MECHANIZMY DZIAŁANIA LEKÓW
Działanie leku nie jest wyłącznie funkcją jego właściwości fizykochemicznych lub jego budowy, warunkującej łączenie się cząsteczki substancji leczniczej z właściwym dla niej „miejscem” w organizmie, lecz uzależnione jest także od wielu zmiennych czynników, np.: masy ciała, wielu i płci chorego współistnienia różnych chorób i stanu ich zaawansowania.
Komórki organizmu przesyłają między sobą sygnały natury chemicznej.
Typy sygnalizacji komórkowej:
autokrynna: uwalniane przez komórkę substancje chemiczne działają na receptory umieszczone na własnej błonie komórkowej (autoreceptory)
parakrynna: komunikacja między komórkami oraz *
endokrynna: na większe odległości, sygnał chemiczny (hormon) uwalniany jest z grupy wyspecjalizowanych komórek (gruczoł) do krwi i z krwią substancje te docierają do komórek zawierających właściwe receptory
*neuronalna: neurony za pomocą wypustek przekazują potencjał czynnościowy, a sygnał elektryczny zamieniany jest na chemiczny w połączeniach między neuronami - synapsach (w synapsie sygnał elektryczny uwalnia neuroprzekaźnik, który działając na receptory w błonie postsynaptycznej generuje potencjał postsynaptyczny
Mechanizmy działania leków:
mechanizm fizykochemiczny działania leków- leki nieswoiste, nie łączą się ze swoistymi receptorami określonych tkanek, ale są obdarzone określonymi właściwościami fizykochemicznymi, np.;
wziewne środki znieczulenia ogólnego (eter dietylowy, halotan)
środki zmniejszające napięcie powierzchniowe (detergenty)
solne leki przeczyszczające (siarczan magnezu)
osmotyczne leki moczopędne (chlorek sodu)
mechanizm chemiczny działania leku- swoisty wpływ na określone struktury komórkowe
„teoria receptorowa”
leki wpływające na kanały jonowe
receptory metabotropowe ( w błonach komórkowych)- receptory sprzężone z białkiem regulatorowym G (receptor serotoninowy, dopaminowy, adrenalinowy)
receptory jonowe ( w których integralną częścią są kanały jonowe) (receptor GABA-ergiczny A)
receptory z udziałem kinazy tyrozynowej
receptory steroidowe
cyklaza adenylowa,
fosfolipaza C,
fosfolipaza A2
cAMP
IP3 (trifosfoinozytol)
DAG (diacyloglicerol)
Ca2+
b. leki wpływające na aktywność enzymów
c. leki hamujące metabolizm komórkowy (antymetabolity)
Mechanizm chemiczny działania leków - leki o tym samym mechanizmie działania są związkami bardzo aktywnymi chemicznie, o swoistym działaniu uzależnionym od budowy chemicznej; działanie to jest wynikiem chemicznego łączenia cząsteczek leku z receptorami lub też wpływem na aktywność enzymów, która najczęściej pod wpływem leku ulega zahamowaniu. Końcowego wyzwolenie efektu farmakologicznego jest wynikiem szeregu procesów biochemicznych.
AD. 2.A. „TEORIA RECEPTOROWA”
Działanie zewnętrznych bodźców, jak np. hormonów neurotransmiterów, czy czynników wzrostu na specyficzne białka receptorowe znajdujące się w błonie cytoplazmatycznej komórki powoduje serię zdarzeń prowadzących do powstania wtórnych przekaźników informacji, a w konsekwencji konkretnych efektów biologicznych.
Przekaźnictwo wewnątrzkomórkowe obejmuje: sygnał, receptor, transdukcja sygnału, efekt biologiczny.
Receptor - biopolimer o strukturze białkowej będący w stanie rozpoznawać określone ligandy (ligandy endogenne np. neuroprzekaźniki, ligandy egzogenne np. leki lub inne substancje) Według tej teorii warunkiem wystąpienia działania farmakologicznego jest połączenie się cząsteczki określonego leku z właściwym dla niego miejscem na błonie komórkowej lub z odpowiednim receptorem. Receptor posiada zdolność tworzenia kompleksu aktywnego z ligandem. Powoduje to szereg zmian strukturalnych i elektronowych w receptorze, a także w ligandzie, co prowadzi do określonego efektu biologicznego. Receptory mają swoisty charakter, tzn., że ich rodzaj i budowa przestrzenna grup czynnych powoduje ścisłe oddziaływanie leku z receptorem.
Receptory zdolne są do rozpoznawania dochodzących do komórek sygnałów, to z kolei uruchamia szereg procesów.
Powinowactwo chemiczne - zdolność oddziaływania leku z receptorem i stopień z jakim lek dopasował się do receptora.
Aktywność wewnętrzna - zdolność leku do wywoływania określonego pobudzenia receptora i wyzwalania efektu farmakologicznego
Agonista wykazuje powinowactwie do receptora i aktywność wewnętrzną
Antagonista wykazuje powinowactwie do receptora ale nie posiada aktywności wewnętrznej (następuje zablokowanie receptora)
Podział receptorów:
A. receptory błonowe
B. receptory cytoplazmatyczne
Receptory metabotropowe
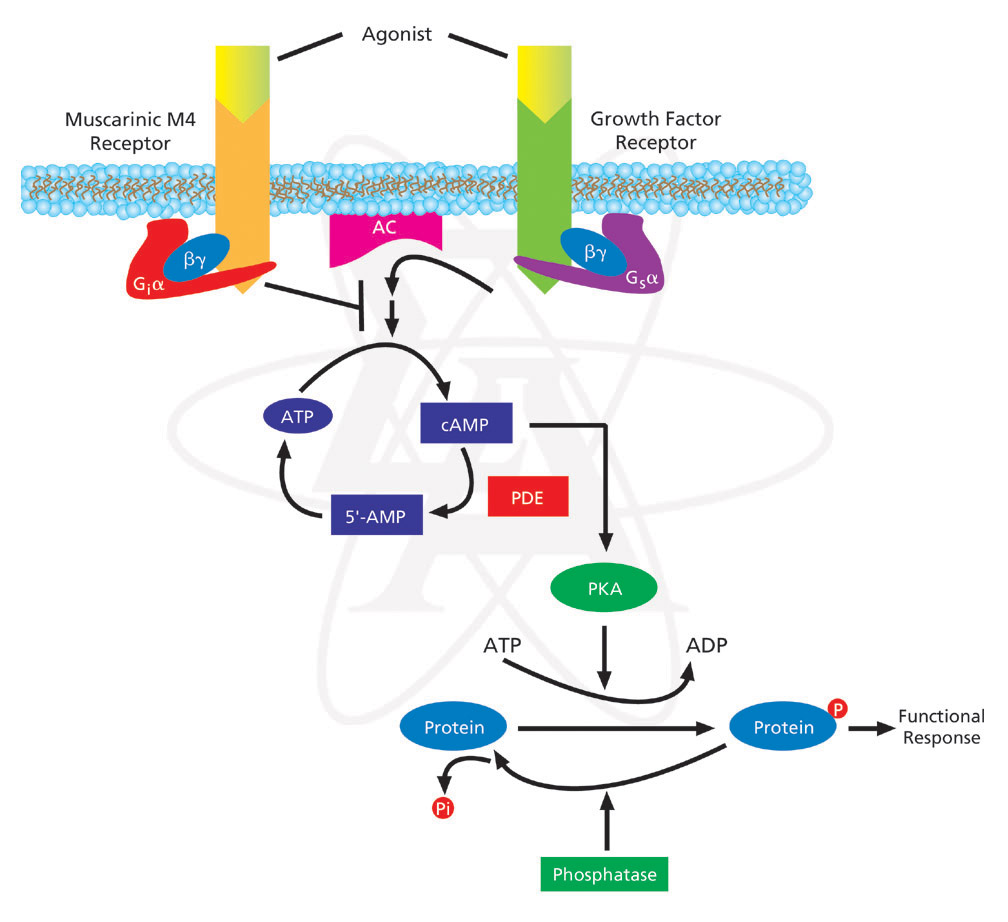
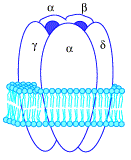
Tworzą największą rodzinę receptorów powierzchni komórki. Cechą wspólną receptorów związanych z białkiem G jest struktura I-rzędowa, która jest utworzona przeważnie z kilkuset jednostek aminokwasowych, tworząca 7 transbłonowych helis (przebijających 7-krotnie błonę komórkową). Białko G umieszczone jest po stronie cytozolowej błony komórkowej i Łożone jest z 3 podjednostek białkowych: α, β i γ. W stanie spoczynkowym wszystkie podjednostki tworzą razem kompleks, a dodatkowo podjednostka α jest związana z GDP (guanozynodifosforan). Agonistami działającymi za pośrednictwiem białka G są m.in.: hormony, prostaglandyny, neurotransmitery, cytokiny. Przyłączenie liganda do receptora powoduje związanie się receptora z białkiem G zastąpienie GDP przez GTP, co prowadzi do rozpadu białka G - odłączenia podjednostki α-GTP.
Podjednostka ta wiąże się z błonowymi białkami - enzymami t.j:
odpowiedzialnymi za syntezę przekaźników drugiego rzędu (wewnątrzkomórkowych sygnałów) takich jak:
GTP połączone z podjednostką α hydrolizuje do GDP i nieaktywna podjednostka łączy się z powrotem z kompleksem i sygnał zostaje wyłączony.
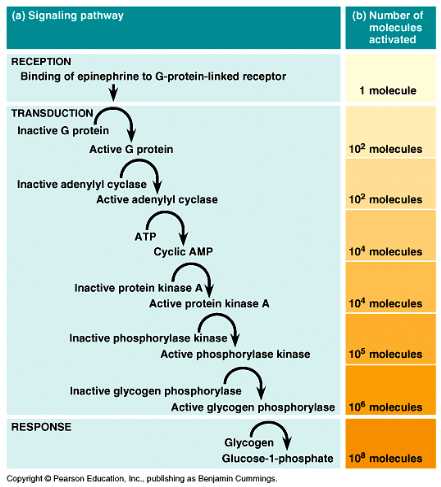
Cyklaza adenylanowa katalizuje syntezę cAMP z ATP. Ten wtórny przekaźnik aktywuje kinazy białkowe, co w efekcie prowadzi do uzyskania efektu metabolicznego.
Wg tego mechanizmu - ! na receptor β-adrenergiczny działa adrenalina (epinefryna) a efektem biologicznym jest rozpad glikogenu do glukozy
Cyclic Nucleotide Metabolism - cAMP
Ścieżka sygnalizacyjna dla adrenaliny (wpływ tego hormonu na enzymy wątrobowe)
Z kolei stymulacja fosfolipazy C uruchamia kaskadę związaną z uwalnianiem wtórnych przekaźników: IP3 i DAG Oba związki zwiększają stężenie Ca2+ poprzez uwalnianie go z wewnątrzkomórkowych magazynów i aktywują kinazę białkową C. Tak wywołują efekty wewnątrzkomórkowe pobudzone !!!receptory α- adrenergiczny czy muskarynowy
Receptoryzwiązane z kanałami jonowymi
Receptory te najczęściej zbudowane są z 5 podjednostek białkowych - przebijając błonę cytoplazmatyczną tworzą uporządkowaną strukturę wewnątrz której znajduje się kanał jonowy. Efektem pobudzenia tych receptorów jest zmiana przepuszczalności błony dla jonów (zmiana potencjału błonowego- depolaryzacja lub hiperpolaryzacja).
Przykłady: !!! receptor GABAergiczny A, receptor nikotynowy
Pentamerowa struktura receptora jonotropowego
Receptory wakazujące aktywność kinazy tyrozynowej lub kinazy serynowo-treoninowej
Związane są z przenoszeniem do komórki sygnału połączenia z receptorem takich agonistów jak np. insuliny, hormonu wzrostu, prolaktyny, cytokin. Receptor ten stanowi pojedynczy łańcuch polipeptydowy pojedynczo przebijający błonę komórkową. W tym przypadku receptor działa jak enzym i po związaniu liganda ulega dimeryzacji (choć cześć tych receptorów może już występować w postaci dimerów) oraz autofosforylacji zawartych w łańcuchu receptora reszt tyrozynowych. Umożliwia to przyłączenie się białek adaptorowych i aktywację kaskady kinaz białkowych.
mechanizm działania na stronie http://faculty.plattsburgh.edu/donald.slish/tyrosinekinase/TK1.html
Receptory steroidowe
!!!To receptory wewnątrzkomórkowe -cytoplazmatyczne lub umieszczone w jądrze komórkowym wpływające na procesy transkrypcji genowej. Powstający kompleks agonista-receptor wiąże się ze specjalnymi regionami DNA i wpływa na transkrypcję pewnych genów Agonistami są związki o charakterze lipofilowym: hormony steroidowe: hormony płciowe, hormony nadnerczy, hormony tarczycy oraz kwas retinowy (aktywna postać witaminy A), witamina D.
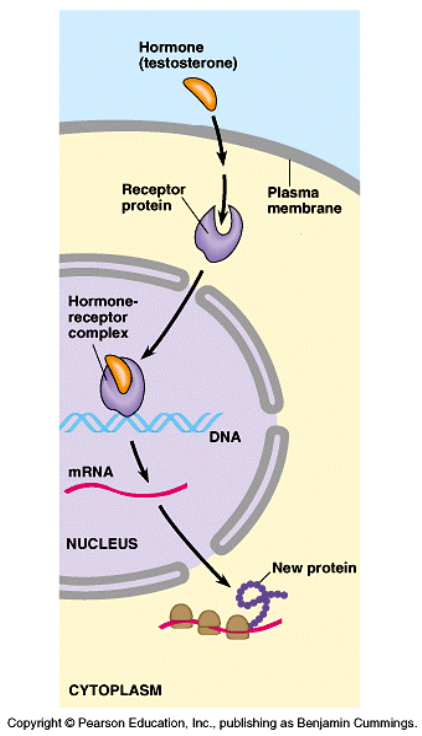
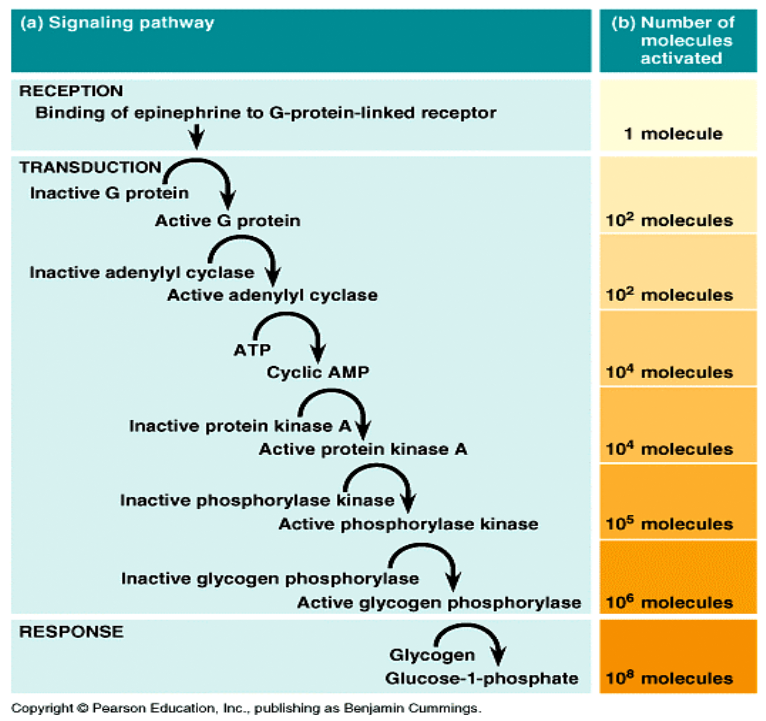
Połączenie cząsteczki agonisty z miejscem wiążącym receptora powoduje odłączenie podjednostki α (G), która aktywuje cyklazę adenylową (enzym zlokalizowany na wewnętrznej powierzchni błony komórkowej). Aktywacja cyklazy adenylowej prowadzi do konwersji adenozynotrójfosforanu (ATP) i powstania cyklicznego 3',5'-adenozynomonofosforanu (c-AMP).
c-AMP aktywuje zależne od c-AMP kinazy białkowe PKA (protein kinase A) w komórkach docelowych. W wyniku tej kaskadowej reakcji dochodzi do zmian fosforylacji białek enzymów, aktywacji różnych kanałów jonowych oraz zmian transkrypcji poszczególnych genów w komórkach.
Budowa receptora metabotropowego
Wyszukiwarka
Podobne podstrony:
Farmakologia ogólna Molekularne mechanizmy działania leków
FARMAKOLOGIA, 00 Mechanizm działania leków
Mechanizm działania leków psychotropowych
Czy nowsze badania nad patomechanizmem stresu i depresji zmienią poglądy na mechanizm działania lekó
Uwagi na temat mechanizmów działania leków przeciwdepresyjnych porównanie leków klasycznych z lekami
Polimorfizm enzymów, receptorów i innych białek biorących udział w mechanizmie działania leków
Mechanizm działania i farmakologia nowych leków z grupy inhibitorów monoaminooksydazy
Mechanizmy działania antybiotyków, materiały farmacja, Materiały 4 rok, farmacja 4 rok part 2, farma
Działanie leków na poszczególne neuroprzekaźniki, Technik farmaceutyczny, Farmakologia
3 TOKSYCZNOŚĆ, MECHANIZMY DZIAŁANIA I?WKOWANIE LEKÓW
Działanie leków na poszczególne neuroprzekaźniki i receptory, Farmacja, farmakologia, farmakologia n
Psychotropowe działanie leków przeciwpadaczkowych, Anastezjologia, Farmakologia
mechanizmy działania leku II, Farmakologia
5 MECHANIZM CHEMICZNY DZIAŁANIA LEKÓW
Mechanizmy działania antybiotyków, materiały farmacja, Materiały 4 rok, farmacja 4 rok part 2, farma
więcej podobnych podstron