,65
19,1
9,0
Trichoderma viridae
0,62-0,71
9,2-21,8
4,8-10,5
Eschericha coli
0,52-0,62
21,8-25,9
10,5-12,5
Pseudomonas
fluorescens
0,45
45,7
21,5
Rozważmy proste równanie stechiometryczne:
CH
i
O
j
+ a NH
3
+ b O
2
→ y CH
r
O
s
N
t
+ z CH
u
O
v
N
w
+ c H
2
O + d CO
2
(S)
(X)
(P)
Stopień redukcji (γ) można zdefiniować następująco:
S:
γ
S
= 4 + i – 2j
X:
γ
X
= 4 + r – 2s – 3t
P:
γ
P
= 4 + u – 2v – 3w
γ
H2O
= γ
CO2
= γ
NH3
= 0
Wyprowadzamy zależność:
γ
S
– 4b = yγ
X
+ 2γ
P
b = ¼ (γ
S
– yγ
X
– 2γ
P
)
Warunkiem wyprowadzenia tej zależności jest znajomość:
wartości stopnia redukcji (γ
i
) związków – a więc ich struktura
ilości wyprodukowanej biomasy (y)
ilości powstałego produktu (z)
Przykład
Obliczyć ilość butanolu wyprodukowanego ze 100 moli glukozy, zgodnie z równaniem
stechiometrycznym
100 C
6
H
12
O
6
+ a NH
3
→ 13 C
4,46
H
16,01
O
3,88
N
0,86
+ x CH
3
(CH
2
)
3
OH + 22 C
3
H
6
O +
0,4 CH
3
(CH
2
)
2
COOH + 14 CH
3
COOH + y CO
2
+ 135 H
2
+ 0,7 C
2
H
5
OH
Wartości niezbędne do obliczeń zebrano w tabeli:
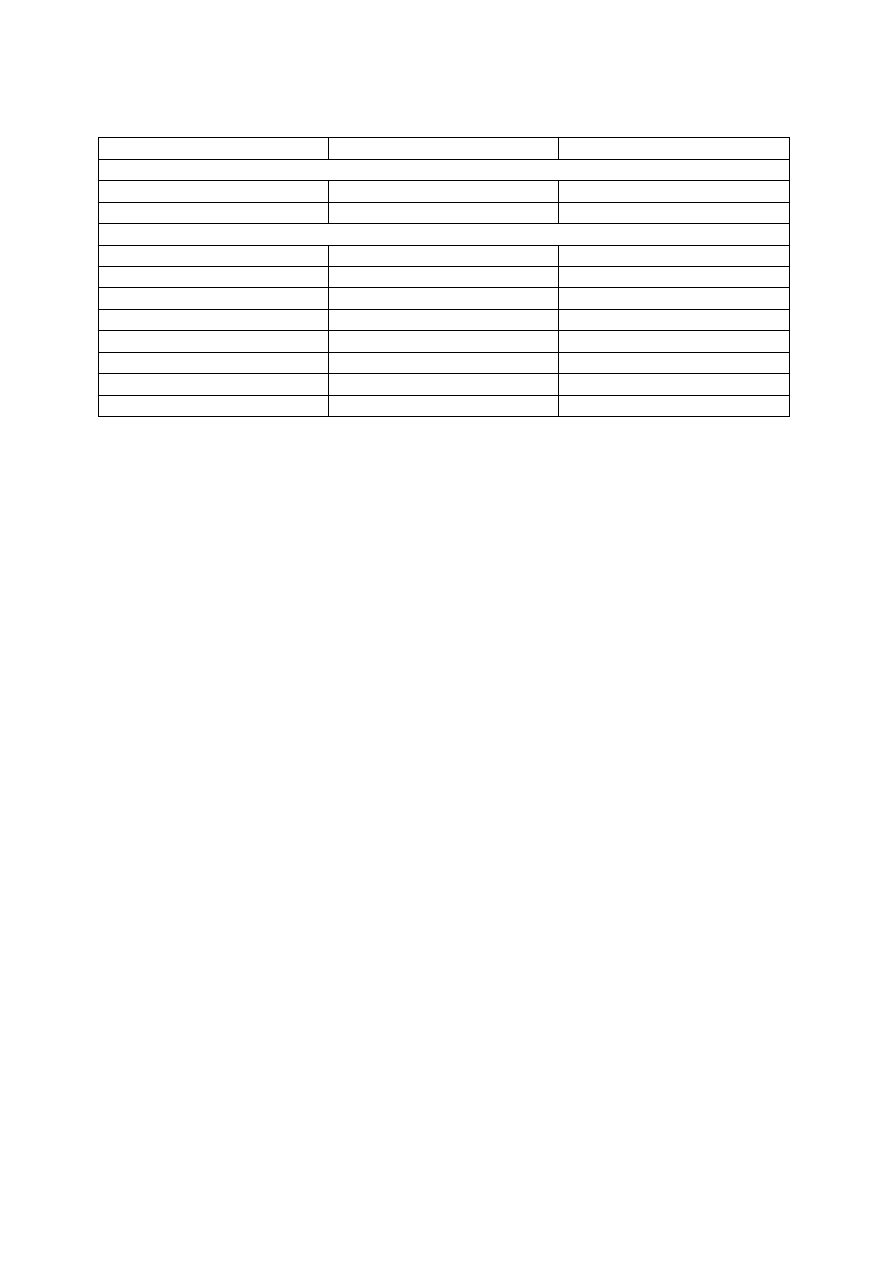
ZWIĄZEK
γ
i
γ
i
∙ ν
i
SUBSTRATY
C
6
H
12
O
6
24
24 ∙ 100 = 2400
NH
3
0
a ∙ 0 = 0
PRODUKTY
X (biomasa)
23,51
23,51 ∙ 13 = 305,63
CH
3
(CH
2
)
3
OH
24
x ∙ 24
C
3
H
6
O
16
16 ∙ 22 = 352
CH
3
(CH
2
)
2
COOH
20
20 ∙ 0,4 = 5
CH
3
COOH
8
8 ∙ 14 = 112
CO
2
0
y ∙ 0 = 0
H
2
2
2 ∙ 135 = 270
C
2
H
5
OH
12
12 ∙ 0,7 = 8,4
SUBSTRATY: γ
i
∙ ν
i
= 2400
PRODUKTY: γ
i
∙ ν
i
= 1053,03 + 24x
2400 = 1053,03 + 24x
x = 56,12
Oznacza to, że ze 100 moli glukozy (18 000g) otrzymujemy 56,12 moli butanolu (4152,88g).
Wynik ten otrzymano bez potrzeby wyznaczana współczynników a oraz y.
Jeśli równanie:
γ
S
– 4b = yγ
X
+ zγ
P
podzielimy przez γ
S
, otrzymamy:
4b/γ
S
+ y γ
X
/γ
S
+ z γ
P
/γ
S
= 1
↑
↑
↑
ε + η + ξ
P
= 1
ε – ułamek ilości elektronów przekazanych na tlen
η – ułamek ilości elektronów przekazanych do biomasy
ξ – ułamek ilości elektronów przekazanych do produktu
Często skład substratu jest nieznany. W takim przypadku metodę obliczania stopnia redukcji
można uzupełnić następującymi, prostymi regułami.
Q
O
= 112,968 kJ/mol
Jest to równoważnik energetyczny 1 mola elektronów przekazanych na tlen.
γ
X
= 4,29 (w przeliczeniu na 1 gramoatom w biomasie)
σ
X
= 0,462g węgla/g suchej biomasy

Dla biomasy o standardowym składzie:
σ
X
= 12/24,6 = 0,488g węgla/g suchej masy
Bilans tlenu – iloraz oddechowy RQ
RQ = ν
C
/ν
O
= [4(1 – y
X/S
– y
P/S
)] / [γ
S
– y
X/S
γ
X
– y
P/S
γ
P
]
Jeśli nie tworzy się produkt: y
P/S
= 0
RQ = [4(1 – y
X/S
)] / [γ
S
– y
X/S
γ
X
]
Iloraz oddechowy dla wzrostu drożdży na:
1. glukozie
C
6
H
12
O
6
+ 6O
2
→ 6CO
2
+ 6H
2
O
RQ = [4(1 – 0,6)] / [4 – 0,6 ∙ 4,2] = 1,08
2. etanolu
C
2
H
5
OH + 3O
2
→ 2CO
2
+ 3H
2
O
RQ = [4(1 – 0,6)] / [6 – 0,6 ∙ 4,2] = 0,49
Przykład
Obliczyć współczynnik wydajności biomasy y
X/S
dla drożdży Pichia sp. hodowanych na a)
glukozie i b) etanolu, wiedząc, że z 1g substratu powstało odpowiednio a) 0,56g suchej masy i
b) 0,61g suchej masy tych drożdży.
c-mol glukozy – 30g
c-mol etanolu – 23g
c-mol biomasy – 25,9g
y
X/S
= Y
X/S
∙ M
S
/M
X
y
X/glukoza
= 0,56 ∙ (30/25,9) = 0,65
y
X/etanol
= 0,61 ∙ (23/25,9) = 0,54
Obliczyć współczynnik zużycia tlenu
a. dla drożdży rosnących na glukozie:
y
O/X
= ¼ (4/0,65 – 4,2) = 0,488 mola tlenu/c-mol biomasy
Y
O/X
= 0,488/25,9 = 0,019 mola tlenu/g suchej masy
b. dla drożdży rosnących na etanolu:
y
O/X
= ¼ (6/0,54 – 4,2) = 1,728 mola tlenu/c-mol biomasy
Y
O/X
= 1,728/25,9 = 0,067 mola tlenu/g suchej masy

Wzrost mikroorganizmów a generowanie ciepła (termodynamika)
Wzrost mikroorganizmów jest wynikiem szeregu powiązanych ze sobą procesów
metabolicznych. Reakcje kataboliczne i anaboliczne są ze sobą sprzężone tak, że energia
wyzwalana w tych pierwszych jest wykorzystywana jako siła napędowa w tych drugich.
Należy pamiętać, że część energii jest tracona (rozpraszana) w postaci ciepła.
W procesach na dużą skalę konieczne jest odprowadzanie (usuwanie) tego ciepła tak, aby
prowadzić hodowlę w temperaturze fizjologicznej.
W małych bioreaktorach ciepło metaboliczne jest łatwo usunąć; w dużych reaktorach
(> 10 000dm
3
), w których zachodzi szybki wzrost mikroorganizmów, konieczne jest
zaprojektowanie efektywnego systemu wymiany ciepła (jego odprowadzania). Temperatura
zawartości bioreaktora musi być utrzymywana z dokładnością do 0,5ºC dla zapewnienia
fizjologicznych warunków wzrostu.
W warunkach wzrostu, gdy powstaje praktycznie wyłącznie biomasa, a ilość powstającego
produktu jest znikoma, reakcję można zapisać równaniem:
C
6
H
12
O
6
+ a NH
3
+ b O
2
→ c CH
1,8
O
0,5
N
0,2
+ e CO
2
+ f H
2
O
Ponieważ zużycie azotu w porównaniu ze zużyciem węgla jest niewielkie, a ponadto azot nie
ulega utlenieniu tak, jak węgiel, równanie uprości się do postaci:
C
6
H
12
O
6
+ b O
2
→ c CH
1,8
O
0,5
N
0,2
+ e CO
2
+ f H
2
O
Bilans cieplny ten reakcji będzie następujący (w przeliczeniu na mole zużywanej glukozy):
Q
(uwalniane ciepło)
= c [(M.cz.
X
) ∙ (-ΔH
X
)] – [(-ΔH
S
) ∙ (M.cz.
S
)]
gdzie (-ΔH
X
) i (-ΔH
S
) to ciepło spalania 1g biomasy lub substratu.
Q/M.cz.
S
= c ∙ M.cz.
X
/M.cz.
S
∙ (-ΔH
X
) – (-ΔH
S
)
Wyrażenie po lewej stronie równania jest ilością ciepła uwalnianego w przeliczeniu na 1g
zużywanego substratu. Natomiast stosunek przyrostu biomasy na jednostkę zużywanego
substratu (pierwszy człon wyrażenia po prawej stronie równania) jest wydajnością biomasy z
substratu.
Równanie przyjmie postać:
Y
Δ/S
= (Y
X/S
)(-ΔH
X
) – (-ΔH
S
)
Po podzieleniu równania przez Y
X/S
:
Y
Δ/X
= (-ΔH
X
) – (-ΔH
S
)/(Y
X/S
)
Podane równania są przydatne do oznaczenia ciepła związanego ze wzrostem biomasy oraz ze
zużywanym substratem. Dane eksperymentalne z szeregu doświadczeń wykazują liniową
zależność pomiędzy ilością uwalnianego ciepła i ilością zużywanego tlenu.

Ciepło spalania biomasy (przykłady):
(-ΔH
S
), kJ/g
Escherichia coli
23,03
Candida lipolytica
21,34
Candida boidinii
20,14
Kluyveromyces fragilis
21,66
Bacillus thuringiensis
22,08
WYKŁAD 7, 29/03/2012
Ustalona doświadczalnie zależność ilości uwalnianego ciepła od ilości zużytego tlenu wynosi
(na 1 mol tlenu):
Y
Δ/O2
= 16,21 kJ/g O
2
Zależność ta pozwala wyliczyć ilości generowanego ciepła na podstawie ilości zużytego tlenu.
W przypadku, gdy oprócz biomasy powstaje dodatkowy produkt, równanie ulegnie
modyfikacji:
Q
(uwalniane ciepło)
= c [(M.cz.
X
) ∙ (-ΔH
X
)] + d [(M.cz.
P
) · (-ΔH
P
)] – [(-ΔH
S
) ∙ (M.cz.
S
)]
gdzie (-ΔH
P
) to ciepło spalania 1 g produktu.
Dzieląc równanie stronami przez M.cz.
S
otrzymamy:
Y
Δ/S
= (Y
X/S
)(-ΔH
X
) + (Y
P/S
)(-ΔH
P
) - (-ΔH
S
)
Dzieląc z kolei przez Y
S/X
, otrzymamy:
Y
Δ/X
= - (ΔH
X
) + (Y
P/X
)( -ΔH
P
) - (-ΔH
S
)/( Y
S/X
)
Y
Δ/S
– wydajność cieplna z substratu
Y
Δ/X
– wydajność cieplna z biomasy
Generowanie ciepła reakcji – przykłady obliczeń
Wartości ciepła reakcji można wyliczyć z dobrym przybliżeniem, znając:
stechiometrię reakcji
wydajność procesu
ciepło spalania biomasy i substratu
Przykład
Należy obliczyć ciepło reakcji hodowli drożdży na glukozie, jako źródle węgla. Równanie
reakcji:
7 CH
2
O + 2,33 O
2
→ C
4
H
6,67
O
2
+ 3 CO
2
+ 3,66 H
2
O
M.cz.
X
= 86,67 g/mol
-ΔH
X
= 1,52·10
3
kJ/100 g
-ΔH
S
= 2,82·10
3
kJ/mol

Zużycie glukozy na węglomol drożdży wynosi:
7 · 30 g = 210 g/c-mol glukozy,
co stanowi:
(210 g)/(180 g) = 1,167 mola glukozy/c-mol drożdży
Wydajność produkcji biomasy:
Y
X/S
= (ΔX)/(ΔS) = (86,67 g)/(210 g) = 0,41
Zatem masa wyprodukowanych komórek drożdży wynosi:
0,41 · 210 g = 86,1 g
Efekt cieplny obliczamy z zależności:
ΔH
reakcji
= 2,82·10
3
kJ/mol · (7 M
CH2O
)/(M
C6H12O6
) - 1,52·10
3
kJ/mol · (86,1 g/100 g) =
= 1,98·10
3
kJ/210 g glukozy = 1,98·10
3
kJ/86,1 g komórek =
= 0,23·10
2
kJ/g komórek
Jeśli skład substratu jest nieznany, to do obliczania efektu cieplnego reakcji można się
posłużyć znajomością stopnia redukcji oraz, wspomnianymi już, prostymi regułami:
Q
O
= 112,97 kJ/mol (równowartość energetyczna 1 mola elektronów przekazanych na O
2
)
γ
X
= 4,29 (w przeliczeniu na gramoatom węgla w biomasie)
σ
X
= 0,462 g C/g suchej masy
Przykład
Ze 100 moli glukozy w obecności tlenu i amoniaku otrzymano 2000 g lizyny i 4000 g
biomasy. Oblicz efekt cieplny reakcji, wiedząc, że stopień redukcji glukozy wynosi w tym
wypadku:
100 · 24 = 2400
Oblicz, jaka część ze stopnia redukcji została przekazana do produktu.
ilość moli lizyny: n
P
= 2000/182,65 = 10,96 mola Lys
γ
P
= 24+14-4-6+1-1 = 28
n
P
· γ
P
= 306,88 moli (część elektronów przekazana do produktu)
Oblicz, jaka część ze stopnia redukcji została przekazana do biomasy.
n
X
· γ
X
= (4000 g · σ
X
· γ
X
)/12 = 660,7 moli
W rezultacie, stopień redukcji zużyty na efekt cieplny X
Q
obliczamy z bilansu:
2400 = X
Q
+ n
P
· γ
P
+ n
X
· γ
X
X
Q
= 1432,42 moli
Pozwala to obliczyć efekt cieplny reakcji:

Q = 112,97 kJ/mol · 1432,42 moli = 161820,5 kJ
Kinetyka wzrostu mikroorganizmów
Z punktu widzenia praktyki – podstawowe znaczenie ma obliczenie:
szybkości namnażania się biomasy drobnoustrojów
szybkości asymilacji składników odżywczych
szybkości wydzielania produktów metabolizmu
W obliczeniu tych parametrów korzystamy z bilansów procesów biochemicznych oraz
równań kinetycznych opisujących przyrost biomasy. We wszystkich ujęciach kinetycznych
rzeczywiste, złożone procesy życiowe komórek oraz rozmnażanie mikroorganizmów
ujmowane są w uproszczeniu i generalnie sprowadzają się do przyrostu biomasy.
Biomasa – ogólnie substancje organiczne komórek mikroorganizmów, roślin lub zwierząt. W
przypadku inżynierii bioprocesowej termin ten odnosi się najczęściej do masy
mikroorganizmów.
Zastosowanie biomasy – jako źródło białka (w postaci paszy lub jako surowiec dla przemysłu
spożywczego). Biomasa często określana jest jako „białko jednokomórkowców” – SCP.
Początki takiego zastosowania – okres I wojny światowej, lata 30 XX wieku, okres II wojny
światowej.
Obecnie przyczyną dużego zainteresowania SCP jest światowy deficyt białka. Na jego
pokrycie nie wystarcza już źródeł tradycyjnych (mączka rybna, sojowa). Uruchomienie
produkcji SCP dla celów paszowych pozwala na zmniejszenie uzależnienia hodowli zwierząt
od importu pasz.
Zalety przemysłu produkcji białek:
szybszy przyrost biomasy mikroorganizmów w porównaniu z roślinami i zwierzętami
wyższa zawartość białka w suchej masie biomasy
szeroki wachlarz utylizowanych surowców, najczęściej odpadowych
wysoka wydajność, niewielka przestrzeń zajmowana przez instalacje produkcyjne
niezależność od zmian pogody, klimatu i sezonowości
możliwość wzbogacania produktu żywnościowego w niezbędne składniki, np.
aminokwasy egzogenne (Val, Leu, Ile, Phe, Tyr, Trp, Lys, Met, Thr), które występują
w niedomiarze w produktach roślinnych

biomasa jest nie tylko źródłem białka, lecz także związków biologicznie czynnych, np.
witamin
Porównanie szybkości przyrostu biomasy różnych organizmów:
ORGANIZM
Czas podwojenia biomasy
Bakterie
1 – 7 godzin
Glony
6 – 50 godzin
Trawy
1 – 2 tygodnie
Kura
3 – 4 tygodnie
świnia
4 – 6 tygodni
Zawartość białek w różnych produktach naturalnych:
SUROWIEC
ZAWARTOŚĆ BIAŁKA
Grzyby mikroskopowe
31 – 50%
Mikroalgi
47 – 63%
Drożdże
47 – 53%
Bakterie
72 – 78%
Mleko
22 – 25%
Wołowina
81 – 90%
Jaja
31 – 38%
Ryż
8 – 9%
Surowce do produkcji biomasy:
SUROWIEC
ŹRÓDŁO
C-1 metan
Gaz naturalny, węgiel kamienny, gazyfikacja
drewna, proces dygestii
C-1 metanol
Gaz syntezowy
C-2 etanol, kwas octowy
Fermentacja alkoholowa, etylen
Modele wzrostu mikroorganizmów
Wyróżnia się następujące grupy modeli:
model wzrostu ustalonego oraz model wzrostu nieustalonego; podział ten wynika z
uwzględnieniem lub nie warunków ustalonych w przemianach metabolicznych;
najczęściej przyjmuje się założenia o ustalonych warunkach wzrostu

modele ciągłe lub korpuskularne; kryterium podziału jest w tym wypadku
uwzględnienie lub nie „ziarnistej struktury biomasy”, tzn. występowanie wydzielania
żywych komórek; w ujęciu ciągłym zamiast populacji mikroorganizmów rozpatruje
się pewną jednorodność (homogenność), „biofazę”, co pozwala na uniknięcie opisu
zjawisk związanych z istnieniem indywidualnych mikroorganizmów
model strukturalny i niestrukturalny; w tym podziale rozpatrywany jest sposób ujęcia
przemian
metabolicznych;
w
modelu
niestrukturalnym
mikroorganizmy
reprezentowane są jednym parametrem – biomasą; wzrost jest rozpatrywany jako
przemiana substratu bezpośrednio w biomasę
Najpowszechniej używa się modelu najprostszego – niestrukturalnych, ciągłych modeli
wzrostu ustalonego.
Drożdże Saccharomyces cerevisiae:
dobrze poznany genom
dobrze zaprojektowane metody genetyczne
model dla innych eukariontów
nie produkuje toksyn; organizm GRAS
około 6000 genów
zdolność do wydzielania białek
możliwość modyfikacji potranslacyjnych
Wzrost wykładniczy mikroorganizmów
X – liczba komórek
n – liczba podziałów komórkowych
X = 2
n
dX/dt = αX = μX
α – współczynnik proporcjonalności
μ – szybkość właściwa wzrostu
Można wyznaczyć stosunek stężenia biomasy do czasu poprzez scałkowanie równania w
przedziale czasu od t
0
do t
1
1/X dX = μ dt
Jeśli w czasie t
0
stężenie biomasy w reaktorze wyrazimy jako X
0
, a w czasie t
1
stężenie
biomasy wyrazimy jako X
1
, to równanie przyjmie postać:

∫ 1/X dX = μ ∫ dt
Całkując obie strony otrzymujemy:
lnX
1
– lnX
0
= μ (t
1
– t
0
)
ln(X
1
/X
0
) = μ (t
1
– t
0
)
Zatem stężenie biomasy po określonym czasie hodowli opisuje równanie:
X
1
= X
0
· e
μ(t1-t0)
Czas podwojenia (okres generacji) a szybkość właściwa wzrostu
Czas podwojenia (t
d
, t
g
, τ
d
) jest wyrażeniem stosowanym powszechnie przez mikrobiologów
do opisu szybkości wzrostu komórek. Jest to czas potrzebny do podwojenia się liczby
komórek; w fazie wykładniczej jest to wartość w przybliżeniu stała. Stosunek między
szybkością właściwą wzrostu (μ) i czasem podwojenia (t
d
) jest następujący:
X
1
→ 2X
1
w czasie t
d
= t
2
– t
1
t
d
= ln2/μ
Przykład
X
0
= 2 g/dm
3
X
1
= 15,96 g/dm
3
t = 90 min
μ = (lnX
1
– lnX
0
)/Δt
μ = 0,023/min = 1,3847/h
t
d
= ln2/μ = 0,69/1,38 = 0,5 h
Zgodnie z modelem wzrostu wykładniczego biomasa będzie ciągle rosła. W rzeczywistości,
dostępność pokarmu nie jest nieograniczona i w chwili, gdy następuje jego ograniczenie,
szybkość wzrostu komórek obniży się lub nawet wzrost ustanie. Powstające produkty
metabolizmu również mogą hamować wzrost. Wzrost wykładniczy nie będzie miał miejsca
także w przypadku niedoboru tlenu lub w przypadku nieodpowiedniego mieszania. W tych
przypadkach wzrost będzie bardziej ograniczony wymianą mas niż limitowany kinetyką.
Ograniczenia w wymianie mas są głównym czynnikiem limitującym wzrost
mikroorganizmów w dużych bioreaktorach.
Nie-wykładniczy wzrost ma miejsce również w przypadku, gdy podziały komórkowe nie
zachodzą ze stałą szybkością. Takie zjawisko jest powszechne w przypadku grzybów
pleśniowych, organizmów wielokomórkowych, niektórych organizmów rozmnażających się
przez pączkowanie oraz wielu typów komórek zwierzęcych.

Powszechnie popełnianych błędem przy analizie procesów fermentacyjnych jest przyjęcie
zależności logarytmicznych, podczas gdy nie występują one w rzeczywistości.
Przemiana podstawowa
SUBSTRAT → WZROST BIOMASY + PODTRZYMYWANIE FUNKCJI ŻYCIOWYCH
Podtrzymywanie funkcji życiowych
MODEL HERBERTA
MODEL PIRTA
Na przemiany podstawowe zużywana jest
część biomasy
Na przemiany podstawowe zużywana jest
część substratu
r
e
= μ
e
· C
X
r
e
= m
s
· C
S
μ
e
– właściwa szybkość przemian
egzogennych
m
s
– współczynnik przemian podstawowych
r
X
= Y
X/S
· r
S
- μ
e
· C
X
r
S
= (1/Y
X/S
) · r
X
+ m
s
· C
S
r
e
– szybkość przemiany podstawowej
r
S
– szybkość asymilacji substratu
WYKŁAD 8, 5/04/2012
Wzrost komórek/przyrost biomasy:
pojęcie przemiany podstawowej
modele Herberta i Pirta
model Herberta – część biomasy jest zużywana na przemianę podstawową
(utrzymanie metabolizmu endogennego); szybkość takiego zużywania biomasy w
procesie endogennym jest proporcjonalna do stężenia biomasy
Ponieważ dla opisu procesów zachodzących w bioreaktorze podstawową miarą jest określenie
przyrostu biomasy; w tym celu istotny jest opis wzrostu pojedynczych komórek. Analiza taka
ma na celu sprawdzenie:
czy nie występują ograniczenia wzrostu
czy nie można przewidzieć ogólnego charakteru zależności dla wzrostu populacji
mikrobów
Zakładamy, że szybkość asymilacji substratu jest zależna od:
stężenia tego substratu
powierzchni komórki
substratem limitującym jest źródło węgla; należy uwzględnić zużycie części substratu
na przemianę podstawową.

dm
K
/dt = a
K
J
S
- μ
e
m
K
m
K
– masa komórki
a
K
– powierzchnia właściwa komórki
J
S
– strumień asymilowanego substratu
μ
e
– właściwa szybkość metabolizmu endogennego
Zależność pomiędzy masą a powierzchnią komórki wynika z kształtu komórki i sposobu jej
wzrostu. W praktyce wyróżniamy dwa przypadki: komórki kuliste i komórki pałeczkowate
(walcowate).
Komórki pałeczkowate – średnica jest stała, wzrost następuje przez wydłużanie się
powierzchni (a
K
). Dla stałego strumienia asymilowanego substratu, właściwa szybkość
wzrostu (μ) jest stała w warunkach ustalonych. Nie będą występowały ograniczenia wzrostu
pojedynczych komórek, ale też np. grzybni.
μ = 4J
S
/d
X
ρ
X
– μ
e
Komórki kuliste – wzrost poprzez powiększanie średnicy. Właściwa szybkość wzrostu
wynosi w tym przypadku:
μ = J
S
[(4Π)
1/3
· (3/ρ
X
)
2/3
] m
K
-1/3
– μ
e
Dla komórek kulistych właściwa szybkość wzrostu maleje wraz ze wzrostem średnicy,
graniczną szybkość wzrostu określa równanie:
m
K,gr
= (36 · Π · J
S
3
)/(ρ
X
· μ
e
3
)
co oznacza taką wielkość, dla której strumień substratu asymilowanego przez jej
powierzchnię jest zużywany wyłącznie na przemianę podstawową (geometryczne
ograniczenie wzrostu).
Czynniki wpływające na wydajność wzrostu mikroorganizmów:
temperatura
pH
dostępność substratu (źródło węgla i azotu)
zapotrzebowanie na tlen i jego dostarczanie
szybkość mieszania zawiesiny komórek
sposób mieszania

Model Monoda – niestrukturalny, ciągły model ustalonego wzrostu w fazie wzrostu
wykładniczego.
μ = (μ
max
· S)/(K
S
+ S)
K
S
– stała nasycenia (mg/dm
3
)
μ
max
– maksymalna właściwa szybkość wzrostu
Podobieństwa między równaniem Monoda i równaniem Michaelisa-Menten dotyczą kształtu
krzywej, postaci funkcji i sposobu wyznaczania parametrów.
Hodowla okresowa
system zamknięty, dobrze mieszany, o stałej objętości
dostępność substratu jest czynnikiem limitującym
wzrost pod koniec fermentacji
po wejściu komórek w fazę stacjonarną wzrostu należy pamiętać, że także inhibicja
przez powstający produkt końcowy prowadzi do zahamowania wzrostu
w typowych warunkach w fazie logarytmicznej stężenie substratu limitującego jest
znacznie wyższe niż K
S
Wyznaczanie K
S
w hodowlach okresowych jest trudne i obarczone błędem, zwłaszcza, jeśli stosuje się
metody bezpośrednie
teoretycznie lepiej zastosować metodę wyznaczania K
S
podobną do wyznaczania
wartości K
m
(Lineweavera-Burka, Eadie-Hofstee); metody te dają również wynik
obciążony błędem w przypadku hodowli okresowych
wiarygodne wyniki uzyskuje się w przypadku hodowli ciągłych (w chemostacie).
Model Monoda – może służyć do opisu (z pewnym przybliżeniem) większości procesów
zachodzących w hodowlach komórkowych. Model ten może być łatwo zmodyfikowany celem
opisu bardziej złożonych zjawisk, m.in. do opisu inhibicji (przez produkt końcowy lub nawet
przez powstającą biomasę). Należy pamiętać, że szereg substratów w wyższych stężeniach
może być toksyczny dla mikrobów (metanol, fenol, aminy).

WYKŁAD 9, 12/04/2012
Równanie Andrewsa:
Równanie Levenspiela:
Pod K
I
można podstawić S (jeśli substrat hamuje wzrost biomasy) lub X (jeśli biomasa
hamuje wzrost biomasy).
Równanie Monoda:
Istnieją mikroorganizmy, których zapotrzebowanie na substrat jest niskie (rosną lepiej przy
niskim stężeniu substratu w pożywce) oraz mikroorganizmy, których zapotrzebowanie na
substrat jest wysokie.
Z powodów ekonomicznych dąży się do zwiększenia współczynnika wydajności fermentacji,
gdyż prowadzi to do obniżenia zużycia surowców przy zwiększeniu ilości produktów, lecz:
nie zawsze prowadzi to do zwiększenia zysków
nie zawsze prowadzi to do zwiększenia produktywności (szybkości tworzenia danego
produktu)
W dużych fermentatorach, podczas procesów aerobowych, tlen jest czynnikiem limitującym
wzrost. Zwiększenie napowietrzania może zwiększyć współczynnik wydajności. W pewnych
sytuacjach zwiększenie napowietrzenia powoduje znaczny wzrost zużycia mocy i wzrost
kosztów może przeważać nad zyskami.
Jeśli w zmodyfikowanym procesie zwiększono współczynnik wydajności, lecz szybkość
powstawania produktów obniżyła się (wydłużył się czas), produktywność obniżyła się.
Jedynie w wypadku, gdy zwiększeniu współczynnika wydajności Y
X/S
lub Y
P/S
towarzyszy
zwiększenie szybkości tworzenia produktu – całkowita produktywność będzie większa.
Rodzaje współczynników wydajności
W procesach fermentacyjnych używa się zasadniczo dwóch podstawowych współczynników
wydajności: Y
X/S
i Y
P/S
(biomasy z substratu i produktu z substratu).

Współczynnik wydajności biomasy jest stosunkiem średniej masy biomasy do masy
wykorzystanego substratu. W hodowli okresowej Y
X/S
oblicza się z wyrażenia:
Y
X/S
= ΔX/ΔS
Analogicznie wyznacza się współczynnik wydajności produktu Y
P/S
:
Y
P/S
= ΔP/ΔS
Niekiedy wydajność produktu podaje się w stosunku do biomasy:
Y
P/X
= ΔP/ΔX
Ten sposób jest przydatny do oceny ilościowej produkcji wtórnych metabolitów.
Proste matematyczne modele procesów fermentacji
Poznaliśmy już model opisujący tworzenie biomasy równaniem:
dX/dt = (μ
max
· S)/(K
S
+ S) · X
Aby model opisywał cały proces fermentacyjny należy w równaniach uwzględnić
wykorzystanie substratu i tworzenie produktu.
Przy założeniu, że powstawanie produktu i biomasy jest bezpośrednio powiązane z
wykorzystaniem substratu (jego ubytkiem) poprzez współczynniki wydajności, można
napisać:
dX/dt = -Y
X/S
· dS/dt
dP/dt = -Y
P/S
· dS/dt
Cały proces fermentacyjny/wzrost mikroorganizmów można opisać trzema równaniami:
dX/dt = (μ
max
· S)/(K
S
+ S) · X
dS/dt = -1/Y
X/S
· (μ
max
· S)/(K
S
+ S) · X
dP/dt = Y
P/S
/Y
X/S
· (μ
max
· S)/(K
S
+ S) · X
Ostatnie równanie jest prawdziwe tylko wtedy, gdy przyrost produktu jest proporcjonalny do
przyrostu biomasy, czyli przyrost produktu jest powiązany ze wzrostem. W procesie
fermentacji często taka zależności nie występuje.
Opisane równania rozwiązuje się zasadniczo metodami numerycznymi, rozwiązując równania
różniczkowe.
Znajomość stałych pozwala na wyznaczenie znormalizowanej kinetyki τ(S). Zmienność X, S
i P jest opisana układem następujących równań różniczkowych.
τ(S) = S/(K
S
+ S)
dX/dt = μ
max
· τ(S) · X
dS/dt = -1/Y
X/S
· μ
max
· τ(S) · X
dP/dt = Y
P/S
· μ
max
· τ(S) · X

W oparciu o model kinetyczny można uzyskać ważną informację – mianowicie określić
długość czasu fermentacji (informacji takiej nie uzyska się w oparciu o wyznaczenie
współczynnika wydajności). Dzięki temu możliwe jest obliczenie ilości cykli produkcyjnych
na jednostkę czasu, a tym samym oszacowanie zysków.
Zalety i wady hodowli okresowych:
proste, dobrze przetestowane rozwiązania
łatwość prowadzenia procesu
względna łatwość zapobiegania zakażeniom
cykl wzrostu jest mało wydajny (faza zwłoki – lag)
jeśli pożądany produkt powstaje tylko w określonej fazie wzrostu, pozostałe fazy
powodują stratę czasu
wymagane jest przygotowanie stosunkowo dużej ilości materiału do zaszczepienia
(inokulum)
konieczność mycia i sterylizacji bioreaktora i urządzeń towarzyszących po każdym
cyklu produkcyjnym (szarży)
Hodowle ciągłe – chemostat Monoda
Reaktory do hodowli ciągłych na dużą skalę mają największe zastosowanie w oczyszczaniu
ścieków:
nie ma konieczności stosowania czystych kultur, nie ma więc zagrożenia zakażeniem
wieloletnia praktyka wykazała, że stosowanie takich oczyszczalni nie stwarza
zagrożenia dla ludzi ani środowiska
z ekonomicznego punktu widzenia, biorąc pod uwagę wielkie objętości utylizowanych
ścieków jest to jedyne sensowne rozwiązanie
Poza oczyszczaniem ścieków, reaktory do hodowli ciągłych na bardzo dużą skalę są używane
do produkcji białka jednokomórkowców (SCP) do celów paszowych. Przykładem może być
wielki bioreaktor koncernu ICI (USA) o objętości 1 000 000 dm
3
, w którym metanol
przekształcany jest przez Methylophilus methylotropus w biomasę.
Charakterystyczne parametry:
szybkość rozcieńczania (D)
czas przebywania (τ)

D = (szybkość przepływu, dm
3
/h) / (objętość reaktora, dm
3
) = F/V (h
-1
)
τ = (objętość reaktora, dm
3
) / (szybkość przepływu, dm
3
/h) = V/F (h)
Szybkość przepływu:
objętościowa
F = V/t
masowa (szybkość przepływu masy)
m = (ρ · F)/t
Produkcja biomasy oraz produktu reakcji:
tworzenie tych składników
usuwanie tych składników
dX/dt = μX – DX
dP/dt = Y
P/S
/Y
X/S
· μX – DP
Wyrażenia określające akumulację substratu zawierają trzy składniki:
wykorzystanie substratu przez mikroorganizmy
dopływ substratu (zasilanie)
usuwanie substratu w odpływie
dS/dt = -1/Y
X/S
· μX + DS
0
– DS
Model matematyczny chemostatu:
dla biomasy:
dX/dt = [(μ
max
· S)/(K
S
+ S)] · X – DX
dla substratu limitującego:
dS/dt = -1/Y
X/S
· [(μ
max
· S)/(K
S
+ S)] · X + D(S
0
– S)
dla produktu:
dP/dt = Y
P/X
· [(μ
max
· S)/(K
S
+ S)] · X – DP
WYKŁAD 10, 19/04/2012
W stanie równowagi:
dX/dt = 0
dS/dt = 0
dP/dt = 0
Zatem:
μX = DX → μ = D
W chemostacie, w stanie równowagi, szybkość właściwa wzrostu może być kontrolowana
przez szybkość rozcieńczania.
Relacje pomiaru szybkości rozcieńczania (D) i stężenia substratu (S): jeżeli μ = D, równanie
Monoda przyjmie postać:
S = (DK
S
)/(μ
max
– D)
Stan równowagi w chemostacie a stężenie biomasy:
dS/dt = -1/Y
X/S
· μX + D(S
0
– S) = 0
X = Y
X/S
(S
0
– S)
X = Y
X/S
[S
0
– (DK
S
)/(μ
max
– D)]
Stan równowagi w chemostacie a stężenie produktu:
dP/dt = 0 = Y
P/X
· μX – DP
P = Y
P/S
(S
0
– S)
P = Y
P/S
[S
0
– (DK
S
)/(μ
max
– D)]
Wszystkie przedstawione równania stosuje się tylko do sytuacji, gdy powstawanie produktu
powiązane jest ze wzrostem.
Powstawanie produktu jest związane
ze wzrostem
Powstawanie produktu nie jest związane
ze wzrostem
Wpływ szybkości rozcieńczania:
wymywanie komórek mikroorganizmów
dX/dt = μX – DX
μX – przyrost biomasy
DX – ubytek biomasy
Równanie równowagowe mas biomasy posiada dwa składniki:
składnik tworzenia (wzrostu) biomasy: μX
składnik usuwania biomasy: DX
Jeżeli szybkość usuwania biomasy przewyższa szybkość jej powstawania (D>μ):
Przy zwiększaniu szybkości rozcieńczania (D) wzrost biomasy spada, przez co
wykorzystywane jest mniej substratu (będzie go więcej w reaktorze). Jeżeli D>μ, nastąpi
wymycie mikroorganizmów. Hodowle ciągłe rozpatrywane były w stanie osiągniętej
równowagi dynamicznej, w której D = μ.
Komórki nie mogą rosnąć szybciej niż wynosi ich maksymalna szybkość wzrostu (lub nie
może być ona określana jako maksymalna). Rozważając równanie dla chemostatu, nie można
mieć ani nieograniczonego, ani ujemnego stężenia substratu.
μ
max
– D ≠ 0 ^ μ
max
– D > 0
Gdy szybkość rozcieńczania jest wyższa niż szybkość właściwa wzrostu, stężenie biomasy w
reaktorze będzie spadać, aż do całkowitego wymycia mikroorganizmów. Szybkość
rozcieńczania, przy której następuje wymywanie mikroorganizmów określamy jako szybkość
krytyczną rozcieńczania (D
kryt
), równą μ
max
.
WNIOSKI:
1. Szybkość rozcieńczania w chemostacie musi być mniejsza od krytycznej wartości
rozcieńczania (D < D
kryt
).
2. Równania stosowane do obliczenia stężeń w stanie równowagi dynamicznej nie mają
zastosowania w przypadku warunków wymywania.
3. W przypadku wymywania komórek z bioreaktora, zmienne przyjmą wartości:
S = S
0
X = 0
P=0
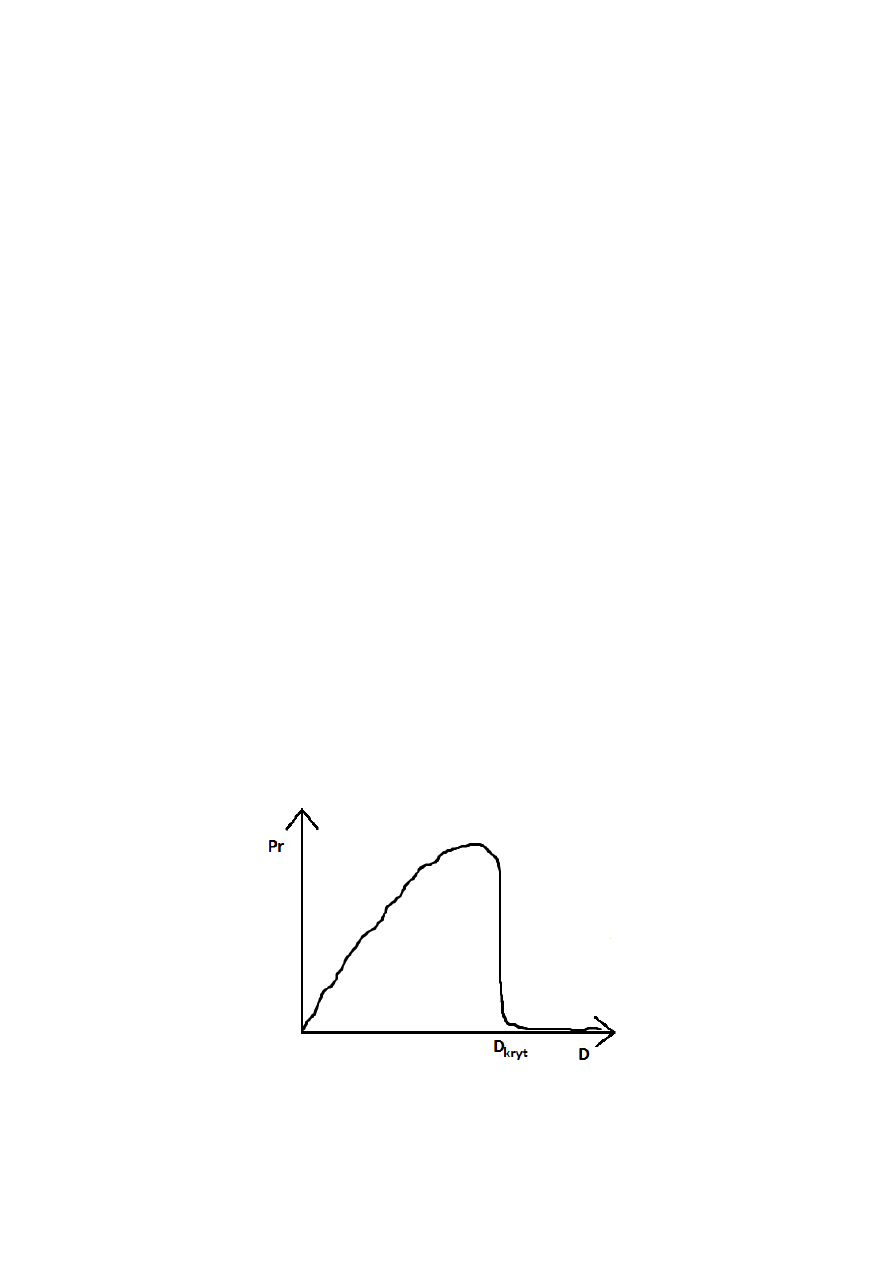
Produktywność w hodowlach ciągłych
Produktywność definiowana jest jako ilość wytworzonego danego składnika na jednostkę
czasu. W przypadku reaktora przepływowego, produktywność (Pr) będzie równa szybkości
przepływu masy składnika.
Dla biomasy:
Pr
X
= m
X
= F · X
Dla produktu:
Pr
P
= m
P
= F · P
W oparciu o model Monoda, produktywność tworzenia biomasy można wyliczyć z równania:
Pr
X
= FY
X/S
[S
0
– (DK
S
)/(μ
max
– D)]
Wybór optymalnej szybkości rozcieńczania
Jeśli wydajność produkcji biomasy jest stała przy różnych szybkościach rozcieńczenia, to
produktywność (Pr) biomasy będzie rosła wraz ze zwiększaniem szybkości rozcieńczania aż
do osiągnięcia wartości D
kryt
, to jest aż do momentu wymycia mikroorganizmów.
W rzeczywistości ani wydajność produkcji biomasy, ani produktu nie jest stała. Stwierdzono
ponadto, że często wydajność tworzenia szeregu ważnych komercyjnie produktów jest
największa wtedy, gdy wzrost jest najwolniejszy. Zatem stosowanie wysokim wartości
szybkości rozcieńczania nie zawsze jest najkorzystniejszą opcją dla tworzenia produktu
(często jest to opcja najmniej korzystna). Należy również wziąć pod uwagę, że zwiększenie
szybkości rozcieńczania (D) prowadzi do zwiększenia stężenia substratu w wypływie.
Zależność produktywności od szybkości rozcieńczania w hodowlach ciągłych:

Równania modelu Monoda można przekształcić tak, aby otrzymać współczynniki wydajności.
Wyznaczanie stałych kinetycznych dla modelu Monoda (transformacja Lineweavera-Burka
dla modelu Monoda):
Szybkość rozcieńczania, dla której produktywność wynosi swoją maksymalną wartość,
nazywamy D
max
. D
max
można obliczyć ze wzoru:
Przy D
max
(lub D
m
) następuje częściowa utrata substratu. D
max
jest bardzo blisko D
kryt
.
Współzawodnictwo mikroorganizmów w chemostacie
Jeśli dwa mikroorganizmy współzawodniczą o ten sam substrat limitujący wzrost w
chemostacie, przeżyje ten, mikroorganizm, który będzie utrzymywał niższe stężenie substratu
w mieszaninie fermentacyjnej.
Jest tak, ponieważ w reaktorze nie może być dwóch różnych stężeń tego samego substratu. W
przypadku współzawodnictwa dwóch mikroorganizmów utrzymywane jest niższe stężenie
substratu (tzn. wygrywa ten mikroorganizm utrzymujący to niższe stężenie). Można obliczyć
właściwą szybkość wzrostu obu mikroorganizmów przy niższym stężeniu substratu.
WYKŁAD 11 → nie mam, bardzo mi przykro…
WYKŁAD 12, 10/05/2012
METABOLIZM
PIERWOTNY
WTÓRNY
TROPOFAZA
podczas zrównoważonego wzrostu (w
sytuacji nadmiaru pożywienia)
IDIOFAZA
przy braku dostatecznej ilości pożywienia
Metabolity wytwarzane podczas idiofazy z acetylo-CoA:
cytrynian
kwasy tłuszczowe
chinony
terpeny
poliketydy (makrolidy)
sterole (steroidy) i inne
Kolorem czarnym zaznaczona biomasa, kolorem czerwonym – metabolit pierwotny, zaś kolorem zielonym
metabolit wtórny.
Tworzenie biomasy i produktu – zależności metaboliczne:
wytwarzanie energii w postaci ATP
wytwarzanie materiału budulcowego
Produkcja biomasy jest efektem skierowania części ATP do syntezy materiału komórkowego.
ATP jest również niezbędny do zapewnienia innych funkcji: homeostazy i ruchu.
Homeostaza jest utrzymywaniem wewnątrzkomórkowej integralności. Obejmuje m.in.
aktywny transport na zewnątrz komórki końcowych produktów metabolizmu, jonów wodoru,
toksycznych składników.
Przełączanie szlaków katabolicznych organizmu powoduje również zmiany wydajności
produkcji biomasy. Szereg organizmów (komórek) ma zdolność fermentacji (proces
beztlenowy), jak i potrafi oddychać (proces tlenowy):
fermentacja (glikoliza beztlenowa):
GLUKOZA → 2 ATP
oddychanie (glikoliza, cykl kwasów trójkarboksylowych, łańcuch oddechowy):
GLUKOZA → CO
2
+ H
2
O + 30-38 ATP
Z równań tych wynika, że fermentacja prowadzi do niższej wydajności produkcji biomasy,
niż oddychanie.
Dwa główne czynniki mogą wymuszać na fakultatywnych beztlenowcach fermentację:
ograniczony dostęp tlenu (efekt Pasteura)

wysokie stężenie rzeczywiście degradowanych substratów (efekt Crabtree lub efekt
glukozy)
W warunkach braku tlenu lub innych związków nieorganicznych podlegających redukcji,
fakultatywne beztlenowce przełączają się na fermentację.
Z porównania schematów fermentacji i oddychania widać, że odtwarzanie NAD
+
w
przypadku fermentacji jest prostsze i szybsze niż w przypadku oddychania. Zdolność
przełączania się z oddychania na fermentację daje przewagę fakultatywnym beztlenowcom
nad bezwzględnymi tlenowcami w kulturach mieszanych, np. w ekosystemach.
Efekt Crabtree:
wiele bakterii prowadzących procesy beztlenowe podlega temu efektowi bardzo
wyraźnie
przy wysokim stężeniu substratu organizmy te produkują mleczan z dużą wydajnością;
bakterie fermentacji homomleczanowej przekształcają glukozę w mleczan prawie
stechiometrycznie w warunkach glikolizy beztlenowej
pierwszorzędnym celem komórek jest wykorzystanie substratu możliwie jak
najszybciej; jeżeli komórka oddycha, szybkość utlenienia NADH może być niższa niż
aktualnie potrzebna – następują wówczas zaburzenia szlaku glikolizy, zaczynają
gromadzić się metabolity, zwłaszcza fruktozo-1,6-bisfosforan i pirogronian
metabolity te są równocześnie aktywatorami kluczowych enzymów uczestniczących w
glikolizie beztlenowej, jak również inhibitorami enzymów odpowiadających za
przekształcenie pirogronianu w acetylo-CoA (inhibitory kompleksu dehydrogenazy
pirogronianowej)
Metabolity wtórne:
szereg enzymów (najczęściej rekombinowanych)
przeciwciała monoklonalne
pigmenty
toksyny
alkaloidy
antybiotyki (wiele spośród nich produkowanych jest tylko w fazie stacjonarnego
wzrostu)
Końcowe produkty fermentacji – mleczan, etanol, cytrynian, octan.

Rozprzężenia metaboliczne
Nagromadzenie się końcowych produktów metabolizmu prowadzi do tzw. rozprzężenia
metabolizmu. Zwiększa się gradient stężeń i komórka jest zmuszona do usuwania produktów
metabolizmu na zewnątrz. W tej sytuacji znacznie więcej ATP jest potrzebne do utrzymania
homeostazy – tym samym mniej może być zużyte do syntez. W tym czasie komórka będzie
produkowała więcej końcowych metabolitów w procesie syntezy ATP. Na koniec produkcja
biomasy ustanie całkowicie, a będą powstawać tylko końcowe metabolity.
Przenoszenie masy – wpływ na wzrost
Dotychczasowe rozważania dotyczące wzrostu mikroorganizmów w różnych systemach
fermentacyjnych (hodowle okresowe, półokresowe, ciągłe) charakteryzowały się szeregiem
uproszczeń. Generalnie założono, że:
szybkość wzrostu komórek w reaktorze jest zależna od charakterystyki kinetycznej
wymieszanie jest idealne (zakłada się, że w takim modelu chemostatu skład odpływu
jest identyczny, jak skład zawartości reaktora; składniki w dopływie, wchodzące do
reaktora są natychmiast równomiernie rozpraszane w całej objętości cieczy w
reaktorze)
wszystkie składniki są roztworami wodnymi
Przedstawione uproszczenia pomijają jeden ważny fakt – przenoszenie masy pomiędzy oraz
wewnątrz faz nie jest natychmiastowe!!! W praktyce wykazano wielokrotnie, że:
kinetyka charakteryzująca biokatalizator nie zawsze jest czynnikiem ograniczającym
całkowitą szybkość reakcji
mieszanie nie zawsze jest bliskie wymieszaniu idealnemu, jak również substrat lub
biokatalizator nie zawsze są w postaci wodnej
transfer cząsteczek zachodzi pomiędzy różnymi fazami
Dobrym przykładem jest transfer tlenu. Tlen atmosferyczny jest dostarczany w postaci
gazowej – w pęcherzykach powietrza. Gazowy tlen musi zostać przetransportowany do fazy
ciekłej, w której ulega rozpuszczeniu – i tylko w takiej postaci jest dostarczany do komórek.
Całkowita szybkość reakcji jest często determinowana nie przez kinetykę katalizy, lecz przez
szybkość przenoszenia mas. W każdej reakcji musi występować ruch mas – jeśli ruch ten
będzie wolny, to i szybkość reakcji będzie niska. Każda reakcja katalizowana obejmuje:
etap przenoszenia mas

etap katalizy
Etap przenoszenia mas może być wolny, jeśli ruch cząsteczek katalizatora/substratu jest
ograniczony lub jeśli stężenie katalizatora/substratu jest niskie. Etap katalizy zależy w dużym
stopniu od czynników środowiska: temperatury, pH i innych.
Rozważając oba etapy:
jeśli szybkość przenoszenia mas jest tak niska, że ogranicza szybkość całkowitą
reakcji – mówimy, że reakcja jest ograniczana przenoszeniem mas
jeśli szybkość przenoszenia mas jest wyższa od szybkości katalizy – reakcja jest
ograniczana katalitycznie
Przenoszenie mas w roztworach
Bez mieszania wymuszonego ruch rozpuszczonych związków w roztworze zachodzi dzięki
spontanicznej dyfuzji lub naturalnej konwekcji. Procesy te są zbyt wolne, aby osiągnąć
wystarczającą szybkość przenoszenia mas dla większości reakcji. Wymuszona konwekcja
(mieszanie) jest zatem powszechnym sposobem na zwiększenie szybkości przenoszenia mas,
przy zastosowaniu różnego typu mieszadeł, pomp lub kompresorów.
Szybkość przenoszenie mas w roztworze zależy od:
intensywności mieszania
lepkości (μ) i gęstości (ρ) cieczy
Z kolei intensywność mieszania zależy od:
kształtu mieszadeł i reaktora
mocy mieszadła
Lepkość i gęstość cieczy zależą od składu roztworu i jego temperatury.
Wraz ze wzrostem intensywności mieszania rosną koszty spowodowane rosnącym zużyciem
energii oraz pojawiają się problemy z destrukcją komórek, enzymów, a niekiedy produktów.
Zatem te czynniki mogą ograniczać stosowanie optymalnych szybkości mieszania.
Odpowiednie zaprojektowanie systemu mieszania oraz reaktora może przyczynić się do
sukcesu lub niepowodzenia całego bioprocesu. Należy pamiętać, że w dużych bioreaktorach
mogą występować martwe strefy, które znacząco wpłyną na obniżenie wydajności.

Szybkość reakcji rośnie wraz ze wzrostem stężenia substratu. W przypadku, gdy substrat
dostarczany jest w postaci niewodnej, musi najpierw ulec rozpuszczeniu. W tym przypadku
stężenie rozpuszczonego w wodzie substratu (a nie jego stężenie całkowite) będzie określało
kinetykę katalizy.
Przenoszenie mas gaz-ciecz jest ważne w procesach biotechnologicznych i dotyczy głównie
transportu tlenu. Obejmuje również procesy usuwania CO
2
oraz procesy destylacji produktów
lotnych.
Przenoszenie mas ciecz-ciecz ma znaczenie w przypadku reakcji w układach dwufazowych,
konwersji związków hydrofobowych (oleje, alkany) oraz w procesach ekstrakcji.
Przenoszenie mas ciało stałe-ciecz ma miejsce w przypadku stosowania osadu komórek
bakterii, komórek immobilizowanych i immobilizowanych enzymów. Transfer taki jest
istotnym czynnikiem w procesach adsorpcji, wymiany jonowej czy też krystalizacji.
Ponieważ ruch cieczy w warstwie granicznej jest wolny, ruch rozpuszczonych związków
poprzez tę warstwę będzie determinowany raczej przez siły dyfuzji, a nie konwekcji, zatem
szybkość tej dyfuzji będzie czynnikiem limitującym.
Etapy transferu rozpuszczonego związku między fazami:
1. rozpuszczony związek wędruje przez pęcherzyk i osiąga granicę faz
2. rozpuszczony związek wędruje przez granicę faz
3. związek wędruje przez warstwę międzyfazową do cieczy otaczającej
Ponieważ szybkość ruchu rozpuszczonego związku przez warstwę międzyfazową jest
określana tylko przez dyfuzję, etap trzeci uważany jest powszechnie za etap ograniczający
szybkość reakcji.
Czynniki wpływające na proces przenoszenia mas między fazami
Ruch cząsteczek poprzez warstwę międzyfazową jest determinowany przez dyfuzję i
zachodzi od wysokiego do niskiego stężenia. Szybkość dyfuzji jest determinowana
gradientem stężeń w warstwie międzyfazowej.
Szybkość można zwiększyć poprzez:
zwiększenie powierzchni międzyfazowej
zwiększenie gradientu stężeń w poprzek warstwy międzyfazowej
obniżenie grubości warstwy międzyfazowej

zwiększenie szybkości ruchu cząsteczek w poprzek warstwy międzyfazowej
Mieszanie a przenoszenie mas między fazami:
intensywne mieszanie wytwarza silne siły ścinające, które rozdrabniają pęcherzyki
gazu i krople
następuje szybsze usuwanie rozpuszczonego związku z warstwy międzyfazowej i
utrzymanie gradientu stężeń
następuje spadek grubości warstwy międzyfazowej
WYKŁAD 13, 17/05/2012
Szybkość przenoszenia mas jest determinowana przez dwa czynniki:
mieszanie
dyfuzję
Opisy matematyczne, dotyczące tego procesu w bioreaktorach z wymuszonym mechanizmem
muszą zatem uwzględniać:
szybkość mieszania
sposób zaprojektowania reaktora
średnicę turbiny
gęstość i lepkość medium
gęstość pęcherzyków
szybkość dyfuzji
W obliczeniach inżynierskich korelacje te charakteryzują różna wartości bezwymiarowe, np.
liczba Reynoldsa, liczba mocy (Newtona).
Wychodząc z założenia, że etapem ograniczającym przenoszenie mas między fazami jest
szybkość dyfuzji związku przez warstwę międzyfazową, możemy opisać to równaniem:
dC
S
/dt = kA(C
S
*
- C
S
)
C
S
– stężenie rozpuszczonego związku w medium otaczającym
C
S
*
- stężenie tego związku w warstwie międzyfazowej
A – całkowita powierzchnia międzyfazowa
k – współczynnik przenoszenia mas
Zwiększenie wartości współczynników k i A będzie prowadziło do zwiększenia szybkości
przenoszenia mas. Współczynniki te są zależne od:

k → temperatury, lepkości medium, wielkości rozpuszczonego związku,
intensywności mieszania, obecności związków interferujących
A → intensywności mieszania, działania sił ścinających, obecności czynników
powodujących zlepianie
Napowietrzanie
Fermentacje tlenowe są podstawowymi procesami biotechnologicznymi, prowadzącymi do
otrzymania ważnych komercyjnie produktów. Dostarczanie tlenu, niezbędnego w tych
procesach, jest dla technologów jednym z największych problemów do rozwiązania. Wynika
to z wspomnianego już faktu, że tlen jest słabo rozpuszczalny w wodzie. Tak więc dostępność
tlenu jest bardzo często czynnikiem limitującym szybkość procesu fermentacji tlenowej.
Często koszty związane z napowietrzaniem hodowli w skali przemysłowej są bardzo
poważnym składnikiem kosztów całkowitych hodowli.
Jeśli mamy do czynienia z prostym układem – zawiesiną pojedynczych komórek, to etapem
limitującym przenoszenie tlenu będzie przenikanie cząsteczek tlenu poprzez warstwę
międzyfazową.
W przypadku dobrego mieszania zawartości bioreaktora, etapem limitującym przenoszenie
tlenu będzie szybkość jego dyfuzji poprzez warstwę międzyfazową.
OTR = dC
O
/dt = k
L
a(C
O
*
- C
O
)
Ponieważ nie jest możliwe dokładne zmierzenie całkowitej powierzchni międzyfazowej, w
praktyce stosuje się łączny termin k
L
a, określający szybkość transferu tlenu na jednostkę
objętości.
Ponieważ nie ma również możliwości technicznej pomiaru stężenia tlenu w warstwie
międzyfazowej, dane te uzyskuje się metodą pośrednią: napowietrzając sterylną zawartość
fermentora, osiągniemy stan równowagi przy maksymalnym (w danej temperaturze) stężeniu
tlenu w roztworze. W tym przypadku stężenie tlenu zarówno w warstwie międzyfazowej, jak i
w roztworze będzie równe, co wynika z równania:
dC
O
/dt = k
L
a(C
O
*
- C
O
)
dC
O
/dt = 0 → C
O
*
= C
O
Maksymalna rozpuszczalność tlenu w cieczy może być wyznaczona w oparciu o równanie
Henry’ego:
C
O
*
= P
O
/H
O
Czynniki wpływające na współczynnik transferu tlenu (k
L
) przez warstwę międzyfazową:
wartość k
L
możemy zwiększyć poprzez zmniejszenie warstwy międzyfazowej
zwiększenie szybkości przenikania tlenu przez tę warstwę
Grubość warstwy międzyfazowej zależy od intensywności mieszania. Szybkość dyfuzji tlenu
przez tę warstwę zależy od lepkości środowiska i temperatury. Wzrost temperatury powoduje
spadek lepkości, ale równocześnie zmniejsza się rozpuszczalność tlenu w roztworze. Jeśli
mieszanie jest zbyt wolne lub przepływ powietrza zbyt duży, zbiera się ono pod turbiną,
pęcherzyki łączą się w duże bąble i spada wydajność natleniania.
Na stopień wykorzystania tlenu zawartego w powietrzu wprowadzanym do ścieków mają
wpływ:
wielkość
pęcherzyków
powietrza,
zależna
od
konstrukcji
elementów
napowietrzających
głębokość założenia elementów napowietrzających
kształt napowietrzanej komory
sposób ułożenia elementów napowietrzających w stosunku do rzuty poziomego
komory
Zatrzymanie gazu (G
H
) określa objętość gazu w stosunku do objętości cieczy w reaktorze.

Wartość G
H
podnosi się wraz ze wzrostem przepływu powietrza, a wartości G
H
i czasu
przebywania (τ) rosną wraz z:
wzrostem lepkości cieczy
spadkiem wielkości pęcherzyków
wysokością reaktora
Należy pamiętać, że dłuższy czas przebywania czy też wyższa wartość współczynnika G
H
nie
zawsze muszą zwiększać wymianę tlenu – ponieważ im dłużej pęcherzyk przebywa w
reaktorze, tym mniej tlenu może zawierać. Małe pęcherzyki szybciej oddają tlen; w praktyce
tak dobiera się warunki napowietrzania, aby pęcherzyki miały średnicę około 3mm.
Stała Henry’ego (H
O
) zależy od temperatury i zawartości soli w roztworze. Przy ciśnieniu
1 atm. ciśnienie parcjalne tlenu wynosi 0,21 atm. Dla czystego tlenu ciśnienie parcjalne w
normalnych warunkach ciśnienia atmosferycznego wynosiłoby 1 atm. A zatem, teoretycznie,
zwiększenie wymiany tlenu można osiągnąć przez natlenianie reaktora czystym tlenem,
zamiast powietrzem.
OUR – szybkość pobieranego tlenu
Stężenie rozpuszczonego tlenu w medium w bioreaktorze jest określane równowagą
pomiędzy szybkością przenoszenia tlenu (OTR) i jego pobieraniem (OUR).
dC
O
/dt = OTR – OUR
W ruchu laminarnym tory cząsteczek mało różnią się od siebie. Pozostające w ruchu medium
można traktować jako zbiór oddzielnych warstw, poruszających się względem siebie z różną
prędkością i nie mieszających się ze sobą. W ruchu turbulentnym ruch cząsteczek płynu
powoduje mieszanie się różnych warstw.

Lepkość cieczy
Lepkość (μ) najogólniej jest definiowana jako miara tarcia wewnętrznego cieczy (siły
międzycząsteczkowego przyciągania cieczy). Na charakterystykę lepkościową cieczy
składają się:
lepkość w zależności od temperatury
wskaźnik lepkości
W technice wyróżnia się:
lepkość dynamiczną – μ
lepkość kinematyczną – ν
Lepkość dynamiczną wyrażamy w jednostkach układu SI:
1 Pa · s = 1 N · s/m
2
= 1 kg/m · s
Lepkość dynamiczna jest często wyrażana w systemie CGS w puazach:
1 P = 1 dyna · s/cm
2
= g/cm · s =
1
/
10
Pa · s
Lepkość kinematyczna to stosunek lepkości dynamicznej danej cieczy do jej gęstości.
Zarówno lepkość kinematyczna, jak i lepkość dynamiczna muszą być wyznaczane w tej samej
temperaturze.
WYKŁAD 14, WYKŁAD 15 → również nie było mnie na nich, więc nie mam notatek…
Wyszukiwarka
Podobne podstrony:
filozofia skrypt w całości, Historia filozofii notatki z wyk?ad?w
Notatki z wyk+Ă©adu[1], Biotechnologia 3 rok 1 i 2 sem, Inżynieria bioprocesorowa
wyk ad 3 14.03.11 5 28.03.11 www.przeklej.pl, FiR, Notatki, Bankowość
Notatki z Kot y i si ownie ma ej mocy (wyk ad)
Prawo wyk ad prawo cywilne 2007, Studenci notatki i wiadomości, Prawo cywilne
Wyk ad 5 6(1)
Wyk ad II
Tkanki wyk ad 1
Ekonomika Transportu wyk+ad 1
Wyk ad Fizyka 2
Wyk ad 04
Na wyk ad id 312279 Nieznany
więcej podobnych podstron