Karolina Jankowska, Tadeusz Trzmiel
Instytut Biochemii Technicznej
Podstawy teoretyczne ekologii biochemicznej
I. WSTĘP
1. Wprowadzenie
Od początków istnienia życia na Ziemi organizmy walczyły pomiędzy sobą o dostęp do światła, wody oraz składników odżywczych. Wygranymi w tej nieustającej walce są zawsze organizmy, które potrafią najlepiej zaadoptować się do warunków panujących w danym środowisku, oraz te które wykształciły najdoskonalsze mechanizmy przetrwania.
W świecie mikroorganizmów doskonałym przykładem takiej adaptacji są wirusy, które poprzez częste mutacje potrafią dopasować się do zmiennych warunków w środowisku ich bytowania np. głośny w ostatnim czasie wirus ptasiej grypy, który poprzez mutację utorował sobie drogę do atakowania ludzi. Innym przykładem mogą być grzyby strzępkowe bądź promieniowce Streptomyces, które posiadają zdolność do produkcji antybiotyków, działających toksycznie na inne mikroorganizmy żyjące w ich środowisku.
Walka o przetrwanie i przedłużenie linii gatunkowej w świecie zwierząt pozwoliła w procesach ewolucyjnych wykształcać coraz to nowsze i doskonalsze mechanizmy obronne u potencjalnych ofiar jak i mechanizmy zabijania u potencjalnych napastników.
Zwierzęta należące do tego samego gatunku wykształciły zdolności do swobodnego komunikowania się w grupie, dawania sygnałów ostrzegających o niebezpieczeństwie lub o gotowości do przyjęcia potomstwa, a także do informowania innych o obecnym nastroju danego osobnika. Cechy te widoczne są nie tylko wśród zwierząt ale przede wszystkim to my ludzie potrafimy w najróżniejszy sposób przekazywać sobie różnego typu informacje i w świecie oddziaływań jesteśmy absolutnymi pionierami.
Również rośliny nie pozostały w tyle w tym wyścigu zbrojeń, prześcigają się w kształtach, kolorach i zapachach kwiatów, które wabią zapylaczy niezbędnych do przedłużenia linii gatunkowej. Rośliny drapieżne wabią swoje ofiary słodkim nektarem lub odpowiednim zapachem. Niektóre rośliny wytwarzają substancje toksyczne aby ochronić się przed roślinożercami, lub wyeliminować inne gatunki roślin zamieszkujących to samo środowisko.
Człowiek od wieków badał przyrodę i starał się wyjaśnić zjawiska w niej zachodzące. Obserwacje zachowania zwierząt i roślin oraz zagadki związane z ogromną różnorodnością związków organicznych występujących w naturze od dawna rodziły pytania, które zadawali sobie naukowcy. Dopiero połączenie wiedzy ekologów i biochemików zaowocowało rzetelną odpowiedzią na wiele problemów.
2. Czym jest ekologia biochemiczna?
Ekologia biochemiczna jest nauką powstałą z połączenia się dwóch dziedzin naukowych: biochemii oraz ekologii. Na pierwszy rzut oka te dwie dziedziny mają ze sobą niewiele wspólnego, ponieważ ekologia opiera się głównie na obserwacjach i badaniach plenerowych natomiast biochemia jest nauką ściśle laboratoryjną.
Połączenie to jednak pozwoliło odpowiedzieć na pytania zarówno w dziedzinie ekologii jak i biochemii, które jak dotąd pozostawały bez odpowiedzi.
Powstanie ekologii biochemicznej dało początek nowemu zagadnieniu, którego przedmiotem jest badanie, jaki wpływ na stosunki ekologiczne w ekosystemach i niszach ekologicznych mają związki chemiczne, będące w głównej mierze produktami metabolizmu wtórnego, a których funkcja jak dotąd nie została zdefiniowana.
Ekologia biochemiczna odsłania kompleksowość oddziaływań i przystosowań koewolucyjnych występujących pomiędzy organizmami żyjącymi we wspólnym środowisku. Nauka ta wyjaśnia również w sposób racjonalny i satysfakcjonujący przyczyny istnienia olbrzymiej różnorodności produktów wtórnego metabolizmu u mikroorganizmów, roślin oraz zwierząt.
3. Pojęcia podstawowe dotyczące ekologii biochemicznej:
Biochemia - nauka zajmująca się budową i funkcjami różnych substancji organicznych o dużym znaczeniu biologicznym, będących substratami, regulatorami oraz produktami w procesach metabolicznych organizmów. Wśród tych substancji znajdują się białka enzymatyczne, białka, węglowodany, lipidy, kwasy nukleinowe, aminokwasy i inne związki występujące naturalnie w środowisku.
Ekologia - nauka zajmująca się głównie obserwacją i badaniem oddziaływań pomiędzy żywymi organizmami w ich środowisku naturalnym, badająca zachowania organizmów różnych gatunków oraz zależności występujące w grupach organizmów tego samego gatunku.
Biosfera - przestrzeń (środowisko) bytowania organizmów.
Ekosystem - układ ekologiczny obejmujący biocenozę będącą zespołem populacji różnych gatunków organizmów żywych oraz biotop czyli środowisko nieożywione będące miejscem bytowania biocenozy. W ekosystemie zachodzi przepływ energii oraz obieg materii.
Ochrona środowiska - przedsięwzięcia podejmowane w celu zapobiegania, ograniczania i usuwania następstw zanieczyszczenia środowiska wynikających z działalności człowieka .
Ksenobiotyki - substancje obce i szkodliwe dla środowiska naturalnego, syntetyzowane sztucznie przez człowieka i nie występujące naturalnie w przyrodzie.
Toksyny - substancje chemiczne wytwarzane przez organizmy żywe (np. niektóre bakterie i pleśnie), mające właściwości trujące dla innych organizmów (np. antybiotyki, mykotoksyny).
Sukcesja ekologiczna - naturalne następstwo (po sobie) wybranych gatunków roślin i zwierząt, a także drobnoustrojów. Przykładem mogą być niektóre grupy mikroorganizmów występują kolejno po sobie prowadząc sukcesywną przemianę materii. Proces kolejnych dominacji drobnoustrojów z różnych grup powtarza się, aż do całkowitego rozkładu substratu. Sukcesja ekologiczna ma miejsce na przykład przy samooczyszczaniu wód.
Metabolity wtórne - substancje o różnorodnej budowie chemicznej wytwarzane podczas metabolizmu komórkowego i wydzielane przez żywe organizmy do środowiska w niewielkich ilościach, nie będące dla nich źródłem energii ani składnikiem strukturalnym, a odgrywające głównie rolę nośników informacji lub regulatorów procesów ekologicznych, w tym w oddziaływaniach pomiędzy żywymi organizmami.
II. ODDZIAŁYWANIA WEWNĄTRZ EKOSYSTEMU
1. Obieg pierwiastków w przyrodzie
Podstawą funkcjonowania ekosystemu jest przepływ i obieg materii. Pjęcie to jest rozumiane poprzez krążenie pierwiastków chemicznych, przemieszczających się ze środowiska abiotycznego do organizmów żywych i z powrotem. Są to tak zwane cykle biogeochemiczne. Najważniejsze z nich to cykl węgla i azotu, a także fosforu i siarki.
Węgiel wchodzi w skład wszystkich związków organicznych, dlatego jego krążenie w przyrodzie jest prawie jednoznaczne z krążeniem materii w ogóle. Nie można tu jednak zapomnieć, że węgiel istnieje również w postaci związków nieorganicznych, które także mają niebagatelne znaczenie w przyrodzie.
Obieg węgla:
Węgiel (C) to podstawowy pierwiastek budulcowy związków organicznych. W żywą materię organiczną zostaje wbudowany w postaci CO2 asymilowanego przez autotrofy (rośliny zielone, niektóre bakterie). Dzięki istniejącym łańcuchom pokarmowym węgiel przedostaje się w postaci roślinnych związków organicznych do konsumentów I rzędu (roślinożerców), a następnie do konsumentów II rzędu (zwierzęta mięsożerne). Ze związków organicznych węgiel wraca do obiegu jako CO2, powstający w procesie oddychania heterotrofów i autotrofów.
Globalny cykl węgla w środowisku jest odzwierciedleniem produkcji pierwotnej i wtórnej. Jednym ze sposobów dostawania się węgla do atmosfery, mającym większe znaczenie dopiero od końca XVIII wieku, jest spalanie paliw kopalnych, czyli przywracanie do obiegu węgla, który został z niego wyłączony przed wielu milionami lat. Wpływ na ilość węgla w atmosferze ma również uwalnianie go ze skał węglanowych. Skały takie były od wielu stuleci wypalane w piecach, zwanych wapiennikami, aby uzyskać wapno palone, przede wszystkim do wyrobu zaprawy murarskiej. Obecnie w podobnych celach, ale na nieporównywalnie większą skalę, prowadzą to działanie cementownie. Człowiek ma ogromny wpływ na obieg węgla w środowisku jednak największy przepływ tego pierwiastka odbywa się pomiędzy atmosferą a roślinnością lądową oraz między atmosferą a oceanami. Te dwa strumienie węgla są niemal równe, a średni czas przebywania atomu węgla w atmosferze wynosi około 3 lat. Wahania zawartości CO2 atmosferycznego są związane z sezonowością jego poboru przez rośliny oraz sezonowością użytkowania paliw i wymiany CO2 z oceanami. Biogeochemiczny obieg węgla opisuje się na podstawie cyklu dwutlenku węgla.
Obieg dwutlenku węgla jest regulowany głównie przez procesy jego wiązania (fotosynteza, chemosynteza, rozpuszczanie się w wodzie) i uwalniania (oddychanie, uwalnianie z gleby i procesów geologicznych zachodzących w skorupie ziemskiej, spalanie paliw). Obieg ten jest utrzymywany głównie przez ustalenie się równowagi pomiędzy dwoma podstawowymi procesami zachodzącymi na Ziemi - fotosyntezą i chemosyntezą a oddychaniem. Zasadnicza część węgla wiązana jest przez rośliny zielone. Zasoby dostępnego dwutlenku węgla zostałyby szybko wyczerpane, gdyby nie uwalnianie tego gazu w procesach oddychania. Rośliny wydalają do środowiska dwutlenek węgla w ilości 1% swojej masy na dobę, ssaki - 3%, ptaki - 25%, a mikroorganizmy aż 500%. Część tego gazu pochodzi z procesów rozkładu materii organicznej i procesów wulkanicznych oraz spalania paliw organicznych. Stan biomasy całej kuli ziemskiej pozostaje względnie stały, jego zmiany spowodowane są działaniami z zewnątrz. W minionych epokach geologicznych na stan równowagi duży wpływ miały zmiany klimatu. W okresie karbońskim klimat podobny był do dzisiejszego równikowego. Dlatego ówczesna roślinność była wyjątkowo bogata, dzięki czemu powstały ogromne złoża węgla kamiennego. Łączna biomasa roślin, w przeciwieństwie do permu, gdzie klimat był na ogół suchy, była bardzo wysoka. Obecnie na zawartość węgla w roślinach ma wpływ niszczenie przez człowieka np. wielkich połaci lasów tropikalnych, jak również coraz szybszy rozwój miast powodujący zagładę terenów zielonych.
Schemat obiegu węgla w przyrodzie:
Obieg azotu:
Drugim ważnym pierwiastkiem biogennym jest azot, należy on do makroelementów i jest podstawowym pierwiastkiem wchodzącym w skład białek i kwasów nukleinowych.
W dużej ilości azoty występuje w atmosferze, stanowi 78% powietrza, niemniej jednak jest on w tej postaci bardzo trudno dostępny dla rośli. Wyjątek stanowią rośliny motylkowe, żyjące w mutualizmie z bakteriami brodawkowymi z rodzaju Rhizobium sp. Oprócz tego wolny azot cząsteczkowy wiążą np. bakterie tlenowe z rodzaju Azotobacter sp. i beztlenowe z rodzaju Clostridium sp. W środowisku wodnym azot wiązany jest przez niektóre gatunki sinic (Nostoc). Rośliny uzyskują azot z gleby i wody, głównie w postaci jonów amonowych lub azotanowych i wbudowują go w białka i kwasy nukleinowe. Białko roślinne jest wykorzystywane przez zwierzęta i innych konsumentów. Po przemianach metabolicznych jest wydalany w postaci amoniaku, mocznika i kwasu moczowego.
Szczątki roślin, zwierząt, grzybów pierwotniaków ulegają bakteryjnemu rozkładowi, w którym azot może zostać uwolniony w postaci amoniaku albo w postaci jonu amonowego. Następnie w warunkach tlenowych bakterie nitryfikacyjne z rodzaju Nitrosomonas sp. utleniają jony amonowe do azotynów. Azotyny są wykorzystywane przez bakterie z rodzaju Nitrobacter sp. i utleniane do azotanów. Część azotanów środowisko traci na drodze szczególnego rodzaju oddychania beztlenowego bakterii denitryfikacyjnych np. Paracoccus denitrificans. Redukują one azotany do azotu cząsteczkowego, który uchodzi do atmosfery. Dzięki burzom możliwe jest przechodzenie wolnego azotu z atmosfery do produktywnego obiegu, ponieważ energia wyzwalająca się podczas wyładowań atmosferycznych jest na tyle duża że powoduje rozszczepienie cząsteczki azotu i umożliwia jego utlenianie się do azotanów.
Schemat obiegu azotu w przyrodzie:
Schemat obrotu azotu w glebie:
Obieg fosforu:
Fosfor, mimo że stanowi zaledwie 0,2% (wagowy udział) masy lądów i oceanów to ma ogromny wpływ na życie prawie wszystkich organizmów żywych. Dlatego też nazwany został pierwiastkiem życia. Fosfor nie wchodzi w skład białek, lecz żadna cząsteczka białka nie może być wytworzona bez udziału fosforu. ,,Wysokoenergetyczne" odwracalne wiązania są uniwersalnym paliwem dla wszystkich reakcji biochemicznych, które zachodzą wewnątrz komórki. Chociaż ani ADP, ani ATP nie zawierają wiele fosforu, jeden atom tego pierwiastka na jedna cząsteczkę adenozyny jest bezwzględnie potrzebny. Żadne procesy życiowe nie byłyby bez niego możliwe. Fosfor jest ważnym i koniecznym składnikiem protoplazmy.
Znaczna większość fosforu występującego w przyrodzie zawarta jest w związkach budujących organizmy i znajduje się w ich płynach ustrojowych oraz w strukturze szkieletowej. Krążenie fosforu w przyrodzie odbywa się w dwóch cyklach: lądowym i morskim. Głównym rezerwuarem fosforu na powierzchni lądu są skały powstałe w dawnych epokach geologicznych. Skały te stopniowo wietrzeją, a uwolnione związki fosforowe dostają się do ekosystemów lądowych oraz morskich. Fosfor znajdujący się w glebie udostępniają roślinom i zwierzętom, bakterie fosforowe przetwarzające martwą materię organiczną (roślinną i zwierzęcą) oraz produkty metabolizmu organizmów znajdujące się w glebie. Bakterie przyczyniają się do rozkładu organicznych form fosforu na fosforany łatwo przyswajalne przez rośliny. Największa ilość tego pierwiastka jest tracona na skutek zmywania go do oceanów i mórz, gdzie osadza się w formie nieprzyswajalnej dla organizmów. Wówczas powrót ten może nastąpić tylko w wyniku ruchów górotwórczych wynoszących dno oceanów ponad powierzchnię wody. Możliwości ponownego włączenia się w obieg są tu niewspółmierne w stosunku do ilości ubywającej. Lokalnie ubytek fosforu następuje w wyniku upraw, a zwłaszcza niektórych gatunków zbóż, które zużywają znaczne ilości fosforu, co powoduje powolne ubożenie gleby. Z tego powodu w celu uzupełnienia niedoborów fosforu, gleby użytkowane rolniczo zasila się nawozami naturalnymi lub sztucznymi. Zbyt duże koncentracje ludności, na osiedlach i w miastach doprowadzają do tego, iż w ściekach emitowanych do rzek znajdują się znaczne nadwyżki fosforu. Najważniejszym źródłem dostępnego fosforu w morzach są organizmy planktonowe, które stanowią pierwszy element cyklu obiegu fosforu w ocenach. Następnie fosfor przechodzi do zooplanktonu jako efekt produkcji wtórnej. Kolejno fosfor jest pochłaniany z pokarmem przez ryby planktonożerne, drapieżne i inne organizmy w kolejnych ogniwach łańcucha pokarmowego. Około 1% fosforu powraca na ląd dzięki połowom ryb stając się źródłem nawozu, który dostarcza fosforu glebie. Tylko przy brzegach Peru ptactwo wodne wybiera z morza około 5,5 milionów ton ryb rocznie przetwarzając je na wielkie ilości guano. Znaczna część fosforu wypada z obiegu morskiego na skutek sedymentacji produktów rozpadu materii organicznej, tam jest on pozyskiwany jedynie przez bentos. Na dno opada też fosfor uwolniony do fosforanów, ponieważ są one słabo rozpuszczalne w wodzie i wytrącają się w postaci osadu. Związany fosfor przepada w osadach dennych i tu praktycznie znika z obiegu.
Ilość wolnego fosforu w przyrodzie jest niewielka, ponieważ wypłukiwanie go ze skał jest powolne, a szybkość asymilacji przez organizmy żywe duża. Ponieważ szybkość uwalniania fosforu jest mniejsza niż jego pobieranie, na wiosnę cały dostępny fosfor zostaje związany przez ciała producentów i konsumentów. Niedostatek tego pierwiastka staje się czynnikiem ograniczającym. W układzie następuje wtedy zwolnienie procesów wiązania, do czasu gdy szczątki organizmów i odchody nie zostaną rozłożone, a przyswajalne formy fosforu nie wejdą ponownie do obiegu.
Cykl biogeocheniczny fosforu niestety jest bardzo podatny na zmiany powodowane przez człowieka. Największe zakłócenia są powodowane przez rolnictwo, ponieważ w zasadzie wszystkie stosowane nawozy mineralne zawierają ten pierwiastek. Fosfor jest wypłukiwany w dużej części z gleb i spływa do jezior i innych zbiorników wodnych, co w konsekwencji powoduje ich eutrofizację.
2. Podział oddziaływań w ekosystemie
Pomiędzy organizmami żyjącymi we wspólnym środowisku na drodze ewolucji wytworzyło się mnóstwo zależności nazwanych w tym rozdziale oddziaływaniami wewnątrz ekosystemu. Organizmy stanowiące biocenozę wpływają na siebie wzajemnie oraz na biotop na ściśle określonych zasadach, w ten sposób formuje się tak zwana równowaga ekologiczna, każda obca ingerencja w ekosystem może zakłócać tą równowagę i doprowadzić do trwałych szkód w środowisku.
Wzajemne zależności w ekosystemie możemy podzielić na trzy grupy:
Pierwsza grupa zależności dotyczy konkurencji pomiędzy organizmami w ekosystemie o terytorium bytowania, dostęp do światła, wody i składników odżywczych.
Druga grupa to oddziaływania mające na celu wzajemną komunikację i przekazywanie sobie różnego typu informacji pomiędzy organizmami.
Trzecia grupa dotyczy oddziaływań w sferze psychologicznej, zależności takie można zaobserwować wśród zwierząt ale przede wszystkim to człowiek wykształcił w sobie możliwość wpływania na innych poprzez praktyki psychologiczne.
Zarówno w pierwszej jak i drugiej grupie ogromną rolę odgrywają substancje chemiczne, a więc możemy podzielić oddziaływania pomiędzy organizmami na dwie podstawowe grupy odnośnie substancji chemicznych biorących udział w tych oddziaływaniach.
Z udziałem substancji i cząsteczek, stanowiących dla otrzymujących je organizmów źródło energii lub składników strukturalnych.
Z udziałem cząsteczek, które odgrywają wyłącznie lub przede wszystkim rolę nośników informacji lub regulatorów procesów ekologicznych przepływu energii i substancji przez ekosystem.
Cząsteczkowe rozpatrywanie relacji w środowisku to główne zadanie ekologii biochemicznej, a więc w kolejnych punktach pracy będę omawiała oddziaływania pomiędzy organizmami pod kątem związków biorących w nich udział.
3. Zależności pokarmowe ekosystemu
Jak już wspominałam życie w ekosystemie to nieustający wyścig zbrojeń i doskonalenie sztuki przetrwania. Dla świata roślin konkurencja dotyczy głównie dostępu do światła, wody i składników mineralnych zawartych w glebie a więc również o miejsce, ale także doskonalenie metod przyciągania zapylaczy. Dla zwierząt to walka o pokarm, wodę, terytorium bytowania i ochrona potomstwa.
Jednak nie zawsze relacje w biocenozie mają charakter walki pomiędzy organizmami. Wyróżniamy trzy typy oddziaływań pokarmowych: dodatnie (nieantagonistyczne: komensalizm, protokooperacja, mutualizm), ujemne (antagonistyczne: amensalizm, konkurencja, drapieżnictwo i pasożytnictwo) i obojętne (neutralizm). Ten ostatni typ interakcji polega na tym, że nie występuje wzajemne oddziaływanie pomiędzy gatunkami, żadna populacja nie podlega wpływom drugiej.
Oddziaływania nieantagonistyczne
Komensalizm: W takim układzie jeden organizm (komensal) czerpie korzyści z życia w zespole z innym (gospodarzem) nie przynosząc drugiemu ani szkody ani korzyści.
Wśród mikroorganizmów komensalizm polega na tworzeniu przez jednego ze współmieszkańców ekosystemu substancji wzrostowych, np. witamin, stymulujących wzrost partnerów, rozkładzie lub wykorzystywaniu w środowisku substancji hamujących wzrost partnerów a także na przeprowadzeniu przez jednego z drobnoustrojów substratów pokarmowych, nieprzyswajalnych przez partnera, w produkty, które już może wykorzystać jako składniki odżywcze. W środowiskach naturalnych, szczególnie w wodzie i glebie, ten rodzaj zależności jest dość powszechny i najczęściej polega na rozkładzie sacharydów lub białek do produktów łatwo przyswajalnych przez partnerów. Mikroorganizmy glebowe, niezdolne do wykorzystania takich sacharydów roślinnych, jak celuloza czy hemicelulozy są zależne od grzybów wydzielających do środowiska enzymy hydrolityczne, rozkładające te substraty; podobnie niektóre mikroorganizmy zdolne tylko do wykorzystywania aminokwasów jako źródła azotu mogą rozwijać się w środowiskach białkowych jedynie z mikroorganizmami o uzdolnieniach proteolitycznych. Zależności komensalne obserwuje się również w mleku pomiędzy drożdżami nie wykorzystującymi laktozy jako źródła węgla i bakteriami mlekowymi.
Komensalizm może polegać na zależnościach pokarmowych ale również na udzielaniu schronienia komensalowi, udostępnienia dogodnego miejsca bytowania, transportu.
Przykładem komensalizmu pokarmowego może być układ pomiędzy pewnym gatunkiem rybika (Lepisma) a gatunkiem środkowoamerykańskich mrówek wędrownych. Owad ten żyje wśród mrówek, korzystając z pożywienia zdobywanego przez nie. Mrówki nie odnoszą z obecności rybika żadnych widocznych korzyści, ani nie ponoszą żadnych strat.
Przykłady komensalizmu:
Zdjęcie 1. Komensal z lasów tropikalnych rybik (Lepisma)
Do komensalizmu można niewątpliwie zaliczyć związek roślin epifitycznych z drzewami, na których one rosną. Epifity nie szkodzą drzewom, korzystają jedynie ze ściekającej wody i substancji mineralnych zmywanych z powierzchni kory i liści, niewykorzystywanych przez drzewo. Poza tym epifity usytuowane na pniu drzew, znajdują się wyżej niż inne rośliny i mają zancznie lepszy dostęp do światła słonecznego. Epifity wykształciły szereg specyficznych przystosowań do nadziemnego trybu życia, jak np. korzenie powietrzne chłonące wilgoć atmosferyczną i bulwy magazynujące wodę u storczyków, układ liści w formie lejka magazynującego wodę deszczową i substancje mineralne u ananasowatych i paproci, czy wreszcie przekształcenia liści tworzących rodzaj własnej "doniczki" u diszidii czy platycerium. W Polsce występuje wiele epifitów w postaci mchów i porostów.
Przykłady epifitów:
Zdjęcie 2. Oplątwa brodaczkowata (Tillandisa usneoides)
Epifity z rodziny ananasowatych:
Zdjęcie 3 i 4. Guzmania monostachya (z lewej), Aechmea (z prawej)
Przykłady innych komensali epifitycznych:
Zdjęcie 5, 6. Katleja z rodziny storczykowatych (u góry) i Diszidia Rafflesa
Przykładem komensalizmu chroniącego może być związek ryb z rodzaju Amphipron z ukwiałami. Ryby te znajdują schronienie pomiędzy ramionami ukwiałów. Przykładem komensalizmu transportowego, w którym gospodarz jest wykorzystywany jako środek transportu, może być ryba remora o zmienionej morfologii płetwy grzbietowej, remora przyczepia się do rekina i razem z nim się przemieszcza.
Inne komensale:
Zdjęcie 7, 8. Ryby Amphipron ocellaris (z lewej)) i Amphipron clarkii żyjące w ramionach ukwiałów.
Zdjęcie 9. Ryba Remora remora przyczepiona do rekina karaibskiego wykorzystuje go jako środek transportu.
Protokooperacja: Układ ten polega na współżyciu przynoszącym obopólne korzyści dla dwóch gatunków nie w pełni od siebie uzależnionych.
Każdy organizm w protokooperacji jest zdolny do życia bez partnera. Protokooperacja może występować okresowo. U mikroorganizmów protokooperacja polega na wzajemnym uprzystępnianiu składników pokarmowych, przykładem może być zespół złożony z bakterii celulolitycznych i bakterii asymilujących azot atmosferyczny, np. z rodzaju Azotobaeter; bakterie Azotobaeter dostarczają partnerom zredukowanych (przyswajalnych) związków azotu, natomiast bakterie celulolityczne, degradując celulozę, zaopatrują zespół łatwo przyswajalne źródło węgla (glukozę); podobne zależności występują w środowiskach, w których współbytują mikroorganizmy zdolne do rozkładania wysokocząsteczkowych substratów (białek, sacharydów, tłuszczy).
Innym typem takiej zalezności może być wzajemna wymiana gazów, najczęściej dotyczy to CO2 i O2, taka zależność łączy heterotroficzne bakterie tlenowe i glony w ściekach, podczas mineralizacji związków organicznych bakterie wydzielają duże ilości CO2, który jest wykorzystywany przez fotoautotroficzne glony i sinice jako źródło węgla, glony natomiast w wyniku metabolizmu fotosyntetycznego zaopatrują partnerów we tlen, ten rodzaj współzależności występuje powszechnie w środowisku wodnym i podczas oczyszczania ścieków.
W podobnym systemie zależne są od siebie bakterie beztlenowe i tlenowe w glebie. W powierzchniowych warstwach gleby, pomimo panowania warunków tlenowych, mogą rozwijać się beztlenowe bakterie, np. z rodzaju CIostridium, możliwe jest to dzięki współżyjącym z nimi drobnoustrojom tlenowym, które wykorzystują tlen i stwarzają warunki beztlenowe, organizmy tlenowe natomiast wykorzystują niektóre produkty beztlenowego metabolizmu partnerów. Innym rodzaj protokooperacji wśród mikroorganizmów polega na wytwarzaniu i wzajemnej wymianie substancji wzrostowych przez partnerów zespołu np. u bakterii juogurtowych. Przykładem zależności protokooperacyjnej u zwierząt może być współpraca pomiędzy ukwiałem i rakiem pustelnikiem. Rak pustelnik jest chroniony parzydełkami ukwiału, natomiast sam pomaga ukwiałowi w zdobyciu pożywienia i służy mu jako środek transportu. Innym przykładem protokooperacji może być układ pomiędzy ptakami bąkojadami i dużymi ssakami roślinożernymi np. z nosorożce, słonie, bawoły, żyrafy, zebry i inne.. W porach skóry tych ssaków żyją larwy pasożytniczych owadów będące pożywieniem dla bąkojadów. Ptaki oczyszczają skórę roślinożerców z pasożytów, a przy okazji zdobywają pokarm.
Niektóre rośliny tj. glistnik, wilczomlecz, kokorycz, fiołek i inne wytwarzają elajosomy na nasionach lub owocach, są to niewielkie ciałka bogate w tłuszcze i białko, stanowią one odżywczy pokarm dla mrówek, które przenosząc elajosomy biorą udział w rozsiewaniu rośliny zjawisko to nazywane jest myrmekochorią.
Przykłady protokooperacji:
Zdjęcie 10. Rak pustelnik (Pagurus bernhardus) i ukwiał.
Zdjęcie 11 i 12. Bąkojad czrwonodzioby (Buphagus erythrorhynchus) oczyszcza skórę roślinożerców.
Mutualizm: Jest to ścisły związek pomiędzy organizmami czerpiącymi z niego obopólne korzyści. W mutualizmie partnerzy nie są zdolni do życia osobno, niekiedy stanowią jeden organizm. Przykładem takiej zależności są porosty zbudowane z grzyba i glonu. Strzępki grzybni stanowią szkielet organizmu, a glony są rozmieszczone na nich warstwowo (heteromerycznie) lub bezładnie (homomerycznie). Glon przez fotosyntezę dostarcza składników pokarmowych.
Ciekawym przykładem mutualizmu jest również współzależność pomiędzy bakteriami i glonami, np. pałeczki z rodzaju Pseudomonas mogą rozwijać się w cytoplazmie Volvox. Pałeczki te mogą również występować w komórkach pierwotniaków. Uważa się, że odgrywają rolę w trawieniu pewnych składników pokarmowych nieprzyswajalnych przez pierwotniaki. U innych pierwotniaków, które zdolne są jedynie do prowadzenia glikolizy z wytworzeniem kwasu mlekowego, stwierdzono, że mogą spełniać funkcję podobną do mitochondriów prowadząc oddychanie tlenowe. Również pantofelki bardzo często łączy symbiotyczna zależność z bakteriami. Najczęściej są to pałeczki lub formy spiralne urzęsione i przenoszone od osobnika do osobnika podczas koniugacji. Niektóre z nich dostarczają pierwotniakom witamin, których nie potrafią same syntetyzować.
Mutualistyczne zależności mogą łączyć również partnera mikrobiologicznego z organizmami wyższymi. Szczególne znaczenie ekonomiczne ma symbioza pomiędzy roślinami motylkowymi i bakteriami z rodzajów Rhizobium i Bradyrhizobium. Podobne stosunki łączą drzewa i krzewy, np. olchę z promieniowcami. Rośliny te nie wytwarzają jednak brodawek korzeniowych.
Specyficznym rodzajem mutualizmu jest mikoryza, polega ona na współpracy pomiędzy grzybami kapeluszowymi a korzeniami rośliny naczyniowej. Grzyby dostarczają roślinie wodę z solami mineralnymi, poprzez specyficzne zdolności metaboliczne udostępniają związki azotowe, fosforowe i substancje wzrostowe(aktywatory wzrostu i kiełkowania ). Przykładami takiego układu mogą być maślak - sosna, koźlarz - brzoza, borowik - dąb.
Kolejnym przykładem mutualizmu jest związek pomiędzy drobnoustrojami stanowiącymi mikroflorę przewodu pokarmowego organizmu wyższego np. przeżuwaczy takich jak krowa, jeleń, wielbłąd. Zwierzęta te nie są zdolne do wytwarzania niektórych enzymów (w szczególności celulolitycznych). Mikroflora mająca tę zdolność odgrywa bardzo ważną rolę w procesach trawiennych, a ponad to dostarcza zwierzęciu wiele cennych witamin np. B i K. Bakterie korzystają w optymalnych dla ich rozwoju warunków: temperatury, środowiska beztlenowego, stałej ilości napływającej pożywki.
Przykładem mutualizmu pomiędzy roślinami i zwierzętami mógł być ścisły związek pomiędzy wymarłym już drontem z wyspy Mauritius Dodo i drzewem Calvaria major. Nasiona tego drzewa były zdolne do kiełkowania dopiero po przejściu przez układ trawienny ptaka. Po wyginięciu ptaka drzewo nie było już zdolne do rozprzestrzeniania się i rozwoju.
Innym rodzajem mutualizmu pomiędzy roślinami i zwierzętami jest zoogamia. Zjawisko to polega na przenoszeniu nasion bądź zapylaniu roślin poprzez działalność owadów latających lub niektórych ptaków (kolibry). Zwierzęta odnajdują w kwiatostanach smaczny nektar a przy okazji przenoszą pyłek kwiatów i przyczyniają się do rozmnażania rośliny.
Przykłady mutualizmu:
Zdjęcie 13 i 14. Porosty nadrzewne Chrobotek cienki (Cladonia macilenta)
(z lewej) i Pawężnica drobna (Peltigera didactyla)
Zapylacze kwiatów:
Zdjęcie 15 i 16. Kolibry z rodziny Archilochus colubris
Zdjęcie 17 i 18. Pszczoła miodna (Apis mellifera) (z lewej) i motyl Paź królowej (Papilio machaon).
Oddziaływania antagonistyczne
Amensalizm: Jest to zależność, w której populacja jednego gatunku hamuje rozwój drugiej populacji nie ponosząc przy tym strat ani nie uzyskuje korzyści. Jeden organizm oddziałuje na drugi pośrednio poprzez wytwarzanie związków, który w naturalny sposób uniemożliwia bądź hamuje wzrost drugiego organizmu. Związki te są najczęściej są wytwarzane na drodze metabolizmu wtórnego.
Między roślinami wyższymi część zależności amensalnych nazywa się allelopatią. Jedna z roślin wytwarza metabolit uniemozliwiający bądź hamujący wzrost drugiej. Przykładem allelopatii jest działanie orzecha włoskiego na niektóre rośliny jak zielne ziemniak, pomidor, niektóre zboża. Podobne działanie mają na siebie sosna i brzoza lub wrzos i dąb.
Wiele mikroorganizmów jest znanych ze zdolności do produkcji antybiotyków; znajdziemy wśród nich nie tylko promieniowce, lecz również grzyby czy bakterie właściwe. Antybiotyki są bardzo silnymi substancjami antydrobnoustrojowymi, hamującymi rozwój lub zabijającymi bakterie, grzyby jak również pierwotniaki i glony. Amensalizm będący wynikiem produkcji substancji antybiotycznych jest również nieraz określany jako antybioza. W środowiskach naturalnych, szczególnie w glebie, rozwój bardzo wolno rosnących promieniowców jest zależny od ich zdolności do wytwarzania antybiotyków. Współistnieć z nimi mogą jedynie szczepy, które posiadają mechanizmy detoksyfikacji (rozkład, przekształcenie ich do substancji nietoksycznych).
Wytwarzanie substancji antagonistycznych może być korzystne dla wytwarzającego je mikroorganizmu; osłabienie szybkości wzrostu wrażliwych partnerów lub ich eliminowanie daje producentowi szansę ekspansji w środowisku. Jest to szczególnie istotne dla mikroorganizmów wolno rosnących, które mają małe możliwości konkurowania z innymi mieszkańcami biocenozy.
Przykłady organizmów amensalnych:
Zdjęcie 19. Orzech włoski wytwarza duże ilości substancji hamującej wzrost roślin zielnych
Zdjęcie 20 i 21. Główni producenci antybiotyków: grzyb strzępkowy Penicillium expansum (z lewej) i promieniowiec Streptomyces sp.
Konkurencja: To interakcja zachodząca pomiędzy osobnikami o podobnych wymaganiach życiowych, zajmujących tę samą przestrzeń i korzystających z tych samych zasobów będących w niedomiarze. Są to osobniki wspólnej niszy ekologicznej. Konkurencja jest tym silniejsza im ściślej pokrywają się potrzeby życiowe. Często dotyczy osobników tego samego gatunku lub gatunków blisko spokrewnionych, lub gatunków niezwiązanych ze sobą ale o podobnych wymaganiach. Pamiętać jednak należy, że konkurencja występuje tylko w takich przypadkach, gdy zasoby substancji potrzebnej dla rozwoju obydwu grup są zbyt małe, aby zabezpieczyć potrzeby współistniejących organizmów. Współzawodniczące osobniki nie szkodzą sobie bezpośrednio, lecz walczą o zaspokojenie własnych potrzeb.
Szansa na wygraną w konkurencji zależy od szybkości wzrostu i rozmnażania, wytrzymałości na czynniki środowiskowe, wydajności energetycznej podczas metabolizowania składników pokarmowych, wymagań w stosunku do substancji wzrostowych, zdolności do gromadzenia substancji zapasowych i wykorzystywania ich, gdy środowisko ubożeje a także zdolności do tzw. ekspansji w środowisku.
U mikroorganizmów najostrzejsza konkurencja występuje między mikroorganizmami o podobnych parametrach wzrostu i podobnych wymaganiach pokarmowych. Sinice będą konkurowały o światło i CO2 z glonami, promieniowce o tlen i składniki odżywcze z pleśniami.
Partner słabszy, wolniej rosnący, o ubogim metabolizmie katabolicznym, wyższych wymaganiach pokarmowych musi przegrać w tej walce. Czynnikiem regulującym skład biocenozy, opartej o system konkurencji, jest najczęściej zawartość substancji organicznych, ilość i rodzaj związków azotowych, dostępność fosforu, tlenu, światła, wody lub przestrzeni życiowej. W mniejszym stopniu konkurencja dotyczy mikroelementów.
Konkurencja o przestrzeń życiową jest stosunkowo rzadka u mikroorganizmów i dotyczy głównie zależności między tlenowcami porastającymi powierzchnię. Rozwój części z nich stanowi barierę przestrzenną dla rozwoju innych. Ten rodzaj konkurencji występuje między różnymi gatunkami grzybów strzępkowych czy promieniowcami.
O zdominowaniu środowiska przez współzawodniczące mikroorganizmy decydują również parametry fizyczne środowiska. Temperatura, dostępność tlenu, aktywność wody w środowisku, intensywność promieniowania świetlnego np. w środowisku wodnym o tym samym składzie chemicznym, lecz z dostępem światła, mikroorganizmy fotoautotroficzne skutecznie konkurują z mikroorganizmami chemoorganotroficznymi, podczas gdy w zbiornikach zacienionych w środowisku dominują te drugie.
Również wytrzymałość na czynniki abiotyczne współdecyduje o możliwościach konkurowania mikroorganizmów. W środowiskach ekstremalnych większą szansę przetrwania w stanie aktywnym mają mikroorganizmy najodporniejsze, zdolne do przeżycia w stanie wysuszonym lub też w bardzo niskich czy wysokich temperaturach. Szczególnie oporne na takie warunki są formy przetrwalne mikroorganizmów.
Konkurencja w świecie roślin i zwierząt prowadzi w efekcie do wyeliminowania najsłabszego gatunku z niszy ekologicznej.
Rośliny żyjące na tym samym obszarze konkurują ze sobą o dostęp do światła, wody i składników mineralnych. Konkurencja między roślinami polega na wytworzeniu metabolitów uniemożliwiających bądź hamujących rozwój innych gatunków, mogących zagrozić roślinie swobodny rozwój.
W przypadku chemicznej konkurencji mamy do czynienia ze wspomnianą już przy amensaliźmie allelopatią. Allelopatia między roślinami widoczna jest szczególnie na polach uprawnych, gdzie rośliny uprawne walczą z szybko rozrastającymi się chwastami.
W przypadku konkurencji pomiędzy zwierzętami znaczenie ma głównie odporność gatunku, ten, który jest silniejszy, o większej tolerancji wygrywa. Przykładem mogą być stada roślinożerców (nawet tego samego gatunku) żyjące na tym samym terenie podczas pory suchej, gdy występuje deficyt pokarmowy.
Drapieżnictwo: Interakcja ta polega na chwytaniu, uśmiercaniu i zjadaniu osobników jednego gatunku przez osobniki drugiego gatunku.
Jest to jeden z głównych procesów regulujących liczebność populacji w biocenozie i warunkujący sprawny obieg materii. Procesy te zachodzą cyklicznie i warunkach naturalnych ( bez inwazji obcych gatunków) utrzymują równowagę ekologiczną w ekosystemie.
Drapieżnictwo wśród mikroorganizmów jest zjawiskiem rzadko występującym, Najbardziej typowym przykładem drapieżnictwa między drobnoustrojami jest odżywianie się pierwotniaków bakteriami. Jest to zjawisko szczególnie widoczne w zbiornikach wodnych, osadach czynnych i ściekach. W tym środowisku drapieżnictwo pierwotniaków jest uznawane jako efekt korzystny, pozwalający na redukcję ilości osadu czynnego. Główną rolę w eliminowaniu bakterii ściekowych przypisuje się orzęskom i wiciowcom. Obecność i odpowiednia ilość pierwotniaków w osadzie czynnym jest uznawana jako wskaźnik dobrze skojarzonej biocenozy. Podobne zależności można również spotkać w glebie, gdzie pierwotniaki żywią się bakteriami, do najpowszechniej spotykanych drapieżców należą wiciowce i ameby. Zależność drapieżca - ofiara występuje również między pierwotniakami i bakteriami w żołądku zwierząt przeżuwających.
Niektóre rośliny mogą być zaliczone do drapieżników. Przykładem może być tłustosz (Pinguicula vulgaris), który jest rośliną drapieżną - owadożerną. Wprawdzie jest fotoautofem, jednak żyjąc na podłożu ubogim w azot, uzupełnia pobór tego pierwiastka polując na zwierzęta. Górną powierzchnię liści, tworzących rozetkę, pokrywają gruczołowate włoski o lepkich główkach: przyklejają się do nich owady, które są potem trawione wydzieliną specjalnych gruczołów, a następnie wchłaniane przez liście. Większość roślin drapieżnych żyje na glebach torfowych, ubogich w łatwo przyswajalne związki azotu, więc na drodze ewolucji wykształciły mechanizmy zabijania, dzięki którym uśmiercają i trawią swoje ofiary. Łupem roślin mięsożernych padają stawonogi (głównie owady), ale także inne małe zwierzęta.
U zwierząt drapieżnictwo jest najlepiej widoczne. Istnieje mnóstwo gatunków drapieżników, a co za tym idzie ogromna różnorodność mechanizmów uśmiercania ofiar. Ofiary jednak nie pozostają bezbronne wobec drapieżników. Można wyróżnić dwa rodzaje obrony przed drapieżnikami: czynną i bierną. Obrona czynna polega na wczesnym ostrzeganiu np. alarmujące okrzyki ptaków, tupanie nogami przez zające, uderzanie o powierzchnię wody ogonem przez bobry, zastosowaniu środków psychologicznych np. stwarzanie fałszywych pozorów fałszywe plamy imitujące oczy u ryb i motyli, sprawianie wrażenia większego niż się jest w rzeczywistości (ryba nadymka), zastosowaniu środków mechanicznych np. kolce u jeży, kopyta i rogi u przeżuwaczy, środków chemicznych poprzez wrzucaniu toksycznych związków na bezpośrednio na napastnika. Obronę bierną dzielimy na: mimetyzm - upodabnianie się barwą, kształtem lub wzorem na ciele do środowiska (mątwy, patyczaki), mimikrę - upodabnianie się w toku ewolucji do zwierząt jadowitych i drapieżnych (niejadowity cylindrowiec koralowy upodobnił się do silnie jadowitego węża koralowego), zmianę barwy skóry dzięki komórkom pigmentowym (kameleon, głowonogi), poprzez kamuflaż stały np. przyklejanie do ciała muszelek, ziaren piasku (skąposzczety, skorupiaki, ryby).
Rośliny drapieżne:
Zdjęcie 22 i 23. Lejowate liście kapturnicy (Sarracenia sp.) owady wpadają do nich, ześlizgując się po gładkich wewnętrznych ściankach.
Zdjęcie 24 i 25. Lepka substancja na rzęskach rosiczki (Drosera aliciae) wabi potencjalne ofiary
Zdjęcie 26 i 27. Muchułówka (Dionaea muscipule) łapie swoje ofiary w specjalnie przystosowane liście
Przykłady drapieżników:
Zdjęcie 28. Doskonały wzrok, ostre szpony i dziób orła bielika pozwalają na wypatrzenie i schwytanie ofiary
Zdjęcie 29 i 30. Wielkie koty są doskonałymi mechanizmami do zabijania, na zdjęciu lampart z ofiarą (u góry) i gepard (poniżej), najszybsze zwierzę na Ziemi w pogoni za ofiarą.
Pasożytnictwo: Jest to interakcja rozumiana jako współzależność, w której jeden z organizmów (pasożyt) osiąga korzyści, natomiast drugi partner (gospodarz) nie ponosi lub ponosi szkody. W pierwszym przypadku pasożytnictwo dotyczy rozkładu martwych organizmów, np. szczątków roślin czy zwierząt. Ten rodzaj pasożytnictwa jest szeroko rozpowszechniony w przyrodzie i jest decydującym dla zapewnienia obiegu pierwiastków.
Drugi rodzaj pasożytnictwa występuje wtedy, gdy gospodarzem jest organizm żywy.
Pasożyt nie jest w stanie rozwijać się poza ciałem gospodarza, w stanie „wolnym” może występować jedynie w formach przetrwalnych.
Pasożyty można podzielić na zewnętrzne i wewnętrzne.
Przykładem tych pierwszych mogą być kleszcze, pchły, wszy, komary, pijawki (u zwierząt), jemioła, kanianka, huba (u roślin). Pasożyty wewnętrzne swój cykl rozwojowy lub jego fazę mogą przeprowadzić tylko wewnątrz organizmu żywiciela, prowadzi to do zmian w budowie pasożyta (zanikanie niektórych narządów, utrata zdolności syntezy niektórych związków np. u bakterii). Przykładami takich pasożytów mogą być: wirusy, tasiemce, robaki, przywry, nicienie, bakterie: prątki gruźlicy, dwoinka rzeżączki, pałeczki czerwonki, u roślin bakterie wywołujące niektóre choroby taki jak czarna zgnilizna kapusty, zgorzel jabłoni, wiele wirusów chorobotwórczych. Również wiele grzybów strzępkowych i drożdżaków może występować w formach pasożytniczych np. drożdże z rodzaju Candida sp., grzyby strzępkowe z rodzaju Aspergillus sp., Fusarium sp. i inne.
Rozwój pasożyta prowadzi często do utraty zdrowia lub całkowitego wyniszczenia organizmu gospodarza.
Przykłady mikroorganizmów pasożytniczych:
Zdjęcie 31 i 32. Bakterie dwoinka rzeżączki (Neisseria gonorrhoeae) (z lewej) i pałeczka dżumy (Yesrinia pestis)
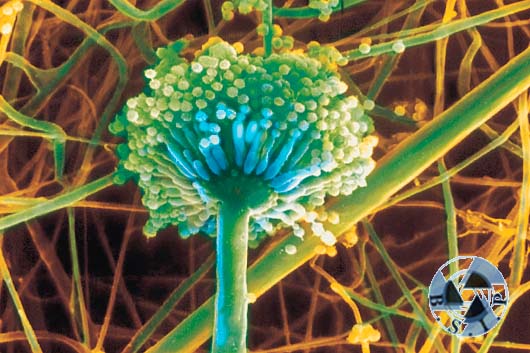
Zdjęcie 33. Pasożyt roślinny grzyb z rodzaju Aspergillus, może czasami atakować także człowieka i wywołać chorobę zwana aspergillozą.
4. Oddziaływania biochemiczno-ekologiczne między gatunkami
Oddziaływania między mikroorganizmami
Drobnoustroje obustronnie w walce pomiędzy sobą wykorzystują biochemiczne środki ataku i obrony. Do tej pierwszej kategorii można zaliczyć enzymy: przede wszystkim lityczne i proteolityczne, powodujące np. lizę ściany komórkowej przeciwnika. Do drugiej kategorii zalicza się antybiotyki wykorzystywane raczej w walce obronnej, choć mogą też być wykorzystane w celu pozbycia się konkurenta z danego ekosystemu.
Przykładowo, bakterie z rodzaju Bacillus są bardzo szeroko rozpowszechnione w świecie (praktycznie biorąc, można je spotkać wszędzie). Z jednej strony wynika to z faktu, iż łatwo adoptują się do różnorodnych warunków środowiskowych i ponadto tworzą przetrwalniki. Z drugiej strony wiele ich gatunków wykazuje zdolność do biosyntezy całej gamy antybiotyków oraz subtilizyny, enzymu o silnych właściwościach proteolitycznych.
Antybiotyki bakterii Bacillus mają budowę peptydową. Jako przykład można wymienić gramicydynę S, pierścieniowy dziesięciopeptyd wytwarzany przez Bacillus brevis.
Bakterie Bacillus brevis wytwarzają jeszcze inne antybiotyki, m.in. tyrocydyny A, B i C, będące również cyklicznymi oligopeptydami, zawierającymi -D-fenyloalaninę, podobnie jak gramicydyna S. Znanych jest obecnie ponad 25 antybiotyków wytwarzanych przez różne szczepy tych bakterii. Innym przykładem antybiotyku o budowie peptydowej jest bacytracyna, wytwarzana przez bakterie Bacillus subtilis. Poszczególne szczepy bakterii Bacillus subtilis produkują ponad 65 antybiotyków o budowie polipeptydowej. Z kolei Bacillus polymyxa wytwarza polimyksyny.
Wszystkie te antybiotyki działają na bakterie gramujemne, gramdodatnie i grzyby. Ich działanie jest różne. Niektóre blokują syntezę ściany komórkowej lub zakłócają funkcje błon biologicznych, inne, mniej liczne, zakłócają replikację, transkrypcję lub translację.
Antybiotyki są wytwarzane również przez wiele innych mikroorganizmów i grzybów, w tym przez promieniowce Streptomyces i pleśnie Penicillium.
Antybiotyki dzieli się w zależności od budowy i od sposobu działania na mikroorganizmy. Ze względu na budowę i mechanizm działania wyróżnia się:
- antybiotyki beta-laktamowe (penicyliny, cefalosporyny, tienamycyna): antybiotyki te mają wiązanie beta-laktamowe, które ulega rozerwaniu pod wpływem enzymów, tzw. beta - laktamaz produkowanych przez niektóre bakterie. Wszystkie antybiotyki beta-laktamowe hamują biosyntezę ściany komórki bakteryjnej, co prowadzi do jej rozpadu. Antybiotyki te są wytwarzane przez pleśnie z rodzajów Penicillum sp., Cephalosporium sp., a tienamycyna przez promieniowca Streptomyces cattleya.
- makrolidy (erytromycyna i pochodne) i linkozamidy: antybiotyki te przenikają przez błony biologiczne i ściany komórkowe, hamują biosyntezę białka w komórce bakteryjnej poprzez łączenie się z podjednostką rybosomu. Wykazują działanie na bakterie Gram dodatnie, a także na beztlenowce. Erytromycyna wytwarzana jest przez promieniowca Streptomyces erythreus.
Erytromycyna
- tetracykliny: mechanizm działania tych antybiotyków polega na hamowaniu biosyntezy białka i procesów energetycznych (przyłączania fosforu) w komórkach bakteryjnych, największe znaczenie odgrywają w leczeniu zakażeń wywołanych przez bakterie atypowe tj. Chlamydiae, Rikettsiae, Mycoplasma. Tetracykliny wytwarzane przez promieniowca Streptomyces aureofaciens.
- aminoglikozydy (streptomycyna): działanie tych antybiotyków polega na upośledzeniu syntezy białek bakteryjnych (powstają białka o nieprawidłowej budowie) oraz uszkodzeniu błony cytoplazmatycznej. W porównaniu z antybiotykami beta - laktamowymi aminoglikozydy mają znacznie węższy zakres działania, nie działają na beztlenowce, a ich zaletą jest to, że oporność bakterii na te antybiotyki powstaje powoli, co zwiększa ich skuteczność, działają głównie na bakterie Gram ujemne. Streptomycynę wyizolowano w roku 1944 z promieniowca Streptomyces griseus.
Streptomycyna
- chloramfenikol: hamuje biosyntezę białek i lipidów w komórkach bakteryjnych. Syntezę chloramfenikolu prowadzi promieniowiec Streptomyces venezualae.
Chloramfenikol
Dodatkowo inny rodzaj oddziaływań może występować w świecie grzybów. Ich metabolity wtórne mogą być wykorzystywane jako substancje sygnalne w układach grzyb-pasożyt kontra ofiara. Np. metabolity wytwarzane przez potencjalną ofiarę mogą stymulować wzrost strzępek grzybni grzyba-pasożyta lub odwrotnie, metabolity grzyba-pasożyta stymulują rozwój strzępek grzybni ofiary w kierunku kolonii pasożyta.
Innym rodzajem metabolitów wtórnych u pleśni są mikotoksyny, których znaczenie nie jest jeszcze do końca poznane. Do tej pory wyodrębniono około 400 mikotoksyn, mają one różnorodne efekty toksyczne i posiadają wysoką odporność na działanie wysokiej temperatury. Mykotoksyny z pewnością są substancjami mutagennymi i teratogennymi. Liczne gatunki pleśni należące przede wszystkim do rodzajów Aspergillus, Penicillium, Fusarium i inne są zdolne do wytwarzania mikotoksyn. Do najbardziej znanych mikotoksyn należą: aflatoksyny, ochratoksyny, trichoteceny, patulina oraz fumonizyny, zearalenon i toksyna T-2. Jak podają źródła naukowe mikotoksyny są wytwarzane w fazie stacjonarnej rozwoju pleśni , od niedawna wiadomo również, że grzyby wydzielają mikotoksyny nie tylko do podłoża na którym rosną ale ich bardzo duża ilość gromadzona jest w konidiach. Założono więc tezę, iż mikotoksyny mogą być rezerwuarem energii dla kiełkujących zarodników. Jednak badania, które przeprowadziłam w trakcie pracy inżynierskiej na szczepach pleśni toksynotwórczych wykazały że konidia nie wykorzystują mikotoksyn w procesie kiełkowania.
Istnieje również podejrzenie, że mikotoksyny mają wpływ na kiełkowanie konidiów innych gatunków pleśni, być może spełniają one rolę inhibitorów kiełkowania, ale zagadnienie to pozostaje w sferze badań.
Nieznane jest działanie mikotoksyn na inne drobnoustroje, ale coraz częściej mówi się o ich niekorzystnym działaniu przede wszystkim na zwierzęta i człowieka, więc omówię je w rozdziale poświęconym oddziaływaniom pomiędzy mikroorganizmami i zwierzętami.
Allelopatia między roślinami wyższymi
Allelopatia jak już wcześniej opisywałam obejmuje oddziaływania pomiędzy roślinami, zachodzące przy udziale związków chemicznych tak zwanych czynników allelopatycznych. Związki te wydzielane są przez rośliny lub glony i mają wpływ na kiełkowanie bądź rozwój innych.
Najlepszym przykładem allelopatii może być oddziaływanie Orzecha czarnego (Juglans nigra) i jego europejskiego krewniaka Orzecha włoskiego (Juglans regia) na niektóre rośliny takie jak ziemniak, pomidor, niktóre zboża, sosna, olcha i inne. Orzech wydziela bardzo duże ilości substancji (4-glikozyd
1,4,5-trihydroksynaftalenu), która w glebie ulega hydrolizie i utlenieniu do naftochinonu o nazwie juglon. Juglon jest rozpuszczalnym w wodzie barwnikiem, powoduje brązowienie rąk podczas krojenia świeżych młodych orzechów, występuje on w częściach zielonych drzewa i zanika w obumarłych tkankach i dojrzałych orzechach. Juglon przede wszystkim działa toksycznie już na nasiona innych roślin, jest inhibitorem ich kiełkowania, powoduje również zamieranie sadzonek w bezpośrednim sąsiedztwie drzewa. Przedostanie się związanej formy toksyny jest uwarunkowane opadami deszczu, który wymywa ją z zielonych częsci rośliny. Dopiero po przekształceniu w glebie toksyna staje się aktywna biochemiczno-ekologicznie. Juglon działa inhibująco i toksycznie już w bardzo niewielkich stężeniach (0,002%), lecz są gatunki takie jak Rubus fruticosus czy Poa pratensis które są odporne na jego działanie i mogą rosnać bez przeszkód w bliskim sąsiedztwie orzecha.
Bardzo wyraźnie widoczy jest allelopatyczny wpływ na rośliny zielne krzewów Salvia leucophylla i Artemisia californica na obszarach makii kalifornijskiej. Działanie krzewów powoduje całkowity zanik wegetacji w odległości od jednego do dwóch metrów od rosnących krzewów. Za zjawisko to odpowiedzialne są toksyny o strukturze terpenów. Zwiąki te, a szczególnie proste monoterpeny takie jak 1,8-cyneol, kamfora, α-pinen, β-pinen, α-tujon i kamfen, z których dwa pierwsze sa najbardziej aktywne występują w dużych ilościach w liściach krzewów, są wydzielane w sposób ciągły i występują w glebie wokół rośliny. Obserwacje allelopatycznego działania tych związków można prowadzić na tym terenie w sposób cykliczny ponieważ makię kalifornijską nawiedzają cykliczne średnio co 25 lat naturalne pożary. Nagromadzone terpeny należą do węglowodorów i w czasie pożaru szybko odparowują i zapalają się pozostawiając glebę oczyszczoną. Na takim podłożu szybko rozwijają się rośliny zielne i trawy, jednak powolny wzrost krzewów powoduje stopniowe zamieranie roslinności wokół krzewów i powstanie gołych powierzchni.
1,8-cyneol kamfora
Zjawisko allelopatii można zauważyć również na polach uprawnych. Trawy i chwasty rosnące wśród roślin zielnych często konkurują z nimi o wodę i światło nie tylko za pomocą szybkości wzrostu i adoptowania się do istniejących warunków ale również dzięki toksycznym substancjom jakie wydzielają do podłoża.
Czynniki allelopatyczne nie muszą mieć tylko i wyłącznie wpływu na inne rośliny, ale również mogą działać stymulująco na wydzielającą je roślinę. Na przykład badania przeprowadzone w Wielkiej Brytanii w 1975 roku przez Newmana i Rovira wykazały, że gatunek Anthoxanthum odoratum rośnie znacznie szybciej w obecności swoich wydzielin niż w każdych innych warunkach. Z kolei zupełnie odwrotne działanie ma kofeina, która wykazuje toksyczność w stosunku do roślin kawy. Zjawisko to nazwano autotoksycznością często obserwowaną w rolnictwie. Niektóre związki allelopatyczne takie jak kofeina wydzielane do gleby trudno poddają się rozkładowi przez mikroorganizmy. Na plantacjach kawy alkaloid purynowy - kofeina zalegająca w glebie jak i w obumarłych liściach nie pozwala na ponowne obsianie terenu ze względu na autotoksyczne działanie.
Biochemiczno-ekologiczne oddziaływania występują również w układach gospodarz-pasożyt wśród roślin.
Pasożyty roślinne mają zdolność wytwarzania organów inwazyjnych zwanych haustoriami, za pomocą których przyczepiają się one do rośliny-gospodarza i mogą korzystać z jej metabolitów. „Wybór” gospodarza przez pasożytniczą roślinę jest uwarunkowany rodzajem metabolitów jakie gospodarz wydziela do środowiska. Przykładami popularnych pasożytów roślinnych mogą być gatunki jemioły (Viscum), kanianki (Custuta), i pasożytów korzeniowych Orobanche i Striga. Warunkiem wykiełkowania i wytworzenia haustoriów przez te pasożyty jest otrzymanie sygnału chemicznego wydzielanego przez nieczego nie podejrzewającego gospodarza. Sygnał ten pozwala pasożytowi rozpoznać organizm gospogarza. Takim stymulatorem kiełkowania nasion Striga jest wyizolowany z bawełny strigol lub podobny do niego strukturalnie sorgolakton wytwarzany przez Sorghum bicolor (głównego gospodarza Striga).
Gdy pasożyt wykiełkuje potrzebuje kolejnego sygnału niezbędnego do wytworzenia haustoriów, czyli wydzielenia związków indukujących wytworzenie organów inwazyjnych. Przykładem takiego działania może być wydzielanie przez roślinę motylkową z gatunku Astragalus dihydrostilbeny o nazwie ksenognizyna A i spokrewnionego z nią izoflawonu o nazwie ksenognizyna B. Substancje te indukują wytwarzanie haustoriów przez pasożyta zgatunku Agalinis purpurea.
Allelopatia jest zjawiskiem powszechnym i może zachodzić w wielu sytuacjach ekologicznych. Głównym powodem występowania tego zjawiska jest ogromna różnorodność metabolitów wtórnych wytwarzanych przez rośliny i ich wpływ na rozwój innych roślin. Znany jest również fakt że niemal wszystkie okrywy nasienne zawierają związki hamujące kiełkowanie nasion, utrzymują one nasiona w stanie uśpienia.
Interakcje roślina wyższa-mikroorganizm
Oddziaływania te mają najczęściej charakter zaatakowania rosliny przez mikroorganizm i wywoływania u niej chorób. Ma tutaj miejsce swoista walka na związki chemiczne: mikroorganizmy wywołują choroby u roslin poprzez wydzielanie związków zwanych patotoksynami (fitotoksynami), natomiast rośliny zabezpieczają się przed inwazją wytwarzając fitoaleksyny.
Patotoksyny (fitotoksyny)
Fitotoksyny są metabolitami mikroorganizmów zdolnymi do wywołania jednego lub więcej symptomów chorobowych w roślinie. Zdolność do syntezy tych związków określa agresywność patogena, jego zdolność do uszkadzania tkanek. Patotoksyny moga być związkami o małej masie cząsteczkowej i te właśnie mają wpływ na wzrost lub powodują więdnięcie rośliny, a także mogą należeć do grupy związków wysokocząsteczkowych takich jak peptydy wywołujące nekrozę czy enzymy powodujące macerowanie tkanek i utratę spójności komórek w roślinach. Zarówno jeden jak i drugi rodzaj patotoksyn może być wytwarzany przez ten sam organizm. Symptomy działania fitotoksyn to często zakłócenia syntezy niektórych enzymów w roslinie, anomalie wzrostowe, więdnięcie czy nekroza. Przykładem pierwszego z wymienionych objawów może być chloroza tytoniu, która jest wynikiem rozkładu chloroplastów. Zjawisko to może być skutkiem akumulacji amoniaku w tkankach, to z kolei jest wywołane przez działanie fitotoksyny peptydowej, wytwarzanej przez bakterie Pseudomonas tabaci zakłócającej syntezę enzymu spełniającego ważną rolę w metaboliźmie azotu - syntetazy glutaminianowej.
Częstymi patogenami roślin są grzyby i tak na przykład gatunek Fusarium oxysporum wytwarza toksyny powodujące więdnięcie pomidora - kwas fuzariowy oraz likomarazmina. Obie toksyny mają właściwości chelatujące metale i ich synteza jest uwarunkowana obecnością jonów niekótrych metali.
Pomidor w walce z grzybem nie pozostał w tyle, niektóre jego odmiany wykształciły zdolność wiązania kwasu fuzariowego z glicyną co powoduje dezaktywację toksyny.
Warto wspomnieć, że mikroorganizmy bardzo często posiadają zdloności do wytwarzania szerokiej gamy enzymów (które w tym przypadku nazwiemy toksynami mokromolekularnymi ze względu na ich budowę). Aktywność enzymatyczna mikroorganizmów wpływa destrukcyjnie na tkanki rosliny zainfekowanej. Macerację tkanek czyli utratę spójności komórek, w wyniku której dochodzi do rozdzielenia się komórek można obserwować na niemal każdyno gnijącym owocu czy warzywie, jest ona wynikiem działania enzymów (celulaz, hemicelulaz i pektynaz) wytwarzanych przez drobnoustroje.
Fitoaleksyny
Fitoaleksyny są w pewnym sensie tarczą obronną roślin przed inwazją drobnoustrojów. Związki te są metabolizowane i kumulowane w tkankach rośliny i mają spełniać funkcję ochronną. Najlepszym przykładem może być tutaj nagromadzenie metabolitów wtórnych w zdrewniałej tkance drzew liściastych. Większość z tych tkanek wykazuje odporność na rozkład, więc można stwierdzić, że związki w nich występujące mają charakter odpornościowy. Na przykład u rodzaju Pinus występuje pinosylwina należąca do grupy hydroksystilbenów, które są odpowiedzialne za odporność rośliny na grzyby. U roślin niedrzewiastych można przytoczyć katechinę i kwas protokatechowy występujące w łuskach cebuli, odpowiedzialne za odporność w stosunku do grzyba Colletotrichum circinas wywołującego antrakozę.
Tę grupę fitoaleksyn można uważać za przedinfekcyjną, czyli związki mające za zadanie ochronić roślinę przed infekcją. Ale istnieją również związki poinfekcyjne tak zwane postinhibityny. W zdrowych tkankach występują one w postaci nieaktywnych glikozydów z których w trakcie inwazji patogena uwalniane są na drodze hydrolizy enzymatycznej lub utlenienia aktywne toksyny. Przykładem mogą tu być glikozydy cyjanogenne, które podczas hydrolizy enzymatycznej uwalniają kwas pruski. U komornicy zwyczajnej (Lotus corniculatus) ten proces zachodzi w następujący sposób:
Uwolniony HCN ma za zadanie chronić roślinę przed dalszą infekcją, lecz na przykład Stemphylium loti toleruje obecność HCN i ponadto wytwarza enzym - hydrolazę formamidową, detoksykującą HCN poprzez przekształcenie go w formamid HCONH2. Wyizolowano dużo fitoaleksyn poinfekcyjnych, których synteza ma spowodować zahamowanie inwazji drobnoustroju, można do nich zaliczyć orchinol z bulw storczyków, ipomeamaron z batatów i inne.
Jednak istnieje siedem zasad sformułowanych przez Mullera i Borgera w 1941roku dotyczących teorii fitoaleksyn:
Fitoaleksyną nazywamy związek, który hamuje rozwój patogena we wrażliwych tkankach i powstaje lub jest aktywowany tylko wtedy, gdy roslina gospodarza wejdzie w kontakt z pasożytem.
Reakcje obronne zachodzą tylko w komórkach żywych
Czynnik hamujący jest odosobnionym związkiem chemicznym, produktem komórki gospodarza
Fitoaleksyna jest związkiem niespecyficznym w swoim toksycznym oddziaływaniu na grzyby, jednakże gatunki grzybów mogą wykazywać zróżnicowaną wrażliwość na ten związek
Podstawowa reakcja komórek wrażliwych i odpornych jest taka sama, różnice między gospodarzem wrażliwym i odpornym polegają na szybkości formowania fitoaleksyn
Reakcja obronna ogranicza się do tkanek zaatakowanych i do ich najbliższego sąsiedztwa
Odporność nie jest dziedziczona, wykształca się ona po próbie infekcji przez patogena. Wrażliwość komórek gospodarza określająca szybkość jego reakcji jest specyficzna i określona genotypowo.
Interakcje roślina wyższa - zwierzę
Pierwszą grupą oddziaływań roślin na zwierzęta można nazwać oddziaływania przyciągające; ma to szczególną rolę gdy mowa jest o zapylaczach czyli głównie owadach ale również ptakach i niektórych gatunkach nietoperzy. Roślina wysyła sygnały do zapylacza w postaci zapachu, barwy kwiatów a na końcu oczywiście kusi smakiem nektaru czy pyłku. Jest to współzależność w której zarówno roślina jak i zapylacz odnoszą korzyści.
Nie byłoby jednak tych sygnałów gdyby nie aspekty biochemiczne. Jest to temat niezwykle rozbudowany z uwagi na ogroną różnorodność zarówno zapylaczy i ich roli jak i indywidualnych gatunkowo sposobów wabienia ich przez rośliny.
Pierwszym znakiem dla owada zapylającego jest barwa kwiatów. Barwy kwiatów takie jak pomarańczowa, czerwona i niebieska a również zółta i biała są uwarunkowane obecnością w kwiatach barwników roślinnych z grupy flawonoidów. Również karotenoidy odpowiadaja w dużej mierze na kolory od żółtego po czerwony. Na przyciąganie zapylaczy mają wpływ również tak zwane wskaźniki lokalizacji nektaru które są w rzeczywistości barwnymi wzorami na kwiatach a ich zadaniem jest kierowanie zapylacza w stronę nektaru i organów płciowych. Zakres widzenia na przykład pszczół miodnych pozwala im na przykład odczytywać różnice w absorbcji promieniowania w zakresie UV flawonów i flawonoli, występujących w białych kwiatach i jako kopigmenty w niebieskich kwiatach. Pszczoły miodne preferują głównie kwiaty o zabarwieniu określanym przez nas jako żółte i niebieskie. Wybierają te kwiaty nawet wtedy, gdy mają do wyboru szeroką gamę kolorów. Z kolei kolibry mają skłonność do wybierania kwiatów czerwonych, na przykład róży chińskiej (Hibiscus). Ptaki te w niektórych środowiskach mogą odwiedzać również białe kwiaty, ale z reguły wolą czerwień, nawet ich upierzenie też często jest podobnie ubarwione aby lepiej zamaskować przed drapieżnikami zbierające nektar ptaki. Motyle preferują barwy jaskrawe natomiast ćmy stonowane. Inne zapylacze są mniej wrażliwe na barwę lub zapylają nocą i to inne czynniki warunkują ich wybór.
Innym bardzo ważnym czynnikiem wpływajacym na przywabienie zapylacza jest zapach. Zapylaczy dziennych przyciagają wonne i odurzające zapachy, natomiast zapylaczy nocnych przyciagają zapachy nieprzyjemne.
Zapachy przyjemne wywołane są przez frakcję występujących w kwiatach olejków eterycznych. Są to związki organiczne w większości mono i seskwiterpeny, a także lotne substancje aromatyczne, proste alkohole alifatyczne, ketony i estry. Zapach to najczęściej mieszanina kilku substancji.
Zapachy nieprzyjemne naśladują odór rozkładającego się białka i kału. Do najbardziej przykrych należą barszcz, ciemiernik i obrazki plamiste. Przyciągają one głównie owady żerujące na odpadach i oborniku. Głównymi składnikami nieprzyjemnych zapachów są monoaminy: metyloamina, etyloamina itp, diaminy: putrescyna i kadaweryna a także indol i skatol, a także niektóre alifatyczne kwasy organiczne takie jak izomasłowy.
Nieprzyjemne zapachy nie wszystkim roślinom jednak służą jako wabik dla zapylacza, ale również jako wobik dla ofiary. Często rośliny drapieżne wykorzystują nieprzyjemny zapach by przyciągnąć owady, które wpadają w pułapkę i zostają wykorzystane jako źródło azotu.
Owady, które porozumiewają się między sobą organicznymi substancjami lotnymi o nazwie feromonów są podatne na zapylanie kwiatów wydzielających związki identyczne bądź bardzo podobne do feromonów. Roślina w pewien sposób oszukuje owada przywabiając go takimi związkami. Na przykład Cassia fitulosa wytwarza ester metylowy eugenolu, który jest jednocześnie feromonem płci u muszki owocowej Dacus dorsalis. Metyloeugenol jest wytwarzany przez niektóre kwiaty jako składnik olejków eterycznychuwalnianych z liści. Muszka jest bardzo wrażliwa na zapach tych kwiatów. Poza zapachem i barwą głównym powodem dla których zwierzęta odwiedzają kwiaty są nektar i pyłek, bogate w cukry, przede wszystkim glukozę, fruktozę i sacharozę oraz niewielkie ilości oligosacharydów. Wiadomo również że nektar zawiera inne cukry w niewielkich ilościach: maltozę, trehalozę, melezitozę oraz co ciekawe w nektarze mozna znależć wszystkie powszechne grupy aminokwasów. Nektary niektórych roślin zawierają również lipidy. Pyłek kwiatowy natomiast bogaty w białka, skrobię, wolne cukry i lipidy.często zawiera również karotenoidy i flawonoidy.
Bardzo interesującym wydaję się fakt, że niektóre rośliny uwalniają tak zwane wabiace związki lotne, szczególnie atrakcyjne dla niektórych pasożytów roślinożerców lub drapieżników. Jest to swoiste wołanie o pomoc w sytuacjach, gdy roślina zostaja zaatakowana przez roślinożerne owady przykładem może być tutaj reakcja bawełny na atak pędziorków.
Druga grupa oddziaływań pomiędzy roślinami wyższymi i zwierzętami odnosi się do wytwarzania przez rośliny substancji toksycznych bądź odstraszających roślinożerców. Mechanizmy obronne roślin można podzielić na fizyczne i chemiczne. Czynnikami fizicznymi mogą być np: twarda epiderma, kolce, igły, ciernie i parzące włoski. Czynniki chemiczne są niezwykle zróżnicowane, istnieje cała gama toksyn wytwarzanych przez rośliny w obronie własnej. Generalnie toksyny roslinne dzielimy na dwie klasy: takie, które w swojej cząsteczce zawierają azot oraz bezazotowe.
Toksyny zawierające azot:
Aminokwasy niebiałkowe - występuje ich około 300, ich występowanie jest najbardziej charakterystyczne dla roślin motylkowych (głównie w nasionach), chociaż występują także u innych, odległych gatunków. Występowanie tych toksyn w nasionach można uzasadnić tym, że nasiona są źródłem wielu odżywczych składników pokarmowych, a roslina chcąc uchronić się przed zjadaniem nasion kumuluje w nich toksyczne substancje.
Przykładami takich związków mogą być L-DOPA pochodzący z nasion Mucuna działający toksycznie na owady, szczególnie na chrząszcze strąkowcowe Prodenia eridania. Innym takim związkiem może być
β-cyjanoalanina w nasionach Vicia wykazyjąca toksyczność wobec ssaków roślinożernych. Niektóre zwierzęta posiadają zdolność detoksykacji niebiałkowych aminokwasów. Na przykład chrząszcze Caryedes brasiliensis, które żywią się nasionami zawierającymi duże ilości kanawaniny. Chrząszcze te dysponują mechanizmem ochronnym przeciwko tej toksynie.
Glikozydy cyjanogenne - są to strukturalnie proste toksyny , ich aktywność zależy od enzymów, rozkładających je do kwasu pruskiego HCN. HCN działa w systemie cytochromowym i prowadzi do zahamowania oddychania, które z kolei przyczynia się do śmierci komórki niedotlenionej. Podobną do HCN toksyczność wobec organizmów wykazują azotyny, a niktóre gatunki roślin np. Astragalus akumulują glikozydy organicznych nitrozwiązków, które są źródłem azotynów.
Glukozynolany - inaczej glukozydy olejków gorczycznych, są to związki blisko spokrewnione z glikozydami cyjanogennymi, występują na przykład u Brassica. Ich toksyczność polega na zaburzaniu pracy żołądka, wywołaniu ślinotoku, biegunki i podrażnienia jamy ustnej.. Toksyczność jest związana z wydzielaniem izotiocyjanianów czyli olejków gorczycznych, które wywołują wyżej wymienone objawy. Glukozynolany mogą również podlegać przegrupowaniu do tiocyjanianów, które powodują powiększenie tarczycy u ssaków.
Alkaloidy - jest to najbardziej znana grupa toksyn roślinnych, są one w większości silnie toskyczne.i teratogenne na ssaki i człowieka. Alkaloidy mogą być strukturalnie podobne do cukrów, przez co mogą hamować aktywność enzymów metabolizmu cukru zwierząt. Do najbardziej znanych alkaloidów należą: solanina (Solanum tuberosum) i strychnina (Strychnos nux-vomica) lunb atropina (Atropa belladonna), deoksymannojirymycyna (Lonchocarpus sericeus) przypominająca mannozę.
Białka - powszechnie uważa się, że białka roślinne nie są toksyczne, jednak kilka z nich jest bardzo niebezpiecznych dla zwierząt i ludzi.Przykładem takim może być abryna występująca w nasionach Abrus precatorius, jej dawka śmiertelna dla człowieka to już pół miligrama. Innym toksycznym białkiem może być rycyna (Ricinus communis).
Toksyny bezazotowe:
Toksyny roślinne nie muszą być skomplikowanymi związkami zawierającymi azot, istnieje wiele związków trujących, które sa terpenoidami lub prostymi węglowodorami. Zaklasyfikowano je do drugiej klasy toksyn - toksyn bezazotowych. I tak należą do niech:
Glikozydy nasercowe - te steroidowe związki są substancjami działającymi na serce i tu przykładem może być ouabaina w Acokanthera ouabaio lub oleandryna z Nerium oleander. Glikozydy nasercowe hamują działanie adenozynotrifosfataz błonowych i wywierają silny wpływ na wymianę niektórych jonów, mają także działanie kardiotoniczne.
Saponiny - przykładem może tutaj być kwas medikagenowy w liściach Medicago sativa, toksyna oddziałująca na ryby i owady.
Chinony - są związkami powodującymi uczulenie na światło, i tak hiperycyna z Hypericum perforatum jest związkiem fotodynamicznym, po dostaniu się do krwiobiegu oddziaływuje ze światłem powodując oparzenia, uszkodzenia i odbarwienia skóry, w efekcie prowadzi do śmierci zwierzecia.
Furanokumaryny - podobnie jak chinony sa to związki fotodynamiczne, ale działają w głównej mierze na owady
Iridoidy - związki te występują w roślinie w stanie wolnym lub w formie glikozydowej. Przykładem iridoidu może być aukubina w liściach Aucuba japonica, wykazująca toksyczniość wobec owadów i ptaków.
Laktony sekswiterpenów - wiele z tych związków wywołuje alergie skórne i dizła odstraszająco, niektóre z nich działają trująco na zwierzęta gospodarskie i tutaj przykładem może być Hymenowina w Hymenoxys odorata.
Taniny - Do tanin zaliczane są polifenole, rozpuszczalne w wodzie, zdolne do wytrącania z roztworów wodnych białek, a także kwasów nukleinowych, alkaloidów i niektórych polisacharydów. Właściwości charakterystyczne dla tanin wykazują związki polifenolowe zbudowane co najmniej z pięciu pierścieni aromatycznych i zawierające dwanaście lub więcej grup fenolowych. Włyróżnia się dwie grupy tych związków:
Taniny hydrolizujące - poliestry kwasu galusowego i jego pochodnych z monosacharydami lub innymi poliolami, a najczęściej z glukozą.
Taniny niehydrolizujące - skondensowane, zwane również proantocyjanidynami, zbudowane z cząsteczek katechin połączonych wiązaniami C-C, które nie ulegają rozerwaniu w warunkach hydrolizy.
Głównymi czynnikami wiązania się tanin z białkami jest hydrofobowa interakcja między aromatycznymi pierścieniami tanin i hydrofobowymi fragmentami - wnękami występującymi w cząsteczkach białek oraz wiązania wodorowe między grupami hydroksylowymi tanin, a grupami karbonylowymi wiązań peptydowych białek. Dodatkowym czynnikiem stabilizującym kompleksy tanin z białkami mogą być wiązania kowalencyjne, które tworzą się między utlenionymi grupami fenolowymi tanin i nukleofilowymi grupami białek takimi jak: -SH, -OH, -NH .
Efektywność wiązania polifenoli z białkami zależy od struktury obydwu reagentów. Biologicznie ważnym efektem interakcji tanin z białkami jest blokowanie aktywności enzymów. Prawdopodobnie główną przyczyną zwiększonej biosyntezy tanin u roślin, w okolicy uszkodzonych tkanek, jest obrona przed destrukcyjnym działaniem enzymów wydzielanych przez atakujące je drobnoustroje. Taniny jako inhibitory enzymów mogą powodować blokowanie enzymów w przewodzie pokarmowym. Dotyczy to zwłaszcza inhibicji enzymów proteolitycznych przez taniny występujące w dużych ilościach w niektórych surowcach stanowiących ważne źródło białka dla organizmu np. nasion roślin strączkowych. Zmniejszenie trawienia białka tych produktów może być również spowodowanetym, że jest ono powiązane z taninami czyli częściowo zdenaturowane. W herbacie duża zawartość tanin powoduje iż ograniczone jest przyswajanie niektórych metali, głównie żelaza, które tworzy z taninami trwałe kompleksy, nie ulegające rozkładowi w przewodzie pokarmowym. Taniny osłabiają właściwości wydzielnicze błony śluzowej żołądka i jelit a więc powodują zaparcia. Taniny blokują także działanie -amylazy w jamie ustnej co powoduje zahamowanie włchaniania niektórych substancji pokarmowych. Taniny odstraszają owady oraz kręgowce przez swój charakterystyczny ściągający efekt, działają również przeciwko drobnoustrojom.
Przykłady tanin:
Toksyny roślinne nie mają tylko funkcji trującej czy odstraszającej w stosunku do zwierząt. Wiele zwierząt, a w szczególności owadów wykorzystuje toksyny do własnych celów, np jako feromon płciowy, jako obronę przed drapieżnikami, mogą być wydzielane przez owady jako czynnik maskujący lub ostrzegawczy. Niektóre owady potrafią prowadzić syntezę de novo toksyny.
Ciekawym zjawiskiem są również zależności hormonalne lub feromonalne pomiędzy roślinami i zwierzętami. Na przykład w układzie pomiędzy owadami i roślinami te drugie wytwarzają dwa typy hormonów owadzich: linienia i juwenilne. Roślina często wytwarza związki o strukturze bardzo zbliżonej do danego hormonu, bądź substancje zakłócające syntezę tych hormonów u owada. Również interkacje hormnonalne - erstogenne mogą zachodzić w przypadku ludzi, owiec, gryzoni i ptaków. Niewielkie ilości ludzkich hormonów męskich (testosteron i androsteron) i żeńskich (estron i estriol) także wykryto w roślinach. Jednak nie jest do końca poznana przyczyna występowania tych związków w roślinach..
Interakcje zwierzę - mikroorganizm
Ten rodzaj oddziaływań można podzielić na dwie grupy. Pierwsza to korzystny dla obu stron układ druga to nierównowaga pomiędzy mikroorganizmem a zwierzęciem czyli stan chorobowy.
W przypadku korzystnego układu pomiędzy mikroorganizmami i zwierzętamia można przytoczyć przykład działania mikroflory przewodu pokarmowego roślinożerców. Uzdolnienia enzymatyczne drobnoustrojów wchodzących w skład tej mikroflory pozwalają zwierzęciu na przyswojenie składników pokarmowych pochodzących z substancji, których on sam nie byłby zdolny hydrolizować enzymatycznie. Takim ekosystemem, gdzie te zależności występują szczególnie wyraźnie, jest przewód pokarmowy. Zwierzęta żywiące się wyłącznie materiałem roślinnym bogatym w błonnik zawierają w swoim przewodzie pokarmowym mikroorganizmy symbiotyczne trawiące ten sacharyd. Najciekawszy zespół mikroorganizmów występuje u przeżuwaczy. Przeżuwacz ich jest w zasadzie wielką komorą fermentacyjną (u krowy o objętości od 80 do 150 dm3). Pasza pozostaje organizmie przez około 9-12 godzin. W tym okresie następuje rozkład celulozy, głównie przez celulolityczne bakterie i orzęski. Uwolniona glukoza podlega fermentacji z wytworzeniem lotnych kwasów tłuszczowych, głównie octowego, propionowego i masłowego oraz gazów takich jak dwutlenek węgla i metan. Powstające kwasy stanowią podstawowe źródło energii dla zwierząt przeżuwających. Poza funkcją trawienną mikroorganizmy przewodu pokarmowego syntetyzują również aminokwasy i witaminy. Najaktywniej celulozę rozkładają bakterie z rodzaju Ruminococcus i Fihrobacter. Proces fennentacji metanogennej prowadzą bakterie z rodzaju Methanobrevibacter i Methanomierobium.
Mikroflora jelitowa człowieka stanowi również bardzo złożony ekosystem, w skład którego wchodzi do około 500 różnych gatunków mikroorganizmów. Mikroorganizmy te zasiedlają blisko 400 m2 powierzchni nabłonka jelitowego, są integralną częścią zawartości światła jelita, jak i trwale są związane ze śluzówką, tworząc specyficzne nisze ekologiczne. Najmniejsza ilość drobnoustrojów występuje w żołądku, zależnie od poziomu pH.
U człowieka tę mikroflorę stanowią głównie gramdodatnie względnie beztlenowe bakterie. W pierwszym odcinku jelita cienkiego ilość drobnoustrojów jest w dalszym ciągu niska i dopiero w odcinku dwunastnicy zwiększa się zarówno ich ilość, jak i różnorodność. Poza gramdodatnimi bakteriami pojawiają się względnie beztlenowe gramujemne bakterie z rodziny Enterobacteriaceae. W jelicie grubym ilość bakterii silnie rośnie i osiąga poziom 1010-1011 komórek/g treści. Wśród tych mikroorganizmów, dominują gramujemne bakterie z rodzaju Bacteroides (ok. 30% całej populacji bakterii jelitowych) oraz gramdodatnie z rodzajów Eubacterium, Bifidobacterium, Ruminococcus i różne gatunki Clostridium, są to gatunki bezwzględnie beztlenowe, bardzo wrażliwe na kontakt z tlenem. Bakterii względnie beztlenowych jest ok. 100 - 1000 razy mniej i znajdziemy wśród nich głównie bakterie z rodzaju Lactobacillus. Tak liczny i różnorodny zespół mikroorganizmów prezentuje ogromny potencjał katalityczny w organizmie człowieka, którego aktywność jest niezmiernie istotna dla jego zdrowia.
Korzystna funkcja obecnych w jelicie mikroorganizmów polega na syntezie witamin, głównie z grupy B, współuczestniczeniu w trawieniu składników pokarmowych oraz na ochronie człowieka przed nadmiernym rozwojem patogenów jelitowych.
Innym przykładem korzystnego oddziaływania pomiędzy mikroorganizmami i zwierzętami może być przypadek związku kałamarnicy Eupryma scolopes z bakterią gatunku Vibrio fischeri. Kałamarnica żeruje nocą i mogłaby łatwo paśc łupem drapieżników gdyby nie strategia polegająca na posiadaniu tak zwanego organu świetlnego, działającego jak rodzaj latarki.Narząd ten działa podobnie do oka, jest wyposażony w torebkę z atramentem, odbijającą światło wytworzone w narządzie. Astruga światła pada na zewnątrz ku dołowi. Kałamarnica wytwarza słabe swiatło bardzo przypominające to, które dociera pod wodę z wieczornego nieba, powoduje to niemal całkowite wtopienie się kałamarnicy w otoczenie. Źródłem światła u kałamarnicy jest luminescencyjna bakteria Vibrio fischeri. Mikroorganizm ten ma zdolność do przekształcania energii chemicznej w świetlną (odwrotnie do fotosyntezy) poprzez specjalny kompleks enzymatyczny zwany lucyferazą.
Reakcja odpowiedzialna za bioluminescencję
Bakteria uzyskuje od kałamarnicy aminokwasy i inne związki organiczne, a prawdopodobie zwierzę dostarcza także bakterii tlen cząsteczkowy. Najciekawszym w tej zależności jest fakt, iż kałamarnia codziennie rano usuwa mikroorganizm z narządu aby wieczorem znów pozwolić na jej osiedlenie.
Isnieje bardzo wiele złożonych lub prostych układów, gdzie mikrooragnizm i zwierzę pozostają w tak zwanej nierównowadze, czy też w przypadku gdy jedno z patrnerów ma niekorzystny wpływ na drugie. Mamy tutaj głównie do czynienia z infekcjami lub intotoksykacjami. Z punktu widzenia ekologii biochemicznej będą nas interesowały intotoksykacje ponieważ w ich przypadku mamy do czynienia z niekorzystnym wpływem toksyn wytwarzanych przez niektóre drobnoustroje na organizm atakowany.
Jak już wspomniałam w rozdziale poświęconym oddziaływaniom pomiędzy mikroorganizmami istnieje wiele gatunków grzybów strzępkowych mających zdolność do syntezy mikotoksyn. Jest to grupa metabolitów wtórnych mających silne działanie toksyczne, w szczególności mutagenne i teratogenne. Do mikotoksyn stwarzających największe zagrożenie zdrowotne dla zwierząt i człowieka należą: aflatoksyny, ochratoksyny, trichoteceny, patulina oraz fumonizyny, zearalenon i toksyna T-2.
Aflatoksyny są syntetyzowane głównie przez niektóre szczepy Aspergillus flavus, większość, o ile nie wszystkie, szczepy Aspergillus parasiticus oraz spokrewnione gatunki Aspergillus nomius, a także przez niezwykle popularnego grzyba Aspergillus niger. Cztery główne toksyny podzielono na dwie grupy, B i G, na podstawie koloru (niebieski, zielony) na jaki fluoryzują w świetle ultrafioletowym po adsorpcji na substracie w fazie stałej. Są to aflatoksyny B1, B2, G1 i G2.W produktach roślinnych najczęściej i w największych ilościach znajdują się metabolity B1 i G1. Aspergillus parasiticus jest dobrze przystosowany do życia w środowisku gleby, natomiast Aspergillus flavus do aktywnego rozwoju na częściach nadziemnych roślin (takich jak kwiaty i liście) i przeważa na kukurydzy, nasionach bawełny i orzechach drzew. Biochemiczna różnica pomiędzy tymi gatunkami polega na tym, że Aspergillus parasiticus może wytwarzać wszystkie cztery aflatoksyny (B1, B2, G1 i G2), natomiast Aspergillus flavus zwykle tylko B1 i B2.
Rodzina aflatoksyn:
Aflatoksyny są zaliczane go substancji kancerogennych i rakotwórczych. Rozróżnia się dwa rodzaje toksyczności aflatoksyn (tak jak u wszystkich mikotoksyn): może ona mieć ostre działanie toksyczne i działanie rakotwórcze. Już małe dawki mogą być śmiertelne dla zwierząt. Najczęściej spotykanymi zatruciami aflatoksynami są: uszkodzenia wątroby, zniszczenie komórek parenchymy i poliferacje przewodów żółciowych. Występuja także krwawienia i zaburzenia funkcji nerwowych, związanych ze skurczami, porażeniami i zaburzeniami równowagi. U młodych zwierząt występuje zahamowanie wzrostu i ubytki na wadze. Obok ostrego zatrucia aflatoksyny mają zdolność do wywoływania nowotworów. Występuje ona wówczas, gdy toksyna przyjmowana jest z żywnością przez długi czas. Toksyna atakuje głównie wątrobę, żołądek a także wywołuje metastazy w płucach i nerkach. U ludzi występowanie ostrych ztruć aflatoksynami jest bardzo małe, ponieważ z reguły nie dochodzi do spożycia pokarmów spleśniałych i zawierających duże dawki toksyn, natomiast o wiele częściej zdarzaja się przypadki chronicznego zatrucia, prowadzącego do marskości i raka wątroby, dotyczy to głównie grup ludności żyjących w klimacie tropikalnym. Największa aktywność kancerogenną wykazują aflatoksyna B1 i jej metabolity.
Innymi groźnymi mikotoksynami są ochratoksyny, jest to metabolit wtórny grzybów strzępkowych wystepujących w klimacie umiarkowanym i chłodnym: Penicillium viridicatum, Penicillium cyclopium, Penicillium verrucosum oraz Penicillium palitans, oraz w klimatach cieplejszych: tropikalnym i subtropikalnym Aspergillus ochraceus, Aspergillus melleus, Aspergillus sulphureus. Ochratoksyny A i B są substancjami silnie rakotwórczymi, powodują uszkodzenia nerek i ogólnie osłabiają system immunologiczny organizmu.
Rodzina ochratoksyn:
U podstaw toksycznego i rakotwórczego działania aflatoksyny B1 i ochratoksyny A i B leży ich zdolność kowalencyjnego wiązania DNA i tworzenie adduktów. Związki te działają hepato- i nefrotoksycznie, czyli wywołują raka wątroby i nerek.
Inną mikotoksyną jest patulina produkowana przez grzyby z rodzaju Penicillium takie jak : Penicillium expansum, Penicillium urticae, Penicillium claviformae i Penicillium vividicatum, a także gatunki Aspergillus elevatus, Aspergillus giganteus, Aspergillus terreus oraz Byssochlamys nivea i Byssochlamys fulva. Patulina działa ogólnotoksycznie na komórki ssaków, jest czynnikiem rakotwórczym, cytotoksycznym i teratogennym, powoduje wysoką umieralność płodów oraz nieodwracalne wady rozwoju zarodków.
Patulina
Poznano równiez inne mikotoksyny takie jak trichoteceny, zearalenon, nazywany też toksyną F-2 i fumonizyny. Cżęsto mikotoksyny są przyczyną ogólnego zmęczenia i niesprawności organizmu, szczególnie widoczne jest to u ludzi, którzy długi czas przebywają w zagrzybionych pomieszczeniach.
Również w przypadku chorobotwórczych bakterii mamy często do czynienia nie tylko z infekcją ale też intotoksykacją, ponieważ patogeneza chorób bakteryjnych zasadza się na dwóch podstawowych mechanizmach. Pierwszy mechanizm związany jest z bezpośrednim atakowaniem przez bakterie komórek i tkanek infekowanego organizmu, drugi zaś z wytwarzaniem toksyn bakteryjnych. Toksyny te są niejednokrotnie bardziej niebezpieczne dla organizmu atakowanego niż sam rozwój drobnoustrojów. Przykładem takiej reakcji jest botulizm, wywoływany przez tak zwany jad kiełbasiany (produkowany przez beztlenową laseczkę Clostridium botulinum, występuje 7 typów laseczek jadu kiełbasianego (od A do G), zróżnicowanych według rodzajów wytwarzanych toksyn. Chorobotwórcze dla człowieka są cztery typy (A, B, E i F). Przetrwalniki jadu kiełbasianego są odporne na działanie wysokich temperatur i wytrzymują gotowanie. Botulizm jest następstwem spożycia zakażonej żywności. Toksyna jest czuła na temperaturę i rozkłada się w czasie gotowania. Zdarza się jednak, że w skutek wytworzenia się ochronnej warstwy z białka lub innych składników żywności toksyna pozostaje nie rozłożona).Toksyna botulinowa jest niezwykle silna i śmiercionośnś trucizną juz w małych ilościach (dawka śmiertelna dla człowieka to 0,0004mg). Egzotoksyna botulinowa jest białkiem złożonym z dwóch połączonych mostkiem dwusiarczkowym łańcuchów polipetydowych: ciężkiego (100 kDa) i lekkiego (50 kDa), aby nabyć swoje wiruletne właściwości neurotoksyna uwolniona przez bakterie w nieaktywnej postaci, musi ulec proteolizie, którą katalizują albo bakteryjne proteazy albo enzymy proteolityczne wydzielane przez żołądek. Zabsorbowana przez śluzówkę botulina przenika do krwiobiegu i wraz z krwią dostaje się do obwodowych zakończeń nerwowych.
Tam blokuje przepływ sygnałów w cholinergicznych synapsach nerowo-mięśniowych (w których przekaźnikiem jest acetylocholina). Łańcuch ciężki toksyny odpowiedzialny jest za swoiste związanie się z błoną neuronów motorycznych i za wprowadzenie łańcucha lekkiego do wnętrza neuronu na drodze endocytozy. Łańcuch lekki posiada aktywność enzymatyczną. Jest zależną od cynku endopeptydazą, która trawi białka SNARE, odpowiedzialne za fuzję pęcherzyka synaptycznego wypełnionego neuromediatorem z błoną neuronu. Nie może dojść wówczas do uwolnienia acetylocholiny do szczeliny synaptycznej i skurczu włókien mięśniowych. W konsekwencji prowadzi to do porażenia i zwiotczenia mięśni. Śmierć następuje zwykle w wyniku paraliżu mięśni oddechowych i zatrzymania akcji serca.
Interakcje w królestwie zwierząt
Pomiędzy zwierzętami dochodzi do interakcji biochemiczno-ekologicznych najcześciej w układach dotyczących drapieżnictwa. Związki chemiczne odgrywaja tutaj ogromną rolę, w tym przypadku nazywa się je allomonami i są powszechnie używane zarówno przez drapieżnika jak i ofiarę.
Drapieżnik substancje toksyczne wydziela w celu unieruchomienia bądź zabicia upolowanej zdobyczy. Niekiedy jednak używa ich także w obronie własnej gdy sam staje się potencjalną ofiarą dla innego drapieżnika.
Producentami allononów są zarówno bezkręgowce jak i kręgowce, a więc ryby, płazy, gady, ssaki. Liczba tych toksyn jest bardzo duża i są one zróżnicowane zarówno pod względen budowy jak i działania. Czasem związki obrony chemicznej mogą mieć właściwości feromonów i w tych przypadkach można mówić o tak zwanych „feromonach obronnych”. Są to najczęściej substancje antyafrodyzjakalne, przenoszone z samca na samicę na przykład u motyla Heliconius erato. Mają one na celu odstraszać zarówno inne samce jak i drapieżniki. Niektóre wydzieliny obronne ofiar sa wyrzucane ze znaczną siłą w stronę napastnika. Przykładem takiego działania może być mechanizm obronny u chrząszczy z rodzaju Brachynus, polega on na wyrzuceniu gorącej chmury toksyn w kierunku napastnika, temperatura chmury może dochodzić do 100C. Zachodzi tutaj reakcja silnie egzotermiczna: hydrochinon, który jest wytwarzany w gruczołach chrząszcza w monemcie wyrzucania ulega w powietrzu utlenieniu do benzochinonu , będącego główną substancją obronną. Pary benzochinonu są substancją drażniącą uszkadzającą głównie oczy napastnika. Reakcja zachodzi z silnym odgłosem przypominającym wystrzał.
Reakcja obronna chrząszcza Brachynus:
Niektóre wydzieliny gruczołów magą wyciekać ze zwierzęcia, bądź potencjalna ofiara opryskuje nimi przeciwnika.
Wśród allomonów kręgowców bardzo interesujące są toksyny węży.
Są to związki bardzo złożone, często mają charakter neurotoksyn, czyli działają na układ nerwowy. Gady te wykorzystują toksyny najczęściej w czasie ataku na ofiarę, mają one na celu głównie sparaliżowanie układu nerwowego zdobyczy a przez to jej unieruchomiene. Często zdarza się, że węże połykają jeszcze żywe ofiary. W składzie toksyn węży wykryto złożone komleksy substancji czynnych bilogicznie: toksyny pre-i postsynaptyczne, kardio i miotoksyny, toksyny krwiotoczne i enzymy proteolityczne. Najwięcej toksyn jednak wytwarzają lub kumulują z pokarmu stawonogi (w tym większą część owady).
Wśród tych związków znajdują się terpenoidy, przykładem może być tutaj gąsiennica borecznika rudego , która w stanie zagrożenia wydziela oleistą substancję, która jest przez gąsiennicę wydzielana i gromadzona z pokarmu.
5. Biochemiczno-ekologiczne oddziaływania wewnątrz gatunku
Omówiłam już wzajemne oddziaływania występujące w ekosystemach pomiędzy różnymi gatunkami. W tym rozdziale zajmę się zależnościami biochemiczno-ekologicznymi pomiędzy osobnikami tego samego gatunku. Omówię jakie związki są głównymi nośnikami informacji na przykład w stadach czy zgrupowaniach zwierząt. Wśród roślin porozumiewanie się w obrębie tego samego gatunku nie jest do końca poznane, można tu jedynie przytoczyć przykład bawełny, która zaatakowana przez pędziorka wysyła substancje lotne mające na celu zaalarmowanie sąsiadujących , jeszcze zdrowych roąlin o niebezpieczeństwie.
W świecie zwierząt, a także wśród niektórych glonów i grzybów oddziaływania wewnatrzgatunkowe są uwarunkowane wydzielaniem feromonów. Związki te pełnią różne funkcje, ale przede wszystkim są sygnałem, informacją dla innych osobników.
Feromony glonów i grzybów
W cyklu życowym glonów i grzybów dużą rolę odgrywa zlewanie się dwóch haploidalnych komórek. Gamety niektórych grzybów wydzielają więc feromony płciowe, będące związkami przyciągającymi gamety męskie. Feromony działające w ten sposób nazywamy atraktantami. Znaleziono je u niektórych grzybów (m.in. z rodzaju Rhodosporidium i Mucor), drożdży (m.in. Saccharomyces cerevisiae) i wielu glonów (zarówno u brunatnic, jak i zielenic). Niektóre z atraktantów płciowych glonów i grzybów przedstawiam poniżej.
Inny rodzaj oddziaływań wewnątrzgatunkowych wywołują autoinhibitory (autotoksyny), które wydzielane do środowiska bytowania glonów, hamują szybkość wzrostu ich komórek, nie dopuszczając do nadmiernego rozmnożenia się populacji. Można traktować ten przypadek, jako autoregulację populacji. Dzięki temu nie ulegają wyczerpaniu zasoby tlenowe lub pokarmowe danego siedliska bytowania żywych organizmów, co mogłoby mieć katastrofalne skutki.
Feromony zwierząt
Feromony pełnią funkcję chemicznych mediatorów zachowań zwierząt tego samego gatunku. Dzielimy je w zależności od wywoływanych efektów na feromony primerowe i uwalniające. Te pierwsze są znacznie słabiej poznane. Pełnią one głównie rolę regulatorów stosunków społecznych u zwierząt socjalnych, wywołując długotrwałe efekty fizjologiczne, które mogą trwać nawet po ustaniu wydzielania feromonu. Przykładem takich feromonów są te, które przyspieszają dojrzewanie płciowe samców lub samic. Feromony uwalniające powodują natychmiastową i gwałtowną reakcję odbierającego je organizmu, wywołując odpowiednią reakcję. I tak wydzielane w odpowiednim czasie związki przywabiają osobnika przeciwnej płci (atraktanty płciowe), informują o ataku przeciwnika, mobilizując do walki lub ucieczki (feromony alarmowe), czy pokazują drogę do gniazda (feromony szlakowe).
Interesującym zjawiskiem mającym miejsce w świecie owadów jest to, że oddzielona od roju pszczoła przywabiana jest przez pojedyńcze żądło innej pszczoły. Zamknięta w pułapce samica Biston betularia przywabia żyjące na wolności samce tego owada. Podobne efekty obserwowane w przypadku innych owadów, w zjawisku przywabiania samców z rodziny Lepidoptera przez samice tych samych gatunków znaczącą rolę odgrywają związki chemiczne. Obserwacje te pomogły w odkryciu pierwszego feromonu płciowego - był to bombykol, atraktant płciowy produkowany przez samice jedwabnika (Bombyx mori). Bombykol przywabia samce jedwabnika w niezwykle niskich stężęniach, Jedwabnik rozpoznaje go za pomocą receptorów umiejscowionych w każdym z 17.000 organów olfakcyjnych (sensilla triodea) ulokowanych w czułkach. Tak więc samiec tego owada rozpoznaje obecność samicy, która może znajdować się w odległości kilkuset metrów od samca. Role kompletnego feromonu wabiącego jedwabnika jest jednak mieszanina dwóch związków - bombykolu i bombykalu, przy czym ten drugi produkowany jest przez samice w znacznie mniejszych ilościach. Feromony są zazwyczaj mieszaninami wielu związków i pojedyńcze składniki tych mieszanin wykazują słabą aktywność. Biorąc pod uwagę fakt, że ponad 4500 gatunków błonkoskrzydłych, żyjących w Europie Środkowej, używa tylko około 150 związków chemicznych jako feromonów płciowych, konieczność używania mieszanin staje się w pełni zrozumiała. Generalnie o feromonach owadów wiadomo dziś znacznie więcej niż o feromonach innych zwierząt.
Feromony primerowe
Nawet w przypadku tak dokładnie poznanego zwierzęcia jakim jest pszczoła miodna (Apis mellifera), nie do końca poznane są mechanizmy regulacji feromonalnej. Wynika to z wielu powodów, a jednym z nich jest fakt, że w ekstraktach gruczołów wydalniczych pszczoły zidentyfikowano ponad 100 prostych związków organicznych. Funkcję feromonów pełni zaś zaledwie kilka z nich. Jednym z najciekawszych zjawisk w biologii owadów socjalnych jest kastowość. W żyjącej w ulu społeczności pszczół miodnych mamy do czynienia z dwiema kastami: królową (która jest jedna) i robotnicami (których zazwyczaj jest od 15.000 do 60.0000 w jednym ulu). Głównym zadaniem królowej, która w ciągu swego życia składa miliony jaj, jest reprodukcja. Robotnice zdobywają pożywienie, karmią larwy i chronią ul. Właściwe funkcjonowanie tej społeczności jest regulowane przez feromony. Poznano co najmniej 15 gruczołów pszczelich wydzielających różnorakie feromony. Feromony wydzielane przez królową wpływają w sposób niezwykle istotny na funkcjonowanie ula, gdyż powodują one spójność stada i regulują wszystkie aspekty aktywności życiowej ula, a co ważniejsze powodują degenerację płciową robotnic. W okresie rojenia w ulu znajduje się kilka królowych - są to nowe królowe. Obecność więcej niż jednej w roju czy ulu powoduje, że toczą one ze sobą walki i tylko jedna zostaje przy życiu. Jeśli zostanie ona zapłodniona przez trutnia (zazwyczaj w 10-15 dniu po narodzinach), zostaje królową do końca swego życia. O dominacji królowej w ulu decyduje tak zwana substancja królewska, wydzielina jej gruczołów. Jest to feromon primerowy, który powoduje, między innymi, degenerację płciową robotnic. Robotnice są bowiem zdolne znosić jaja i niekiedy to czynią. Jaja te są jednak pożerane przez pielęgniarki, gdyż nie są oznakowane substancją królewską (jest to druga primerowa funkcja tej substancji). Głównym składnikiem substacji królewskiej jest kwas 9-okso-(E)-2-decenowy. To on powoduje zahamowanie wykluwania się nowych królowych. Substancja królewska jest jednak mieszaniną co najmniej pięciu związków o skomplikowanym sposobie działania.
Kwas 9-okso-(E)-2-decenowy pełni także inną rolę - jest on feromonem uwalniąjącym, atraktantem płciowym, gdyż przywabia trutnie do pszczoły w trakcie jej lotu godowego. Jest on jedynym feromonem, którego receptory bez wątpliwości zidentyfikowano w czułkach trutni. Gruczoły robotnic nie produkują substancji królewskiej. Robotnice wydzielają podobne dziesięciowęglowe kwasy (substancja królewska to trzy diewięciowęglowe kwasy) o nieznanej dotychczas funkcji biologicznej. Takie same kwasy i w podobnym stężeniu wydzielane są również przez gruczoły królowej.
Związki występujące w substancji królewskiej pszczoły miodnej:
Feromony primerowe, podobne do feromonów pszczoły, odgrywają najprawdopodobniej kluczową rolę w regulacji stosunków społecznych u większości owadów socjalnych. Niestety ich izolacja i zdefiniowanie struktury chemicznej nastręcza o wiele więcej trudności niż ma to miejsce w przypadku feromonów uwalniających. Trzmiel (Bombus terrestris) to jeden z najprymitywniejszych owadów socjalnych. W przeciwieństwie do pszczół rozwój kolonii trzmiela nie przebiega wcale harmonijnie. Królowa po zimowej hibernacji zakłada gniazdo i składa w nim jaja. Wykluwa się z nich 5 - 20 robotnic, które rozbudowują gniazdo. Pierwszy etap rozwoju to bezkonfliktowe współżycie królowej i robotnic. Królowa składa jaja, a robotnice dbają o gniazdo. Gdy populacja robotnic osiąga stan krytyczny kolonia osiąga następną fazę rozwoju. Robotnice przestają być zdegenerowane płciowo i, mimo obecności królowej, składają haploidalne (niezapłodnione) jaja, z których wylęgają się samce. Co więcej, robotnice stają się agresywne w stosunku do królowej i niekiedy nawet wypędzają ją z gniazda. Powszechnie uważa się, że ten drugi etap rozwoju związany jest z powolnym zanikiem produkcji feromonu królewskiego przez królową. Feromon ten jest złożoną mieszaniną wielu substancji chemicznych. Analiza wydzielin gruczołów trzmiela wykazała, że składa się ona z ponad 500 różnorakich związków chemicznych różnego typu, z czego jedna trzecia to węglowodory, a jedna trzecia to estry kwasów tłuszczowych. Niestety, jak dotychczas nie udało się stwierdzić, które z nich mogą wchodzić w skład substancji królewskiej trzmiela.. Ekstrakty otrzymane z kilkunastu gatunków trzmieli różnymi rozpuszczalnikami mają bardzo podobny skład i zawierają po kilkaset prostych zwiazków chemicznych. Podział funkcji spłecznych osiąga niezwykły poziom skomplikowania w społeczności mrówek. Przykładem mogą być tu mrówki z rodziny Atta, które zbierają świeże liście i hodują na nich grzyby. W kaście robotnic można wyróżnić mrówki wyspecjalizowane w hodowli grzyba, przetwarzaniu liści oraz te poszukujące liście. Także w przypadku mrówek uważa się, że degenaracja płciowa robotnic jest wynikiem działania feromonów primerowych. Wiele gatunków mrówek charakteryzuje się harmonijną koegzystencją kilku królowych, które wzajemnie ignorują swoją obecność. W koloniach palearktycznych mrówek Leptothorax gredleri, zaraz po okresie hibernacji, królowe ustalają swoją pozycję socjalną, walcząc z innymi. Wpływa to na funkcjonowanie gniazda i często zdarza się, że przedłużające się walki między królowymi są przyczyną ucieczki robotnic wraz z larwami z gniazda. W przypadku nieefektywnego działania feromonów królewskich zdarza się, że i świat robotnic ma budowę hierarchiczną, a wysoko ulokowane w hierarchi robotnice mogą składać jaja w obecności królowej. Takie przypadki zaobserwowano wśród mrówek z rodziny Leptothorax.
Feromony szlakowe mrówek Atta:
Ślepe mrówki Aenictus nie żyją w gniazdach, lecz przenoszą się z miejsca na miejsce, organizując biwaki (tymczasowe gniazda). Z tych miejsc postoju organizują one zmasowane rajdy w poszukiwaniu pożywienia. Jedna kolumna mrówek liczy zazwyczaj kilkadziesiąt tysięcy robotnic. Oczywistym jest, że muszą one znakować chemicznie drogę w trakcie rajdu. W tym celu używają feromonu szlakowego, którym jest antranilan metylu. Okazuje się, że prócz feromonu szlakowego (typowego feromonu uwalniającego) mrówki te produkują również feromon primerowy, który powoduje mobilizację robotnic i ich grupowanie się w kolumny rajdowe. Jest nim nikotynian metylu, wydzielany przez gruczoły mrówek w bardzo niewielkiej ilości. Związek ten powoduje, że mrówki poruszają się po ścieżkach utworzonych przez feromon szlakowy jescze w kilka godzin po jego zniknięciu ze środowiska.
Feromony uwalniające
Feromony uwalniające używane są przez wszystkie organizmy jako przekaźniki informacji przeznaczone dla osobników tego samego gatunku. Informacja niesiona przez feromon uwalniający powoduje natychmiastową reakcję odbierającego ją organizmu. Zatem kontrolują one zarówno ich rozwój, jak i zachowanie się. Najlepiej poznaną grupą tych feromonów są atraktanty płciowe, a mniej poznane są feromony agregacyjne, związki znakujące szlak czy terytorium, sygnały alarmowe, deterenty owipozycji (związki powodujące zaniechanie składania jaj przez owady), czy też substancje powodujące uwalnianie gamet. Większość z tych feromonów to skomplikowane mieszaniny związków chemicznych o różnorakiej budowie. Stężenie tych związków w środowisku jest bardzo niskie. Łączenie się zwierząt w pary poprzedzone jest zazwyczaj drastycznymi zmianami zachowania i fizjologii. Jednym ze sposobów znajdowania partnera jest produkowanie feromonów - atraktantów płciowych. U owadów sygnalizacja chemiczna jest najważniejszym składnikiem zachowań godowych. Jest tak dlatego, że sygnał chemiczny dociera do odbiorcy bez względu na oświetlenie (także i w nocy) i konfugurację terenu. Energetyczny wysiłek niezbędny dla biosyntezy i emisji feromonów jest niewielki. Niedogodnością jest fakt, że sygnały te rozprzestrzeniane są w powietrzu statystycznie i owad nie ma wpływu na kierunek ich dyfuzji ani nie może szybko zamienić wysłanego już sygnału na inny (na przykład dać znać, że pojawił się wróg). Podobne zalety mają sygnały dźwiękowe i wiele gatunków owadów wabi partnera charakterystycznym „śpiewem”. Ćmy używają niemal wyłącznie sygnalizacji chemicznej i niemal wyłącznie samice emitują feromony płciowe. Najważniejszą cechą atraktantów płciowych jest to, że przywabiają one tylko i wyłącznie samce tego samego gatunku. Zapobiega to krzyżowaniu się owadów różnych, choć spokrewnionych gatunków. Dlatego też są to zazwyczaj mieszaniny związków o ściśle określonych składach ilościowych. Samice ciem produkują feromony płciowe wyłącznie w nocnym i to w ściśle określonych porach. Jest to ważne, ponieważ w większości przypadków ćmy żyją krótko. Samce zwykle zapładniają wiele samic. Charakterystyczną cechą samców tej ćmy są wielkie „owłosione” czułki (anteny) pozwalające na identyfikację niewielkich stężeń feromonów.
Feromonem płciowym Idea leucone jest danaidon. Związek ten znaleziono również w wydzielinach gruczołów innych motyli z tej rodziny. Ekstrakty tych gruczołów zawierają zazwyczaj do 60 składników chemicznych. Żaden ze zidentyfikowanych związków nie wykazywał jednak aktywności przywabiającej samce. Badania na feromonem Idea leucone - głównie metodą chromatografii gazowej i spektroskopii mas - pozwoliły znaleźć dwa następne składniki feromonu wabiącego tego motyla, a więc lepiej zrozumieć specyficznośc gatunkową tego feromonu.
Dobrym przykładem do badań jest praca opisująca próbę identyfikacji feromonu szlakowego mrówki Dolichoderus thoracicus. Ta azjatycka mrówka buduje gniazda w lasach pomiędzy liśćmi roślin. Mrówki te posiadają co najmniej sześć gruczołów wydzielniczych. Feromon szlakowy produkowany jest przez jeden z nich. Ekstrakt tego gruczołu to osiemnaście aldehydów, z których pięć wykazywało wysoką aktywność jako feromony szlakowe, jeden był średnio aktywny, zaś dwa wywoływały słabą odpowiedź. Pozostałe aldehydy nie są używane do znakowania szlaku. Z kolei korniki, które powodują poważne straty w drzewostanie lasów całego świata aby osiągnąć cel, jakim jest kolonizacja drzewa i pokonać bariery ochronne, jakie to drzewo stwarza, przeprowadzają zmasowany atak Zazwyczaj kolonizacji drzewa towarzyszy atak przyniesionych przez korniki grzybów. Hamują one biosyntezę żywicy - głównej substancji obronnej rośliny. Zatem efektywność ataku zależy wprost proporcjonalnie od liczby atakujących owadów. Rolę czynnika zwołującego owady pełni feromon agregacyjny składający się z terpenów produkowanych przez kornika i terpenów będących składnikami żywicy. Biosynteza feromonów agregacyjnych jest stymulowana obecnością składników żywicy. Większość korników kolonizuje różne gatunki drzew, a ich żywice zawierają różne składniki co stanowi, że system komunikacji chemicznej tych owadów jest wysoce skomplikowany. Warto zdać sobie sprawę, że atmosfera lasu jest pełna różnorodnych zapachów, a każdy centymetr kwadratowy powietrza zawiera miliony różnych substancji chemicznych. W tej sytuacji niezwykle zaskakującym jest, że ten sam gatunek kornika Ips pini ze stanu Idaho reaguje na działanie jednego izomeru ipsendolu, podczas gdy żuki ze stanu Nowy Jork reagują jedynie na działanie mieszaniny enancjomerów tego związku i to tylko wtedy, gdy jej skład jest ściśle określony.
Feromony produkowane są przez prawie wszystkie organizmy, od mikroorganizmów poczynając, na ludziach kończąc. Swoistą sensacją było odkrycie kilka lat temu feromonu płciowego słonia azjatyckiego. Samica słonia, kiedy jest gotowa do zapłodnienia, wydziela z moczem prosty ester nienasycony - octan (Z)-dodecen-7-ylu. Jest to o tyle zaskakujące, że to największe zwierzę lądowe używa jako feromonu płciowego tego samego związku, co niektóre świerszcze oraz ponad 100 gatunków motyli i ciem. Takie zjawisko nie jest jednak wyjątkowe w przyrodzie. Na przykład feromon płciowy wielu korników jest niezwykle podobny do feromonu agresji myszy. Myszy tworzą wysoce hierarchiczne struktury społeczne i pozycja samca jest wprost proporcjonalna do poziomu jego agresji. Feromon ten produkowany jest przez samce walczące o samice. Samce oddzielone od samic nie produkują tego feromonu i nie są aktywne płciowo. Innym przykładem może być substancja produkowana przez roślinę Nepeta cataria. Ekstrakt tej rośliny, a właściwie jego główny składnik - nepetalakton, powoduje dramatyczną zmianę zachowania kotów, tygrysów i lwów. Pod jego wpływem zwierzęta te zaczynają taniec godowy. Zaskakującym jest, że mieszanina nepatalaktonu i jego zredukowanej formy jest standardowym atraktantem płciowym mszycy Aphis fabae. Istnienie ludzkich atraktantów płciowych nie podlega dzisiaj dyskusji, intensywne badania sugerują, że feromonem płciowym mężczyzn jest mieszanina pochodnej androsteronu i prostego kwasu nienasyconego emitowana wraz z potem. To, że feromony te wydzielane są z potem, nie brzmi przyjemnie, a dodatkowo na ironię zakrawa fakt, że higiena osobista i stosowanie perfum (także i tych polecanych jako podnoszące atrakcyjność płciową mężczyzn) może usuwać naturalny atraktant płciowy z bukietu zapachowego człowieka. W niektórych kulturach panuje opinia, że stosowanie perfum zabija naturalny zapach i atrakcyjność skóry.
26
N2
N03-
N02-
NH4+
Aminokwasy
Białka
nitrogeneza
denitryfikacja
utlenienie
nitryfikacja
amonifikacja
aminacja
deaminacja
proteoliza
biosynteza
Wyszukiwarka
Podobne podstrony:
Podstawy Ekologii
Podstawy Ekologii, Studia, 1-stopień, inżynierka, Ochrona Środowiska, Ekologia
zagadnienia z podstaw ekologii HIL6M54BEDM2SJZDKLYP6QBMDXOM2MTNGL37INY
Podstawy ekologii cw. 1, Studia (Turystyka i Rekreacja), I semestr, Podstawy ekologii
Podstawy ekologii cw 4
Podstawy ekologii cw.6, Studia (Turystyka i Rekreacja), I semestr, Podstawy ekologii
Zarys Ekologii biochemicznej, Ekologia
Ekologia biochemiczna częściowe opracowanie na koło, Studia, Ochrona środowiska
jadczyk,ekologia, Podstawy ekologii krajobrazu
Różnorodność biologiczna, bio-chem-eko, Ekologia biochemiczna
1- Podstawy ekologii, Ekologia
Podstawy ekologii cw.2, Studia (Turystyka i Rekreacja), I semestr, Podstawy ekologii
Ekologia biochemiczna - Feromony, Ekologia
Podstawy ekologii cw.5, Studia (Turystyka i Rekreacja), I semestr, Podstawy ekologii
podstawy biotechnologii biochemia prezentacja
Podstawy ekologii, pedagogika społeczna
PODSTAWY?ZPIECZEŃSTWA EKOLOGICZNEGO wykłady
Podstawy Ekologii
więcej podobnych podstron