DSCN2110 (2)

lYii włączonych ccntromerem. Rozchodzą sic w ten sposób ęhromosmi# u

W większości przypadków pierwszy podział meiotvcznv kończy sit* t«.Lr.

morki, tworzące izw. d i a d ę. Po krótkiej interfazie, ale bez replikacji DNA, 0l jądra diady zaczynają drugi podział mejotyczny (II), który nic różni się w istocie od podziału mitotycznego. Na zakończenie metafazy II następuje podział cenlromeru łączącego dotąd obie chromatydy w każdym chromosomie i chromatydy te — obec-nie już jako chromosomy siostrzane rozchodzą się w anafazie 11 do dwóch biegu, nów wrzeciona. Po telofazie II powstają 4 haploidalne jądra, a po cytokinezie || czter\ haploidalne komórki tworzące tetradę.
Pomiędzy I a II podziałem mejotycznym może brakować niektórych stadiów, np. cytokinezy I (w mejozie poprzedzającej rozwój pyłku u większości dwuliściennych). interfazy, a nawet telofazy 1 i profazy II. tak że w niektórych przypadkach (np. u Trillium) chromosomy po anafazie I przechodzą bezpośrednio do metafazy II.
Przebieg mejozy przedstawiony jest schematycznie na rysunku 5.12.
Jak wynika / przedstawionego wyżej przebiegu mejozy. rozmaitość kombinacji genetycznych w komórkach haploidalnych powstałych w rezultacie mejozy jest spowodowana (I) wymianą odcinków chro-matyd miedzy chromosomami homologicznymi w pachytenie, (2) przypadkowym przekazaniem chromosomów homologicznych pochodzących od jednego lub drugiego rodzica do jąder potomnych w ana-tazie I W komórkach diploidalnych pochodzących od zygoty liczba kombinacji zwiększa się dalej w wyniku przypadkowego łączenia się gamet w procesie syngamii.
5.3. ROZMNAŻANIE PŁCIOWE GLONÓW I GRZYBÓW
Przemiana faz jądrowych
U wszystkich rozmnażających się płciowo organizmów występuje w cyklu życiowym przemiana faz jądrowych, haploidalnej i diploidalnej. Faza haploidalna trwa od mejozy do syngamii, diploidalna od syngamii do mejozy:
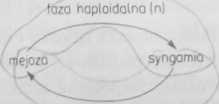
fazo diploidalna (2n)
Najprościej przedstawia się ta sprawa u roślin jednokomórkowych. Już u tych organizmów spotykamy różne rodzaje syngamii, od izogamii do oogamii. Omówimy je na przykładzie zielenie jednokomórkowych z rodziny Chlamydnmonadaceae. Si, lo formy wjęiowcowe swobodnie pływające w wodzie i rozmnażające się bez-płciowa przez, pojedyncze lub wielokrotne podziały mitotyczne_Sa one haplontami. tj. liczba chromosomów w jądrze komórek wegetatywnych jest haploidalna. W pewnych warunkach, zwłaszcza zmieniających się w kierunku niekorzystnym dla życia glonu, te jednokomórkowe organizmy rozmnażają się płciowo. U gatunków izo-gamicznych w większości przypadków gamety nie różnią się kształtem i budową od zwykłych komórek wegetatywnych. U Chlamydomonas euga/neios wygląda to tak. że wegetatywne dotąd komórki tworzą gęste skupienia, nabierają właściwości gamet i kopulują parami ze sobą (rys. 5.14 A—C). W innych przypadkach, np. u Chlamy-domonas sleinii, przez wielokrotny podział protoplastu powstają 32 lub 64 dwu-wiciowe izogamety (rys. 5.15 A—-D). Po uwolnieniu się z rodzicielskiej ściany komórkowej gamety te gotowe są do syngamii, przy czym jednak zarówno tu. jak i u Chla-mydomonas eugametos, występuje zróżnicowanie izogamet na formy 4- i — sprawiające, że kopulacja nie może nastąpić między jakimikolwiek dwiema gametami (np. pochodzącymi od tego samego osobnika), a tylko między gametami różnego „znaku”.
Brak zróżnicowania gamet jest tu więc tylko pozorny, dotyczy jedynie cech budowy, tzogamety występują Taktycznie w dwóch odmianach różniących się pewnymi cechami biochemicznymi. Istnieją dowody, że wzajemne przyciąganie się gamet i możliwość kopulacji polega tu na komplementarnych różnicach dotyczących przynajmniej częściowo biochemicznych cech wici.
U heterogamicznego Chlamydomonas braunii w jednych komórkach powstaje przez wielokrotny podział 8 lub 16 mikrogamet, a w innych 2 lub 4 makrogamety. Oogamia występuje u Chlorogonium oogamum, gatunku blisko spokrewnionego z rodzajem Chlamydomonas (rys. 5.16). Protoplast pewnych osobników zaokrągla się i po opuszczeniu ściany komórkowej staje się nieruchomą komórką jajową, w innych zaś dzieli się kilkakrotnie na 8, 16 lub 32 małe dwuwiciowe plemniki, które również wydostają się z zamykającej je ściany komórkowej. W wodzie plemniki otaczają komórkę jajową, a jeden wnika do środka i dokonuje zapłodnienia.
W przypadku izogamii lub heterogamii, jeżeli obie gamety miały po 2 wici, to powstała zygota przez pewien czas pływa jako komórka 4-wiciowa, a następnie traci wici i opada na dno zbiornika wodnego (rys. 5.15 E.F). Gdy występuje oogamia. zygota od początku nie ma wici. Zygota otacza się następnie grubą, odporną ścianą i staje się komórką przetrwalnikową (zygosporą), która przechodzi w stan spoczynku (rys. 5.14 D, 5.15 F). W tym stanie może przetrwać okres suszy, niskiej temperatury itp. Z nastaniem odpowiednich warunków zygosporą zaczyna się rozwijać, przy czym pierwszy jej podział jest podziałem mejotycznym, w wyniku którego powstają 4 haploidalne dwuwiciowe komórki (rys. 5.14 E, 5.15 G), które następnie mogą rozmnażać się bezpłciowo na drodze podziałów mitotycznych.
Inne glony jednokomórkowe, okrzemki, są diplontami i w tej fazie rozmnażają się bezpłciowo przez podział. Jak pamiętamy (por. str. 88). w związku
Wyszukiwarka
Podobne podstrony:
0000013 (9) W ciągu 16 lat leczono w ten sposób 320 z 336 przypadków świeżych złamań kości udowej, u
M a t e m a t k a k r o k p o k r o k u I Page 01 (38) Stosując skalę ciągłą, nasze spostrz
W ten sposób, w ocenie większości obserwatorów i opiniotwórczych instytucji zakończyła się faza
W ten sposób, w ocenie większości obserwatorów i opiniotwórczych instytucji zakończyła się faza
374 (14) Najczęściej stosuje się sieci /budowane z 3 warstw neuronów w ten sposób, że wejście dla pi
CIMG2768 nie podaje się im wody, zmuszając w ten sposóbl spożywania większej ilości pokarmu (pozosta
Warto zauważyć, że obliczana w ten sposób wartość c jest dodatnia. Interpretujemy to jako falę rozch
skanowanie0015 ÓD rENOMEŃOira^pÓ^®b^SraŃexAil5ZS«D W .ten. sposób istota pqe^i-
Nr 9PRZEGLĄD ELEKTROTECHNICZNY277 torka, którego obwód pierwotny włączony jest w ten sposób, że
85661 IMGy69 1 przedsionkowy. W ten sposób, podczas tego prostego zadania, w proces uczenia się włąc
więcej podobnych podstron