DSCN6274 (Kopiowanie)

256 fitti/nyw . rrprły/tmurn t!łu kaiulyiluU'n* nu ak/idrmlr mr<ly< /,

P
i
© garno*
J_
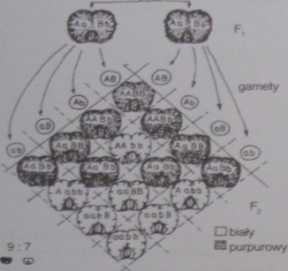
genów dominujących A i H. Każdemu / łych genóy, odpowiada allcl rcccsywny (a tub b). Osobnik będący homozygotą rcccsywną pod względem któregokolwiek z łych genów nic wylwar/a barwnika (pokolenie V na ryc. 6-22). Spotkanie się obu genów dominujących umożliwia natomiast syntezę barwnika (P, i cześć potomstwa F na ryc. 6-22). Można sobie wyobrazić, że zależności pomiędzy genami A i H są takie, jak przedstawiono na ryc. 6-23. Geny takie nazywamy genami albo współdziałającymi.
albo współdziałającymi.
Choć współdziałają one w wytworzeniu jednej cechy, jednak dziedziczą się niezależnie. Rozkład fenotypowy 9:7 jest modyfikacją znanego nam już rozkładu 9:3:3; I (II prawo Mendla). A zatem geny te muszą leżeć na różnych chromosomach. Jeżeli chcemy otrzymać groszki utrzymujące cechę purpurowej barwy kwiatów, należy krzyżować osobniki o genotypie AABB.
Współcześnie przeprowadzone badania wykazały, że w wytworzeniu barwy kwiatów biorą udział nie dwie. lecz wiele par genów. Proces wytwarzania barwnika jest
Ryc. 6-22. Współdziałanie genów na przykładzie bowiem typowym ciągiem przemian metabolicznych,
dziedziczenia barwy płatków groszku a zatem barwa kwiatu jest wynikiem współdziałam pachnącego ILathyrus odaratus). Opis w tekście. licznych genów związanych z syntezą różnych związków
P - pokolenie rodzicielskie. F,. F, - kolejne i wpływających na ich rozmieszczenie w komórkach
pokolenia mieszańców (JD).
rośliny (por. też biosyntezę barwnika mclaniny z fenyloalaniny u człowieka - ryt. 6-41).
Podobnie rzecz przedstawia się u zwierząt. Na przykład barwa sierści myszy, królików, świnek morskich i innych gryzoni determinowana jest współdziałaniem licznych, nicailclicznych genów, które oddziałują nic tylko na syntezę barwnika (ryc. 6-41), lecz i na jego rozmieszczenie we włosach. Poznaliśmy już działanie jednej pary tych genów: dominującego B powodującego czarne zabarwienie sierści i recesywnego - b - barwy brunatnej (ryc. 6-16). Okazało się jednak, że do wytworzenia barwnika niezbędne są też inne geny. Przede wszystkim gen C, determinujący syntezę mclaniny w sierści. Gdy tego genu nic ma. synteza barwnika nic następuje: homozygoty cc są białymi albinosami niezależnie od zestawu pozostałych genów (ryc. 6-24). Jeżeli - jak to ilustruje ryc. 6-24 skrzyżujemy mysz-albinosa (homozygotę bbcc) z homozygotyczną czarną myszą (BBCC). to w F uzyskamy osobniki wyłącznie czarne (BbCc). Ponieważ jednak są to heterozygoty, wytwarzają czety rodzaje gamet: BC, Be. bC i be. Przez wzajemne ich łączenie w drugim pokoleniu uzyskujemy 9 mysz) fenotypowo czarnych. 3 fenotypowo brązowe i 4 fenotypów o białe.
Ale i jeszcze inne geny współdziałają w wytworzeniu barwy sierści gryzonia. Np. myszy płowe (o barwie -dzikiej") mają w komórkach geny B i C (a więc ich sierść winna być czarna), lecz oprócz ■ach posiadają inny gen A. Wywołuje on nierównomierne ubarwienie włosa: mc lam na gromada ■sę w jego szczytowej części oraz u podstawy, a pomiędzy tymi warstwami występuje pasek niemal be/tmrmmy. Efekt optyczny to właśnie płow a (..dzika") sierść. Osobniki nie posiadające genu A tocz jrga allci a sąjednolicie ubarwione (np. czarne).
Podobnie z ubarwieniem brunatnym osobników posiadających geny C i M>. W obecność gsms A ich włosy są nierównomiernie zabarwione (sierść „cynamonowa”), zaś w obecności wykąc mm d&tm równomiernie (sierść brązowa). Te zależności zestawiono w lab 6 III
Wyszukiwarka
Podobne podstrony:
DSCN6274 (Kopiowanie) 256 liitilnyju - rrprlyUiriuin iflti ktmdytiut/iw nu ubuJrm/U mfllyfjnr © 1© ]
DSCN6264 (Kopiowanie) 246 Biologia - rrprlylorium dla kandydatów no akademie mrdono b aMIK III HM m
DSCN6200 (Kopiowanie) 184 Biologia - rrpetylorium dla kandydatów na akademie medyczn* 184 Biologia -
DSCN6200 (Kopiowanie) 184 Biologia - repetytorium dla kandydatów na akademie medyczne oddając lam dw
DSCN6201 (Kopiowanie) B 1>CO,*2H,S (CH,0)*2S»H,0 2) 3CO,*2S*5
DSCN6201 (Kopiowanie) Światło chloro ADP* P, *-ATP ^chlorom* A cylochromy ATP ADP *P/ Ryc, 5 • 23.
DSCN6202 (Kopiowanie) 186 Biologia - repetytorium dla kandydatów na akademie mrdynj» 5.2.4. Fotofosf
DSCN6202 (Kopiowanie) 186 Biologia - repetytorium dla kandydatów na akademie medytan 5.2.4. Fotofosf
DSCN6203 (Kopiowanie) , wwłnfem 1*7 Na.SĄ + H.O + łOj -► NajSO^ + H;S04 + 883 kJ/mol. Natomiast prze
DSCN6203 (Kopiowanie) 187 (IMMCk Nr!^;Oj + HjO+20j -► Na2S04+HjSO, + 883 kJ/mol. Natomiast przedstaw
DSCN6205 (Kopiowanie) 188 Biologia - rr/fctylorium dla kandydatów na akademi0 mv,iv< 188 glikogon
DSCN6205 (Kopiowanie) MS Biologia - repetytoriom dla kandydatów na akademii- głikogen Ryc. 5 - 25. G
DSCN6206 (Kopiowanie)
DSCN6206 (Kopiowanie) Whtab<>li:"i .3.1. Przemiany pirogronianu I Pirogronian powstaje z
DSCN6207 (Kopiowanie) 190 Biologia - repetytorium dla kandydatów na akademie medyarm 190 Biologia -
DSCN6207 (Kopiowanie) 190 Biologia - repetytorium dla kandydatów na akademie medyezne 5.3.2. Cykl Kr
DSCN6208 (Kopiowanie) powstały związek - szczawiobursztynian - jest nietrwały i w obecności tej same
DSCN6208 (Kopiowanie) powstały /.wiązek szczawiobursztynian - jest nietrwały i w obecności tej samej
DS
więcej podobnych podstron