MATERIAŁ NA WEJŚCIÓWKĘ Z GENETYKI
Kwasy nukleinowe (skład chemiczny i struktura kwasów nukleinowych)
Kwas nukleinowy składa się z połączonych podjednostek, którymi są nukleotydy.
Każdy nukleotyd zbudowany jest z zasady azotowej połączonej wiązaniem N- glikozydowym z cukrem (pentozą) oraz grupy fosforanowej
|
Zasada pirymidynowa |
Zasada purynowa |
Budowa |
Sześcioczłonowy pierścień pirymidynowy |
Sześcioczłonowy i pięcioczłonowy pierścień |
Nazwy |
C (cytozyna) T (tymina) w RNA T= U (uracyl) |
A (adenina) G (guanina) |
!!! Uracyl od Tyminy różni się grupą metylową w pozycji C5 heterocyklicznego pierścienia tyminy.
|
Ryboza |
Deoksyryboza |
Występowanie |
RNA |
DNA |
Obecność grupy hydroksylowej w pozycji 2' |
występuje |
brak |
Zasada purynowa lub pirymidynowa połączona jest z cukrem przez wiązaniem N- glikozydowe, które utworzone jest:
C1 pentozy a N1 pirymidyny
C1 pentozy N9 puryny
Zasadę azotową połączoną z cukrem nazywamy nukleozydem
Nukleozyd z przyczepioną grupą fosforanową nazywamy nukleotydem (ester fosforanowy nukleozydu)
Sekwencje zasad azotowych w kwasach nukleinowych zapisuje się w kierunku 5'- 3'
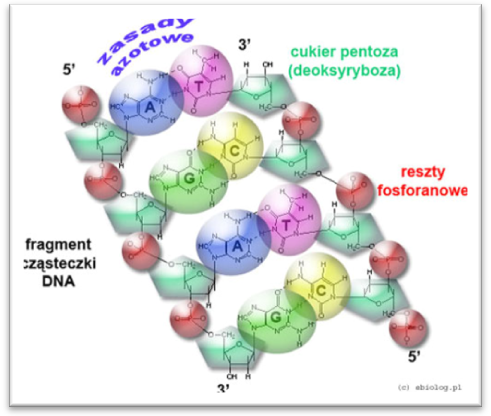
Cząsteczka DNA zbudowana jest z dwóch nici polinukleotydowych
Nici te łączą się w podwójnej helisie wiązaniami wodorowymi między zasadami azotowymi.
A= T, G= C- komplementarne pary zasad
Adenina z tyminą tworzy dwa wiązania
Guanina z cytozyną tworzy trzy wiązania
Jedna nić skręcona jest w kierunku 5'- 3' druga nić w kierunku 3' - 5'
Długość jednego całkowitego skrętu wynosi 3,4 nm a średnica 2 nm
Na jeden skok helisy przypada 10 par nukleotydów
Helisa jest prawoskrętna!
SEMIKONSERWATYWNA REPLIKACJA
Replikacja polega na enzymatycznej syntezie nowej nici DNA
Każda potomna nić DNA powstaje na matrycy nici macierzystej
Każda z potomnych cząsteczek DNA składa się z jednej nici macierzystej i jednej zsyntetyzowanej de Novo.
Miejsce rozdzielenia nici nazwano widełkami replikacyjnymi.
Nić wiodąca syntetyzowana jest bez przerw
Nić opóźniona syntetyzowana jest w postaci krótkich fragmentów DNA (fragmenty Okazaki)
STRUKTURA PRZESTRZENNA KWASÓW NUKLEINOWYCH
Struktura pierwszorzędowa- kolejność (sekwencja) nukleotydów w łańcuchu polinukleotydowym
Struktura drugorzędowa- dwa zwinięte helisowo łańcuchu polinukleotydowe
Struktura trzeciorzędowa- specyficzna konfiguracja w przestrzeni wielkocząsteczkowego, podwójnego heliksu DNA
Struktura czwartorzędowa- oddziaływanie wzajemne cząsteczek DNA, mających swoją określoną strukturę I-II- i III- rzędową.
STRUKTURY PODWÓJNEJ HELISY |
|||
|
B- DNA |
A-DNA |
Z-DNA |
Duży rowek |
Obecny- średnica 2,2 nm |
Obecny - trudniej dostępny niż w helisie B |
Brak |
Mały rowek |
Obecny- średnica 1,2 nm |
Obecny - ale uległ prawie całkowitemu zanikowi |
Obecny |
Liczba par zasad przypadających na jeden skok |
10,4 |
11 |
12 |
Kierunek skręcenia |
prawoskrętna |
prawoskrętna |
lewoskrętna |
Inne |
- |
Szersza i krótsza niż helisa B |
Szkielet cukrowo- fosforanowy przypomina kształtem literę Z |
Materiał genetyczny, kod genetyczny
MATERIAŁ GENETYCZNY
Występują 3 typy DNA:
Nici DNA ulegające natychmiast denaturacji
stanowi on ok. 10- 15% genomu ssaków
zgromadzony jest w okolicy centromerów chromosomów- odpowiada on prążkom C czyli heterochromatynie konstytutywnej i nie ulega ekspresji)
Nici DNA ulegające denaturacji wolniej
stanowi on od 20- 40 % genomu ludzkiego
charakterystyczne są dwie rodziny sekwencji:
SINE- krótkie rozproszone powtórzenia
LINE- długie rozproszone powtórzenia
Nici DNA ulegające denaturacji bardzo wolno
Ogólna budowa genów kodujących białka
region 5' flankujący
części niekodujące (introny)
region 3' flankujący
w pozycji -70 do -80 par zasad występuje sekwencja CAAT, do której przyłącza się jeden lub więcej białkowych czynników transkrypcyjnych
w pozycji -25 do -30 znajduje się sekwencja TATA- przyłącza się tu polimeraza RNA II
najczęściej w miejscu inicjacji transkrypcji znajduje się zasada purynowa.
transkrypt pierwotny powstaje z przepisania sekwencji kodujących- eksonów i niekodujących intronów.
intron- sekwencja przepisana a następnie wyeliminowana przez wycięcie w trakcie dojrzewania tran skryptu pierwotnego
KOD GENETYCZNY
Jest to sposób zapisania informacji dziedzicznej w DNA- dotyczy ona głównie kolejności nukleotydów w łańcuchu DNA.
Wyróżniamy 4 litery kodu genetycznego: A, G, T, C. Za ich pomocą możemy pisać słowa kodu genetycznego, którymi są triplety czyli trójki nukleotydów.
Jeden aminokwas jest kodowany przez trójkę zasad w DNA (triplet) co odpowiada komplementarnej trójce zasad w mRNA zwanej kodonem.
Kodony oznaczające ten sam aminokwas nazywane są synonimowymi.
Kodony zawierające informację o aminokwasie noszą nazwę kodonów sensownych.
Istnieją 3 kodony, pełniące funkcję sygnałów interpunkcji- kodony nonsensowne- kodony STOP ( UAA, UAG, UGA)
Cechy kodu genetycznego:
Trójkowy- jeden aminokwas jest kodowany przez triplet nukleotydów w DNA a zatem przed kodon w mRNA
Uniwersalny- dany kodon koduje ten sam aminokwas u wszystkich organizmów żywych oraz wirusów
Niezachodzący- nukleotydy nie zachodzą na siebie, każdy nukleotyd wchodzi w skład tylko jednego kodonu
Bezprzecinkowy- nie ma znaków przecinkowych pomiędzy kolejnymi trójkami i kod odczytywany jest w sposób ciągły
Wieloznaczny- dany aminokwas może być kodowany przez więcej niż jedną trójkę nukleotydów
Podstawowe pojęcia genetyczne (gen, allel, fenotyp, genotyp, cecha recesywna i dominująca, krzyżówki: Parentes, wsteczna, testowa)
Gen - podstawowa jednostka dziedziczności. Jest odcinkiem DNA zawierającym informację o budowie najczęściej jednego białka (o kolejności aminokwasów w pojedynczym łańcuchu polipeptydowym)
Allel - gen, który warunkuje różne formy tej samej cechy. Allele są wariantami jednego genu warunkującymi przeciwstawnośd danej cechy lub cech. Allele jednego genu oznacza się jako różne warianty jednej litery (A,a)
Allel dominujący - ujawnia swoją cechę w fenotypie
Allel recesywny - niewidoczny w fenotypie
Fenotyp - ogól widocznych cech morfologicznych, fizjologicznych, biochemicznych danego osobnika.
Genotyp - ogół genów danego osobnika
Cecha dominująca- dominującym określa się allel i odpowiadającą jemu cechę, który przejawia się u wszystkich heterozygotycznych osobników pierwszego pokolenia F1
Cecha recesywna- allele i odpowiadające im cechy maskowane przez geny dominujące nazywane są recesywnymi lub ustępującymi.
KRZYŻÓWKI
Krzyżówka Parentes
Krzyżówka wsteczna- polega na kojarzeniu heterozygot pokolenia F1 z którymkolwiek z typów rodzicielskich zarówno z cechą dominującą jak i recesywną.
Krzyżówka testowa - krzyżówka heterozygoty Aa z homozygotą recesywną aa. W wypadku dominacji krzyżówka ta pozwala stwierdzić heterozygotyczność analizowanego układu.
Prawa Mendla
I PRAWO MENDLA
(PRAWO CZYSTOŚCI GAMET)
Zgodnie z pierwszym prawem Mendla podczas podziału mejotycznego komórki następuje rozdział odpowiadającej sobie pary genów (alleli). Do każdej gamety przechodzi tylko jeden allel z danej pary.
Do każdej gamety dostaje się tylko jeden allel z dwóch warunkujących daną cechę!!
II PRAWO MENDLA
(PRAWO NIEZALEŻNEGO DZIEDZICZENIA SIĘ CECH)
Cecha uwarunkowana jedną parą genów (alleli) dziedziczy się niezależnie do cechy uwarunkowanej drugą parą genów (alleli), w związku z czym w pokoleniu F2 obserwujemy rozszczepienie fenotypów w stosunku 9:3:3:1.
Poszczególne cechy dziedziczą się niezależnie od siebie i mogą tworzyć różne kombinacje!!
Sprzężenie genów, zjawisko crossing - over
SPRZĘŻENIE GENÓW
Gdy geny warunkujące dwie rozpatrywane cechy leżą na tej samej parze chromosomów homologicznych, wówczas cechy te dziczą się zależnie (sprzężenie genów). Jeśli geny leżą blisko siebie na chromosomie to wykazują całkowite sprzężenie.
Sprzężenie genów - łączne przekazywanie genów i uwarunkowanych przez nie cech fenotypowych, spowodowane tym, że geny te są zlokalizowane blisko siebie w tym samym chromosomie. Sprzężenie zostaje zerwane wówczas, gdy nastąpi wymiana odcinków (crossing over) między homologicznymi chromosomami.(Morgan 1910).
CROSSING- OVER
Crossing over jest zjawiskiem zachodzącym w trakcie podziału mejotycznego przy wytwarzaniu komórek rozrodczych. Jest to wymiana odpowiadających sobie odcinków chromosomów homologicznych, mogąca powodować zerwanie sprzężeń pomiędzy genami, umożliwiając tym samym ich nowe kombinacje.
Proces ten zachodzi w profazie I mejozy (pachytenie) i polega na tworzeniu połączeń chiazm między chromatydami, a następnie rozrywaniu tych połączeń, ale tak, że następuje wymiana ich odcinków. W wyniku crossing-over dochodzi do rozszczepienia genów sprzężonych i powstania nowych sprzężeń. Częstość tego procesu zależy od odległości pomiędzy rozpatrywanymi genami w chromosomie: im bliżej są one położone, tym silniej są sprzężone i mniejsze jest prawdopodobieństwo rozdzielenia ich pomiędzy dwa różne chromosomy homologiczne
Wymiana między dwiema niesiostrzanymi chromatydami chromosomów homologicznych
Kodominacja, dominacja zupełna i niezupełna
Kodominacja polega na występowaniu alleli, które nie są związane stosunkiem dominacji- recesywności. Produkty tych alleli są wytwarzane jednocześnie i niezależnie; każdy z nich znajduje odbicie w fenotypie.
Zjawisko ko dominacji obserwuje się u ludzi przy dziedziczeniu grup krwi.
Kodominacja - stan, w którym oba allele danego locus ujawniają się w heterozygocie niezależnie od siebie (np. relacje między allelami grup krwi A i B)
Dominacja zupełna uwidacznia się już w heterozygotach, jeśli jeden z dwóch różnych alleli danego genu maskuje obecność drugiego. Jeśli np. gen A decyduje o wysokiej łodydze natomiast gen a decyduje o niskiej łodydze to heterozygota Aa wykształci rośliny o wysokich łodygach (uwidacznia się w heterozygotach, jeśli z dwóch rożnych alleli danego genu jeden maskuje obecność drugiego. )
Dominacja niezupełna- uwidacznia się w heterozygotach, jeśli allel R nie maskuje całkowicie obecności allela R'. Przykładem mogą być heterozygoty RR' o różowych kwiatach otrzymane ze skrzyżowania homozygoty RR o czerwonych kwiatach z homozygotą R'R' o kwiatach białych (uwidacznia się w heterozygotach, jeśli allel dominujący A nie maskuje całkowicie obecności allelu recesywnego)
Współdziałanie genów (epistaza, penetracja i ekspresja, poligeny, plejotropia, komplementacja);
EPISTAZA
Dotyczy dwóch lub więcej par alleli
W parze alleli gen dominujący całkowicie lub częściowo hamuje ujawnienie się genu recesywnego
Ekspresja genu dominującego jest tłumiona działaniem produktu innego genu znajdującego się odmiennym locus lub w innym chromosomie
Gen hamujący to gen epistatyczny
Gen maskowany to hipostatyczny
Wyróżniamy epistazę dominującą i epistazę recesywną.
PENETRACJA I EKSPRESJA
Penetracja (przenikliwość) to częstość ekspresji określonego genu wyrażona w procentach lub w wartościach liczbowych (pełna penetracja 100 % lub 1,0)
Ekspresja to ujawnienie się produktu danego genu, odpowiedzialnego za daną cechę. (niektóre geny wykazują zmienną ekspresję czyli zmienny stopień natężenia cechy)
POLIGENY
Inaczej geny kumulatywne- są to geny z różnych par alleli, zajmujące różne loci w chromosomach, wpływające na wytworzenie tej samej cechy.
U ludzi wzrost, IQ, barwa skóry jest determinowana przez poligeny
PLEJOTROPIA
Polega na warunkowanie przez jeden zmutowany gen kilku pozornie niezwiązanych z sobą cech fenotypowych.
KOMPLEMENTACJA
Polega na dopełniającym działaniu produktów różnych genów
Geny komplementarne działają wspólnie- produkt jednego genu ma wpływ na produkt drugiego genu
U człowieka działanie komplementarne wykazują geny decydujące o zabarwieniu włosów i skóry.
Cykl komórkowy (fazy cyklu)
Podział- mitoza (M)
Interfaza: G1, S, G2,
FAZA G1- faza wzrostu- między końcem mitozy a syntezą DNA
Komórka wchodząc w tą fazę jest dwukrotnie mniejsza od komórki macierzystej
Intensywne procesy anaboliczne, wzmożona wymiana chemiczna z otoczeniem, zwiększenie ruchliwości i wrażliwości na bodźce.
Zwiększenie masy i objętości komórki
Komórki w tej fazie syntetyzują cykliny A, C, D i E
W późnej fazie G1 występuje punkt restrykcyjny R, który zmusza komórkę do realizowania następnych etapów cyklu.
FAZA S- faza syntezy DNA
Czas trwania wyznaczony jest przez replikację DNA.
Masa i objętość komórki zwiększa się w porównaniu do wielkości komórki poprzedniej fazy
Rozpoczyna się syntetyzowanie cykliny B oraz synteza histonów.
FAZA G2- między końcem syntezy a rozpoczęciem mitozy
Druga przerwa w syntezie DNA
Zachodzi synteza białek wrzeciona podziałowego (tu buliny) oraz cykliny B
Nadprodukcja składników potrzebnych do odtwarzania błony komórkowej
W późnej fazie- aktywuje się kinaza białkowa CDK przyczyniająca się do późniejszego zaniku błony jądrowej i kondensacji chromosomów
FAZA G0- stan spoczynkowy komórki
W fazie G0 komórka traci umiejętność powielania materiału genetycznego i dzielenia się.
Komórki fazy G0 mają bardziej skondensowaną chromatynę, inne mRNA i swoiste białka.
Np. pod wpływem niedotlenienia komórki wątroby przechodzą z fazy G0 do fazy G1 dzieląc się i odbudowując miąższ wątroby- taki sam system działania mają komórki nowotworowe.
Mitoza
Mitoza- podział komórki somatycznej gdzie z jednej powstają dwie identyczne genetycznie komórki.
KARIOKINEZA
PROFAZA
Przejście od fazy G2 do M
Widoczne kondensujące chromosomy
Pod koniec profazy każdy chromosom to dwie identyczne (siostrzane) chromatydy połączone centromerem
Centriole rozchodzą się parami do biegunów komórki, pomiędzy nimi wytwarza się wrzeciono podziałowe
Przy centromerach wytwarzają się kinetochory od których odchodzą włókna
Zanika jąderko lub jądro
METAFAZA
Po zaniku otoczki jądrowej chromosomy ustawiają się w płaszczyźnie równikowej (PROMETAFAZA)
Chromosomy tworzą płytkę metafazową (stawiając się centromerami wrzeciona w płaszczyźnie równikowej)
W wyniku kondensacji powstają chromosomy metafazowe
Chromosomy pod koniec fazy układają się na obwodzie wrzeciona podziałowego w miejscy kineto choru- tworzą się pary homologiczne.
ANAFAZA
Rozpoczyna się całkowitym rozdzieleniem się chromatyd z każdego chromosomu
Przemieszczają się od ku biegunom komórki
TELOFAZA
Rozpoczyna się w momencie gdy chromatydy umiejscowione są na biegunach i de kondensują.
Chromatydy wydłużają się i ich struktura ulega rozproszeniu
Rozpoczyna się intensywna synteza rRNA
Włókna wrzeciona podziałowego zanikają
Przy udziale siateczki śródplazmatycznej odtwarzana jest otoczka jądrowe wokół chromatyd.
Przywrócona jest aktywność metaboliczna komórki
CYTOKINEZA
Podział cytoplazmy zaczyna się od przewężenia
Bruzda podziałowa pogłębia się,
Zacisk przewężenia zatrzymuje się na pęczku ściśniętych włóknie wrzeciona mitotycznego, które tworzą tzw. ciałko środkowe.
W procesie tym biorą udział również pęcherzyki i zbiorniki siateczki śródplazmatycznej.
Błona komórkowa powstaje z elementów wytworzonych w fazie G2
Mejoza, nondysjunkcja
Mejoza zachodzi tylko w komórkach szlaku płciowego. W podział mejotyczny może wejść tylko komórka diploidalna (2n). W wyniku tego procesu powstają gamety o haploidalnej liczbie chromosomów (1n)
I PODZIAŁ MEJOTYCZNY
PROFAZA I
LEPTOTEN
Chromosomy przybierają postać wydłużonych nitek
Każdy z chromosomów mejotycznych jest połączony z otoczką jądrową na obu końcach za pomocą płytki przyczepowej.
Chromosom składa się z dwóch siostrzanych chromatyd, ale nie można ich rozróżnić, bo przylegają do siebie bardzo ściśle.
ZYGOTEN
Rozpoczyna się proces zwany koniugacją- tworzenie par pomiędzy chromosomami homologicznymi
Powstaje kompleks synaptonemalny- składa się z długiego, środkowego rdzenia białkowego, który dopasowuje się do struktury chromosomu homologicznego (układ drabinkowy, połączenie na zasadzie zamka błyskawicznego)
Para chromosomów homologicznych nazywana jest biwalentem! (można też nazywać ją tetradą ze względu na 4 siostrzane chromatydy)
PACHYTEN
Powstają węzły rekombinacyjne na przebiegu kompleksu synaptonemalnego, których zadaniem jest ułatwienie procesu wymiany między chromosomami.
Zachodzi tutaj crossing- over- wymiana między dwiema niesiostrzanymi chromatydami chromosomów homologicznych
Miejsce w którym zachodzi crossing- over tworzy chiazmę!
DIPLOTEN
Chromosomy homologiczne rozdzielają się
Kompleks synaptonemalny rozpuszcza się
Biwalenty są połączone poprzez jedną lub dwie chiazmy
Chromosomy są aktywne w syntezie RNA
DIAKINEZA
Zmniejszona jest synteza RNA
Chromosomy ulegają kondensacji, grubieją i oddzielają się od otoczki jądrowej
Kinetochory chromosomów tworzących biwalent, zlewają się ze sobą
METAFAZA I
Chromosomy biwalente układają się w płaszczyźnie równikowej
KInetochory połączone są mikrotubulami wrzeciona.
ANAFAZA I
Rozpoczęcie rozdzielania biwalentów na dwie pary chromatyd, z których każda ma jeden centromer.
Z każdego chromosomu biwalentnego do biegunów komórki przesuwa się jeden chromosom
Chiazmy przesuwają się ku końcom ramion chromosomów (terminalizacja)
TELOFAZA I
Powstają dwie komórki z których każda zawiera haploidalną liczbę chromosomów z diploidalną ilością DNA
II PODZIAŁ MEJOTYCZNY
Jest to mitoza
W wyniku tego podziału powstają dwie komórki z których każda ma 23 pojedyncze chromosomy oraz ok. 3 pg DNA na jądro.
NONDYSJUNKCJA
Czynnik patogenetyczny prowadzący do aneuploidii, polega na nierozdzieleniu się chromosomów lub chromatyd podczas podziału komórki w trakcie mejozy lub mitozy.
Nondysjunkcja podczas I podziału mejotycznego jest wynikiem braku segregacji (nierozdzielenia się) chromosomów homologicznych
Nondysjunkcja podczas II podziału mejotycznego jest wynikiem braku segregacji chromatyd siostrzanych.
Czynnikiem wpływającym na nondysjunkcje jest wiek matki
Czynnikami są również: niewielkie dawki promieniowania, hormony pochodzenia egzogennego, fungicydy, herbicydy, alkohol
Przyczyną błędów są również leki:
anestezjologia- halotan
leki uspokajające- diazepam
przeciwnowotworowe- winblastyna, winkrystyna
przeciwgrzybiczne- gryzeofluwina
Wyszukiwarka
Podobne podstrony:
tematy ćwiczeń i zakres materiału na wejściówki 2011-12, semestr 1
materiały na wejściówke
opracowanie na wejściówkę z genetyki
inżynieria genetyczna - ściąga na egzamin, Zootechnika (UR Kraków) - materiały, MGR, Inżynieria gene
cw 4(1), II rok, II rok CM UMK, Giełdy, od Joe, histologia i embriologia, HISTOLOGIA, Materiały na ć
Inzynieria materialowa- pytania na wejsciowki do Zywicy, Studia, I o, rok II, semestr III, inżynieri
jebana histologia, II rok, II rok CM UMK, Giełdy, od Joe, histologia i embriologia, HISTOLOGIA, Mate
Przykładowe pytania na wejściówkę z laborek z ceramiki, AGH, Semestr 9, Współczesne materiały inżyni
materialy na diagnoze, Wyklad VI diagnoza
materialy na zajecia historia sejmu staropolskiego
biologia zakres materiau na egz Nieznany (2)
Proces brzeski, materiały na losy 1921-1945
więcej podobnych podstron