Ilościowa charakterystyka informacji:
Dla zbioru wszystkich występujących w układzie sygnałów można w umowny sposób określić ilość informacji przypadającej na poszczególny sygnał. Do chwili odebrania sygnału istnieje nieokreśloność, niepewność, który sygnał wystąpi. Odebranie określonego sygnału z danego zbioru usuwa niepewność a tym samym dostarcza pewnej informacji. Ilość informacji odpowiada zlikwidowanej niepewności. Największą ilość informacji otrzymamy, gdy istnieje największa niepewność przed jej uzyskaniem, a nie otrzymamy jej wcale gdy owej niepewności nie mamy. Im liczniejszy jest zbiór tym bardziej niepewny jest wybór z tego zbioru określonego sygnału. Za miarę niepewności wyboru danego sygnału przyjęto ujemny logarytm z prawdopodobieństwa wyboru sygnału:
I= -log p
Każdą dowolnie złożoną wiadomość, liczbę itp. można przedstawić za pomocą ciągu dwóch tylko różnych znaków, użycie kodu dwójkowego. Przy tym ilość informacji obliczamy ze wzoru:
I= -log2p
Taka ilość informacji wynosi 1 i stanowi dwójkową jednostkę informacji. Nazwano ją bit, nadaje się do określania ilości dowolnej informacji. Z każdej możliwej liczby zdarzeń można przeprowadzić wybór dowolnego zdarzenia drogą wykluczeń przez podziały dwójkowe.
Przy zdarzeniach niejednakowo prawdopodobnych ilość informacji przypadających na sygnał możemy obliczyć ze wzoru Shannona:
I= - p1log2p1 - p2log2p2 …..-pnlog2pn
Informacja a entropia (demon Maxwella)
Entropia jest miarą nieuporządkowania układu, a informacja jest miarą nieokreśloności. Przyjęto średnią ilość informacji I nazwać entropią H zbioru zdarzeń, czyli I=H, wtedy wzorem na entalpię H jest poniższy wzór:
H= - p1log2p1 - p2log2p2 …..-pnlog2pn
Gdy zjawisko jest pewne(jedna możliwość) to entropia ma wartość zerową. Natomiast ma wartość maksymalną gdy zjawisko jest najbardziej niepewne, tj. gdy prawdopodobieństwo poszczególnych zdarzeń są sobie równe.
Podstawowe pojęcia z teorii informacji
Kod to odpowiedniość polegająca na tym, ze każdemu elementowi jednego zbioru odpowiada element zbioru drugiego. Celem kodowania jest takie przystosowanie nadawanego ze źródła sygnału, aby mógł być zrozumiały przez odbiorcę. Kod może być zupełnie dowolny np. języki, układy cyfr, lub niezmienny- kod genetyczny, natomiast sama informacja pozostaje niezmienna. Kod to zbiór m wyrazów, który tworzy pewną wiadomość. Wiadomość ta może zostać przesłana przez n sygnałów (sygnał - proces fizyczny stanowiący materialny nośnik informacji np. dźwięk, litera, liczba). Wyróżniamy kody ziarniste (np. cyfrowe lub literowe) lub ciągłe (poprzez zmiany natężenia dźwięku, światła).
A także:
Kod amplitud - amplitudy iglic powstałego potencjału w czasie zależą proporcjonalnie od siły bodźca zewnętrznego - im silniejszy bodziec tym wyższa amplituda iglicy (ogromna redundancja, podatność na błędy, nie występuje u organizmów żywych).
Kod dwójkowy - amplitudy iglic powstałego potencjału są stałe, jednak ilość iglic w czasie zależy od rodzaju bodźca zewnętrznego (wytłumienie lub nadmiar iglic przynosi ten sam efekt, nie występuje u organizmów żywych).
Kod przedziałów - amplitudy iglic powstałego potencjału są stałe, jednak odstępy między iglicami w czasie zależą od siły bodźca zewnętrznego - im słabszy bodziec tym większa odległość między iglicami ( nie występuje u organizmów żywych).
Kod częstości - amplitudy iglic powstałego potencjału są stałe jednak ich liczba w jednostce czasu zmienia się w zależności od siły bodźca zewnętrznego(najbardziej odporny na szumy, występuje u organizmów żywych)
Pojemność informacyjna układu
Graniczna ilość informacji jaką układ może zgromadzić, wprowadzić lub przekazać jeśli dysponuje „m” różnymi sposobami przy czym wiadomość składa się z „n” symboli, niekoniecznie różnych. Ilość różnych wiadomości układu oblicza/ się ze wzoru:
N=mⁿ
Miarę pojemności układu obliczmy ze wzoru:
log2N= Q=nlog2m
Rzeczywista miara ilości informacji zawartej w wiadomości uwzględnia osobliwości występujących symboli, bo większość kodów dopuszcza jedynie niektóre kombinacje.
Jednostką pojemności inf. układu, w którym wiadomość składa się z 2 różnych symboli [0,1] jest bit.
Pojemność informacyjna pamięci
Pojemność informacyjna pamięci wynosi około 10^20bitów
Nadmiar informacji (redundacja) -ilość informacji przekraczająca wymagane do rozwiązania problemu minimum
R= 1- Hrzecz/Hmaks
gdzie:
H rzecz- entropia rzeczywista
H maks- entropia maksymalna
Gdyby redundancja miała wartość zerową, oznaczałoby to, że moglibyśmy otrzymywać zrozumiały tekst przy dowolnym układzie liter, co jest niemożliwe. Zwiększeniu nadmiarowości informacji sprzyja stosowanie pewnej frazeologii lub żargonu zawodowego. Przykładami redundancji są historie chorób spisywanych przez lekarzy (więcej informacji, niż potrzeba do określenia diagnozy)
Entropia względna informacji inaczej dywergencja Kulbacka-Leiblera (odnosząca się do teorii i informacji i statystyki) jest to miara stosowana do określenia rozbieżności między dwoma rozkładami prawdopodobieństwa p i q . Czasami zwana jest też odległością Kulbacka-Leiblera, w rzeczywistości nie jest to prawdziwą metryką, gdyż nie jest symetryczna i nie spełnia nierówności trójkąta.
Dla rozkładów dyskretnych wyrażana jest wzorem:
dKL(p,q)= ƹp(i)log2p(i)/q(i)
Dla rozkładów ciągłych:
dKL(p,q)= ƪp(x)log2[p(x)/q(x)]*dx
Entalpia ta przyjmuje same wartości nieujemne, przy czym 0 tylko wtedy gdy porównywane rozkłady są identyczne
Scharakteryzuj schemat przesyłania informacji. Omów prędkość przepływu informacji.
Na początku dla odbiornika informacji, źródło informacji stanowi maksymalną nieokreśloność, niepewność. Źródło przekazuje pewne informacje zarówno werbalne jak i niewerbalne, które są zakodowane w pewien ogólnie przyjęty sposób-kod. Celem kodowania będzie jej transformacja, czyli przemiana na ciąg sygnałów zrozumiałych dla odbiorcy. Czasami spotykamy się z transformacją wielokrotną, np. przy wyrażaniu myśli w języku obcym. Między źródłem a odbiorcą wytwarza się kanał łączności, którą może być np. fala dźwiękowa, linia telefoniczna czy łącze radiowe. Do kanału łączności mogą się dostawać różnorodne szumy, które zakłócają przekazywane informacje. Na końcu kanału łączności znajduje się odbiornik sygnałów (np. człowiek), którego zadaniem jest wyodrębnienie spośród czynników zakłócających właściwych sygnałów, a następnie przeprowadzenie dekodowania, przy zastosowaniu transformacji odwrotnej. Na końcu informacja jest przekazywana do odbiornika informacji (np. przez narządy zmysłu do mózgu). Prowadzi to do powstawania sprzężnego między źródłem informacji a odbierającym informację.
Prędkość przepływu informacji.
Różnica między entropią bezwarunkową źródła H(X) a entropią warunkową H(X/Y), czyli entropią źródła wiadomości kiedy jest znany sygnał na wyjściu.
v= [H(X)- H(X/Y)]/t
Jednostką jest bit na sekundę bit/s
W warunkach dobrego odbioru H(X/Y) jest zbliżone do zera i wtedy prędkość przesyłania jest równa entropii źródła, co oznacza, że całkowita ilość dociera do odbiornika. Stwierdzono, że prędkość odbioru i przetwarzania informacji z otoczenia przez człowieka nie przekracza 25bitów na sekundę.
Prawa rządzące przepływem informacji (sterowanie a regulacja)
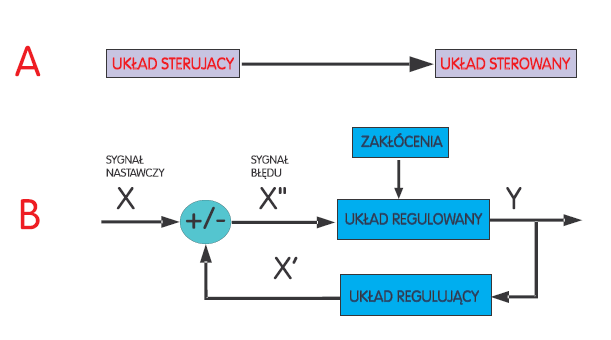
Pojęcie sterowania- kojarzy się z prowadzeniem łodzi, statku, samolotu lub z przykładem dotyczącym sterowania wojskiem, zakładem pracy czy grupą ludzi. Można wyróżnić dwa układy: sterowany i sterujący.
Proces sterowania można zrozumieć jako wywieranie przez układ sterujący działania na układ sterowany dla osiągnięcia zamierzonego celu. Sterowanie odbywa się nie za pomocą energii, lecz poprzez informację zawartą w sygnałach. Układ sterowania jest układem otwartym to znaczy takim, w którym skutek działania nie ma wpływu na przyczynę go wywołującą a wykonanie przez układ sterowany czynności nie jest kontrolowane przez układ sterujący.
Układy regulacji umożliwiają utrzymanie homeostazy w organizmie. Układy regulacyjne w organizmach żywych są przystosowane do przesyłania (lub odbierania) sygnałów tylko jednego znaku (od stanu zerowego do maksymalnego) i dlatego zawsze muszą w nich istnieć dwa typy antagonistycznych układów:
jeden reagujący na błędy dodatnie
drugi reagujący na sygnał błędów ujemnych
W układach regulujących stałość środowiska wewnętrznego tego rodzaju funkcje wykonuje wegetatywny układ nerwowy. Realizuje on czynności regulacyjne za pomocą dwóch rodzajów nerwów: układu współczulnego (pobudzającego) i przywspółczulnego.
UKŁAD REGULACJI
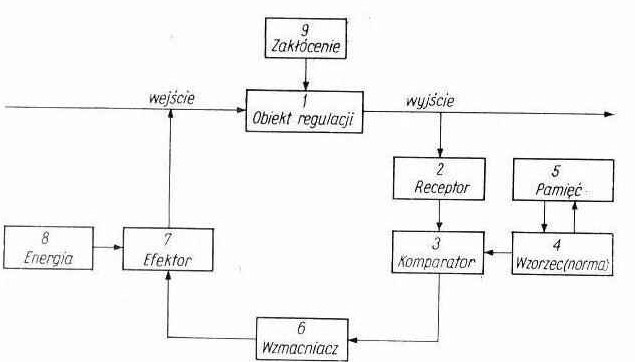
Sterowanie
Regulacja
Sposoby przetwarzania informacji w przyrodzie (podaj przykłady)
Przetwarzanie informacji można rozumieć przystosowanie do określonych potrzeb człowieka. Będzie to polegało na doprowadzeniu informacji do postaci ułatwiającej podjęcie decyzji.
Proces przetwarzania informacji zawartej w symptomach odbywa się w umyśle i dlatego nie jest wolny od znacznych nieraz zakłóceń (szumów), które wiążą się z czynnikami emocjonalnymi lub psychologicznymi niewystarczającym doświadczeniem i zakłócają proces właściwego przetwarzania informacji.
Rozważmy jeszcze kilka konkretnych przypadków skomplikowanego przetwarzania danych, z którymi spotykamy się w układach biologicznych.
Eferentny efekt ruchu oka.
Mimo że mięśnie naszego oka są stale w ruchu, a w związku z tym obraz powstały na siatkówce przesuwa się po niej bez przerwy, człowiek nie dostrzega tego przerwanego ruchu otoczenia. Można bardzo łatwo odróżnić nieruchome przedmioty od ruchomych. Ta cecha organizmu związana jest z określonym przetwarzaniem informacji aferentnych, to znaczy informacji dochodzących od oka do ośrodkowego układu nerwowego). Ruch oka wywołany jest przez polecenie eferentne wysłane z ośrodkowego układu nerwowego. Polecenie to dochodzi jednak nie tylko do odpowiednich mięśni, ale rozchodzi się jednocześnie po drogach aferentnych prowadzących z siatkówki do mózgu. W mózgu dokonuje się odejmowanie nakazanego kąta obrotu gałki oka od obserwowanego przez oko kąta przesunięcia obrazu na siatkówce. Jeśli obserwowany obiekt nie porusza się, to jego obraz na siatkówce musi przesunąć się o tyle, o ile gałka oczna przesunęła się na skutek polecenia okoruchowego. Wynik dokonanego w mózgu odejmowania obu kątów jest w tym przypadku równy zeru. Przy ruchu obiektu względem obserwatora różnica obu kątów jest rożna od zera, a wielkość tej różnicy zawiera w sobie pewną ilość informacji. Hipotezę tę łatwo można sprawdzić. Należy patrzeć na nieruchomy przedmiot jednym okiem i przesunąć gałkę oka przez lekki nacisk palcem. Ponieważ w tym przypadku nie dochodzi do mięśnia oka polecenie motoryczne, powstaje więc wrażenie ruchu przedmiotów nieruchomych znajdującym się w naszym otoczeniu.
Całkowanie przez mrówkę zmiennej wielkości mierzonej przy określaniu kierunku.
Mrówka poruszająca się przez pewien czas po drodze o zmiennym kierunku po osiągnięciu celu wędrówki może wrócić do swego mrowiska po drodze prostoliniowej i może powtórnie dotrzeć do tego. celu również po linii prostej łączącej cel wędrówki z mrowiskiem. Można wykazać doświadczalnie, że zwierzę kieruje się w swej wędrówce informacjami o położeniu łatwo dostrzegalnych/ optycznie punktów, a przede wszystkim informacjami położeniu słońca. W czasie akcji poszukiwania celu mrówka rejestruje bez przerwy wielkość kąta między kierunkiem ruchu i kierunkiem ku dobrze widocznym punktom. W ośrodkowym układzie nerwowym mrówki dokonuje się jednocześnie całkowanie tej ujemnej wielkości regulowanej. Wynik całkowania podzielony przez czas szukania celu daje dokładnie kąt zawarty między kierunkiem do celu i kierunkiem ku obranemu znakowi optycznemu. Załóżmy, że droga mrówki do celu składa się z prostoliniowych odcinków o tej samej długości, .ale o rożnych kierunkach. Kąt, jaki tworzy droga mrówki do celu z kierunkiem ku punktowi wybranemu przez mrówkę, jako punkt orientacyjny, równy jest średniej wartości sumy wszystkich kątów, jakie tworzą poszczególne odcinki drogi z kierunkiem ku punktowi wybranemu za znak optyczny. Jeśli odcinki drogi są nieskończenie małe, to operacja powyższa sprowadza się do całkowania. Ośrodkowy układ nerwowy mrówki musi więc być zdolny do wykonania tego skomplikowanego działania i musi przy tym uwzględnić jeszcze zmiany położenia słońca podczas ruchu mrówki, jeśli wybranym punktem orientacyjnym było słońce.
Sterowanie wzrostu grzyba Phycomyces przez światło.
Skomplikowane biologiczne układy regulacji i sterowania nie muszą być koniecznie związane z istnieniem wysoko rozwiniętego układu nerwowego. Badania przeprowadzone na grzybie Phycomyces wykazały, ze zmiana natężenia światła wywołuje bądź zwiększenie, bądź zmniejszenie prędkości wzrostu jego grzybni. Światło o stałym, obojętnie jakim natężeniu, powoduje średni wzrost grzyba około 3 mm na godzinę. Sterowanie procesem wzrostu grzyba wymaga skomplikowanego przetwarzania efektów pobudzającego działania światła. Przypuszczalny przebieg tego procesu pokazany jest w układzie blokowym. Sygnał receptora świetlnego dociera do obszaru jego działania bezpośrednio bądź pośrednio przez kanał, który opóźnia bieg sygnału o pewien stały czas (stała czasowa). Różnica między opóźnioną w czasie i aktualną wartością wielkości regulowanej jest miarą wielkości zmiany natężenia światła i można ją wykorzystać bezpośrednio jako wielkość sterującą wzrostem. Takiemu schematowi przebiegu procesu sterowania można podporządkować również odpowiedni model biochemiczny.
Te trzy przykłady procesów regulacji i sterowania, wybrane z obszernego-zbioru zbadanych dotychczas tego rodzaju procesów, powinny nas przekonać, że przetwarzanie danych w układach biologicznych może odbywać się w bardzo różnorodny sposób. Obok tego niezwykłego działania układu regulacji sterowania na stosunkowo niskim poziomie fizjologicznym istnieją procesy regulacji i sterowania w o wiele bardziej skomplikowanych zjawiskach związanych z psychologią
Z przykładów, które przytoczyliśmy dotychczas, wynika, że przetwarzanie danych odbywa się na każdym stopniu rozwoju układu biologicznego, a więc nie tylko w układach posiadających układ nerwowy.
Sprzężenie zwrotne (funkcje przenoszenia, układy oscylacyjne)
Sprzężenie zwrotne - oddziaływanie sygnałów stanu końcowego (wyjściowego) procesu (systemu, układu), na jego sygnały referencyjne (wejściowe). Polega na otrzymywaniu przez układ informacji o własnym działaniu (o wartości wyjściowej). Matematycznym, jednoznacznym opisem bloku gałęzi zwrotnej jest transmitancja. Informacja ta może być modyfikowana przez transmitancję bloku gałęzi zwrotnej.
Sprzężenie zwrotne ujemne stanowi fundamentalny mechanizm samoregulacyjny. Z cybernetycznego punktu widzenia ma ono za zadanie utrzymanie wartości jakiegoś parametru na zadanym poziomie. Zachodzi ono wtedy, gdy jakiekolwiek zaburzenia powodujące odchylenie wartości parametru od zadanej wartości w którąkolwiek stronę indukują działania prowadzące do zmiany wartości parametru w stronę przeciwną (stąd nazwa "ujemne"), a więc do niwelacji (kompensacji) efektu tego odchylenia.
Sprzężenie zwrotne dodatnie polega na tym, że w sytuacji zakłócenia jakiegoś parametru w układzie, układ ten dąży do zmiany wartości parametru w kierunku zgodnym (stąd - "dodatnie") z kierunkiem, w którym nastąpiło odchylenie od "zadanej" wartości.
Funkcje przenoszenia statyczne i dynamiczne
• Funkcja przenoszenia y = f(x) Funkcja przenoszenia o charakterystyce
statycznej dynamicznej-układ inercyjny I rzędu,
(1-e -(t/A)
• Wzmocnienie lub tłumienie zmiana w reakcji na bodziec = y = yo
układu: tgα=G
A = stała czasowa -określa czas narastania wielkości do wartości
yo = Gxo
Połączenie szeregowe dwóch układów daje układ inercyjny II rzędu, a n układów -n rzędu
STATYCZNE DYNAMICZNE
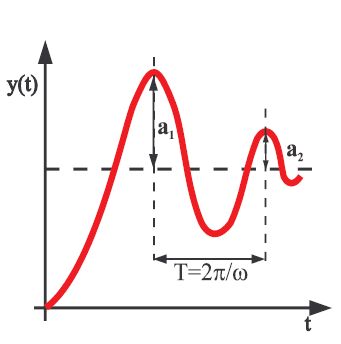
Układy oscylacyjne- w układach oscylacyjnych elementy nie tylko wykorzystują energie ale mogą też chwilowo ją magazynować jako energię potencjalną.
Układy takie oprócz współczynnika tłumienia także częstość oscylacji (
)
Poziom do którego parametr dąży.
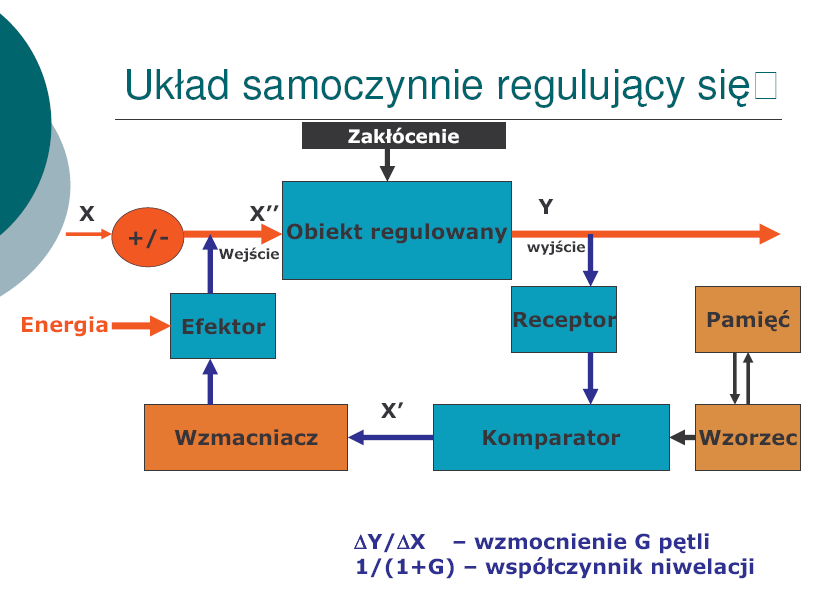
Schemat układu, w którym zachodzi autoregulacja przepływu informacji.
Autoregulacja- proces, który umożliwia utrzymanie stałości parametru wyjściowego niezależnie od działania czynników zewnętrznych. W układzie samoczynnej regulacji każde odchylenie kontrolowanego parametru od stanu równowagi, czyli tzw. sygnał błędu, stanowi informację odbieraną przez detektory (receptory), która wywołuje reakcje prowadzące poprzez odpowiednio działające człony do jego likwidacji.
Np. regulacja poziomu glukozy we krwi, autoregulacja ciśnienia tętniczego krwi
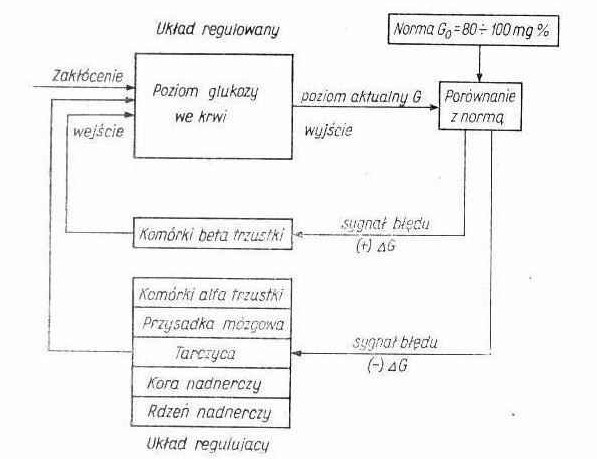
Scharakteryzuj homeostazę
Homeostaza- zdolność organizmu do utrzymania stałości środowiska wewnętrznego mimo zmian zachodzących w środowisku zewnętrznym. Organizm ludzki wymienia z otoczeniem materię i energię w sposób kontrolowany tak, aby zachować względną stałość środowiska wewnętrznego. Claude Bernard stwierdził, że zachowanie stałości środowiska wewnętrznego jest warunkiem swobodnego i niezależnego życia. Aby utrzymać stan homeostazy organizm wybiórczo reaguje na bodźce zewnętrzne - reakcja na wszystkie bodźce zewnętrzne prowadziłaby do zbyt intensywnego napływu informacji i zachwiania stanu równowagi (homeostazy).
Znaczenie polaryzacji wiązań kowalencyjnych dla właściwości fizykochemicznych wody
Wszystkie właściwości wody można wytłumaczyć dzięki jej polarnej strukturze.
Wraz ze wzrostem polaryzacji wiązań rośnie temperatura topnienia i wrzenia.
Oddziaływania elektrostatyczne pomiędzy cząsteczkami wody decydują o ich właściwościach
fizykochemicznych.
Woda jest dipolem. Między tlenem a wodorem cząsteczki sąsiedniej wytwarza się wiązanie wodorowe (mostek wodorowy), które prowadzi do powstawania asocjatów, którym woda zawdzięcza swoje właściwości. Z konieczności dostarczenia dodatkowej energii na rozerwanie wiązań wodorowych wynikają wysokie wartości parametrów fizykochemicznych:
Ctop = 333 kJ/kg
Cpar = 2260 kJ/kg
Cmol = 75 J/(mol*K)
Cwł = 4,18 kJ/(kg*K)
Przestrzenny rozkład chmury elektronowej cząsteczki wody prowadzi do tetraedrycznego rozkładu ładunku elektrycznego. Dzięki temu, cząsteczka może łączyć się z 4 kolejnymi cząsteczkami wody - tworząc heksagonalną strukturę lodu. Jest to struktura luźna, dużo w niej przestrzeni nie wypełnionej, dlatego lód jest kruchy, porowaty.
Inne właściwości wody:
a) gęstość - do 4 oC rośnie, później maleje
b) lepkość spada wraz ze wzrostem temperatury
c) przewodzenie dźwięku, ściśliwość - rosną wraz z temperaturę
Opisz znane Ci struktury uporządkowane ośrodka wodnego
Struktura tetraedryczna - każda cząsteczka otoczona jest przez cztery inne, zaangażowane w 4 wiązania wodorowe.
bardzo krucha
dynamiczna
atomy wypełniają 34% przestrzeni
proton wspólny dla 2 cząsteczek oscyluje
każda cząsteczka może być 2 razy donorem i 2 razy akceptorem
Struktura heksagonalna lodu
struktura luźna (66% przestrzeni niewypełnionej)
dynamiczna wymiana cząsteczek jest dowodem na to, że dyfuzja zachodzi w temperaturze 0 stopni
W stanie ciekłym występują 2 typy uporządkowanych form wyższych - minimalne układy przestrzenne:
struktura kulista (klatrat) - utworzona z 20 cząsteczek tworzących 12 pięcioboków wokół innej, np. organicznej cząsteczki
struktura pentagonalna - dwa pięcioboki na krawędzi połączone z sześciobokami
z zawiniętymi narożnikami. Z nich powstają formy clusters. Model Nemethy, Scheraga
W modelu tym przyjmuje się, że woda składa się z obszarów cluster, w których cząsteczki powiązane są wiązaniami wodorowymi, przestrzeń między nimi wypełniona jest nie powiązanymi wodorowo cząsteczkami wody.
W temperaturze 0 oC chmura ma zawierać 90 , a w temp. 10 oC, 57 cząstek wody (70% cząstek powiązanych wodorowo). Teoria ta tłumaczy zmiany objętościowe wody w zależności od temperatury (objętość właściwa w stanie stałym o 10% większa niż w stanie ciekłym - najmniejsza objętość i największa gęstość w temp. 4 oC
Znaczenie oddziaływań cząsteczek wody podczas tworzenia uporządkowanej struktury układów biologicznych
Scharakteryzuj przewodnictwo równoważnikowe oraz promienie hydratacji jonów.
Promień hydratacji jonów
Promień hydratacji maleje ze wzrostem ciężaru cząsteczkowego. Tłumaczy to różnice przenikalności potasowców przez błonę komórkową Ciągły wypływ K+ przez błonę zapewnia stały potencjał spoczynkowy. Cząsteczki o dużym promieniu hydratacyjnym z trudnością przeciskają się przez pory i kanały znajdujące się błonie komórkowej.
Hydratacja cząstek biopolimer warunkiem zachowania natywnej struktury. Zmiany w stopniu uwodnienia oraz wynikające zaburzenia strukturalne można śledzić za pomocą spektroskopii dielektrycznej.
Jon znajdujący się w roztworze otoczony jest cz. H2O, których ilość zmienia się przy ruchomości jonów. Im mniejszy promień jonu - tym większy promień hydratacji.
Przewodnictwo równoważnikowe to przewodnictwo takiej porcji elektrolitu, która zawiera 1 mol ładunków dodatnich i 1 mol ładunków ujemnych. Objętość tej porcji to objętość równoważnikowa Vr.
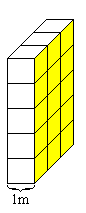
Przewodnictwo równoważnikowe zdefiniowane jest jako przewodnictwo Vr sztuk jednostkowych kostek elektrolitu ustawionych jak na rysunku. Gradient potencjału elektrycznego przyłożony do równoległych ścianek (żółte).
jednostki
Przewodnictwo równoważnikowe w funkcji stężenia czyli podział na elektrolity mocne i słabe.
Oznaczenia :
oraz
|
|
Taki typ elektrolitu, gdzie słabo zależy od stężenia i to zależy w sposób to elektrolit mocny. |
Taki typ elektrolitu, gdzie zależy od stężenia wyraźnie i to zależy krzywoliniowo to elektrolit słaby. |
Przykład: wodny roztwór HCl |
Przykład: wodny roztwór CH3COOH |
Przyczyny spadku przewodnictwa równoważnikowego przy wzroście stężenia: |
|
1.Jonów coraz gęściej i w wędrówce w przeciwne strony zaczynają sobie przeszkadzać. 2. (powstaje taka sama ilość jonów z 1 mola substancji, bo zawsze 100%dysocjacji) |
1.Jonów coraz gęściej i w wędrówce w przeciwne strony zaczynają sobie przeszkadzać.
2.Im większe stężenie, tym mniej jonów z 1 mola substancji powstaje (cofa się dysocjacja). |
Przenikalność dielektryczna wody
Przenikalnosć elektryczna- wielkość fizyczna, oznaczana grecką literą ε (epsilon), charakteryzująca właściwości elektryczne środowiska. Dla substancji izotropowych jest to wielkość skalarna, równa stosunkowi indukcji pola elektrycznego do natężenia tego pola:
W ciałach anizotropowych przenikalność jest tensorem drugiego rzędu (drugiej rangi). Wymiarem przenikalności elektrycznej w układzie SI jest F·m-1 (farad na metr).
Warstwa podwójna Helmholtza i warstwa dyfuzyjna Sterna
Przy omawianiu oddziaływań wewnątrz cząsteczkowych i międzycząsteczkowych zwracaliśmy uwagę na rożne sposoby polaryzacji elektrycznej cząsteczek. Powierzchnie graniczne, istniejące w układach biologicznych, np. błony otaczające komórki lub organelle, traktowaliśmy jako nośniki ładunków elektrycznych.
Ładunki te przyciągają inne (np. jony). Dokoła takiej struktury, jak również wokół naładowanej lub spolaryzowanej cząsteczki, tworzy się więc warstwa ładunków o znakach przeciwnych.
Obszar, w którym znajduje się warstwa ładunków pewnego znaku i warstwa ładunków znaku przeciwnego, nazywamy elektryczną warstwą podwójną. Wprowadziliśmy pojęcie warstwy hydratacji jonów. Jest to szczególny przypadek warstwy podwójnej.
Tego rodzaju warstwy wprowadził po raz pierwszy Helmholtz. Założył on po prostu, że naprzeciw stałej warstwy ładunku struktury znajduje się uporządkowana warstwa ruchomych nośników ładunku. Warstwa taka, zwana podwójną warstwą Helmholtza, ma określoną grubość.
Wprowadziliśmy pojęcie potencjału elektrycznego (
). W punkcie o współrzędnej r jest on równy pracy, którą trzeba wykonać, aby przesunąć ładunek jednostkowy z nieskończoności do tego punktu.
Potencjał jako funkcja r wywołany jest przestrzennym oddziaływaniem wszystkich nośników ładunku. Przy tworzeniu się warstwy podwójnej następuje zmiana pierwotnego potencjału stałego ładunku warstwy granicznej. (rysunek z wykładu) Pokazany jest przebieg funkcji
(r). Dla podwójnej warstwy Helmholtza funkcja y (r) jest liniowa. Taki układ warstw można bowiem traktować jako kondensator płaski, którego jedna płytka jest powierzchnią graniczną struktury, a druga powierzchnią utworzoną przez stałe jony warstwy podwójnej.
Jak już wyjaśniliśmy natężenie pola w kierunku r określone jest w następujący sposób:
Aby obliczyć ψ, należy podstawić w równaniu wartość
zamiast E i scałkować otrzymane równanie (całkowanie jest procesem odwrotnym' do różniczkowania). Zakładając, że żadna z wielkości znajdujących się po prawej stronie tego równania nie jest funkcją odległości (również D!), otrzymamy:
gdzie K jest stałą całkowania.
Stała całkowania zniknie, gdy utworzymy różnicę potencjałów = 2-1.
Funkcja
jest więc istotnie funkcją liniową r.
Okazało się, że w niektórych przypadkach hipoteza Helmholtza jest nieprawdziwa.
Gdy Gouy i Chapman badali dyfuzyjną warstwę podwójną, którą pod pewnym względem można w zasadzie przyrównać do atmosfery Ziemi. Za pomocą prawa rozkładu Boltzmana autorzy ci wyznaczyli rozkład ruchomych nośników ładunku w rożnych odległościach od powierzchni, o stałym ładunku, czyli na rożnych poziomach energetycznych.
Główna myśl podanej przez nich teorii jest następująca: ruchome nośniki ładunku dążą w zasadzie do utworzenia podwójnej warstwy Helmholtza. Na przeszkodzie stają jednak ruchy cieplne. Efektem ich działania byłby całkowicie równomierny rozkład ładunku. Struktura dyfuzyjnej warstwy podwójnej jest wypadkową obu tych procesów.
Jak widać potencjał nie jest w tym przypadku funkcją liniową r i sięga głębiej w roztwór niż przy podwójnej warstwie Helmholtza.
Teoria dyfuzyjnej warstwy podwójnej jest zgodna z doświadczeniem dla roztworów o niewielkim stężeniu. Przy większych stężeniach pojawiają się duże odstępstwa od tej teorii i dlatego trzeba było wprowadzić dalszą jej modyfikację. Lepszą zgodność z doświadczeniem daje teoria warstwy podwójnej Sterna.
Stern połączył teorie Helmholtza i Gouy' -Chapmana założeniem, że jedna część ruchomych nośników ładunku, zgodnie z teorią Helmholtza, tworzy warstwę znajdującą się naprzeciw ładunków ustalonych. Ładunek tej warstwy nie jest jednak równy ilościowo stałemu ładunkowi powierzchni i wobec tego na większych nieco odległościach od tej powierzchni istnieją chmury jonów związane z tą warstwą. Rozdział ruchomych nośników ładunku na te dwie składowe części warstwy podwójnej Sterna zależy od stężenia jonów w roztworze.
Przy bardzo małym stężeniu wszystkie nośniki ładunku znajdują się w warstwie dyfuzyjnej. Wraz ze wzrostem stężenia jonów warstwa Helmholtza jest coraz silniej obsadzana przez ruchome nośniki ładunku. Nie możemy tu przedstawić ilościowo tego rozkładu ładunków. Potencjał, jest w tym przypadku bardziej skomplikowaną funkcją r.
Można dokładnie określić zarówno grubość warstwy dyfuzyjnej, jak i grubość podwójnej warstwy Sterna. Można również zdefiniować efektywną grubość tych warstw. Jest to taka grubość, która odpowiada odległości płytek kondensatora o tej samej pojemności.
Zmiany potencjału warstwy podwójnej Sterna zależą w dużym stopniu od różnych chemicznych sił wiązania. W pobliżu powierzchni może pojawić się nadwyżka jonów przeciwnego znaku wywołana absorpcją cząstek spolaryzowanych, która w przypadku ekstremalnym może spowodować odwrócenie potencjału pola. Istnienie warstw podwójnych należy brać pod uwagę przy wszelkich rozważaniach molekularno-biologicznych.
Istnieniem tych warstw tłumaczymy fakt, że stężenie cząsteczek spolaryzowanych lub jonów na powierzchniach granicznych jest inne w roztworze znajdującym się w spoczynku niż w roztworze, który np. zamieszamy. Fakt ten ma szczególnie duże znaczenie przy obliczaniu strumieni lub potencjałów dyfuzyjnych w oparciu o różnice stężeń między dwoma składnikami rozdzielonymi błoną. Istnienie warstw podwójnych należy również uwzględniać przy badaniu przepływu na poziomie molekularnym.
Warstwy podwójne grają ważną rolę przy wyprowadzaniu zmiennych napięć elektrycznych z obiektów biologicznych za pomocą elektrod. Elektryczna warstwa podwójna na katodzie metalicznej ma właściwości kondensatora. Opór pojemnościowy takiego kondensatora i opór omowy tej warstwy granicznej stanowią łącznie o wartości impedancji elektrody. Pojemność właściwa wypolerowanej powierzchni platynowej zanurzonej w roztworze biologicznym, przy częstości 1 kHz wynosi około
. Istniejące na elektrodach warstwy podwójne są poważnym źródłem błędów pomiarów fizjologicznych.
Przewodnictwo wody (ruchliwość jonów, elektroforeza)
Ruchliwości kationów i anionów nie są jednakowe i oznacza się je uk (u+) i ua (u-). Ruchliwość jonów wzrasta z rozcieńczeniem i ma wartość maksymalną dla roztworu nieskończenie rozcieńczonego, co nazywa się graniczną ruchliwością jonu: uo+ i uo-.
Ruchliwość kationu
U+=Z+/k•η
Prędkość
V+=(Z+/k•η)E= U+•E
U=V/E
[m2/volt•s]
V+- prędkość jonów o ładunku dodatnim
E -natężenie pola elektrycznego
U+- wskaźnik ładunku jonu (określa liczbę i znak ładunku jonu)
η - współczynnik lepkości
k- stała
Elektroforeza - technika analityczna, rzadziej preparatywna, stosowana w chemii i biologii molekularnej, zwłaszcza w genetyce. Jej istotą jest rozdzielenie mieszaniny związków chemicznych na możliwie jednorodne frakcje przez wymuszanie wędrówki ich cząsteczek w polu elektrycznym.
Cząsteczki różnych substancji różnią się zwykle ruchliwością elektroforetyczną. Parametr ten jest w przybliżeniu wprost proporcjonalny do ładunku elektrycznego cząsteczki i odwrotnie proporcjonalny do jej wielkości. Zależy także od kształtu cząsteczki.
Szybkość wędrowania cząsteczek zależy przede wszystkim od ich wielkości, posiadanego ładunku i masy cząsteczkowej, co zezwala na rozdział układów o różnej wielkości i budowie cząsteczek
Potencjał przepływu
Rozważaliśmy elektryczną warstwę podwójną utworzoną na powierzchniach granicznych jako wynik działania elektrostatycznych sił przyciągania i fluktuacji termicznych. Nie uwzględniając fluktuacji statystycznych, powierzchnię graniczną z taką warstwą podwójną możemy traktować jako konkretną, obojętną elektrycznie strukturę układu. Sytuacja zmienia się jednak, gdy w układzie występują zakłócenia wywołane przez siły wewnętrzne, np. podczas ruchu roztworu względem powierzchni granicznej lub pod działaniem zewnętrznego pola elektrostatycznego. W tym przypadku nośniki ładunku części dyfuzyjnej podwójnej warstwy Sterna zostają odrywane i ładunek znajdujący się na 'Powierzchni granicznej nie jest zrównoważony. Pomiędzy powierzchnią graniczną i wnętrzem roztworu powstaje potencjał elektryczny, zwany potencjałem elektrokinetycznym lub potencjałem z (dzeta). Dającym się zmierzyć efektem tego procesu są: potencjał przepływu, elektroosmoza, elektroforeza i potencjał elektroforetyczny.
Potencjał przepływu pojawia się, gdy ciecz przepływa przez kapilarę, na której ściankach istnieje stały ładunek. Jeśli ścianka naładowana jest np. ujemnie, to dodatnie jony roztworu przesuwają się w kierunku warstw cieczy znajdujących się w pobliżu ścianek. Szybkość przepływu wody nie jest taka sama w całym przekroju kapilary: w środku prędkość przepływu jest większa niż przy ściankach. Dlatego jony dodatnie, znajdujące się w pobliżu ścianek, będą płynęły wolniej niż jony ujemne, znajdujące się w środkowej warstwie
kapilary. Z tego właśnie powodu jony ujemne będą przechodziły przez kapilarę szybciej niż dodatnie. Jeśli pory błony są kapilarami, to po jednej stronie błony nagromadzi się więcej jonów ujemnych niż po drugiej. W ten sposób wytworzy się duży spadek potencjału elektrycznego. Potencjał ten działa silnie elektroforetycznie na jony i wywołuje kompensujący prąd jonów. Podobnie, jak przy potencjale dyfuzyjnym, prąd kompensujący
działa tak, że ładunek znajdujący się po obu stronach błony makroskopowo jest taki sam. Jednakże ze względu na opór elektryczny błony nie można nigdy osiągnąć takiego stanu równowagi i zawsze istnieje bardzo niewielka różnica ładunków po obu stronach błony, której rezultatem jest potencjał przepływu przy przewodnictwie właściwym błony σ stałej dielektrycznej D oraz różnicy ciśnień p po obu stronach błony wynosi:
ΔΨs = − (D εrε0ξΔπ)/(ησ)=(Δpkηu)/(ησεrE)
Gdzie:
x- oznacza potencjał elektrokinetyczny,
h- lepkość roztworu,
ε0- przenikalność elektryczną próżni
εr- przenikalność elektryczna ośrodka.
Wzór ten jest jednak słuszny tylko wtedy, gdy promień porów jest duży w stosunku do efektywnej grubości podwójnej warstwy Sterna. Pory większości błon biologicznych nie spełniają tego warunku. Przy tego rodzaju kapilarnych porach przewodnictwo elektryczne jest funkcją odległości od ścian kapilary. Zmienia się również wartość stałej dielektrycznej w pobliżu ścianek pod wpływem panującego tam silnego pola elektrostatycznego (E od 105 do 106 V/cm).
Przy wyznaczaniu potencjału przepływu możemy ominąć niektóre z tych trudności, jeśli nie będziemy mierzyć bezpośrednio potencjału przepływu, lecz prąd ładunków powstały przy przepływie rozpuszczalnika. Możemy tego dokonać za pomocą techniki prądów zwarcia, stosowanej często przy pomiarze aktywnego transportu jonów przez błony biologiczne.
Przykładem badań osobliwych cech potencjału przepływu w błonach biologicznych o bardzo wąskich porach są pomiary przeprowadzone na pęcherzyku żółciowym królika. Badania te rzuciły światło na właściwości porów, przez które przepływa roztwór. Przy badaniu cienkich błon stosowano różnice ciśnień osmotycznych, a nie hydrostatycznych. W przeprowadzonym doświadczeniu błona pęcherzyka żółciowego rozdzielała dwa roztwory o tym samym składzie elektrolitycznym, ale o rożnej zawartości sacharozy. Gdy roztwór znajdujący się po stronie błony śluzowej (mukoza) był hipertoniczny, to woda płynęła od strony błony surowiczej (seroza) do śluzowej. Potencjał przepływu był wtedy dodatni po stronie błony śluzowej. Wskazywałoby to, że przez błonę przenika więcej jonów dodatnich niż ujemnych. Mogłoby się wydawać, że jest to sprzeczne z teorią, która mówi, że ściany tych porów są naładowane ujemnie, a więc jony ujemne płynące środkiem kanału powinny płynąć szybciej niż kationy znajdujące się w pobliżu ścian. Istotnie, ujemny ładunek ścian wpływa bezpośrednio na skład roztworu w porach. Ponieważ jednak całość musi być elektrycznie obojętna, więc we wnętrzu kanału w roztworze elektrolitu występuje większe stężenie kationów niż anionów o tyle, a ile wymaga tego konieczność skompensowania stałego ładunku ściany kapilary. Przy wyjątkowo wąskich porach, o dużej względnej powierzchni ścianek gęstość stałego ładunku jest tak duża, że równowaga jonów w roztworze jest silnie zakłócona. W porach, których ścianki naładowane są ujemnie, stężenie. Kationów jest o wiele większe niż anionów. Dlatego przy przepływie roztworu przez takie pory przechodzi przez nie więcej kationów niż anionów, odpowiednio do różnicy stężeń. Jeśli potencjał przepływu może być wywołany ruchem wody, to i odwrotnie potencjał elektrokinetyczny może wywołać przepływ wody.
Tego rodzaju zjawisko nazywa się -elektroosmozą. Nie wiemy jeszcze czy ma ono jakieś
znaczenie w procesach biologicznych, np. w transporcie wody. Badania Bowlinga wykazały np., że elektroosmoza może być siłą napędową przy transporcie pewnych substancji w roślinach kwiatowych. Płytki siatkowe tych roślin działają jak porowaty filtr posiadający na powierzchni pewien ładunek.
Bardzo pomocna w rożnego rodzaju badaniach jest elektroforeza. Przez elektroforezę lub kataforezę rozumiemy ruch naładowanych cząstek w polu elektrycznym. Badania elektroforetycznego ruchu jednokomórkowców (bakterie, drożdże, pierwotniaki, erytrocyty) pozwoliły określić właściwości powierzchniowe ich błon w różnych warunkach. Załóżmy, że erytrocyty mają w przybliżeniu kształt kuli. Siła tarcia, powstała przy ich ruchu, określona jest wtedy w przybliżeniu przez prawo Stokesa. W rozważaniach podobnych do tych, które przeprowadziliśmy przy wyznaczaniu promienia hydratacji można zastąpić tę siłę tarcia przy jednostajnym ruchu erytrocytów siłą pola elektrycznego, która jest iloczynem natężenia pola i ładunku powierzchniowego. Traktując powierzchnię erytrocytów jako kondensator kulisty możemy oszacować potencjał elektrokinetyczny bezpośrednio z ruchu elektroforetycznego komórek, który możemy mierzyć bezpośrednio obserwując przy pomocy mikroskopu ruch komórek w polu elektrycznym. Fritze otrzymał dla potencjału erytrocytów krwi ludzkiej w roztworze o sile jonów równej 0,024 wartość - 40 mV. Przez ekstrapolację do siły jonowej równej zero potencjał' zmienia się do -79 mV. Siłą jonową roztworu nazywamy wyrażenie:
n ½ Σ ci z2i i=1
Błona komórkowa znajdująca się w obszarze obojętnym jest naładowana ujemnie w stosunku do otoczenia przez spolaryzowane grupy swych cząsteczek. Potencjał elektrokinetyczny zależy w dużym stopniu od stężenia elektrolitu i wartości pH roztworu oraz od obecności innych cząsteczek w otoczeniu komórki. Długie cząsteczki białka, np. fibrynogenu, mogą zmniejszać potencjał elektrokinetyczny na skutek pokrycia przez nie
powierzchni naładowanych błon. Potencjał t; erytrocytów zapobiega ich aglutynacji, ponieważ jednakowo naładowane cząsteczki odpychają się.
Wspomniany poprzednio potencjał elektroforetyczny pozostaje w pewnym sensie w stosunku odwrotnym do elektroforezy. Przy wymuszonym ruchu naładowanych elektrycznie cząstek, np. w polu grawitacji, powstaje potencjał między cząsteczkami zawiesiny i środkiem, w którym się one znajdują.
Warstwy monomolekularne i podwójne lipidów (ocena grubości)
Charakterystyczna cechą związków lipidowych jest amfifilowość, czyli dwojaki sposób wiązania się z wodą. Jest to następstwem faktu, że reszty kw. tłuszczowych mają charakter hydrofobowy, natomiast znajdująca się po przeciwnej stronie cząsteczki gr. fosforanowa jest grupą hydrofilową.
W ośrodkach wodnych strukturą energetycznie korzystną dla fosfolipidów jest struktura, w
której polarne reszty kwasów tłuszczowych nie mają kontaktu z wodą. Strukturami takimi są:
micele - tworzone przez lipidy z jednym łańcuchem wodorotlenowym
liposomy - powstające przy odpowiednim stężeniu lipidów np. w wyniku
naświetlania r-ru, ultradźwiękami lub wstrząsaniadwumolekularna warstwa lipidów - powstaje na granicy dwóch faz polarnych,
końcówki hydrofobowe oddziałują na siebie siłami van der Waalsa
Otrzymywanie błon bimolekularnych:
Otrzymywanie błony o pow. sferycznej
Na końcu cienkiej igły połączonej ze strzykawką umieszcza się kroplę r-u lipidów w niepolarnym,
lotnym rozpuszczalniku a następnie przez powolne wyciskanie r-ru wodnego ze strzykawki uzyskuje
się błonę.
Otrzymywanie błon niesymetrycznych
Złożenia 2 błon lipidowych utworzonych na pow. wody dokonuje się tego przez zanurzenie
przegrody z otworem w fazę wodna; obie błony powierzchniowe odkładają sie na ściankach
przegrody zlepiając sie ze sobą na pow. otworu.
Na otwór w środkowej ściance pojemnika wypełnionego woda lub roztworem wodnym
elektrolitu nanosi sie za pomocą pipety, mikrostrzykawki lub pędzelka niewielka ilość
roztworu lipidu w rozpuszczalniku niepolarnym . otrzymana w ten sposób błonę, początkowo
o dużej grubości obserwuje sie w świetle odbitym. Dopóki grubość błony jest porównywalna
z długością fali światła widzialnego, widoczne są na jej powierzchni barwy interferencyjne.
Obszar, w którym błona jest bimolekularna, odbija znacznie mniej światła , przez co
widoczny jest jako czarna plama w otoczeniu jasnej, barwnej błony - stad nazwa „czarna
błona lipidowa”
Cząsteczki zajmują taką odległość, aby nakład energii na oddziaływania był jak najmniejszy.
W zależności od tej odległości wyróżniamy warstwy:
gazowa
ciekła rozciągnięta
ciekła skondensowana
stała
Pomiaru grubości błony lipidowej można dokonać 2 metodami:
Metoda spektrofotometryczna (mierzy się współczynnik odbicia światła od błony) wiązka światła o natężeniu I0 i długości fali λ pada na błonę lipidową pod pewnym kątem.
Część światła odbija się od pierwszej warstwy, część przechodzi przez błonę, część odbija się od drugiej warstwy a następnie interferuje z wiązką odbitą od warstwy pierwszej. Następuje wygaszenie.
Badamy natężenie IR światła odbitego i obliczamy współczynnik odbicia światła R.
R = IR/I0
Następnie znając współczynnik załamania błony n i współczynnik załamania r-u wodnego obliczamy grubość błony x:
R = 4 [(n - no) / (n + no)]2 · sin2 (2πnx/λ)
W metodzie tej mierzona jest całkowita grubość błony, tzn. wewnętrznej warstwy hydrofobowej i obu warstw zewnętrznych hydrofilowych.
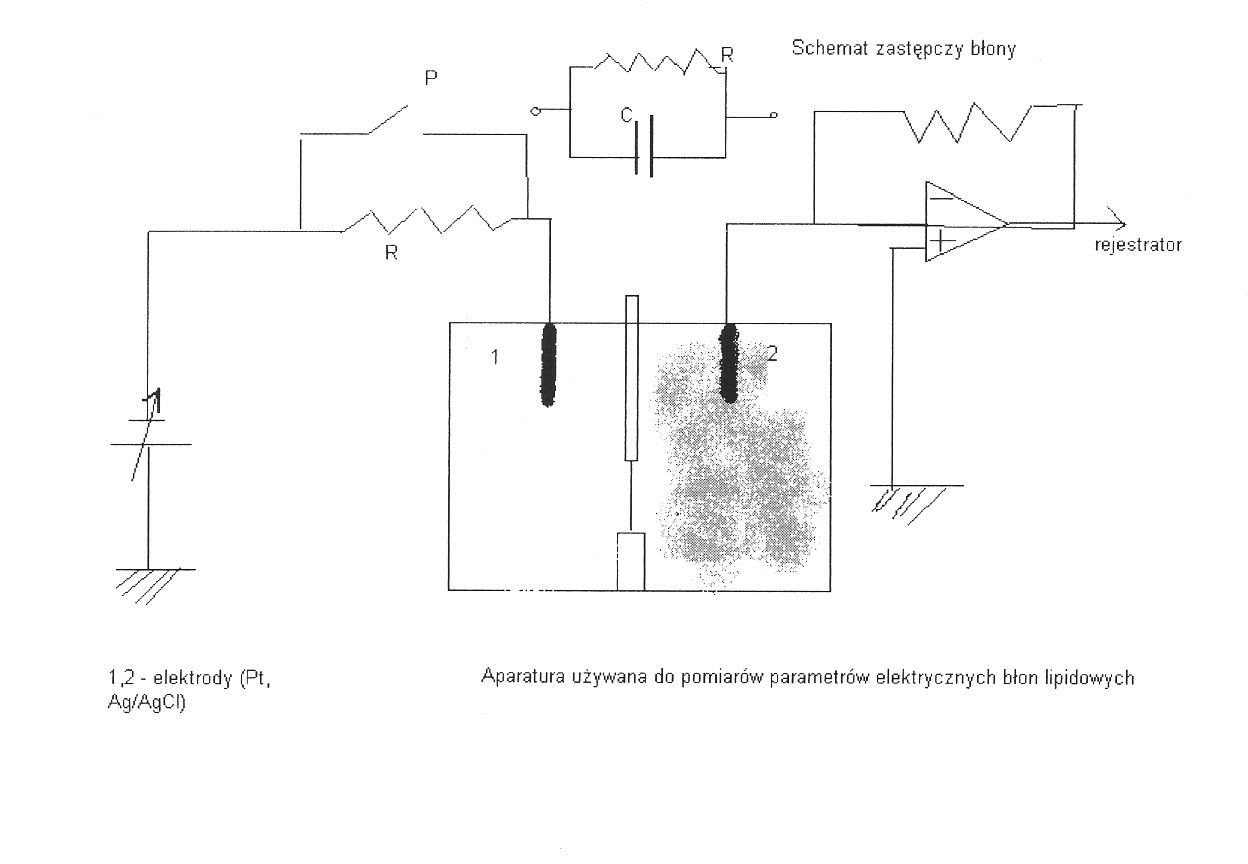
Metoda elektryczna (mierzy pojemność elektryczną właściwą)
Pomiaru pojemności błony mozna dokonać umieszczając odpowiednie elektrody w roztworach po obu stronach błony i uzywając mostka RC. Włączanie i wyłączanie napięcia podawanego na elektrodę 1 przez opornik R wywołuje impulsy prądu zapisywane na rejestratorze. Pojemność błony mozna obliczyć z szybkości zaniku impulsów prądu. Grubość błony oblicza się przyjmując, ze stanowi ona, wraz z roztworami wodnymi, kondensator płaski. Zakłada się przy tym wartość stałej dielektrycznej błony typową dla węglowodorów nasyconych ε = 2,3 ÷ 2,5.
C = εoεrS/d
Gdzie:
d - grubość warstwy błony
S - pow. okładek kondensatora
Z pomiaru pojemności otrzymuje się grubość jedynie hydrofobowego wnętrza błony, a to dlatego, że tylko ta część błony stanowi obszar niedostępny dla jonów i wody i spełnia rolę dielektryka oddzielającego 2 fazy wodne.
Właściwości błony biologicznej i błon bimolekularnych
Każda komórka otoczona jest błoną komórkową- rodzajem granicy, która pozwala na zachowanie integralności komórki przez zachowanie charakterystycznego dla niej składu chemicznego, różniącego się często w istotny sposób od stanu jej otoczenia. Wewnątrz komórki błony otaczają organelle tworzące wyodrębnione obszary, w których zachodzą specyficzne reakcje biochemiczne. Kontakt między wnętrzem a środow. zewn komórki czy innej struktury ograniczonej błoną jest możliwy dzięki zdolności błon do wybiórczego przepuszczania określonych substancji. Ponadto przepuszczalność błony podlega procesom
regulacji i może ulegać zmianom pod wpływem określonych bodźców chemicznych bądź elektrycznych.
Według modelu mozaikowego błonę biologiczną tworzy podwójna warstwa lipidowa. Apolarne łańcuchy lipidowe skierowane są do wnętrzna błony , tworząc jej hydrofobowe wnętrze, natomiast fragmenty posiadające elektryczny moment dipolowy skierowane są na zewnątrz błony w kierunku środowiska wodnego. Z tak utworzoną matrycą związane są białka:
białka integralne przenikające poprzez błonę
białka peryferyjne związane z powierzchnią błony
Białka błonowe pełnią kluczową rolę w funkcjonowaniu komórki: są odpowiedzialne za wybiórczy transport przez błonę określonych substancji, tworząc system kanałów jonowych i system transportu wbrew istniejącym bodźcom termodynamicznym, pełnią też rolę receptorów, neuroprzekaźników i antygenów.
Płynności błony zależy od:
składnika lipidowego : czy są nasycone czy nienasycone, długie czy krótkie łańcuchy
dynamiki lipidów i uporządkowania łańcuchów arylowych kwasów tłuszczowych, zależnych m.in. od temp. głównego przejścia fazowego o
oddziaływań białko - lipid (zjawisko lipidu granicznego), lipid- lipid (stosunek zawartości cholesterolu do fosfolipidów i SM do PC)
dualizm działania cholesterolu:
usztywnienie błony w fazie ciekłokrystalicznej
upłynnienie jej w fazie żelu

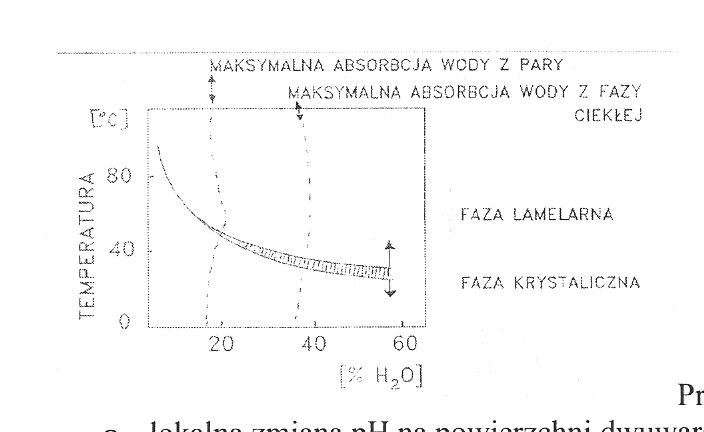
Przejście fazowe lipidów (czynniki wpływające)
Niezwykle ważną właściwością lipidów tworzących dwuwarstwy jest zdolność do zmiany fazy - możemy wyznaczyć temp. przejść fazowych dla różnych błon lipidowych. W błonie biologicznej takie zmiany występują lokalnie i umożliwiają obniżenie oporności, zwiększając przepuszczalność.
Czynniki wywołujące zmianę fazy:
stopień uwodnienia
temperatura
a) im dłuższy łańcuch acylowy- tym wyższa
b) im więcej wiązań podwójnych - tym nizsza
poniżej temp. przejścia fazowego lipidy tworzą strukturę krystaliczną, powstają
formy żelowate (fosfolipidy ściśle upakowane brak ruchu)powyżej temp. przejścia fazowego lipidy tworzą struktury ciekło - krystaliczne (charakteryzują się rozluźnieniem upakowania, w ich budowie uczestniczy woda)
temp przejścia fazowego dla lipidów, które występują w błonach biologicznych
wynosi ok. 40 C
Czynniki regulujące:
cholesterol - jego dodatek obniża temp głównego przejścia fazowego , dzięki niemu
można uzyskać fazę wysoko uporządkowaną i płynną
pH, obecność Ca2+ i innych substancji, które mogą być wbudowane w błonę
związane z budową lipidu
stopień nasycenia ( więcej nienasyconych kwasów - temp przejścia fazowego niższa)
wielkość kwasu tłuszczowego (im dłuższy łańcuch tym temp wyższa)
Przejścia fazowe można pobudzić:
lokalną zmianą pH na powierzchni dwuwarstwy
związaniem jonów Ca2+
wbudowaniem polarnych cząsteczek peptydów i białek
zmianą temp.
Warstwy Langmuira-Blodgetta
Etapy otrzymywania:
1. Podłoże hydrofobowe- (materiały dzięki którym można uzyskać warstwę: spreparowane szkło, metale Ag, Au) zanurzamy materiał, cz. hydrofilowe błony oddziałują, odkłada się 1 warstwa, przy wyciąganiu płytki odkłada się kolejna
2. Podłoże hydrofilowe - (materiały dzięki którym można uzyskać warstwę: czyste szkło, metal pokryty tlenkiem) 1 warstwa odkłada się dopiero po wyciągnięciu płytki do góry, powtarzając ten proces wielokrotnie, można otrzymać warstwy L-B zbudowane z setek warstw.
Warstwy Langmuira - Blodgetta są to warstwy wielocząsteczkowe tworzone na podłożu stałym. Możemy otrzymać warstwy o różnej orientacji cząsteczek. Ważne, aby w procesie ich nakładania ciśnienie powierzchniowe Π było stałe - jest to niezbędne, aby zachować identyczną strukturę następnej warstwy. Amfifilowe właściwości lipidów, dzięki którym mogą one tworzyć stabilne warstwy monomolekularna na powierzchni wody, umożliwiają również stworzenie w sposób kontrolowany warstw wielocząsteczkowych na podłożu stałym. Dzięki tej metodzie można otrzymać białka integralne o nienaruszonych centrach aktywnych.
Dynamika dwuwarstwy lipidowej (powstawanie domen)
Domeny lipidowe to zgrupowania lipidów o tym samym ładunku, mające charakter silnie polarny.
Dwuwarstwa lipidowa jest strukturą niezwykle dynamiczną. Zmiana pH w jej obrębie prowadzi do tworzenia się silnie polarnych domen lipidowych. Gdy pH maleje - rośnie liczba ujemnie naładowanych cząsteczek lipidów, które tworzą domeny lipidowe; gdy pH rośnie - maleje liczba ujemnie naładowanych cząsteczek lipidów i powstałe wcześniej domeny lipidowe ulegają rozpadowi.
Przykładem substancji, która zdolna jest tworzyć domeny jest kwas fosfatydowy w lecytynie.
Do domen lipidowych mogą przyłączać się jony wapnia. Przyłączenie się jonu wapniowego do cząsteczki kwasu fosfatydowego powoduje usztywnienie dwuwarstwy lipidowej w miejscu dołączenia się jonów wapniowych.
Asymetryczne rozmieszczenie lipidów w błonie komórkowej (znaczenie biologiczne)
Asymetria składu lipidowego - skład lipidowy błony jest różny w różnych komórkach czy organellach, poza tym obie monowarstwy tej samej błony mają różne rodzaje lipidów, asymetria polega na ilościowych różnicach zawartości poszczególnych klas lipidów - np. w błonie erytrocytalnej lipidy z choliną (PC, SM) są w zewnętrznej monowarstwie błony, podczas gdy lipidy za terminalną grupa aminową (PE i PS) SA w monowarstwie zewnętrznej. Kwasy tłuszczowe PC i SM SA bardziej nasycone niż PE i PS więc oprócz asymetrii rozmieszczenie głów polarnych fosfolipidów jest też asymetria dystrybucji kwasów tłuszczowych , która powoduje, że wewnętrzna monowarstwa jest bardziej płynna + dodatkowa różnica ładunku między monowarstwami, bo PS jest naładowana ujemnie. Asymetria fosfolipidów jest generowana już w miejscu powstania błon (retikulum endoplazmatyczne), poprzez odpowiednie białka ER cząsteczki fosfolipidów przemieszczane są z 1 monowarstwy do 2.
Funkcja asymetrii:
niektóre enzymy korzystają z niej, np. białkowa kinaza c - PS w formie aktywnej związana z cytoplazmatyczną powierzchnia błony w miejscu koncentracji PS, potrzebuje ujemnego ładunku tego tłuszczu
rola fosfolipidów inozytolowych w generowaniu wewnątrzkomórkowych przekaźników informacji (są one w cytoplazmatycznej monowarstwie błony)
Zmiany strukturalne w dwuwarstwie po związaniu z białkami lub peptydami
Zmiana konformacji lipidów w bezpośrednim sąsiedztwie białka po związaniu z białkami w pobliżu białka integralnego następuje usztywnienie dwuwarstwy lipidowej, przechodzi z żelu w zol ( przejście z silnie upakowanej struktury krystalicznej w ciekło - krystaliczną ), zmiana przepuszczalności, pobudzenie przejść fazowych
Zmiana temperatury przejścia
- polimyxyna obniża o 20stopni temp przejścia, wiele antybiotyków ma taki mechanizm co
ułatwia potem transport K+ z wnętrza komórki
- normalnie lipidy mają temp. topnienia znacznie wyższą od temp ciepłoty ciała im więcej
składników w błonie tym ta temp zmniejsza się i zbliża się do 36,6 lub jest od niej niższa.
Zaburzenia w budowie bimolekularnej warstwy lipidowej
- defekty błony - powoduje stan napięcia który może ulec rozładowaniu jeśli wolna przestrzeń między cząsteczkami lipidu zostanie napełniona błoną lub rozpuszczonymi w wodzie substancjami, w przypadku zmian fazowych nie nastąpi całkowita degradacja i zapadnięcie komórki, gdyż błona biologiczna rozpościera się na szkielecie z włókien aktyny i miozyny (cytoszkielet). Relaksacja - rozładowanie w wyniku którego następuje przemieszczenie domen do części hydrofobowej. Dylatacja - wbudowane białko tworzy micele (oddziałuje z częściami hydrofilowymi warstwy)
Transport wody przez błonę
Woda transportowana jest przez błonę na zasadzie osmozy. Błona ma charakter półprzepuszczalny, czyli przepuszcza rozpuszczalnik (wodę), a nie przepuszcza substancji rozpuszczonej. Transport ten jest spowodowany różnicą potencjałów chemicznych wody po obu stronach błony, powoduje to transport wody przez błonę w kierunku od roztworu o większym do roztworu o mniejszym potencjale chemicznym wody, czyli do roztworu o większym stężeniu substancji rozpuszczonej.
Polaryzacja błony komórkowej - potencjał spoczynkowy
Potencjał elektryczny wnętrza komórki jest różny od potencjału jej środowiska zewnętrznego. Można to wykazać doświadczalnie za pomocą mikroelektrod, z których jedną wkłuwa się do wnętrza komórki, a drugą umieszcza na jej powierzchni.
Różnica potencjałów zmierzona woltomierzem o bardzo dużym oporze wewnętrznym wynosi
około 50 ÷ 100 mV, w zależności od rodzaju komórki.
Mechanizm potencjału spoczynkowego wynika z:
ujemnego potencjału Donnana
różnej przepuszczalności błony dla jonów (potencjał dyfuzyjny)
aktywnego transportu jonów
Potencjał spoczynkowy jest efektem współdziałania kilku mechanizmów. Potencjał spoczynkowy jest uzależniony od trzech rodzajów jonów: potasu, sodu i chloru. Ponieważ stężenia jonów sodu i potasu na zewnątrz i wewnątrz komórki są odpowiednio bliskie siebie co do wartości, a jony chloru są w równowadze, największy wpływ wywierają jony potasu, dla których błona ma największą półprzepuszczalność.
U podstaw zmiennej przepuszczalności błony komórkowej leżą molekularne zmiany w strukturze kanałów jonowych, które są zbudowane z podjednostek białkowych będących strukturalnymi białkami błony.
Potencjał spoczynkowy dotyczy wyłącznie komórek w stanie niepobudzonym
Jest uzależniony od stężeni K+ Na+ Cl- A- których stężenie nie jest jednakowe wewnątrz i na zewnątrz komórki
Potencjał wnętrza komórki jest ujemny w stosunku do jego otoczenia (-40 - -100 mV)
Potencjał spoczynkowy neuronu -90mV
Potencjał spoczynkowy to różnica potencjałów (napięcie) między obiema stronami błony plazmatycznej niepobudzonej komórki pobudliwej.
Wszystkie napięcia na błonie wyraża się jako różnice potencjału wnętrza komórki do potencjału po stronie zewnętrznej. Potencjały spoczynkowe mają wartości ujemne, które w komórkach nerwowych wahają się między -65 mV a -90 mV. Powstanie potencjału spoczynkowego jest spowodowane przede wszystkim tendencją jonów potasu do przepływania zgodnie z gradientem stężenia tych jonów z wnętrza na zewnątrz błony komórkowej. Powoduje to pozostanie niewielkiego nadmiaru ładunków ujemnych po wewnętrznej stronie błony. Inne jony (np. sodu) jedynie w niewielkim stopniu wpływają na wartości potencjału spoczynkowego. Siła elektrochemiczna, powodująca ruch jonu poprzez błonę komórkową, jest różnicą między potencjałem spoczynkowym a potencjałem równowagi dla danego jonu. Potencjał równowagi jonu jest to taki potencjał, przy którym wpływ tego rodzaju jonów z komórki jest równy ich wpływowi do jej wnętrza.
Równowaga Donnana
Przez błony komórkowe na ogół mogą przenikać elektrolity o niskich masach cząsteczkowych, lecz błony te są nieprzepuszczalne dla elektrolitów makrocząsteczkowych. W szczególności przez błony te może przechodzić wiele soli nieorganicznych, lecz nie mogą przez nie przenikać białka. Jeżeli jony wchodzące w skład soli nie są transportowane aktywnie, ich rozkład po obu stronach błony w obecności zdysocjowanych makrocząsteczek jest określony przez warunek równowagi Donnana.
W układzie tym będzie miał miejsce wypadkowy przepływ NaCl przez błonę, póki nie ustali się stan równowagi, określony przez następujące warunki:
roztwory po obu stronach błony muszą być elektronie obojętne
potencjał elektrochemiczny elektrolitu przenikającego przez błonę musi być jednakowy w obu przedziałach.
X moli jonów przeszło przez błonę
Błona ładuje się przy określonej różnicy potencjałów -V, ruch jonów zostaje wówczas
zatrzymany i ustala się stan równowagi [przy określonym stężeniu jonów po obu stronach
błony].
Potencjał błonowy wyraża się wzorem:
różnica potencjału Donnana w stanie równowagi w roztworach rozcieńczonych wynosi kilka mV, a w stężonych kilkanaście mV
osiągniecie stanu równowagi oznacza śmierć dla komórki
Wzór ten odpowiada stanowi równowagi, nie wiec dziwnego, ze nie daje poprawnych
wyników dla wytłumaczenia powstania potencjały spoczynkowego błony. Teoria Donnana
nie uwzględnia transportu aktywnego.
Donnana równowaga, równowaga membranowa, równowaga ustalająca się między 2 roztworami przedzielonymi przegrodą (membraną) półprzepuszczalną; wykorzystywana przy oczyszczaniu koloidów z soli mineralnych; odgrywa istotną rolę w procesach zachodzących w organizmach żywych.
Porównać graficznie równanie Nernsta i Goldmanna - uzasadnić słuszność
Porównanie jak opisuje potencjał spoczynkowy wzór Goldmana a jak Nernsta na przykładzie zmiany potencjału w zewnątrzkomórkowego stężenia jonów K+ ponieważ K+ są głównymi jonami wewnątrzkomórkowymi możemy łatwo zmieniać stosunek molowy K+ we wnętrzu od strony zewnętrznej poprzez zmiany środowiska w którym komórki są zawieszone gdy będziemy zwiększać stężenie K+ we wnętrzu komórki i mierzyć potencjał spoczynkowy zobaczymy zmianę potencjału. punktami na wykresie są zaznaczone pomiary potencjału spoczynkowego dla różnych stężeń K+ na zewnątrz komórki.
Przy wysokich stężeniach K+ na zewnątrz równanie Nernsta stosunkowo dobrze opisuje doświadczenie, przy niskich następuje rozbieżność i tu właściwy jest wzór Goldmana wzór Nernsta odpowiada stanowi równowagi komórki, co jest dużym uproszczeniem, ponieważ żyjąca, działająca komórka nie może się znaleźć w takim stanie . Wzór Goldmana nie odpowiada stanowi równowagi, lecz zastanej w danej chwili różnicy stężeń.
Omów znane ci typy kanałów jonowych.
Główne cechy kanałów:
zbudowane z wielu homologicznych podjednostek lub domen [4,5,6]
por przebiega zgodnie z osią symetrii kanału i jest wyściełany przez α helisy, boczne łańcuchy wystające z elementów strukturalnych, decydują o selektywności kanału
stopień selektywności zależy od średnicy największej części poru
wszystkie kanały mają stany otwarte i zamknięte i tym różnią się od prostych porów wodnych
czynnikami otwierającymi kanały są:
zmiana napięcia po obu stronach błony
bodźce mechaniczne
związanie odpowiedniej cząsteczki sygnałowej
bardzo duża szybkość przenoszenia substancji (100 razy większa niż z udziałem białek nośnikowych) - 106 jonów/s
obszary zewnętrzne podjednostek są hydrofobowe: potrzebują najmniej energii, żeby stworzyć kompleks z lipidami
podjednostki lub odcinki peptydowe ułożone koliście w dwuwarstwie lipidowej tworzą por; średnica porów jest tym większa im więcej jest podjednostek
o selektywności decyduje charakter bocznych aminokwasów skierowanych do wnętrza kanały; jeśli posiadają reszty + to kanał przepuszcza aniony, jeśli - to kationy
im mniejsza średnica tym większa selektywność kanału
otwieranie i zamykanie może być sterowane efektorami alosterycznymi (k.j.bramkowane ligandami) potencjałem elektrycznym (k.j. bramkowane potencjałami)
Typy:
kanały jonowe bramkowane ligandami
receptory jonowe klasy I charakteryzują się b, odpowiedzią układu eżektorowego; należą do
nich różniące się budową chemiczną receptory aktywowane zewnątrzkomórkowo np. receptory neuroprzekaźników jak i wewnątrzkomórkowo: fotoreceptory, receptory węchu, receptory ATP te aktywowane zewnątrzkomórkowo zbudowane są z kilku, najczęściej 5 podjednostek (łańcuchów polipeptydowych z których każdy ma 4 domeny transbłonowe) najlepiej poznanym z tej grupy jest nACh (cholinergiczny receptor nikotynowy) receptor acetylocholiny
Budowa receptorów jonowych aktywowanych wewnątrzkomórkowo jest słabo poznana
wiadomo, że zbudowane są z 6 domen transbłonowych
kanały jonowe bramkowane potencjałem (napięciowo zależne)
wyspecjalizowane struktury białkowe, uczestniczące w przenoszeniu jonów, ich wysokie przewodnictwo (107 j/s) zależy od potencjału błonowego
do najlepiej poznanych jest kanał potasowy, sodowy, wapniowy
niezależnie od różnic w budowie tych kanałów (izolowanych z tkanek różnych gatunków zwierząt) podjednostka przenosząca jony jest strukturalnie podobna
Podjednostka przewodząca kanału sodowego stanowi białko transbłonowe, którego łańcuch
24 razy przebija błonę komórkową tworząc strukturę o średnicy 5 nm
Stwierdzono, że w białkach tworzących kanały jonowe następują przemieszczenia ładunków elektronów w wyniku obniżenia różnicy potencjałów kom. Jest to pierwszy etap aktywacji kanałów, prowadzący do takich zmian kształtu lub wzajemnego usytuowania podjednostek białkowych, iż kanały otwierają się dla jonów. Po pewnym czasie (ok. 1 ms dla kanałów sodowych , dłuższym dla kanałów potasowych) następuje ich zamknięcie niezależnie od wartości zewnętrznego pola elektrycznego. Kanał pozostaje wówczas w fazie nieaktywnej (retrakcji) i dopiero po upływie pewnego czasu może znów ulec aktywacji.
Charakterystyka kanałów sodowych
w czasie pojedynczego cyklu otwarcia przez kanał sodowy przemieszcza się około 100 jonów
kanał sodowy reaguje szybciej na bodziec elektryczny niż kanał potasowy, co powoduje wzrost, a następnie spadek potencjału czynnościowego (powstanie igły potencjału)
Bezpośrednie i pośrednie bramkowanie
Neurotrasmitery mogą działać bezpośrednio i pośrednio na kanały jonowe regulujące przepływ jonów. Bezpośrednie bramkowanie odbywa się w receptorach jonotropowych. Bramkowanie pośrednie odbywa się poprzez aktywację receptorów metabotropowych. Receptory metabotropowe aktywują białko G, które aktywuje kaskadę procesów modulujących aktywność kanału jonowego.
Wyjaśnij mechanizm powstawania potencjału czynnościowego
Gdy napięcie depolaryzacyjne przekracza pewną wartość krytyczną, zwaną progową (20 - 40mV),
pojawia się potencjał czynnościowy. Bezwzględna wartość ujemnego potencjału wnętrza komórki gwałtownie spada do zera - błona ulega depolaryzacji, po czym przepolaryzowuje na znak dodatni, żeby po osiągnięciu wartości maksymalnej (ok. +40 mV) szybko spaść nawet poniżej wartości spoczynkowej i dopiero wrócić do wyjściowej. Gwałtowna zmiana potencjału przy pobudzeniu komórki nerwowej jest wynikiem szybkiego wzrostu przepuszczalności błony dla kationów, przede wszystkim sodu, a następnie podobnie szybkiego zmniejszenia tej przepuszczalności. W pierwszej fazie jony sodowe gwałtownie dyfundują do wnętrza komórki neutralizując jej potencjał ujemny, a następnie zmieniają ten potencjał na dodatni względem płynu międzykomórkowego, na skutek wytworzenia własnego potencjału dyfuzyjnego. Mniej więcej od chwili osiągnięcia maksimum potencjału dodatniego błony kom. zaczyna się zmniejszać jej przepuszczalność dla jonów Na+, natomiast wzrasta przepuszczalność dla K+.
Po ustaleniu się przepuszczalności błony na poziomie normalnym potencjał spoczynkowy stabilizuje się również na poziomie - 90 mV
im mniejsza średnica tym większa selektywność kanału
otwieranie i zamykanie może być sterowane efektorami allosterycznymi (k.j.bramkowane ligandami) potencjałem elektrycznym (k.j. bramkowane potencjałami)
Typy:
kanały jonowe bramkowane ligandami
receptory jonowe klasy I charakteryzują się b, odpowiedzią układu eżektorowego; należą do
nich różniące się budową chemiczną receptory aktywowane zewnątrzkomórkowo np. receptory neuroprzekaźników jak i wewnątrzkomórkowo: fotoreceptory, receptory węchu,
receptory ATP te aktywowane zewnątrzkomórkowo zbudowane SA z kilku, najczęściej 5 podjednostek (łańcuchów polipeptydowych z których każdy ma 4 domeny transbłonowe) najlepiejpoznanym z tej grupy jest nACh (cholinergiczny receptor nikotynowy).
Porównaj kinetykę czasową potencjałów czynnościowych znanych Ci komórek (komórek nerwowych, mięśnia szkieletowego, mięśnia sercowego) oraz węzła zatokowo- przedsionkowego.
Cały przebieg potencjału czynnościowego dla komórki nerwowej jest rzędu kilku milisekund, dla komórki mięśnia szkieletowego rzędu kilkudziesięciu, a sercowego nawet kilkuset milisekund. Przebieg potencjału czynnościowego komórki mięśniowej serca różni się wyraźnie od potencjału czynnościowego kom. nerwowej lub mięśnia szkieletowego. Po gwałtownej depolaryzacji błona przepolaryzowuje się, tak że potencjał wewnętrznej strony błony przyjmuje wartość do +20 mV. Stan przepolaryzowania utrzymuje się przez dłuższy czas (ok. 150 ms), potencjał utrzymuje się na poziomie +20 - (-10) mV po czym spada i błona repolaryzuje się do stanu spoczynkowego. Potencjał czynnościowy węzła nie utrzymuje się na stałym poziomie, a stopniowo rośnie; gdy potencjał błonowy osiągnie wartość progu pobudliwości następuje depolaryzacja, po czym powrót do stanu wyjściowego. Węzeł zatokowo - przedsionkowy zachowuje się podobnie do układu wykonującego drgania relaksacyjne.
Omów jak przenosi się fala depolaryzacyjna wzdłuż włókna nerwowego
Depolaryzacja błony komórki nerwowej w jakimś miejscu przy nie zmienionym potencjale sąsiednich odcinków błony wywołuje przepływ prądu skierowany tak, aby wyrównać różnicę potencjałów. Powoduje to obniżenie potencjału sąsiedniego obszaru błony i wyzwala jej dalszą, samorzutną depolaryzację. W ten sposób wzdłuż włókna przemieszcza się fala zmian potencjału. W przypadku nerwu osłoniętego osłonką mielinową - prądy płyną skokowo od jednego przewężenia Ranviera do drugiego (zwiększa to szybkość przemieszczania).
Różnica miedzy synapsami aktywującymi i hamującymi
Zależnie od rodzaju przekaźnika (mediatora) zawartego w danej synapsie oraz rodzaju kanałów w błonie następnej komórki , przekazywany impuls może być albo aktywujący (zmniejszający potencjał komórki następnej) albo hamujący (podwyższający potencjał następnej komórki). Aktywacja lub hamowanie mają charakter impulsowy.
Synapsy pobudzające:
Dwa rodzaje receptorów glutaminianowych:
AMPA: alfa-amino-3-hydroksy-5-methyl-4-isoxazolepropionic acid
NMDA: N-methyl-D-sparatate
Odpowiedzi receptora AMPA i NMDA przy różnych wartościach potencjału. Separacja aktywności dwóch rodzajów receptora jest uzyskiwana blokadą receptora NMDA przez APV.
Synapsy hamujące:
Dwa rodzaje receptorów GABA (gamma-aminobutyric acid) (kwas gamma - aminomasłowy)
GABAA
GABAB
Odpowiedzi receptora AMPA, GABAA i GABAB
Zinterpretuj prawo ciągłości strumienia oraz prawo Bernoulliego i ich znaczenie
Prawo ciągłości strumienia cieczy - mówi, że przez każdy przekrój naczynia w tym samym czasie przepływa taka sama objętość cieczy, czyli przepływ jest stały i niezależny od przekroju naczynia.
Założenia:
Przepływ cieczy odbywa się w taki sposób, że wpływa tylko przez jeden koniec naczynia, a wypływa drugim.
Wewnątrz naczynia nie ma dodatkowych źródeł cieczy ani odpływów
Omów czynniki wpływające na lepkość krwi
Lepkość krwi zależy od liczby hematokrytowej, temperatury, przekroju naczynia, w którym płynie i od szybkości przepływu. Przy wzroście liczby hematokrytowej ponad 60% lepkość krwi znacznie wzrasta, wiąże się to z dużym zagęszczeniem krwinek tworzących często kolumny dysków. Lepkość krwi zwiększa się z obniżeniem temperatury. W temperaturze 0°C jest w około 2,5 raza większa niż przy 37°C.
Lepkość krwi w przewodach o średnicy większej od 0,3 mm nie zależy od powierzchni przekroju naczynia. Natomiast dla średnicy mniejszych zmniejsza się w miarę jak zmniejsza się przekrój. W przewodach cienkich większego znaczenia nabiera niejednorodność krwi i związana z nią nieciągła struktura cieczy (zawiesina krwinek). Zjawisko akumulacji osiowej polega na tym, że krwinki gromadzą się raczej bliżej osi naczynia unikając niejako jego ścian. Przy ścianach ciecz płynie wolniej, środkiem prędzej, w poprzek przekroju rury istnieje spadek prędkości. W skutek różnicy prędkości cieczy w poprzecznym przekroju naczynia krwinki zostają wprowadzone w ruch obrotowy. Ruch obrotowy krwinek pociąga za sobą ciecz nadając jej ruch cyrkulacyjny wokół krwinki, co jest powodem rozrzedzenia linii prądu przy odściennej stronie krwinki, a ich zagęszczenie od strony odosiowej. W przewodach o średnicy 0,1 do 0,2 mm lepkość zmniejsza się ze wzrostem szybkości, osiągając przy pewnej prędkości wartość stałą. Natomiast samo osocze (lepkość względna około 1,8) zachowuje się jak ciecz niutonowska. Przy prędkościach spotykanych w warunkach fizjologicznych krew zachowuje się jak ciesz newtonowska i lepkość jej od prędkości prawie nie zależy. Niemniej zjawisko akumulacji powoduje, że lepkość jest mniejsza, warstwa cieczy między ścianą a strumieniem krwinek zmniejsza bowiem tarcie.
Jakie znasz współczynniki lepkości krwi
Współczynnik lepkości bezwzględnej - równy liczbowo sile potrzebnej do podtrzymania ruchu warstw cieczy o przepływie laminarnym; jest to stosunek naprężenia stycznego do szybkości ścinania:
η = F/S : ^U/^X,
gdzie:
F/S- naprężenie styczne,
^X/^U- szybkość ścinania (gradient prędkości warstw).
^ - delta
Współczynnik lepkości kinetycznej:
ηkin =η / ρtemp
gdzie:
η- współczynnik lepkości bezwzględnej,
ρtemp - gęstość krwi w danej temperaturze.
Współczynnik lepkości względnej:
ηwzg= η / η0
gdzie:
η - współczynnik lepkości bezwzględnej,
η0 - współczynnik lepkości bezwzględnej wzorcowej cieczy (najczęściej wody).
Właściwy współczynnik lepkości
ηwłaściwy = 2,5 × η V/V0, gdzie:
η - współczynnik lepkości bezwzględnej,
V - objętość uwodnionej cząsteczki,
V0 - objętość nieuwodnionej cząsteczki.
Omów znaczenie czynnika geometrycznego dla przepływu krwi
Przepływem cieczy lepkich w naczyniach rządzi prawo Poiseuille'a, które wyraża się następującym wzorem:
^I= pr4/8ηl * ^P
gdzie:
^I - natężenie przepływu strumienia cieczy
r - promień przekroju poprzecznego naczynia
η - współczynnik lepkości bezwzględnej
l - długość naczynia
^P - różnica ciśnień na końcach przewodu
^-delta
W podanym wzorze pr4/8ηl - to odwrotność oporu naczyniowego przepływu R. W związku z tym:
R= 8/p * η * l/r4
gdzie:
l / r4 - czynnik geometryczny.
W naczyniach sztywnych R jest stały w związku z tym natężenie przepływu zależy tylko od różnicy ciśnień na końcach naczynia. W naczyniach sprężystych, takich jak tętnice i w pewnym stopniu żyły, ciśnienie, z
jakim krew działa na ściany naczyń powoduje ich chwilowe rozciągnięcie w związku z czym zmienia się promień przekroju poprzecznego tych naczyń. Ponieważ długość naczyń nie ulega zmianie, stosunek l / r4, nazywany czynnikiem geometrycznym ulega zmniejszeniu, a wraz z nim maleje opór naczyniowy R przepływu krwi. Wraz ze spadkiem oporu naczyniowego przepływu krwi rośnie natężenie przepływu strumienia krwi przesz naczynie. Natężenie przepływu w naczyniach sprężystych zależy więc nie tylko od różnicy ciśnień na końcach naczynia, ale również od czynnika geometrycznego dla tego naczynia. Należy pamiętać, że naczynia krwionośne działają ciśnieniem sprężystym ściany na krew, co uniemożliwia strumieniowy krwi nadmiernie odkształcić naczynie.
Scharakteryzuj fale tętna i tony serca
Tony serca - efekty akustyczne towarzyszące pracy serca, powstają w wyniku drgania zastawek wywołanego przez uderzenie w nie krwi podczas skurczu i rozkurczu serca. Wyróżnia się cztery tony serca, z których dwa są fizjologiczne (I i II) i występują u wszystkich ludzi, a dwa pozostałe, tzw. tony dodatkowe (III i IV) mogą występować w stanach patologicznych lub u osób zdrowych (szczególnie u dzieci).
ton pierwszy, skurczowy (systolityczny) jest wynikiem gwałtownego zamknięcia się zastawek oddzielających przedsionki serca od komór (zastawek przedsionkowo-komorowych). Ton skurczowy jest najlepiej słyszalny w piątej przestrzeni międzyżebrowej po obu stronach mostka.
ton drugi, rozkurczowy (diastoliczny) jest wynikiem zamknięcia zastawek oddzielających komory i tętnice (zastawek półksiężycowatych). Ton rozkurczowy jest najlepiej słyszalny w drugiej przestrzeni międzyżebrowej po obu stronach mostka.
ton trzeci powstaje podczas wypełniania się i rozszerzania komór serca podczas rozkurczu, jest najlepiej słyszalny na koniuszku serca. Występuje jako ton fizjologiczny u dzieci lub (częściej) w przypadku powiększenia prawej lub lewej komory.
ton czwarty powstaje podczas skurczu przedsionków
Tony serca badane są za pomocą stetoskopu, w celu stwierdzenia prawidłowej pracy serca.
Fala tętna
Rytmiczne skurcze tętna wprowadzają do układów tętniczych tak dużego, jak i płucnego, w odstępach czasu około 0,8s, takie same objętości krwi, około 70 cm3- pojemność wyrzutowa serca w spoczynku. Dzięki dużemu oporowi obwodowemu krew ta nie od razu zostaje włączona w obieg krążenia, lecz rozciąga podatne ściany aorty tak, że tuż za sercem tworzy się wybrzuszenie. Energia kinetyczna, którą krew otrzymała przy wyrzucie z serca zostaje przemieniona w energię potencjalną sprężystości odkształconej aorty. Siły sprężyste ścian naczynia przywracają mu w danym miejscu stan początkowy; przepychając porcję krwi powodują rozdęcie aorty w sąsiedztwie. W międzyczasie ponowny skurcz serca ponawia odkształcenie. W ten sposób odkształcenia sprężyste, wywoływane rytmicznie skurczami serca, przenoszą się ruchem falowym wzdłuż tętnic, aż zostaną stłumione w łożysku małych naczyń. Fala odkształceń sprężystych w ten sposób wywołana nosi nazwę fali tętna. Szybkość fali tętna wyliczona z równania ruchu falowego wyraziłaby się wzorem:
E - moduł z Younga ścian naczynia (zależy od ciśnienia i od wieku)
e - grubość ściany
- gęstość
r - promień przekroju
Półempiryczny wzór
F - współczynnik empiryczny dla aorty człowieka przy ciśnieniu rozkurczowym 70 mmHg wynosi 0,6-0,7
Prędkość fali tętna nie pokrywa się z prędkością przemieszczenia się krwi. Fala tętna rozchodzi się z prędkością około 5m/s do 8m/s, podczas gdy średnia prędkość krwi w aorcie jest na ogół mniejsza od 0,5m/s.
Długość fali tętna obliczona wzorem λ=cT, wynosi λ=5m/s*0,8s=4m. Wynika stąd, że amplituda odkształcenia znajdzie się „u progu” naczyń oporowych, gdzie zostanie stłumiona, zanim następne odkształcenie zostanie wywołane przez kolejny skurcz serca.
Tętnice odkształcają się stosunkowo dużym modułem sprężystości objętościowej. Duży opór
obwodowy pozwala w większych tętnicach utrzymać wysokie ciśnienie średnie. W związku z tym ściany tych tętnic są stale rozciągnięte i są magazynem energii potencjalnej sprężystości, który rytmicznie jest uzupełniany pracą serca. Ten magazyn energii potencjalnej sprężystości dużych tętnic nazywają powierzchnią, jego zadanie jest bowiem takie samo jak powierzchni w urządzeniach hydraulicznych. Zbiornik powietrza w takich urządzeniach amortyzuje wahania ciśnień wywoływanych działaniem pompy. Rolę tego zbiornika pełni układ tętniczy (ciśnieniowy) - odciąża pracę serca. Ściany żył- łatwo zmieniają objętość, pełnią rolę zbiornika pojemnościowego, w części żylnej mieści się 70% krwi.
Prawo Poiseuille'a.
Prawo Hagena-Poiseuille'a - prawo fizyczne opisujące zależność między strumieniem objętości cieczy a jej lepkością (która wynika z tarcia wewnętrznego), gradientem ciśnień (który jest bodźcem termodynamicznym powodującym przepływ płynu), a także wielkościami opisującymi wielkość naczynia (długość, promień przekroju poprzecznego).
Przy stacjonarnym (tj. niezmiennym w czasie), laminarnym przepływie nieściśliwego, lepkiego płynu w cylindrycznym przewodzie (tj. w rurze o stałym, kołowym przekroju), strumień objętości przepływu (objętość przepływającego płynu na jednostkę czasu) proporcjonalny jest do gradientu ciśnienia wzdłuż przewodu, a zatem i do różnicy ciśnień na końcach przewodu.
Praca i moc serca
Cykl pracy serca (cykl hemodynamiczny serca) jest indukowany przez układ bodźcoprzewodzący serca, który pobudza kardiomiocyty do skurczu w odpowiedniej kolejności wymuszając przepływ krwi. Na układ bodźcoprzewodzący wpływa impulsacja z układu autonomicznego regulując rytm serca i dostosowując go do aktualnych potrzeb ustroju.
Za początek cyklu pracy serca powszechnie przyjmuje się pauzę. W czasie pauzy przedsionki i komory serca są w stanie rozkurczu i krew pod wpływem gradientu (różnicy) ciśnień przelewa się z żył głównych i płucnych do przedsionków.
Następnie dochodzi do skurczu przedsionków, zwiększając ciśnienie w przedsionkach i powodując dopchnięcie jeszcze porcji krwi do komór, objętość komór po skurczu przedsionków nazywa się objętością późnorozkurczową, a ciśnienie panujące w komorach ciśnieniem późnorozkurczowym lub obciążeniem wstępnym.
Ciśnienie w komorach wzrasta powyżej ciśnienia w przedsionkach i następuje zamknięcie zastawek odpowiednio trójdzielnej po prawej i mitralnej po lewej stronie serca i uderzenie krwi o zastawki od strony komór. Zamknięcie zastawek wywołuje efekt akustyczny w postaci pierwszego tonu serca. Następnie rozpoczyna się skurcz komór nie powodujący zmiany objętości krwi zawartej w komorach jest to tzw. skurcz izowolumetryczny. W czasie skurczu izowolumetrycznego narasta napięcie ścian komór serca, co powoduje wzrost ciśnienia w komorach. Gdy ciśnienie przekroczy ciśnienie odpowiednio w pniu płucnym i aorcie następuje faza wyrzutu i pewna objętość krwi zostaje wypchnięta do pnia płucnego i aorty, jest to tzw. objętość wyrzutowa. Po fazie wyrzutu ciśnienie w komorach zaczyna spadać co powoduje zamknięcie zastawek pnia płucnego i aortalnej i wywołuje drugi ton serca.
W komorach po wyrzucie pozostaje zawsze pewna ilość krwi- jest to objętość późnoskurczowa a ciśnienie panujące w komorze nazywane jest ciśnieniem późnoskurczowym. Rozpoczyna się rozkurcz komór. W początkowej fazie rozkurczu ciśnienie w komorach jest jeszcze wyższe niż w przedsionkach i zastawki przedsionkowo-komorowe są zamknięte, ta faza rozkurczu nazywana jest rozkurczem izowolumetrycznym. Gdy ciśnienie w komorach spadnie poniżej ciśnienia w przedsionkach zastawki otwierają się i krew przelewa się z przedsionków do komór i cały cykl powtarza się.
Praca serca to inaczej praca zewnętrzna, związana bezpośrednio z uruchomieniem krwi. Składa się na nią praca wykonana przeciw ciśnieniu panującemu w aorcie i tętnicy płucnej oraz praca związana z nadaniem krwi energii kinetycznej.
Omów równanie fali głosowej i podstawowe jej parametry
Fala głosowa jest falą podłużną. W najprostszym przypadku fali harmonicznej prostej każdy punkt ośrodka objętego ruchem falowym tzw. pola akustycznego, wykonuje drgania harmoniczne proste, równolegle do kierunku rozchodzenia się fali, z tym, że im dalej od źródła fali, tym bardziej opóźnione są fazy drgań. Jeżeli fala rozchodzi się w danym ośrodku z prędkością c, to zanim ona dotrze do punktu odległego o x od źródła, upłynie czas t=x/c. Wychylnie y punktu odległego o x od źródła jest w chwili t takie samo jak wychylenie źródła (x=0) w chwili wcześniejszej t - x/c.
Jeżeli źródło drga według wzoru y=A sin ωt, to punkt odległy o x od źródła drga według:
- równanie fali i może wyglądać też tak:
ωx/c -wyraża opóźnienie fazy punktu x w stosunku do źródła.
Wprowadzając długość fali λ jako drogę, którą fala przebędzie w ciągu jednego okresu T,
czyli:
λ = cT albo λ =c/v
T - okres periodyczności czasowej
λ - okres periodyczności przestrzennej, czyli odległość między dwoma punktami fali,
których fazy różnią się o 2 Π
Prędkość fali wyraża się zależnością

Co rozumiesz przez ciśnienie fali, gęstość energii i natężenie dźwięku
Fala głosowa rozchodzi się jako fala ciśnień, towarzyszy jej zagęszczanie i rozrzedzanie ośrodka. W czasie t i w odległości x od źródła drgającego panuje ciśnienie:
P = Pm cosω (t - x/c )
ၵ = ω A cos(ωt + φ)
Pm = c ၵ
ၵ - prędkość akustyczna
c - prędkość rozchodzenia się fali głosowej (prędkość fazowa)
Natężenie dźwięku
Dla oceny energii przenoszonej przez falę dźwiękową wprowadza się natężenie dźwięku (fali) I. Natężenie dźwięku definiuje się stosunkiem mocy P, przenoszonej przez falę do powierzchni S ustawionej prostopadle do kierunku rozchodzenia się fali
I=P/S
P - moc przenoszona przez falę
S - pole powierzchni prostopadłej do kierunku rozchodzenia się
I = G c
[I] = [J/m2 s] = [W/m2]
Gęstość energii G=E/V
Energia zawarta w określonej objętości pola akustycznego
Oporność akustyczna fali i współczynnik odbicia
Wielkość Z- analogiczna do oporności elektrycznej- wyraża tzw. oporność akustyczną przewodu przekroju S przenoszącego falę- bywa też nazywana impedancją akustyczną. Oporności akustyczne odgrywają ważną rolę przy przechodzeniu fali z jednego ośrodka do drugiego. Na granicy ośrodków zachodzi częściowo odbicie fali.
Współczynnik odbicia, wyrażający się stosunkiem natężenia I, fali odbitej do natężenia I0 fali padającej, gdy fala pada prostopadle na płaszczyznę odbijającą, można przedstawić zależnością:
Ir/I0=(ρ1c1 -ρ2c2/ρ1c1+ ρ2c2)2
Fala przechodzi z jednego ośrodka do drugiego bez odbicia, jeżeli oporności właściwe fali będą równe, czyli ρ1c1= ρ2c2
Oporność akustyczna:
Z =ρc/S
Scharakteryzuj granice słyszalności, próg czułości, próg bólu i próg zmiany
Granice słyszalności 16 Hz - 20 000 Hz
Drgania o częstotliwości niższej niż 16 Hz- infradźwięki, jak i o częstotliwości wyższej od 20 000 Hz- ultradźwięki- nie są słyszalne przez ucho ludzkie.
Próg zmiany ( najmniejsza dostrzegalna zmiana częstotliwości) ( od 0,3% do 1%) Najniższy dla częstotliwości w zakresie 500 Hz do 4 000 Hz
Próg zmiany częstotliwości tj. najmniejsza dostrzegalna zmiana częstotliwości, jest dla częstotliwości w granicach 500- 4000 Hz w przybliżeniu stały i wynosi ok. 0,3%. Oznacza to, że ucho jest zdolne zauważyć zmianę wysokości, jeżeli częstotliwość słyszanego tonu zmieni się o 0,3% np. ton 1000 Hz musiałby się zmienić o 3Hz, żeby zauważona została zmiana wysokości. Przy częstotliwościach niższych i wyższych próg zmiany częstotliwości jest wyższy. Dla tonu 8000 Hz wynosi w przybliżeniu 1%, tyleż samo wynosi dla tonu 50 Hz, zależy jednak w znacznym stopniu także od natężenia.
Próg bólu - wartość ciśnienia akustycznego, przy której ucho odczuwa wrażenie bólu. Jest ona słabo zależna od częstotliwości i wynosi 140 dB dla dźwięków sinusoidalnych oraz 120 dB dla szumów. Próg bólu dla różnych częstotliwości dźwięku wyznacza krzywa progowa ograniczająca od góry obszar słyszalności.
Wrażenie bólu wywołane jest reakcją mięśni bębenka i kosteczki ucha środkowego na impulsy wysokiego ciśnienia akustycznego. Reakcja ta ma na celu ochronę aparatu słuchowego przed ewentualnymi uszkodzeniami.
Ból działa obezwładniająco, toteż emitery dźwięku o natężeniu przekraczającym próg bólu stosowane są m.in. w służbach porządkowych podobnie jak armatki wodne.
Czułość słuchu, próg czułości słuchu, próg słyszalności, zdolność słuchu do wykrywania najcichszych dźwięków- dla różnych częstotliwości dźwięku wyznacza krzywa progowa ograniczająca od dołu obszar słyszalności. Przez czułość słuchu rozumiemy również czułość różnicową, czyli zdolność do wykrywania najmniejszych różnic wysokości, głośności i barwy dźwięku (progi różnicy).
Jaką funkcje pełni ucho zewnętrzne
Kierunek dźwięku:
z przodu lub z tyłu - sekwencja czasowa wywołana bezpośrednim i wtórnym pobudzeniem kanału słuchowego zmienia się,
z lewej lub z prawej strony - przesunięcie czasowe pomiędzy sygnałami (nie większe niż 500 ms)
Ucho zewnętrzne stanowi komorę o częstotliwości rezonansowej ok. 2,5 kHz, wzmacniającą dźwięki o ω w zakresie 2-5 kHz do 15-20 dB
Kanał słuchowy - natężenie drgań słupa powietrza maksymalne gdy l równa się 4 długościom kanału
l = c/ω, czyli ωmax = c/4, to jest ωmax = 3173 Hz
Dlatego dla tej długości jest najniższy próg czułości, a efekt wzmocnienia wynosi 15 dB i wynika z długości kanału oraz z tego, że l jest porównywalna z rozmiarami czaszki
Jaka funkcje pełni ucho środkowe
Ucho środkowe służy zwiększeniu ciśnienia wywieranego przez podstawę strzemiączka na płyn ślimakowy w stosunku do ciśnienia, które działa na błonę bębenkową. Jest to możliwe dzięki temu, że powierzchnia błony bębenkowej, która dotyka pierwszej kosteczki słuchowej - młoteczka, jest ok 17 razy większa niż błona okienka owalnego (ta styka się z ostatnią kostką - strzemiączkiem). Pomaga w tym również układ młoteczek kowadełko, będący klasyczną dźwignią o ramionach różnej długości, gdyż ramię młoteczka jest ok 1,2 - 1,4 raza dłuższe niż ramię kowadełka. Te czynniki powodują, że wzmocnienie dźwięku w uchu środkowym wynosi ok 22 razy. Dla dźwięków 70-90 dB występuje odruch strzemiączkowy, prowadzący do obniżenia natężenia dźwięku. W tym pomagają mięśnie naprężacz błony bębenkowej i strzemiączkowy. Ich napięcie ogranicza drgania kosteczek słuchowych i zmniejsza ich amplitudę. Do ucha środkowego należy także trąbka Eustachiusza. Ta służy do wyrównania ciśnienia w jamie ustnej z ciśnieniem na zewnątrz organizmu.
Jest to otwór łączący jamę bębenkową z gardłem, jego światło otwiera się podczas ziewania, połykania i wymawiania głosek u e i p k. Niedrożna trąbka Eustachiusza upośledza słyszenie, natomiast stale otwarta sprawia, że głośno słyszymy nasz własny głos.
Scharakteryzuj teorie Helmholtza analizy dźwięku w uchu wewnętrznym
Teoria rezonansowa Helmholtza (1863), inaczej zwana teorią harfy, zakłada iż błona podstawna zbudowana z cienkich włókien (strun słuchowych), krótszych u podstawy ślimaka i reagujących na składowe wysokoczęstotliwościowe oraz dłuższych, na końcu ślimaka reagująca na składowe niskoczęstotliwościowe. Każde z włókien nastrojone jest na określoną częstotliwość rezonansową dzięki czemu odpowiada tylko wybranej składowej zawartej w docierającym do słuchacza sygnale. Włókna po rozedrganiu pobudzają właściwe sobie komórki nerwowe ślimaka, dzięki czemu powstają impulsy elektryczne, które dalej przekazywane są do ośrodka słuchowego w mózgu. Teoria ta pomija udział wyższych pięter drogi słuchowej w procesie analizy dźwięku.
Scharakteryzuj teorie Bekesego
Teoria wędrującej fali (George von Bekesy) mówi o tym, że powstała w cieczach ślimaka błona hydrodynamiczna wytwarza w błonie podstawnej falę wędrującą z wyraźnie zaznaczonym maksimum wychylenia. Największe wychylenie, w zależności od częstotliwości padającego dźwięku przypada na różne miejsca w błonie podstawnej. Przemieszcza się ona ku szparze osklepka.
Rola błony podstawnej w analizie dźwięku
Błona podstawna znajduję się wewnątrz ślimaka. Wprawiana jest ona w ruch przez ruch cieczy znajdującej się w ślimaku. Na niej znajduje się narząd spiralny, gdzie zachodzi wstępna analiza dźwięku oraz zmiana sygnału mechanicznego w ciąg impulsów bioelektrycznych.
Rola komórek zmysłowych wewnętrznych i zewnętrznych w analizie dźwięku
Narząd Cortiego, narząd spiralny - właściwy narząd słuchu znajdujący się w ślimaku w przestrzeni
zwanej schodami środkowymi (przewód ślimakowy). Rozciąga się on wzdłuż przewodu ślimakowego poza jego częścią zwaną kątnicą przedsionkową. Położony jest na błonie podstawnej. Przypomina "wał", który po swojej wewnętrznej stronie zwrócony jest w kierunku bruzdy (blaszki) spiralnej wewnętrznej. Przyśrodkowo od narządu znajduje się rąbek spiralny.
Składa się on z dwóch rodzajów komórek:
komórki zmysłowe - są to komórki rzęsate (zwane inaczej komórkami słuchowymi, rzęskowymi lub włoskowatymi). Zgrupowane są one w rzędy: komórki rzęsate wewnętrzne - 1 rząd i komórki rzęsate zewnętrzne 3 rzędy.
komórki tworzące zrąb narządu, które pełnią między innymi funkcje "szkieletu" utrzymującego komórki rzęsate we właściwym położeniu:
komórki filarowe wewnętrzne i zewnętrzne tzw. filary. Oba filary są nachylone ku sobie w części górnej i łącząc się wierzchołkami ograniczają tzw. tunel wewnętrzny (Cortiego) będący trójkątnego kształtu. Wypełniony jest on płynem zbliżonym składem do przychłonki zwanym kortylimfą lub chłonką Cortiego (trzecią chłonką)
komórki falangowe wewnętrzne i zewnętrzne, (komórki Deitersa), są to komórki podporowe, na których spoczywają komórki rzęsate. Pomiędzy filarami zewnętrznymi a komórkami falangowymi zewnętrznymi znajduję się przestrzeń Nuela (przestrzeń przytunelowa), spiralny kanał mający połączenie z tunelem Cortiego przez szczeliny pomiędzy filarami zewnętrznymi.
komórki graniczne wewnętrzne, (komórki Helda)
komórki graniczne zewnętrzne, (komórki Hensena)
komórki podporowe wewnętrzne i zewnętrzne, (komórki Claudiusa)
Najbardziej bocznie w narządzie Cortiego położony jest tunel zewnętrzny, wypełniony także kortylimfą. Tuż zanim znajduję się rowek spiralny zewnętrzny. Nad komórkami rzęsatymi wewnętrznymi i zewnętrznymi znajduje się błony pokrywowej, galaretowata masa rozpoczynająca się od rąbka spiralnego i pokrywająca także bruzdę spiralną wewnętrzną. Odcinek zewnętrzny błony jest znacznie grubszy od wewnętrznego. Składa się ona z włókien zanurzonych w bezpostaciowej masie zawierającej mukopolisacharydy i glikoproteiny. Na jej dolnej powierzchni odpowiadającej położeniu komórek rzęsatych wewnętrznych znajduję się rowek zwany pasmem Hensena.
Źródła promieniowania jonizującego
NATURALNE ŹRÓDŁA PROMIENIOWANIA JONIZUJĄCEGO
•promieniowanie kosmiczne (w wyniku którego powstają głównie tryt, beryl i izotop węgla 14C)
•promieniowanie emitowane przez pierwiastki zawarte w skorupie ziemskiej, a w konsekwwnie izotop potasu 40K), oraz radon uwalniany ze ścian budynków, wody i naturalnych paliw podczas ich spalania.
SZTUCZNE ŹRÓDŁA PROMIENIOWANIA JONIZUJĄCEGO
•aparatura rentgenowska - diagnostyczna i terapeutyczna oraz przemysłowa, a także kineskopy telewizyjne, prostowniki próżniowe średnich i wielkich mocy, betatrony, akceleratory
•izotopy promieniotwórcze - "bomby" kobaltowe lub cezowe, igły radowe, mierniki lub czujniki stosowane w przemyśle (gęstościomierze, pojemnościomierze, wagi izotopowe, czujniki dymu itp.), znakowanie radioizotopowe
Najczęściej spotykanymi rodzajami promieniowania jonizującego elektromagnetycznego jest promieniowanie X i promieniowanie gamma.
Promieniowanie jonizujące jest nieodłącznym składnikiem naszego środowiska, mającym doniosły wpływ na rozwój i ewolucje życia na Ziemi.
Promieniowanie to powstaje w wyniku szeregu procesów:
promieniowania kosmicznego, będącego w istocie rozpędzonymi do ogromnych energii protonami, jądrami helu, a także innymi atomami docierającymi nieustannie z Kosmosu oraz z atmosfery Słońca, które zderzając się z atomami atmosfery ziemskiej prowadzą do powstawania wtórnych źródeł promieniowania,
samorzutnego rozpadu niestabilnych atomów promieniotwórczych, wchodzących w skład naszego naturalnego otoczenia,
rozpadu promieniotwórczych pierwiastków wprowadzonych do naszego otoczenia w sposób niekontrolowany, np. w wyniku prób z bronią jądrową, czy też katastrof w energetyce jądrowej,
rozpadu paliwa jądrowego w reaktorach jądrowych lub przyspieszania cząstek do dużych prędkości w akceleratorach,
przy przejściach elektronów na wewnętrzne powłoki elektronowe atomu, powstające wówczas promieniowanie rentgenowskie wykorzystywane jest w technice i diagnostyce medycznej.
Wielkości opisujące dawki promieniowania jonizującego (Grey, REM)
Do określania natężenia promieniowania i jego dawki na jaką jest narażony człowiek służy wiele jednostek, niektóre z nich to:
W fizyce i chemii podstawową jednostką charakteryzowania promieniowania jest energia ich cząstek mierzona eV (elektronowoltach). 1eV=1,602*10-19 J. Jednakże do określania skutków działania promieniowania na organizmy żywe nie wystarczy tylko podanie energii cząstek, na działanie których był narażony.
Jednostki natężenia promieniowania.
Jednostki natężenia promieniowania to jednostki określające aktywność promieniotwórczą, poprzez podanie intensywności jej promieniowania (gęstości strumienia).
1 Ci - jeden kiur, nazwa tej jednostki pochodzi od nazwiska Marii Skłodowskiej-Curie. Za wzorzec ilościowy przyjęto intensywność promieniowania jednego grama czystego izotopu radu-266, w którym zachodzi 3,7*1010 rozpadów promieniotwórczych na sekundę. Kiur nie jest wprawdzie jednostką układu SI, ale jest niekiedy używany, zatem warto pamiętać co oznacza.
Stosowaną obecnie i umieszczoną w układzie SI jednostką intensywności promieniowania jest Bekerel: 1Bq. Aktywność jednego bekerela posiada ciało w którym na jedną sekundę dochodzi do jednego aktu rozpadu promieniotwórczego. Nazwa ta pochodzi od nazwiska odkrywcy promieniowania: Henriego Becquerela. Aktywność jednego kiura jest równa aktywności 3,7*1010 bekerela.
Jednostki dawki pochłoniętej energii
Jednostki dawki pochłoniętej energii określają jaką energię promieniowania pochłonęło ciało o danej masie.
W układzie SI jednostką dawki pochłoniętej energii promieniowania jonizującego przez materię jest jeden Grej (Gy). 1Gy=1J/1kg.
Dawną jednostką jest rad 1Gy=100rad=0,01J/kg=100erg/g (erg-jednostka pracy ciepła i energii w układzie CGS (układ jednostek fizycznych oparty na centymetrze, gramie i sekundzie) 1erg=10-7J
Jeszcze jedną jednostką określającą oddziaływanie promieniowania z materią jest LET, określa stratę energii danej cząstki w stosunku do przebytej przez nią drogi. LET zależy nie tylko od rodzaju promieniowania, ale również od zdolności hamowania ośrodka absorbującego. LET to skrót od angielskich słów: linear energy transfer, które oznaczają: liniowy współczynnik przenoszenia energii.
Jednostki dawki ekspozycyjnej
Dawka ekspozycyjna mówi nam o ładunku który został wytworzony przez promieniowanie jonizujące w jednostce masy danego ciała. Obecnie używaną i umieszczoną w układzie SI jednostką dawki ekspozycyjnej jest kulomb na kilogram (C/kg). Dawniej używaną jednostką jest rentgen (1R=2,58*10-4 C/kg).
Moc dawki
Moc dawki promieniowania to stosunek wartości tej dawki do czasu w którym została otrzymana. Moc dawki jest to wielkość opisująca szybkość napromieniowania.
Moc dawki pochłoniętej: Gy/s lub Gy/h, albo rad/h.
Moc dawki ekspozycyjnej:A/kg albo R/h. 1R/h=7,166 · 108 A/kg.
Jednostki dawki równoważnej
Konsekwencje jakie będzie miał dla nas kontakt z promieniowaniem zależą nie tylko od tego na jaką dawkę promieniowania byliśmy narażeni, ale również jaki był jego rodzaj i jaka tkanka została napromieniowana.
Do określania biologicznych skutków napromieniowania ożywa się tzw. dawki równoważnej mierzonej w siwertach (Sv), lub dawniej w remach (rem).
W związku z tym, że nie każde promieniowanie działa równie destrukcyjnie na organizmy żywe do porównywania dawki pochłoniętej i dawki równoważnej używa się tzw. czynnika jakości promieniowania
QF - quality factor (współczynnik skuteczności biologicznej). Np. dla promieniowania gamma 1Sv=1Gy. Dla innych rodzajów promieniowania, działających o wiele silniej na organizmy żywe współczynnik QF przyjmuje wartości wiele większe (np. promieniowanie alfa QF=25). Sv=Gy*QF, rem=rad*QF, 1Gy=100radów, zatem 1Sv=100remów.
Stadia radiacyjne
Etapy radiolizy wody
Radioliza - całokształt przemian chemicznych zachodzących w obiekcie w wyniku pochłonięcia promieniowania jonizującego. Odmianą radiolizy jest radioliza impulsowa dotycząca skali czasowej rzędu mikrosekund.
Radioliza to proces chemiczny wywołany działaniem cząstek i promieniowania wysokoenergetycznego.
Etapy radiolizy
I - akt pierwotny-proces przekazania energii cząsteczkom ośrodka 10-18 - 10-16 s.
II - reakcje wtórne między produktami 10-16 - 10-4 s. Dyfuzja powstałych produktów poza obszar tunelu jonizacji, wzbudzenie cząsteczek.
III- reakcje jonów i cząsteczek o energii termicznej 10-4 - 10-1 s.
Przykładem radiolizy jest radioliza wody.
Do najważniejszych reakcji radiacyjnych należy radioliza wody. Po pochłonięciu kwantu promieniowania (hv) przez cząsteczkę wody następuje jej jonizacja:
H2O + hv → H2O+ + e-
Jony H2O+ ulegają rozpadowi:
H2O+ → H+ + OH
a elektrony - hydratacji i następnie rozkładowi:
H2O + e- → H2O- → H + OH-
Gdy elektron nie zdoła oddalić się poza obszar elektrostatycznego oddziaływania jonu macierzystego H2O+, ulega wychwytowi i wówczas cząsteczka wody (H2O*), która rozpada się na rodniki:
H2O+ + e- → H2O* → H + OH
Rodniki OH i atomy wodoru rekombinują:
OH + OH → H2O2 oraz H + H → H2.
W rezultacie otrzymuje się bardzo reaktywne rodniki OH i H oraz cząsteczki H2O2 i H2, które mogą reagować z substancjami znajdującymi sie w wodzie. Skutki chemiczne zależą od wielkości pochłoniętej przez układ dawki promieniowania. Dawką nazywa się ilość pochłoniętej energii przez określoną masę substancji (jednostką jest grej: 1 Gy = 1J/1kg).
Rozkład wody pod wpływem cząstek α zaobserwowała Maria Skłodowska-Curie podczas pracy z roztworami soli radu. Ponieważ produktami tej reakcji były wodór i tlen, proces ten nazwała radiolizą, przez analogię do elektrolizy wody, której produktami są także wodór i tlen. Obserwacja ta dała początek chemii radiacyjnej. W odróżnieniu od reakcji fotochemicznej, o których przebiegu decydują rodzaj i charakter wiązań między atomami w cząsteczce, w reakcjach radiacyjnych dominuje proces jonizacji, zależny przede wszystkim od gęstości ośrodka.
Zmiatacze wolnych rodników
Zmiatacze wolnych rodników, akceptory wolnych rodników, związki chemiczne łatwo wchodzące w reakcję z wolnymi rodnikami, w której wyniku powstają wolne rodniki zmiataczy, o mniejszej reaktywności.
Przykłady: cysteina, witamina C, witamina E, jod, chinony, jony NO2-, Fe3+.
Niektóre zmiatacze wolnych rodników pełnią funkcję ochronną w komórkach organizmów żywych podczas procesów utleniania i proteolizy. Stosowane są w badaniach reakcji wolnorodnikowych w fotochemii, chemii radiacyjnej i biochemii.
Typy reakcji rodnikowych
Reakcje rodnikowe i ich przebieg:
Reakcje rodnikowe nie są tak powszechne jak reakcje polarne, są jednak bardzo ważne w chemii organicznej, szczególnie w przypadku pewnych procesów przemysłowych. Z procesami z udziałem rodników spotykamy się praktycznie na każdym kroku. Procesy spalania oraz procesy zachodzące w atmosferze oraz środowisku naturalnym i prowadzące do ich zanieczyszczenia, procesy zachodzące w układach biologicznych towarzyszące stanom chorobowym oraz procesom starzenia się organizmów, to tylko niektóre z przykładów, których jesteśmy świadkami lub doświadczamy ich osobiście.
Historycznie, pierwszy rodnik został zidentyfikowany przez Gomberga w roku 1900 w formie trójwartościowego związku, trifenylometylu Ph3C. Trzeba było jednak czekać prawie 40 lat, aby reakcje rodnikowe były po raz pierwszy wykorzystane przez chemików w syntezie chemicznej. W kolejnych dekadach XX wieku udowodniono istotny udział reakcji rodnikowych w układach biologicznych ora ich znaczenie w medycynie.
Chociaż większość rodników jest elektrycznie obojętna, są one bardzo reaktywne, ponieważ zawierają atom z nieparzysta liczba elektronów na jego powłoce walencyjnej zamiast trwałego oktetu gazu szlachetnego. Oktet elektronowy na powłoce walencyjnej może powstać na wiele sposobów. Na przykład: rodnik może oderwać atom (albo grupę atomów) z innej cząsteczki, tworząc w konsekwencji obojętna cząsteczkę i nowy rodnik. Sumaryczny wynik jest reakcją substytucji rodnikowej.
Reakcje substytucji rodnikowej przebiegają zwykle w trzech etapach: inicjacji (zapoczątkowania), propagacji (kontynuowania) oraz terminacji (zakończenia).
Inicjacja - Etap ten rozpoczyna reakcję tworzenia reaktywnych rodników. Rozważmy przypadek reakcji rodnikowej - chlorowanie metanu. Stosunkowo słabe wiązanie Cl-Cl ulega homolitycznemu rozerwaniu pod wpływem naświetlania światłem UV. Tworzą się dwa reaktywne rodniki chlorowe i następuje kolejny etap.
Cl−Cl hν→ 2 Cl•
Propagacja - Gdy w środowisku reakcji pojawi się niewielka liczba rodników chlorowych, zachodzi etap propagacji (wydłużenia łańcucha). Gdy rodnik chlorowy reaguje z cząsteczką metanu, odrywa od niego atom wodoru i tworzy cząsteczkę HCl oraz rodnik metylowy. Rodnik metylowy reaguje następnie w drugim etapie propagacji z cząsteczką Cl2, tworząc trwały produkt - chlorometan, i nowy rodnik chlorowy, który ponownie uczestniczy w pierwszym etapie propagacji.
CH4 + Cl• → CH3• + HCl
CH3• + Cl2 → CH3Cl + Cl•
Gdy sekwencja reakcji zostanie zapoczątkowana, reakcja przebiega przez powtarzające się etapy, co stanowi o charakterze tej reakcji - jest to reakcja łańcuchowa.
Terminacja (zakończenie reakcji) - Niekiedy dwa rodniki mogą połączyć się ze sobą i utworzyć trwały produkt. Gdy do tego dojdzie cykl reakcyjny zostaje zerwany i łańcuch reakcyjny się kończy. Taka terminacja zachodzi jednak rzadko, ponieważ stężenie rodników w dowolnym momencie reakcji jest bardzo małe i prawdopodobieństwo, że dwa rodniki zderzą się ze sobą jest również niewielkie.
Cl• + Cl• → Cl2
CH3• + Cl• → CH3Cl
CH3• + CH3• → C2H6
Przedstawiona reakcja jest reakcją rodnikową, ponieważ zachodzi według mechanizmu wolnorodnikowego. Jest reakcją fotochemiczną, ponieważ w fazie inicjacji wymagany jest dostęp światła. Jest też reakcją łańcuchową, ponieważ procesy propagacji napędzają się wzajemnie powtarzając się cyklicznie. I w końcu jest to reakcja substytucji, czyli podstawienia, bo jeden atom jest wymieniany na inny.
To jednak nie koniec. Kto zabroni rodnikowi chlorowy może podstawić także chlorometan. Mogą więc zachodzić dalsze podstawienia.
CH3Cl + Cl• CH2Cl• +HCl
CH2Cl• + Cl2 CH2Cl2 +Cl•
CH2Cl2 + Cl• CHCl2• + HCl
CHCl2• +Cl2 CHCl3 + Cl•
CHCl3 + Cl• CCl3• + HCl
CCl3 + Cl2 CCl4 + Cl•
Z chloru i metanu powstają: CH3Cl, CH2Cl2, CHCl3, CCl4 , a także produkty uboczne HCl w tym wyższe węglowodory jak C2H5 oraz ich chloropochodne, oczywiście w śladowych ilościach.
Opisana reakcja substytucji rodnikowej stanowi jedynie jeden z wielu różnych procesów, w których mogą uczestniczyć rodniki. Jednak podstawa, na której opierają się wszystkie reakcje rodnikowe jest taka sama. Wiązania ulegają rozerwaniu lub powstają przy udziale rodników o nieparzystej liczbie elektronów.
Alternatywnie, rodnik może przyłączać się do alkenu, wykorzystując jeden elektron wiązania podwójnego alkenu, co prowadzi do powstania nowego rodnika. Sumaryczny wynik jest reakcją addycji rodnikowej.
Najczęściej rodniki powstają w wyniku reakcji oderwania atomu wodoru od cząsteczki związku organicznego. Jeżeli następuje oderwanie pierwszorzędowego atomu wodoru, to spowoduje powstanie pierwszorzędowego rodnika, oderwanie drugorzędowego atomu wodoru powoduje utworzenie rodnika drugorzędowego itd. Otóż zauważono, że względna łatwość odrywania się atomu wodoru, przedstawia się następująco.
3o > 2o > 1o > CH3
I odpowiednio, trwałość wolnych rodników jest następująca
3o > 2o > 1o > CH3
Z tego wynika, że im trwalszy jest wolny rodnik, tym łatwiej się tworzy. Takie uogólnienie jest szczególnie użyteczne, ponieważ trwałość rodników decyduje o orientacji i reaktywności podczas wielu reakcji, w których powstają wolne rodniki.
Ilościowa charakterystyka bezpośredniego działania promieniowania jonizującego (promienioczułość)
Zasadnicza ilościowa charakterystyka rażącego działania promieniowania jonizującego podczas przebywania i działania w terenie skażonym substancjami promieniotwórczymi lub w strefie wybuchu jądrowego. Stopień rażącego działania dawki promieniowania na organizm ludzki zależy od jej wielkości, wyrażonej w rentgenach. Jednorazowa dawka promieniowania wielkości 50 R nie powoduje ujemnych skutków. Dawka promieniowania 50 - 100 R wywołuje zmiany we krwi i pierwsze objawy choroby popromiennej, lecz nie powoduje utraty zdolności bojowej. Dawka promieniowania 100 - 200 R wywołuje chorobę popromienną, wskutek której część porażonych może utracić zdolność bojową na kilka dni lub tygodni. Dawka promieniowania 200 - 400 R powoduje chorobę popromienną, długotrwałą utratę zdolności bojowej, a nawet wypadki śmiertelne. Wielkość dawki promieniowania mierzy się za pomocą dozymetrów.
Ilościowa charakterystyka pośredniego działania promieniowania jonizującego
Działanie promieniowania jonizującego na rozcieńczone roztwory białek sprowadza się do reakcji produktów radiolizy wody z rozpuszczonymi cząsteczkami. Zakładamy, że liczba powstających rodników wody jest proporcjonalna do dawki promieniowania, a rodniki te nie oddziałują ze sobą. Rozpatrujemy zatem 2 przypadki:
A. cząsteczki rozpuszczonej substancji wchodzą tylko jednorazowo w reakcję z rodnikami wody - liczba zmienionych cząstek jest liniową funkcją dawki:
(dawkę, przy której uległy przekształceniu wszystkie cząsteczki substancji rozpuszczonej B. drugi przypadek odnosi się do biopolimerów, które mogą wchodzić w reakcje z rodnikami wody wielokrotnie, tzn. produkty radiolizy wody reagują z jednakowym p-po zarówno ze zmienionymi, ja i z nie zmienionymi cząsteczkami białka.
Jeżeli zmiana wywołana jest pojedynczym efektywnym aktem oddziaływania, to ilość zmienionych makrocząsteczek w funkcji dawki opisana jest wzorem:
D37 - dawka powodująca inaktywację 63% cząstek
Rodniki aminokwasowe i białkowe
Rodniki (lub wolne rodniki) - atomy lub cząsteczki posiadające jeden lub więcej wolnych (niesparowanych) elektronów, czyli posiadające spin elektronowy różny od 0. Rodniki powstają np. na skutek homolitycznego rozpadu wiązań chemicznych, tj. takiego, w wyniku którego przy obu częściach rozerwanej cząsteczki zostaje po jednym z elektronów, które wcześniej tworzyły wiązanie. Może ono następować pod wpływem naświetlania promieniowaniem ultrafioletowym, promieniowaniem rentgenowskim, przez bombardowanie elektronami, w wyniku niektórych reakcji redoks, a także w wyniku termicznego rozpadu. Rodniki są zwykle obojętne elektrycznie i zazwyczaj bardzo reaktywne. W "typowych" reakcjach z udziałem rodników ich stężenie w mieszaninie reakcyjnej jest zwykle dość niskie, ze względu na ich dużą reaktywność.
NIE WIEM O CO CHODZI W TYM PYTANIU W SUMIE! Może o to, że w wyniku powstawania rodników dochodzi w jakimś stopniu to zniszczenia struktury białek, rodniki aminokwasów mogą łączyć się ze sobą tworząc różne, nowe białka- nie spełniające swojej docelowej funkcji…
Czynniki wpływające na promienioczułość
Promieniowrażliwość komórki zależy od wielu czynników:
właściwej promieniowrażliwości komórki,
jednorodności populacji komórek,
zdolności do naprawy uszkodzeń popromiennych,
fazy w cyklu komórkowym (wieku komórki),
stopnia jej utlenowania,
stopnia jej uwodnienia.
Wrażliwość komórek na działanie promieniowania jonizującego jest tym większa, im większa jest ich aktywność podziałowa i tym mniejsza, im większy stopień ich zróżnicowania.
Promienioczułość a pojęcie tarczy
Promieniowrażliwość - wrażliwość komórek na promieniowanie, czyli odsetek komórek, które giną po podaniu dawki testowej promieniowania.
Promienioczułość, promieniowrażliwość, wrażliwość biologiczna komórek i tkanek na działanie promieniowania jonizującego; wysoką p. wykazuje układ krwiotwórczy (szpik, węzły chłonne, śledziona) i nabłonek płciowy gonad, jak też nowotwory z nich się wywodzące (wskazanie do radioterapii), niską — tkanki układu nerwowego i tkanki podporowe
W każdym organizmie istnieje kilka wrażliwych objętości lub tarcz, w których musi nastąpić odpowiednia liczba aktów adsorpcji (trafień) by efekt popromienny mógł się ujawnić. Liczba tarcz danej struktury nie jest znana, zniszczenie promieniowaniem jednej tarczy odpowiedzialnej za określoną funkcję powoduje wypadnięcie tej funkcji, np. komórka traci zdolność podziału.
Zatem oba te terminy są ze sobą w pewnym stopniu powiązane.
Podstawy efektu tlenowego

EFEKT TLENOWY
- Bardzo istotnym czynnikiem modyfikującym wrażliwość nowotworu na energię promienistą jest stopień utlenowania guza. Zjawisko to nosi nazwę efektu tlenowego: wrażliwość nowotworu na radioterapię jest tym większa, im lepiej jest on utlenowany. Iloraz dawki promieniowania jonizującego, wymaganej do zniszczenia komórek pozostających w niedotlenieniu, do dawki wywołującej analogiczny efekt przy prawidłowym utlenowaniu nosi nazwę współczynnika wzmożenia tlenowego (WWT). Ma on większą wartość w wypadku stosowania promieniowania elektromagnetycznego (fotonowego). Oznacza to, że hipoksja ma wówczas istotniejsze znaczenie.
Zmiany popromienne w kwasach nukleinowych
Promieniowanie jonizujące wnikające do ustroju żywego powoduje radiolizę wody zawartej w tkankach. Uwolnione w jej wyniku rodniki tlenowe i wodorotlenowe rozrywają wiązania wodorowe pomiędzy parami zasad purynowych i pirymidynowych w łańcuchach kwasów nukleinowych (DNA i RNA), powodując uszkodzenie cząsteczki. Skutkiem tego oddziaływania są mutacje genetyczne lub martwica komórek. Szczególnie wrażliwe na oddziaływanie rodników tlenowych i wodorotlenowych są te odcinki DNA które ulegają procesowi replikacji, a zatem w komórkach które ulegają podziałowi mitotycznemu (znajdują się w fazie S, G2 lub M cyklu komórkowego). Fakt ten tłumaczy słuszność prawa Bergonie i Tribondeau, określającego wrażliwość tkanek na promieniowanie. Prawo to głosi, że promieniowrażliwość tkanek jest wprost proporcjonalna do aktywności proliferacyjnej danej tkanki i odwrotnie proporcjonalna do stopnia jej zróżnicowania (dojrzałości).
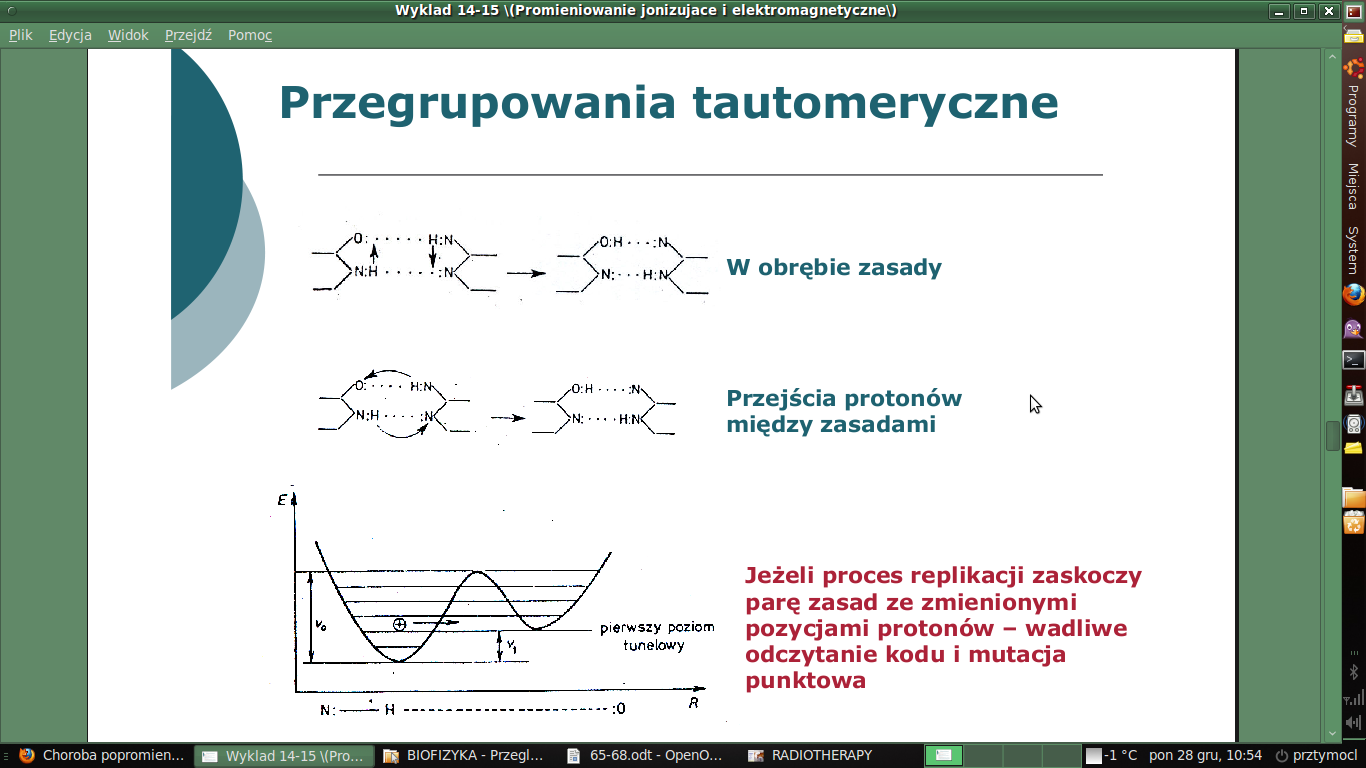
Aberracje chromosomów po napromienieniu (biodozymetr)
Mutacje chromosomowe powstałe w ściśle określonym punkcie chromosomów na skutek bezpośredniego lub pośredniego działania promieniowania jonizującego. Zwykle są to zmiany nieodwracalne. Promieniowanie powoduje poprzeczne pęknięcie całego chromosomu. Oderwane fragmenty łączą się ponownie (często odwrotnymi końcami), albo łączą się z fragmentami innych chromosomów. Mogą też nie połączyć się wcale. Struktura takich chromosomów zostaje zmieniona, następuje mutacja. Aberracje popromienne chromosomów mogą być potęgowane przez czynniki zewnętrzne, np. temperaturę.
Typy aberracji w których tracone są różne odcinki chromosomów

Przyczyny dużej wrażliwości komórek na napromienienie
Duża ilość dostarczonej energii do komórki w postaci promieniowania powoduje powstanie wolnych rodników które niszczą ultrastrukturę komórki, w tym enzymy, DNA, białka strukturalne, bez których komórki obumierają bądź przekształcają się w komórki nowotworowe.
Promieniowrażliwość komórki zależy od wielu czynników:
* właściwej promieniowrażliwości komórki,
* jednorodności populacji komórek,
* zdolności do naprawy uszkodzeń popromiennych,
* fazy w cyklu komórkowym (wieku komórki),
* stopnia jej utlenowania,
* stopnia jej uwodnienia.
- Wrażliwość komórek na działanie promieniowania jonizującego jest tym większa, im większa jest ich aktywność podziałowa i tym mniejsza, im większy stopień ich zróżnicowania (prawo Bergoniego i Tribondeau). Można przyjąć, że nowotwory o różnej histogenezie cechują się promieniowrażliwością zbliżoną do tkanek, z których się wywodzą.
- Jednostką dawki pochłoniętej promieniowania jest gray (Gy); 1 Gy odpowiada energii jednego dżula, pochłoniętej w masie 1 kilograma.
Podstawy radioochronny (chemiczne radioprotektory)
Radioprotektory są to substancje tiolowe, które podane przed napromienieniem obniżają efekty popromienne.
Radioochronę chemiczną dzielimy ze względu na efekty, którym przeciwdziała.
Przeciwdziałanie efektowi bezpośredniemu, polegające na donacji elektronów. Środkiem chemicznym w tym rodzaju radioochrony jest cysteamina.
Przeciwdziałanie efektowi pośredniemu może polegać na:
Przeciwdziałaniu potencjalizacji chemicznej poprzez:
Substytucję związków wysokoenergetycznych
Ochronę biostruktur
Inaktywację enzymów hydrolitycznych
Przeciwdziałanie potencjalizacji biochemicznej poprzez:
Hamowanie tworzenia się rodników na drodze: hipoksemii, ischemii i hipoksji
Inaktywację rodników na drodze: redukcji (środek chemiczny używany to cysteamina) i rozkładu (za pomocą substancji EDTA)
Scharakteryzuj funkcje błon biologicznych, podaj przykłady
Błony biologiczne biorą udział we wszystkich przejawach aktywności komórek (jest to pojęcie szersze niż błona komórkowa).
Do funkcji błon biologicznych należy:
tworzenie fizycznych granic, kontrola składu komórki, nadanie organellom komórkowym pewną suwerenność - błona biologiczna organellum wyodrębnia je z cytoplazmy,
selektywna przepuszczalność, transport ograniczonej liczby cząsteczek - białka integralne, kanały jonowe, pompy, receptory zanurzone w błonach biologicznych umożliwiają transport bierny bądź aktywny,
stanowienie granic faz, przekazywanie sygnałów chemicznych i energii z jednego przedziału do drugiego - wewnętrzna błona biologiczna mitochondrium stanowi granicę faz dla elektronów podczas fosforylacji oksydacyjnej, błony mogą prowadzić egzocytozę,
zapewnienie optymalnych warunków działania enzymów, pomp jonowych, receptorów - błony biologiczne stanowią rusztowanie, w którym umocowane są ww. struktury, oddzielają od siebie środowiska o różnym pH jak np. lizosom i cytoplazmę, dlatego zarówno w lizosomie jak i w cytoplazmie działają inne enzymy dostosowane do różnego pH.
Omów sposoby przetwarzania informacji przez sieci nerwowe
W skład sieci nerwowej wchodzą nerwy obwodowe oraz komórki ośrodkowego układu nerwowego.
Jej funkcje to: analiza odbieranych bodźców zewnętrznych, przechowywanie informacji w pamięci, kojarzenie i porównywanie informacji i sterowanie czynnościami organizmu.
Przetwarzanie informacji w sieci nerwowej odbywa się następującymi sposobami:
komórka B zostanie pobudzona wówczas, gdy została pobudzona komórka A
bezpośrednio z nią połączona - implikacja,
komórka C zostanie pobudzona, gdy zostanie pobudzona jedna z komórek A lub B bezpośrednio z nią połączonych - implikacja z alternatywą
komórka C zostanie pobudzona, gdy zostaną pobudzone obie komórki A i B bezpośrednio z nią połączone. Związane to jest z wysokim progiem pobudliwości neuronu C - implikacja z koniunkcją
komórka C zostanie pobudzona, gdy zostanie pobudzona komórka A bezpośrednio z nią połączona i nie zostanie pobudzona komórka B, połączona z komórką A za pomocą komórki inhibitorowej In - implikacja z koniunkcją i negacją.
Omów mechanizmy pamięci
Pamięć to odtwarzanie impulsów dochodzących do ośrodkowego układu nerwowego po pewnym, nieraz długim, czasie. Podłożem pamięci krótkotrwałej są impulsy elektryczne krążące po obwodach zamkniętych składających się z komórek nerwowych. Impuls dochodzący z zewnątrz do pętli składającej się z neuronów od czasu do czasu dochodzi do neuronu, który aktywowany jest dodatkowo zewnętrznym impulsem.
Pamięć krótkotrwała może być trwale usunięta poprzez: szok elektryczny, silny mechaniczny, raptowne impulsy elektryczne, przerwanie dopływu tlenu, oziębienie tkanki nerwowej. Informacje z pamięci krótkotrwałej są przekazywane do pamięci długotrwałej i mogą być tam przechowywane nawet dziesięciolecia. Podłoże pamięci nie jest znane. Jedna z hipotez zakłada utrwalenie się połączeń miedzy neuronami, szczególnie łatwo przewodzącymi impulsy. Inna hipoteza zakłada modyfikacje biosyntezy kwasów nukleinowych i białek przez impulsy elektryczne pamięci krótkotrwałej. Pamięć długotrwała nie zanika przy przejściowym zaburzeniu czynności elektrycznych mózgu.
Transmisja dźwięku z ucha środkowego do wewnętrznego
Strzemiączko pod wpływem uderzeń młoteczka wprawia w ruch błonę okienka owalnego. Wywołuje to ruch cieczy w ślimaku. Wtłaczanie błony okienka owalnego wywołuje ruch błony okienka okrągłego w kierunku ucha środkowego (OTOSKLEROZA).
Zmieniające się ciśnienie przenosi się na perylimfę schodów przedsionka, a stamtąd, poprzez błonę przedsionkowa do endolimfy oraz - przez szparkę osklepka do schodów bębenka.
Ruch cieczy wprawia z kolei w ruch błonę podstawna, na której znajduje się narząd spiralny.
W narządzie spiralnym odbywa się wstępna analiza dźwięku oraz zamiana sygnału mechanicznego (ruch błony podstawnej) na ciąg impulsów bioelektrycznych.
Omów na czym polega zmiana sygnału mechanicznego na elektryczny w komórkach zmysłowych
ZAMIANA SYGNAŁU MECHANICZNEGO NA ELEKTRYCZNY
Teoria rezonansowa Helmholtza (1863) - inaczej zwana teorią harfy, zakłada iż błona podstawna jest zbudowana z cienkich włókien (strun słuchowych), krótszych u podstawy ślimaka i reagujących na składowe wysokoczęstotliwościowe oraz dłuższych, na końcu ślimaka reagująca na składowe niskoczęstotliwościowe. Każde z włókien nastrojone jest na określoną częstotliwość rezonansową, dzięki czemu odpowiada tylko wybranej składowej zawartej w docierającym do słuchacza sygnale. Włókna po rozedrganiu pobudzają właściwe sobie komórki nerwowe ślimaka, dzięki czemu powstają impulsy elektryczne, które dalej przekazywane są do ośrodka słuchowego w mózgu. Teoria ta pomija udział wyższych pięter drogi słuchowej w procesie analizy dźwięku.
Drgania mechaniczne:
- swobodne - np. ruch wahadła matematycznego - „pozostawiony samemu sobie”
- wymuszone - pod wpływem zmieniającej się okresowo siły
Rezonans - gdy częstotliwość siły wymuszającej równa jest częstotliwości drgań swobodnych - amplituda drgań wymuszonych jest największa.
Helmholtz: ton - na zasadzie rezonansu - pobudza te „włókienka”, które mają częstotliwość drgań własnych równą częstotliwości transmitowanego do ślimaka dźwięku.
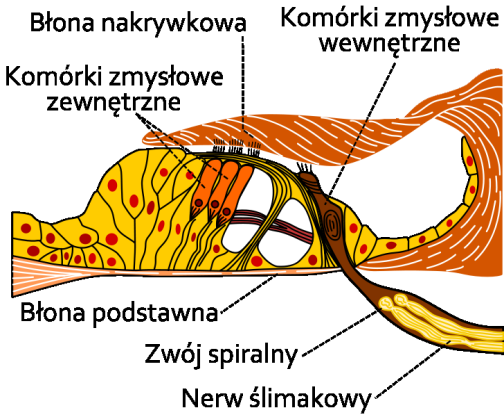
Budowa narządu spiralnego (Cortiego):
WKZ
_ Właściwe detektory ruchu
błony podstawnej
_ Ok.3 500 (w 1 rzedzie)
_ 20 włókien tworzy synapsy na 1 komórce (95%
unerwienia aferentnego (mielinowego, szybko transmitujacego dane)
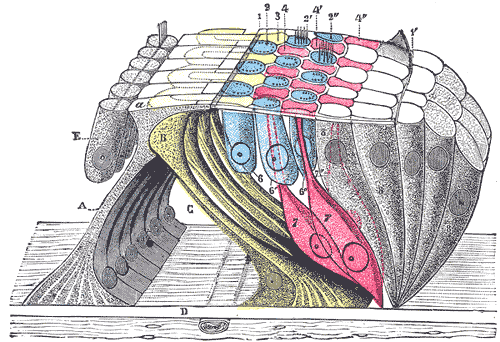
ZKZ
• Potrzebne do wzmocnienia
odpowiedzi WKZ
• Ok.20 000 (w 3 rzędach)
• 1 włókno zasila kilkanaście
komórek. Transmiterem
• eferentnym jest acetylocholina.
ich rzęski sa wtopione w błonę
nakrywkowa
Komórki zmysłowe narządu spiralnego - zamiana sygnału mechanicznego na elektryczny. Ruch błony podstawnej w górę i dół wywołuje cykliczne zbliżanie układu Cortiego i błony pokrywkowej - rzęski
przeginają się raz w jedna raz w drugą stronę. Przeginanie to połączone jest z otwieraniem i zamykaniem kanałów jonowych znajdujących się w rzęskach.
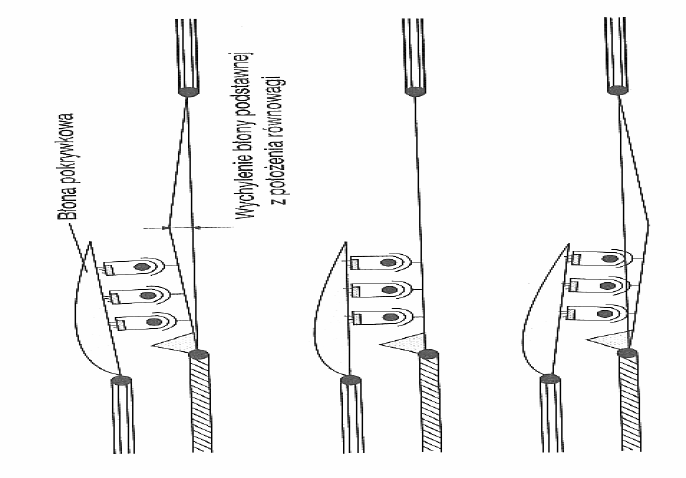
Zamiana sygnału mechanicznego na elektryczny - UDZIAŁ KATIONÓW
1. Przez otwarte kanały napływają K+, których stężenie w endolimfie jest bardzo wysokie.
2. Dochodzi do depolaryzacji błony rzęsek, co jest przyczyna aktywacji kanałów Ca2+.
3. Napływ Ca2+ indukuje fuzje pęcherzyków zawierających neurotransmiter (prawdopodobnie jest nim glutaminian), który wywołuje depolaryzacje błony postsynaptycznej.
Metody otrzymywania dwuwarstw lipidowych i pomiar ich grubości
Dwuwarstwa lipidowa - to spontanicznie powstająca w roztworach wodnych niektórych lipidów (głównie fosfolipidów) błona zbudowana z dwóch warstw cząsteczek tych związków chemicznych. Dwuwarstwa lipidowa stanowi podstawową część każdej błony biologicznej. Bez jej istnienia nie mogłyby funkcjonować żadne komórki żywe.
W pewnym zakresie stężeń, w roztworach wodnych niektóre rodzaje lipidów spontanicznie samoorganizują się w dwuwarstwę, która z formalnego punktu widzenia jest liotropową fazą ciekłokrystaliczną o budowie lamelarnej (Lα). Zjawisko to wynika z entropowo uzasadnionej dążności długich cząsteczek posiadających na jednym z końców grupę hydrofilową a na drugim hydrofobową do kierowania części hydrofilowej w stronę wody, a hydrofobowej w stronę innych cząsteczek lipidowych. W rezultacie dwuwarstwa lipidowa składa się z dwóch przeciwnie uporządkowanych warstw cząsteczek lipidu, z końcami węglowodorowymi (hydrofobowymi) zwróconymi do środka warstwy i polarnymi, hydrofilowymi grupami fosforylowymi na zewnątrz. Podobnie zachowują się również cząsteczki syntetycznych detergentów, np: mydła, które jednak zazwyczaj tworzą dużo mniej od lipidów stabilne dwuwarstwy.
SKŁAD LIPIDOWY BŁON - rozmieszczenie w dwuwarstwie błony
Zewnętrzna monowarstwa:
•Głównie lipidy cholinowe
Wewnętrzna monowarstwa:
•Ujemnie naładowana (grup NH2 - obecność PS - miejsce koncentracji PKC)
•Bardziej płynna - (kwasów nienasyconych)
WYZNACZANIE GRUBOSCI DWUWARSTWY LIPIDOWEJ (na podstawie pomiaru jej pojemności elektrycznej):
1. Błona stanowi nieprzepuszczalną dla jonów barierę (dielektryk o stałej dielektrycznej typowej dla wyższych węglowodorów nasyconych, e = 2,3 - 2,5), rozdzielająca 2 roztwory elektrolitów, stanowiące okładki kondensatora.
2. Grubość błony oblicza się przyjmując, że stanowi ona wraz z roztworami wodnymi kondensator płaski, którego pojemność wyliczana jest z szybkości zaniku impulsów w błonie.
3. Otrzymywana wartość to grubość jedynie hydrofobowego wnętrza błony.
36
Wyszukiwarka
Podobne podstrony:
~$RACOWANE ZAGADNIENIA NA EGZAMIN Z BIOFIZYKI 2009 2010 (dla tych doc
OPRACOWANE ZAGADNIENIA NA EGZAMIN Z BIOFIZYKI, BIOFIZYKA
Opracowanie Zagadnień na egzamin Mikroprocki
Opracowanie zagadnień na egzamin z MO
Przemiany geopolityczne (opracowane zagadnienia na egzamin)
Opracowane zagadnienia na egzamin
Andragogika opracowane zagadnienia na egzamin
opracowane zagadnienia na egzamin, ►► UMK TORUŃ - wydziały w Toruniu, ►► Socjologia, Praca socjalna,
Dydaktyka [opracowane zagadnienia na egzamin], Metodyka nauczania, język polski, teksty i notatki, e
Opracowanie zagadnień na egzamin z judaizmu, 2. GENEZA JUDAIZMU, Religia patriarchów
Konflikty opracowanie zagadnien na egzamin 2
opracowane zagadnienia na egzamin piachy
Opracowanie Zagadnień na egzamin Mikroprocki ściąga
Nauka?ministracji Opracowanie zagadnień na egzamin z NA
ZAGADNIENIA NA EGZAMIN Z MECHANIKI TECHNICZNEJ II DLA SEMESTRU III, sem III, +Mechanika Techniczna I
Zestaw 1, Opracowane zagadnienia na egzamin
Zestaw 15, Opracowane zagadnienia na egzamin
ściąga opracowane zagadnienia na egzamin piachy
Zestaw 22, Opracowane zagadnienia na egzamin
więcej podobnych podstron