DSCN6274 (Kopiowanie)

256
liitilnyju - rrprlyUiriuin iflti ktmdytiut/iw nu ubuJrm/U mfllyfjnr
© 1© ]
© | © gamoty
garrwty
9:7 • 09
ca * B T
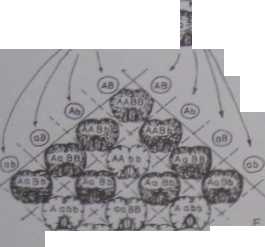
genów dominujących A i Ii. Każdemu /. łych genów odpowiada allcl rcccsywny (a lub h). Osobnik będący homozygolą reccsywną pod względem któregokolwiek z tych genów nic wytwarza barwnika (pokolenie l‘ na ryc. 6-22). Spotkanie się obu genów dominujących umożliwia natomiast syntezę barwnika (F, i cześć potomstwa l;. nu ryc. 6-22). Można sobie wyobrazić, że zależności pomiędzy genami A i B są takie, jak przedstawiono na ryc. 6-23. Geny takie nazywamy genami uzupełniającymi sic albo współdziałającymi Choć współdziałają one w wytworzeniu jednej cechy, jednak dziedziczą się niezależnie. Rozkład fenotypowy 9:7 jest modyfikacją znanego nam już rozkładu 9:3:3:1 (II prawo Mendla). A zatem geny te muszą leżeć na różnych chromosomach. Jeżeli chcemy otrzymać groszki utrzymujące cechę purpurowej barwy kwiatów, należy krzyżować osobniki o genotypie AABB
(wpww»jr
Ryc. 6-22. Wsjiółdziąlanie genów na pizykladzic dziedziczenia barwy płatków groszku pachnącego (Juuhyrus odoralu.il. Opis w tekście. P - pokolenie rodzicielskie. F(. F, - kolejne jmkolenia mieszańców (JD).
Współcześnie przeprowadzone badania wykazały, że w wytworzeniu barwy kwiatów biorą udział nic dwie. lecz wiele par genów. Proces wytwarzania barwnika jest bowiem tyjjowym ciągiem przemian metabolicznych, a zatem barwa kwiatu jest wynikiem wsjjółdziatama licznych genów związanych z syntezą różnych związków i wpływających na ich rozmieszczenie w komórkach rośliny (por. też biosyntezę barwnika mclaniny z fenyloalaniny u człowieka ryc. 6-41).
Podobnie rzecz przedstawia się u zwierząt Na przykład barwa sierści myszy, królików, świnek morskich i innych gryzoni determinowana jest współdziałaniem licznych, nicallclicznych genów, które oddziałują nie tylko na syntezę barwnika (ryc. 6-41), lecz i na jego rozmieszczenie we wlotach. Poznaliśmy już działanie jednej pary tych genów: dominującego B powodującego czarne zabarwienie sierści i rccesywnego - b - barwy brunatnej (ryc. 6-16). Okazało się jednak, że do wytworzenia barwnika niezbędne są też inne geny. Przede wszystkim gen C. determinujący syntezę mclaniny w sierści. Gdy tego genu nic ma. synteza barwnika nie następuje: homozygoty cc są białymi albinosami niezależnie od zestaw u pozostałych genów (ryc. 6-24). Jeżeli jak to ilustruje ryc. 6-24 skrzyżujemy mysz-albinosa(homozygotębbcc)z homozygotycznączarną myszą(BBCC). to* F uzyskamy osobniki wyłącznic czarne (BbCc). Ponieważ jednak są to heterozygoty, wytwarzają c/en rodzaje gamet: BC, Bc. bC i be. Przez wzajemne ich łączenie w drugim jtokolcniu uzyskujemy 9 my vy fenotypowo czarnych. 3 fenotypowo brązowe i 4 fenotypowo białe.
Ale i jeszcze inne geny współdziałają w wytworzeniu barwy sierści gryzonia. Np. myszy płowe (« barwie -dzduej") mają w komórkach geny B i C (a więc ich sierść winna być czarna), lecz oprócz ■tch posiadają inny gen - A. Wywołuje on nierównomierne ubarwienie włosa: mclanma gromadzi aię w jego szczytowej części oraz u podstawy, a pomiędzy tymi warstwami występuje pasek niemal bezbarwny. Efekt optyczny to właśnie płowa („dzika") sierść. Osobniki nie posiadające genu A lecz jego alleł a są jednolicie ubarwione (np. czarne).
Podobnie z ubarwieniem brunatnym osobników posiadających geny C i bb. W ubacansn —uAicbwfcisy tą nierównomiernie zabarwione (sierść .cynamonowa”), zaś w obecności wy łącffi* równonwernie (sierść brązowa). Te zależności zestawiono w lab, 6 III
Wyszukiwarka
Podobne podstrony:
DSCN6274 (Kopiowanie) 256 fitti/nyw . rrprły/tmurn t!łu kaiulyiluU n* nu ak/idrmlr mr<ly< /, P
DSCN6264 (Kopiowanie) 246 Biologia - rrprlylorium dla kandydatów no akademie mrdono b aMIK III HM m
DSCN6247 (Kopiowanie) 1 oiMi:w nyd „topl^
DSCN6259 (Kopiowanie) ’8o cel •lany a; ^PnyatW Praedkny46,^ •kacjęisWiyw ‘weposzca^ eden ze spowk,
DSCN6126 (Kopiowanie) j Struktura motekutama istot śyu/ysrh IW j Struktura motekutama istot śyu/ysrh
DSCN6200 (Kopiowanie) 184 Biologia - rrpetylorium dla kandydatów na akademie medyczn* 184 Biologia -
DSCN6200 (Kopiowanie) 184 Biologia - repetytorium dla kandydatów na akademie medyczne oddając lam dw
DSCN6201 (Kopiowanie) B 1>CO,*2H,S (CH,0)*2S»H,0 2) 3CO,*2S*5
DSCN6201 (Kopiowanie) Światło chloro ADP* P, *-ATP ^chlorom* A cylochromy ATP ADP *P/ Ryc, 5 • 23.
DSCN6202 (Kopiowanie) 186 Biologia - repetytorium dla kandydatów na akademie mrdynj» 5.2.4. Fotofosf
DSCN6202 (Kopiowanie) 186 Biologia - repetytorium dla kandydatów na akademie medytan 5.2.4. Fotofosf
DSCN6203 (Kopiowanie) , wwłnfem 1*7 Na.SĄ + H.O + łOj -► NajSO^ + H;S04 + 883 kJ/mol. Natomiast prze
DSCN6203 (Kopiowanie) 187 (IMMCk Nr!^;Oj + HjO+20j -► Na2S04+HjSO, + 883 kJ/mol. Natomiast przedstaw
DSCN6205 (Kopiowanie) 188 Biologia - rr/fctylorium dla kandydatów na akademi0 mv,iv< 188 glikogon
DSCN6205 (Kopiowanie) MS Biologia - repetytoriom dla kandydatów na akademii- głikogen Ryc. 5 - 25. G
DSCN6206 (Kopiowanie)
DSCN6206 (Kopiowanie) Whtab<>li:"i .3.1. Przemiany pirogronianu I Pirogronian powstaje z
DSCN6207 (Kopiowanie) 190 Biologia - repetytorium dla
DSCN6207 (Kopiowanie) 190 Biologia - repetytorium dla kandydatów na akademie medyezne 5.3.2. Cykl Kr
więcej podobnych podstron