
Ćwiczenie V. „Panta rhei” - zastosowanie równań
różniczkowych do modelowania procesów biologicznych
• Strona internetowa ćwiczeń:
http://www.home.umk.pl/~henroz/matm1112
Załóżmy, że mamy
taksonomiczną grupę zwierząt lub roślin
. Niech „
e
”,
będzie
prawdopodobieństwem, że wymrą
one w procesie ewolucji, a
„c” –
niech będzie prawdopodobieństwem specjacji tej grupy
w ewolucji.
Doświadczalnie, jest to do udowodnienia, że ogólne przys-tosowanie tej
grupy do życia w danych warunkach
(„fitness”) spada wraz z upływem
czasu
. Liczba zjawisk
specjacji, spada
w czasie, a liczba zjawisk
wymierania odpowiednio wzrasta
. Załóżmy, że
zależ-ność pomiędzy
obydwoma ww. zjawiskami jest liniowa
, tzn., szyb-kości specjacji i
wymierania są odpowiednio
odwrotnie i wprost proporcjonalne do czasu
.
Możemy dzięki temu założeniu opisać
zmianę liczby gatunków w czasie
.
Zmiana ta jest
różnicą pomiędzy ogółem zjawisk specjacji i wymierania
.
Proces taki może być opisany przez
równaniem różnicowe
, lub – jeżeli
weźmiemy pod uwagę b. małe odstępy czasu – przez tzw.
równanie
różniczkowe
.
Równanie różniczkowe wyznacza zależność pomiędzy nieznaną fun-kcją, a
jej pochodnymi
. Rozwiązanie równania różniczkowego polega

na znalezieniu funkcji f
, której
pochodne spełniają to równanie
. W praktyce
rozwiązanie równania różniczkowego
polega na sprowadze-niu go do
standardowej formy,
a następnie na scałkowaniu jednej (prawej) lub
obydwu stron równania
. W rozpatrywanym przykładzie zmiany liczby
gatunków w obrębie danego taksonu/grupy w czasie
(w wyniku procesów ewolucji), – oznacza zmianę, N – liczbę gatunków, t –
czas, e – prawdopodobieństwo wymierania, c – prawdopodobieństwo
specjacji:
N = N
t+1
– N
t
= (ctN
t
– etN
t
)t
; po zamianie równania
różnicowego na różniczkowe:
dN/dt = ctN – etN
. Czy możliwe jest obliczenie
liczby gatunków w dowolnym czasie? Tak, o ile rozwiążemy równanie
względem N, tzn. gdy rozwiążemy równanie różniczkowe. Dzięki prostemu
przekształceniu (dzielenie obu stron przez N i mnożenie przez dt)
uzyskujemy:
dN/N = (c – e)tdt
. Teraz całkujemy obydwie strony równania
(podobnie, jak na ćw. poprzednim) i uzyskujemy:
Jak w ćw.
poprzed-
nim,
N
0
u-
zyskano
ustawiając
t = 0
. Dwa przykłady wykresów takiej funkcji zostaną pokazane na
następnym przeźroczu. Widać tu, że nawet
mała różnica w szybkości
wymierania i specjacji
może doprowadzić
do znacznego wzrostu
liczby gatunków
w obrębie taksonu lub do jego
szybkiego wymarcia
.
2
2
2
2
2
2
1
2
0
ln( )
2
c e
c e
t
t
c e
N c
t
c
N Ce
N e
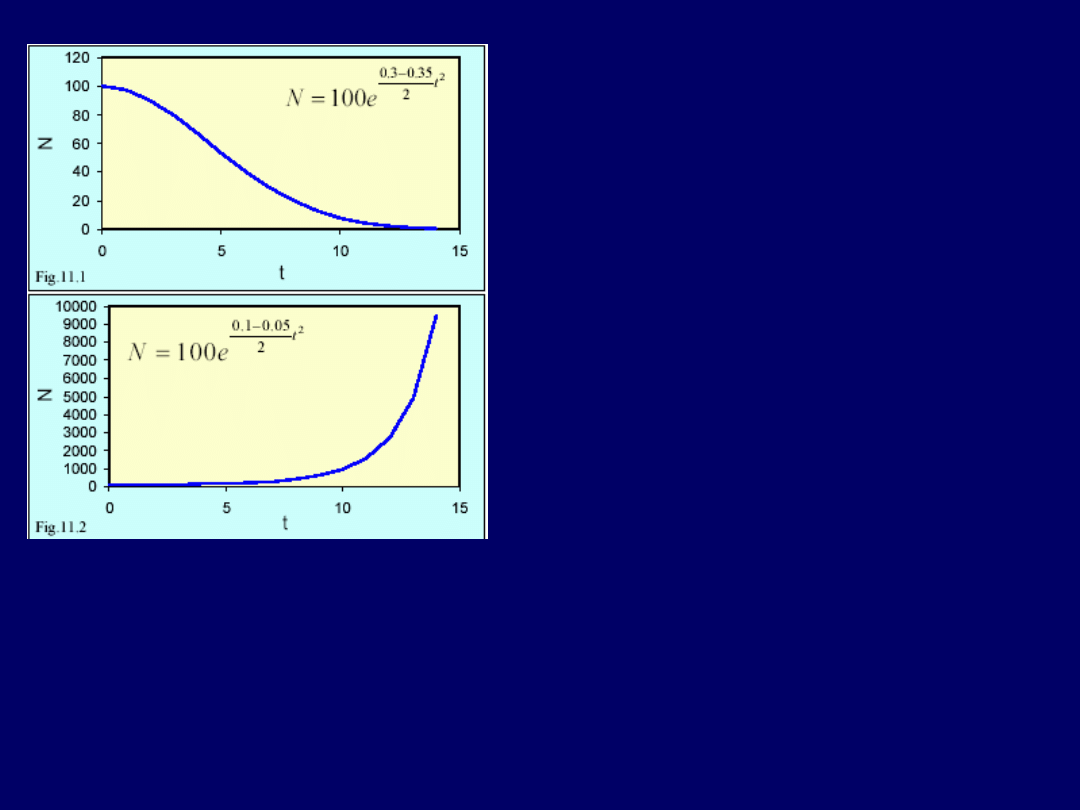
Dynamika wymierania i specjacji:
Wniosek: w przyrodzie
oby-
dwa te procesy są modyfikowane
przez dodatkowe mechanizmy
,
które nie zostały uwzględnione w
ww. prostym modelu. Rozwiązane
powyżej równanie, jest
liniowym
równaniem różniczkowym pierw-
szego rzędu
. I-go rzędu, ponieważ
pochodna była I-go rzędu (I-sza) i
liniowe, ponieważ nie wystąpiły
wykładniki wyższe niż 1 przy N.
Równania różniczkowe I rzędu
są
bardzo ważne w biologii
i moż-
na je zapisać w formie ogólnej:
dy/dx + f(x)y = g(x)
, gdzie f i g są
funkcjami zależnymi od x. Równa-
nie jest zwane
homogenicznym
(jednorodnym), jeżeli
g(x) = 0
dla wszystkich wartości x. Jeżeli roz-
patrzymy tylko to jednorodne równanie, to po przekształceniu uzys-
kujemy:
dy/y = –f(x)dx
. Po scałkowaniu obu stron i ew. dodatkowych
przekształceniach uzyskujemy:
y = Ce
–f(x)dx
. Stała C jest często ozna-czana
przez doprowadzenie x do odpowiedniej wartości (najczęściej 0) i
rozwiązanie względem C.
Jeżeli równanie jest niejednorodne –
sytuacja skomplikowana
.
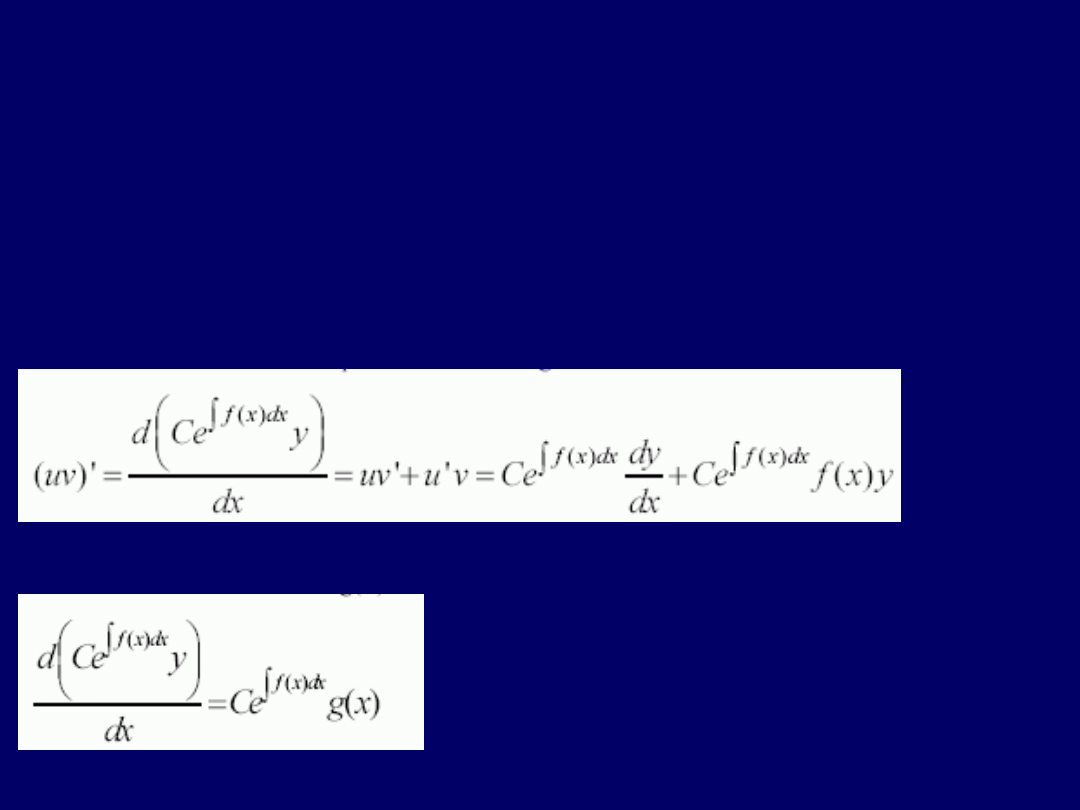
Stosujemy wtedy wzór na
pochodną iloczynu: (uv)’ = u’v + uv’
. Musimy
znaleźć taką
funkcję a(x)
, przez którą moglibyśmy pomnożyć nasze liniowe
równanie różniczkowe I-go rzędu i w odniesieniu do czego możnaby później
zastosować wzór na pochodną iloczynu. Dlatego
funk-cja a(x), musi spełniać
następujący warunek: a(x)dy/dx = a(x)f(x)y + d(a(x)y)/dx
. Równanie to ma
taką samą strukturę, jak wzór na pochod-ną iloczynu, a ponadto:
da/dx =
a(x)f(x)
. Jest to jednorodne równanie różniczkowe I-go rzędu, którego
rozwiązanie podano powyżej. Ustala-my:
a(x) = Ce
f(x)dx
, a następnie
podstawiamy to do naszego wzoru na pochodną iloczynu i uzyskujemy:
Pamiętając, że:
dy/dx + f(x)y = g(x),
wnosimy, że
g(x) powinna spełniać
następujący warunek
: Teraz możemy
scałkować obie strony
tego równania
, uzyskując:
Ce
f(x)dx
y =
Ce
f(x)dx
g(x)dx + C
1
Ostatecznie po przekształceniu tego rów-
nania przez podzielenie przez I-szą całkę:
y = e
–f(x)dx
e
f(x)dx
g(x)dx + ce
–f(x)dx
. Jest to

najbardziej
ogólne rozwiązanie liniowego równania różniczkowego I-go rzędu
.
Wygląda ono skomplikowanie, ale w większości przypadków jest
łatwe do
wykorzystania
, o ile funkcje f(x) i g(x) nie są zbyt skompliko-wane.
Przykład praktyczny:
stężenia niektórych hormonów i wielu leków w
krwioobiegu
mogą być modelowane jako
suma szybkości produkcji lub
indukcji i szybkości rozkładu
. Załóżmy, że lek podawany jest z mniej lub
bardziej stałą szybkością „g”. Równocześnie jest on metabolizowa-ny –
proporcjonalnie do jego stężenia. Proces ten można opisać nastę-pującym
równaniem:
dF/dt = g – fF
. Zmiana stężenia leku F w czasie t jest zwykłą
różnicą pomiędzy szybkością podawania (g) a szybkością
wykorzystywania/zużywania fF. Jest to
niejednorodne (inhomogenous)
równanie różniczkowe I-go rzędu z g = f(t) = const. i c = g(t) = const
. Aby
wyliczyć stężenie leku w dowolnym czasie t, wykorzystujemy roz-wiązanie
ogólne. Potrzebujemy: –
f(t)dt = –
fdt = – (ft + C
1
); po podsta-wieniu:
e
f(t)dt
g(t)dt =
(e
ft+C1
)gdt = g(1/f)e
ft+C1
+ C
2
. Podstawiamy te wy-niki do
rozwiązania ogólnego i uzyskujemy:
Stałe C, to wszyst-
ko stałe całkowania.
Pozostałą stałą K można
oznaczyć przyjmując t=0.
Dlatego
K = F(0)–c/g
.
Stąd ostateczny wynik może-
my zapisać następująco:
1
1
1
(
)
ft C
ft C
ft C
ft
g
g
F Ce
e
e
Ke
f
( )
(0)
ft
g
g
F t
F
e
f
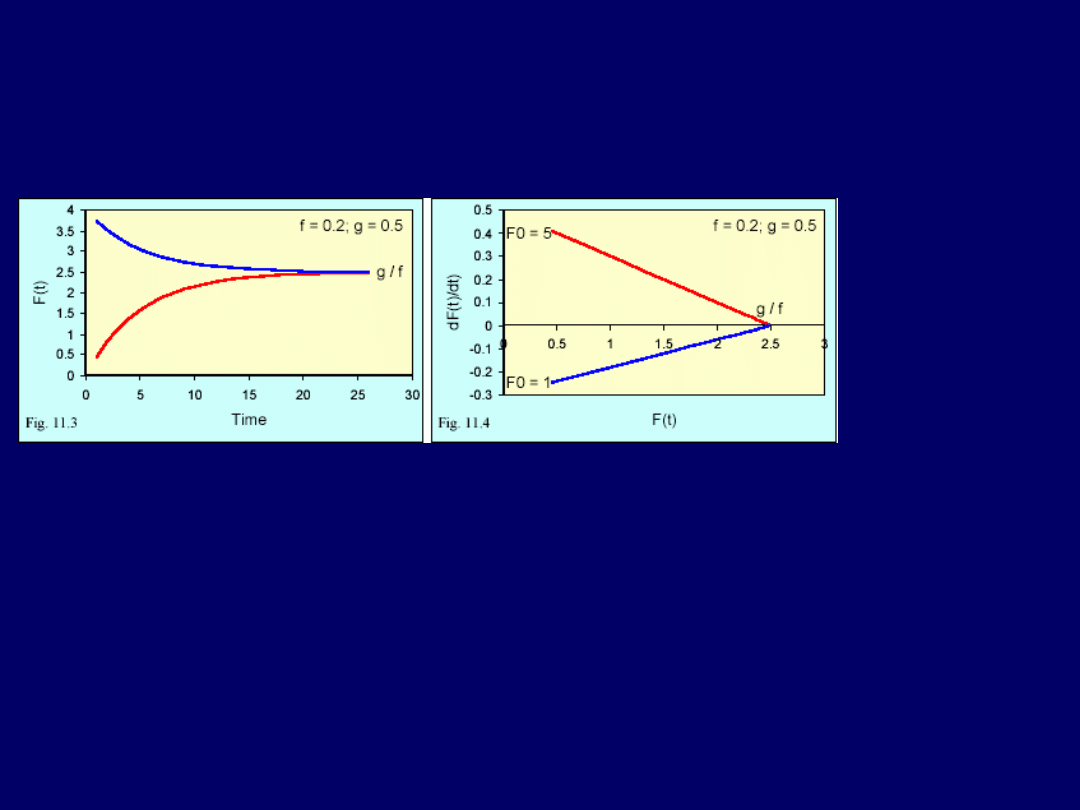
Wynik ten jest ważny, ponieważ jest to
ogólne rozwiązanie liniowego
równania różniczkowego I-rzędu, gdzie obie funkcje: f(t) i g(t) są stałymi
.
Można również zastosować
program matematyczny do rozwiązania na-
szego problemu
(Mathematica, wxMaxima – cz. praktyczna). Lewa cz.
rysunku obok przedstawia
zależność F(t) od czasu
. Bez względu na stę-
żenie począt-
kowe, proces
zmierza do
stężenia sta-
bilnego.
Stę-
żenie to jest
zwane pun-
ktem równo-
wagi
. Punkt ten wyliczamy, ustawiając
dF/dt = 0
. To daje nam bezpoś-
rednio:
F
równowagi
= g/f
, co jest widoczne na rys. Prawa cz. rys. przedsta-
wia tzw.
diagram fazowy procesu
.
dF/dt jest funkcją liniową
i bez wzglę-
du na stężenie początkowe, pierwiastkiem tej funkcji jest
F(t) = g/f
.
Inny przykład praktyczny:
wykładnicze równanie wzrostu
. Równanie to
jest
realistyczne tylko dla b. niskich wielkości populacji
. W rzeczywis-
tości są pewne
górne granice, zwane „zdolnościami przepustowymi”
(„carrying capacities”)
i sprawiają one, że powyżej ich,
rzeczywisty
wzrost populacji staje się wolniejszy dla wyższych wielkości populacji
.
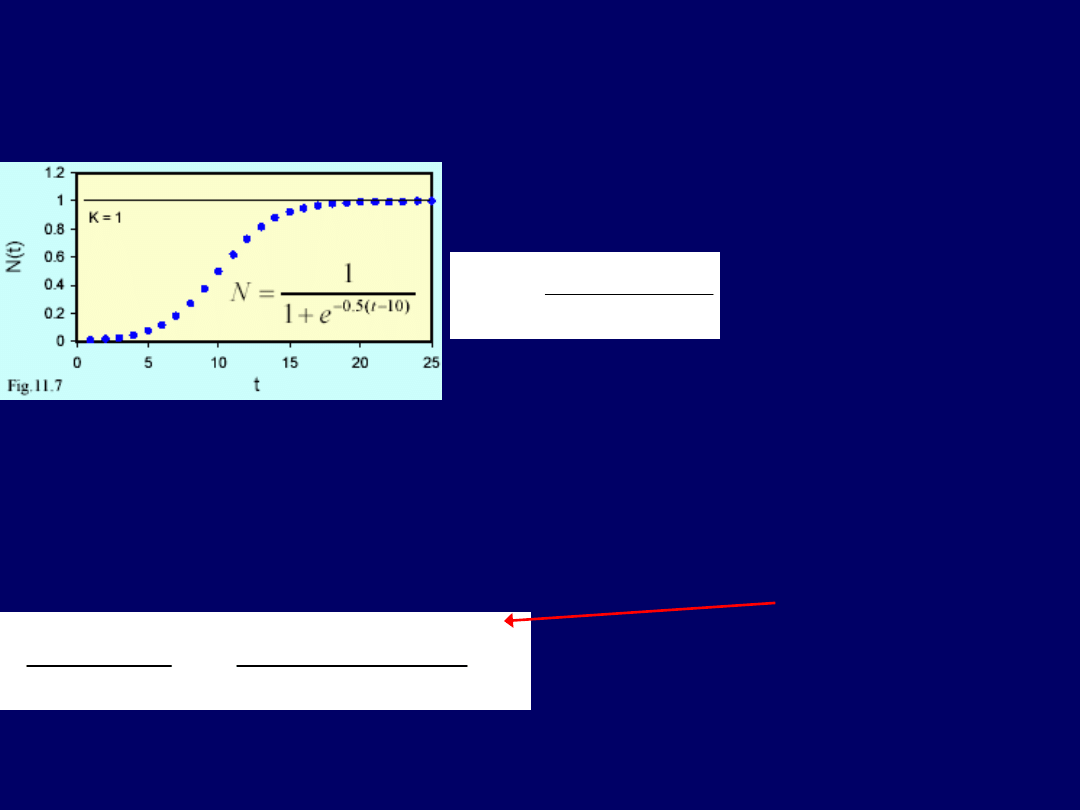
Rys poniżej przedstawia populację drożdży Saccharomyces cerevisiae,
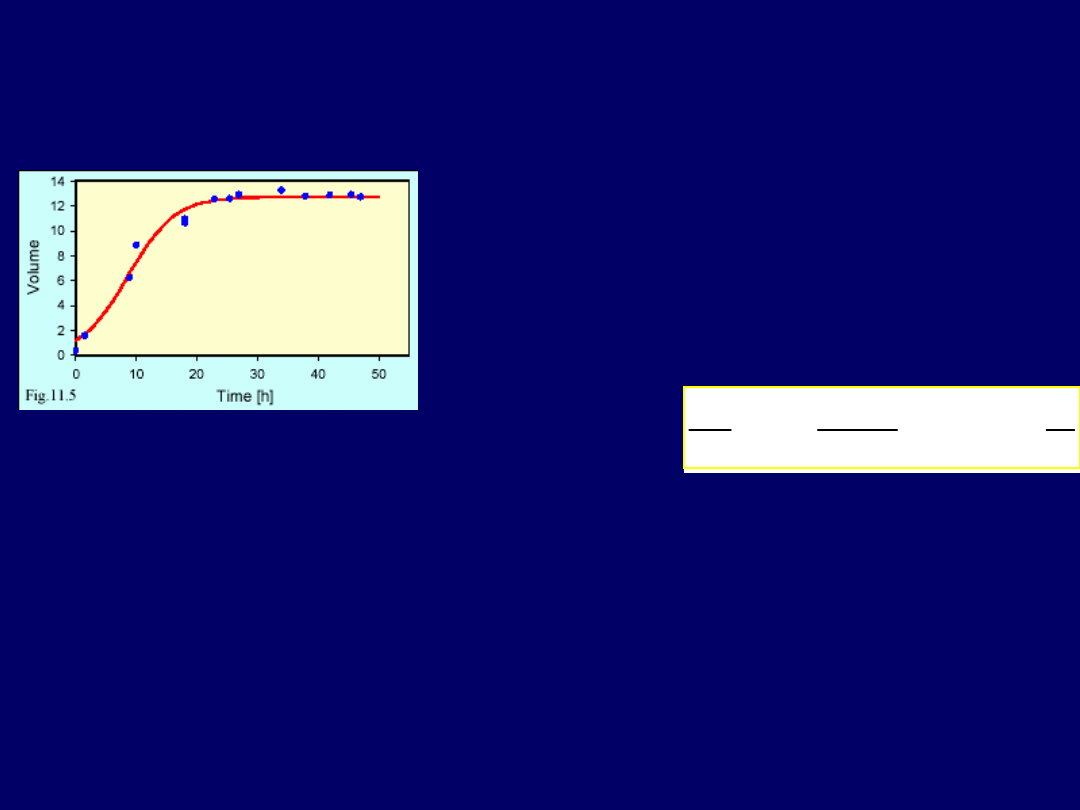
hodowanych w chemostacie, które
na początku doświadczenia rosły
szybko – zgodnie z wykładniczym modelem wzrostu
. Lecz
im wyższa
była wielkość populacji (objętość kolonii), tym wolniejszy był wzrost
drożdży
. Jak modelować taki proces? Bierzemy wtedy wyjściową,
wykładniczą funkcję wzrostu i
dodajemy
„drugie wyrażenie” („second term”), które
zmniejsza szybkość wzrostu populacji,
w miarę, jak N zbliża się do wartości gra-
nicznej
. Jeżeli określimy tę
górną granicę
wielkości populacji przez K
, to cały proces
możemy modelować równaniem:
Gdy N staje się
coraz większe,
K–N dąży do zera i drugi
czynnik [(K–N)/K] też dąży do zera
i szyb-
kość wzrostu populacji spada. Dla
niskiego N, komponent (K–N)/K
jest bliski 1
i cały proces przypomina nasz wyjściowy wzrost wykład-
niczy. Powyższe równanie jest najprostszą formą modelowania tego
typu procesu i zwane jest
logistycznym procesem wzrostu lub rów-
naniem Verhulst’a-Pearl’a
. Wykres na przeźroczu następnym pokazu-
je, że szybkość wzrostu jest b. mała dla małych wartości t i spada
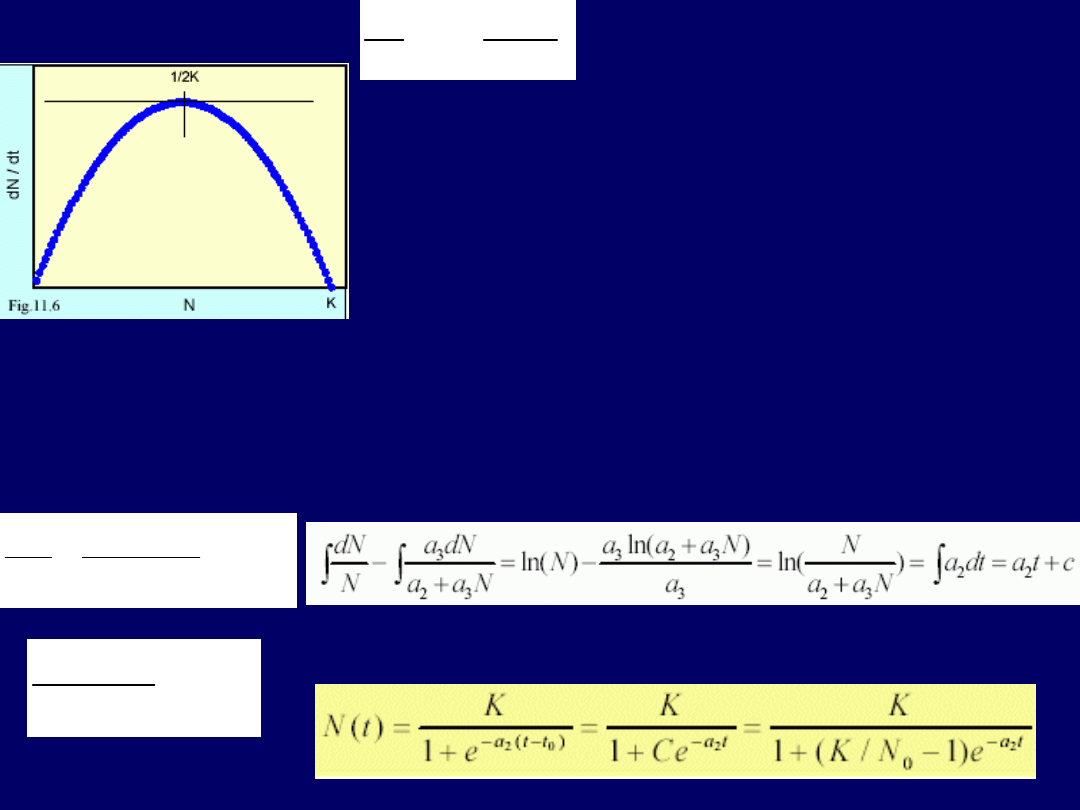
do zera dla N=K.
Maksymalna szybkość wzrostu występuje przy
N = K/2
. Powyższe logistyczne równanie wzrostu nie daje nam możli-
wości wyliczenia N. W tym celu musimy rozwiązać odpowiednie rów-
dN
K N
N
rN
rN rN
dt
K
K
nanie różniczkowe.
Aby je rozwiązać,
aproksymu-
jemy funkcję z jej rozwinięcia
w szereg Taylora
. Zakładamy, że funkcja N(t)
daje się rozwinąć w szereg Taylora. Dlatego
dN/dt musi być funkcją algebraiczną w formie:
dN(t)/dt = a
1
+ a
2
N(t) + a
3
N(t)
2
+ a
4
N(t)
3
...
Równanie wzrostu populacji ma
2 pierwiastki:
dla N=0 i dla N=K
. Najprostszym równaniem o
2 pierwiastkach jest
funkcja kwadratowa
. Dla-
tego możemy
„obciąć” rozwinięcie funkcji
w szereg Taylora już po 3-cim składniku
, uzyskując:
dN/dt = a
1
+ a
2
N + a
3
N
2
. Dla N = 0, uzyskujemy: a
1
= 0 i dlatego może-
my ten składnik opuścić:
dN/dt = a
2
N + a
3
N
2
. Na tym etapie,
dzielimy
całe równanie przez N(a
2
+ a
3
N)
. Dzięki temu staje się ono przeksz-
tałcone:
Taka postać jest
łatwiejsza do scałkowania
:
Stąd:
Teraz można przekształcić to równanie,
przyjmu-
jąc a
2
/a
3
= K
i w ten sposób uzyskać:
dN
K N
rN
dt
N
3
2
2
3
a dN
dN
a dt
N a
a N
2
2
3
a t
N
Ce
a
a N
W powyższym równaniu,
t
0
odpowiada populacji o wielkości N
0
w cza-
się t = t
0
.
K – „zdolność przepustowa
” („the carrying capacity”), jest
górną granicą N i można ją
łatwo obliczyć ustawiając dN/dt = 0
. Przy-kład
takiej funkcji jest przedstawiony na rys. poniżej. Dla przykładu z
drożdżami („2 rysunki wstecz”), udało się
dopasować krzywą funkcji o następują-
cym równaniu:
W genetyce lub w
epidemiologii,
zjawis-
ka mutacji lub infekcji
są zjawiskami masowymi
. Oznacza to, że
efekt końcowy określany jest przez cał-
kowitą liczbę czynników indukujących mutację lub infekcję
. Dlatego
powinniśmy znać sumę wszystkich tych czynników. Zakładamy, że cały
badany
proces daje się opisać za pomocą logistycznego modelu wzros-tu
.
Stąd, całkowita liczba czynników może być równa
całce, której od-powiada
pole powierzchni pod krzywą funkcji logistycznej
. Wyliczamy to za pomocą
programu matematycznego, uzyskując: Ostatni przykład
zmierza do
ogólnego rozwiązania
kwadratowego równania różnicz-
kowego I-szego rzędu
. Możemy
takie równanie zdefiniować następująco:
dy/dx = ay + by
2
. Na początku
oznaczamy punkt równowagi i uzyskujemy
dla y’=0: y(x) = –a/b
.
0,26
12,74
( )
1 9,32
t
N t
e
0
0
(
)
(
)
ln(1
)
1
a t t
a t t
K
K
e
dt
C
e
a
Korzystamy znów z programu matematycznego (Mathematica, wxMaxi-ma) i
uzyskujemy
rozwiązanie ogólne
: Wynik – tzw.
uogólnione logis-
tyczne równanie wzrostu
, a zarazem – rozwiązanie tzw.
autonomicznego równania różniczkowego
. Można spraw-
dzić, że dla wysokich wartości x, funkcja ta
asymptoty-
cznie zbliża się do wartości stanu równowagi: –b/a
. Przy
użyciu Mathematica, można przekształcić to rozwiązanie i przedstawić je w
kategoriach funkcji sinh i cosh.
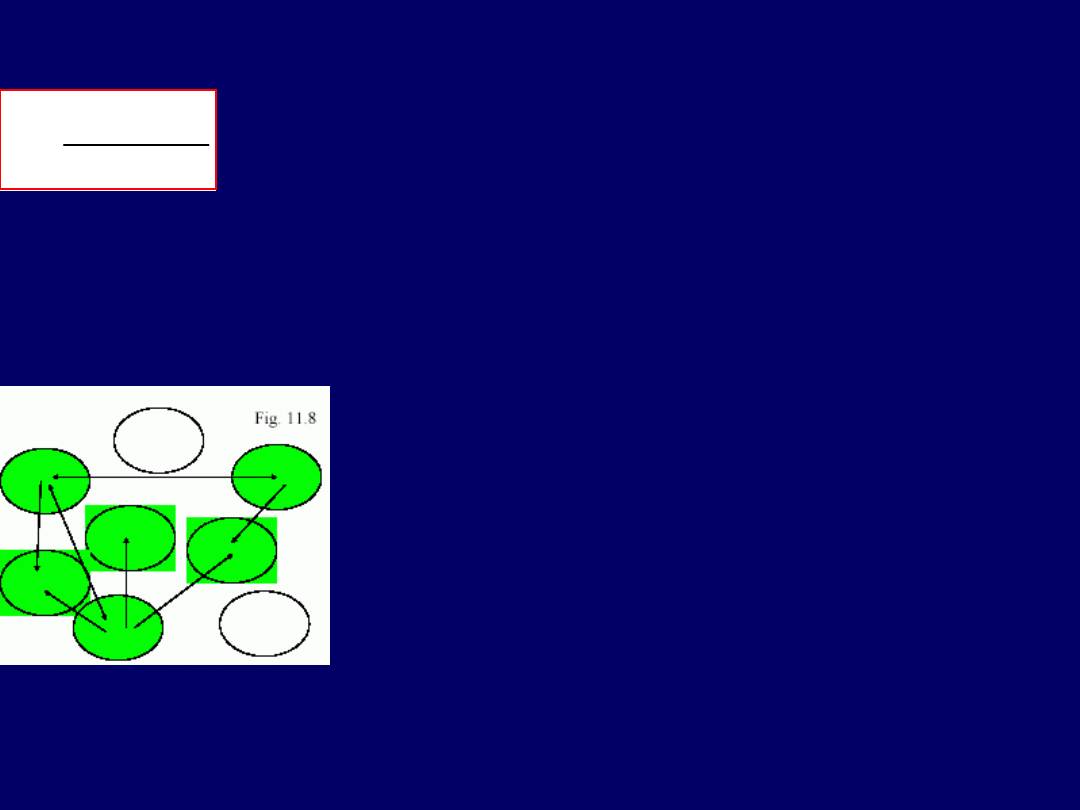
Inny przykład z genetyki i ekologii: załóżmy, że mamy
populację ga-
tunków zwierzęcych lub roślinnych, które zasiedlają „skrawki siedlis-ka”
(„habitat patches”)(
rys. – poniżej). Przyjmujemy, że wyjściowa częs-
tość zasiedlonych fragmentów siedliska wynosi
p
, a wyjściowa częstość niezasiedlonych frag-
mentów siedliska, to
q = (1-p)
. Częstości są zaw-
sze wybierane w taki sposób, aby ich całkowita
suma = 1 (lub 100%). Członkowie populacji
mig-
rują
. Są oni zdolni do
kolonizowania niezasied-
lonych fragmentów, ew populacje na zasiedlo-
nych fragmentach wymierają
. Taka populacja,
która jest rozdzielona na szereg „skrawków/frag-
mentów” siedliska, powiązanych ze sobą zjawiskami migracji,
nazywa-na jest
metapopulacją
. Problemem do rozwiązania jest
modelowanie
zmian p (liczba zasiedlonych fragmentów) w zależności od czasu
. Zmia-na p
w czasie, to:
dp/dt
. Definiujemy teraz
, jako tempo kolonizacji
1
ac ax
ac ax
ae e
y
be e

(imigracji) i
v
– lokalne tempo wymierania (obie wartości wyrażane jako
prawdopodobieństwo). Tak więc
zmiany w p mogą być teraz modelo-wane
jako suma 2 niezależnych procesów: zmian spowodowanych ko-lonizacją i
zmian wywołanych lokalnym wymieraniem
. Zakładamy, że
wzrost liczby
zasiedlonych skrawków jest proporcjonalny do faktycznej liczby zasiedlonych
skrawków (p) i do liczby pustych skrawków q=1-p
. Stąd:
dp/dt = p(1–p)
. Z
kolei
spadek p, powinien być proporcjonalny do lokalnej szybkości
wymierania
. Stąd:
dp/dt = –vp
. Teraz możemy mode-lować cały proces za
pomocą prostego równania:
dp/dt = p(1–p) – vp = –p
2
+ p(– v)
. Jest to
bdb. znany
model metapopulacji Richarda
Levinsa
, zaproponowany w 1969 r. Aby oznaczyć liczbę zasiedlonych
skrawków dla stanu równowagi, ustawiamy:
dp/dt = 0 i uzyskujemy
bezpośrednio: p = 1 – v/
.
Metapopulacja utrzymuje się przy życiu tylko
wtedy, gdy: v <
. Załóżmy, że v jest odwrotnie proporcjonalne do po-
wierzchni fragmentu siedliska (A). Oznacza to, że na większych skraw-kach,
prawdopodobieństwo wymierania spada, ze względu na większą populację.
Stąd
v 1/A
. Załóżmy dalej, że
szybkość kolonizacji () spa-da wraz ze
wzrostem odległości pomiędzy skrawkami (D)
. Stąd:
1/D
. Uzyskujemy
ostatecznie:
dp/dt = (1/D)p(1–p) – (1/A)p
. Wniosek I-szy:
liczba zasiedlonych siedlisk będzie wzrastała wraz ze wzrostem śred-niej
powierzchni skrawka
(z powodu spadku szybkości wymierania). Wn. II-gi:
liczba zasiedlonych siedlisk będzie spadała wraz ze wzrostem
odległości pomiędzy skrawkami
(ze względu na ograniczone procesy
kolonizacji). Przedostatnie równanie jest wg przyjętej terminologii
kwadratowym równaniem różniczkowym I-go rzędu
. Poniżej – podane
ogólne rozwiązanie równania ostatniego
:
Brak jest jednak początko-
wej wartości C. Przy
t = 0,
p = p
0
– częstość począt-
kowa. Biorąc to pod uwa-
gę,
uzyskujemy C
:
Logarytm w liczniku musi być > 0
.
Stąd właściwe rozwiązanie musi
spełniać warunek:
p
0
> 1 – v/
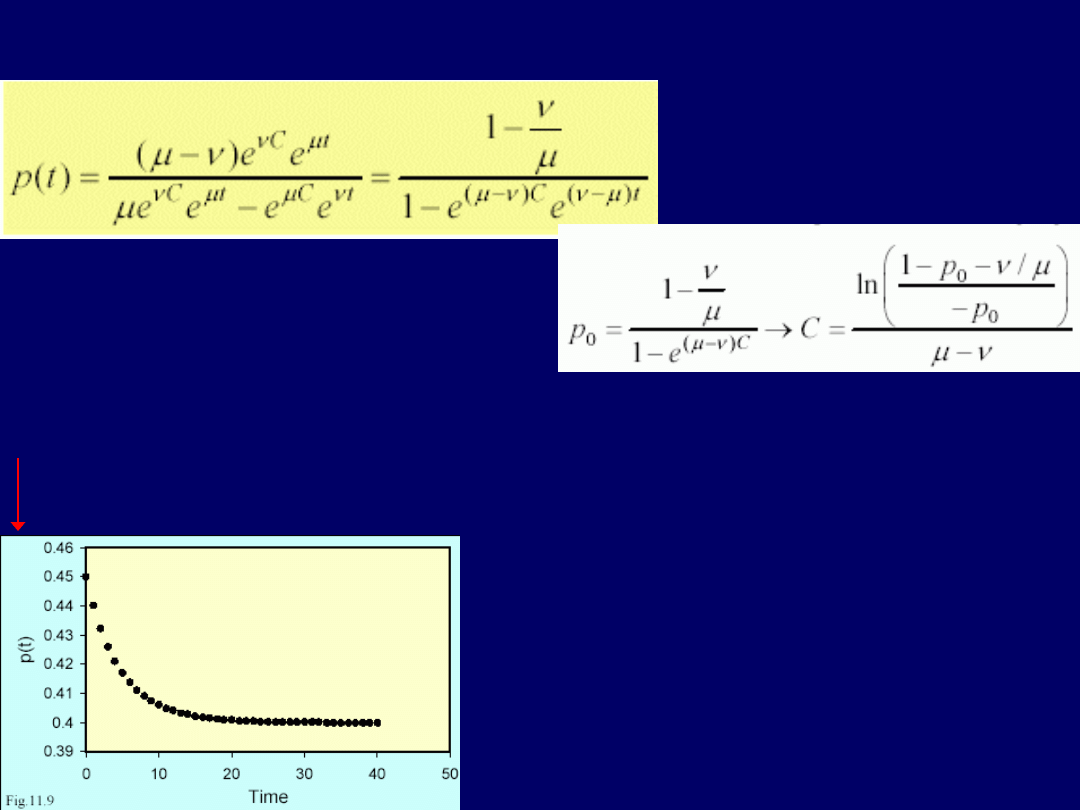
. Po-
niżej, przedstawiony jest wykres
zależności proporcji zasiedlonych skrawków [p(t)] w zależności od cza-
su: .
Tylko 10 pokoleń wystarcza do osiągnięcia stanu równowagi przy: p(t) =
1 – 0,3/0,5 = 0,4
. Stąd na podstawie warunków początkowych mo-żna wnosić,
że
40% fragmentów/skrawków siedliska zostanie zasied-
lonych
. Nie zawsze jednak jest to tak
proste i
całki albo przyjmują bardzo
skomplikowaną postać lub nawet nie
można wyrazić całki w tzw. formie zam-
kniętej
. Można spróbować metody uzys-
kiwania
przybliżenia całek
. Aby to zro-
bić,
rozwijamy funkcję w szereg Taylora
i „obcinamy” szereg
przy odpowiednim

wyrazie.
Całkę można wtedy łatwo wyliczyć ręcznie
, ponieważ będzie
potrzebna następująca całka: Dla
logistycznego równania wzros-
tu
, program Mathematica wylicza
automatycznie
rozwinięcie w sze-
reg Taylora
. „Obcinamy” po 3-cim
wyrazie i uzyskujemy następujące
przybliżenie całki:
Można porównać wyniki
: dokładne rozwiązanie dało dla a = 1, K = 1
i t
0
= 0 pomiędzy t = 0 a t = 3 wartość N = 2,355. Przybliżenie dało:
N = 1,45. Stąd nasz
błąd wynosi: (2,355 – 1,45)/2,355 = 0,38 = 38% (..nie
najlepiej.....!)
.
Przy aproksymacji z wykorzystaniem rozwinięcia funkcji w szereg Taylora,
wynik znacznie zależy od tego
jak szybko szereg osiąga zbież-ność
; szybka
zbieżność oznacza wyniki bliskie dokładnego rozwiąza-nia. Można to
sprawdzić np. sporządzając
wykres zależności sumy wy-razów szeregu od
kolejnych numerów wyrazów
.
W powyższym przykładzie
aproksymacja do szeregu Taylora zawiod-ła
.
Dlaczego? Ponieważ
szereg jest naprzemienny
(wyrazy o znakach: +, –, + –,
+, –, etc.). „Obcięcie” za wyrazem o niewielkim numerze kolej-
nym (porządkowym) pociąga za sobą
bardzo wysokie błędy
. Praktyka
sugeruje, że powinniśmy wykorzystywać jako aproksymacje tylko te
1
(
)
(
)
1
n
n
a
a x b dx
x b
C
n
0,5
0,5
3
6
4
7
0,5
3
0
0
0
1
1
0,492
2
8
8 56
|
x
x
x
x
x dx
dx x
0
3
3
3
2
4
0
0
0
0
0
(
)
(
)
(
)
(
)
(
)
(
)
1
4
4
48
4
8
192
a t t
K
K aK
a K
K
aK
a K
dt
t t
t t dt
t t
t t
t t
C
e

rozwinięcia w szereg, które zawierają
wyrazy wyłącznie dodatnie lub ujemne,
nadają się do wykorzystania jako aproksymacje funkcji
. Roz-
winięcia wielu znanych funkcji często dają szeregi, które „
z trudem są
zbieżne
” (wyjątek – prosta funkcja wykładnicza: y = e
x
). Inny przykład:
funkcja
y = (1 – x
3
)
0,5
, w zakresie: 0 < x < 1. Mathematica wylicza b.
skomplikowaną całkę. Jednakże
szereg Taylora jest bardzo prosty i osiąga
zbieżność bardzo szybko
. Całka w granicach od 0 do 0,5 wynosi:
Rozwiązanie
numeryczne przy użyciu Mathematica
dało prawie identy-czny
wynik:
0,492041 + 3,33067 10
–16
.
0,5
0,5
3
6
4
7
0,5
3
0
0
0
1
1
0,492
2
8
8 56
|
x
x
x
x
x dx
dx x

Wskazówki do wykonania zadań praktycznych ćw. 5.
• Wskazówki do zadania 1:
Ogólna postać równania różniczkowego, to:
dy/dx = f(x)y
. Przystępu-jąc do
jego rozwiązania, zazwyczaj
dzielimy obie jego strony przez y i mnożymy
przez dx
, wskutek czego uzyskujemy:
dy/y = f(x)dx
.
Całkujemy teraz obie
strony
, uzyskując:
ln(y) = f(x)dx + C
. Po prze-kształceniu:
y = Ce
f(x)dx
. C
można wyznaczyć, przyjmując x
0
= 0;
wtedy (o ile wykładnik przy e zostanie sprowadzony do 0): C = y
0
i
ostatecznie:
y = y
0
e
f(x)dx
. Dla I-szego równania:
dy/dx = ay | :y *dx
dy/y = adx
dy/y =
adx
ln(y) = ax + C
y = Ce
ax
= y
0
e
ax
.
Dla II-go równania:
dy/dx = y/x | :y *dx
dy/y = (1/x)dx
dy/y =
(1/x)dx
ln(y) = ln(x) + C
y = Cx
.

Wskazówki do zadania 2:
Uruchamiamy program wxMaxima. Pierwsze równanie różniczkowe:
dy/dx =
e
x–y
, wprowadzamy do pola „INPUT:” następująco:
‘diff(y,x)=%e^(x-y)
, co widać na zrzucie ekranu: Następnie naciska-
my
<Enter>,
uzyskując:
Po tym, wracamy do pola przyciskami
i
klikamy w przycisk „Solve ODE”
, który
otwiera
okno dialogowe rozwiązywania równań różniczkowych
:
Klik
Wynik – okno dialogowe –
na następnym przeźroczu
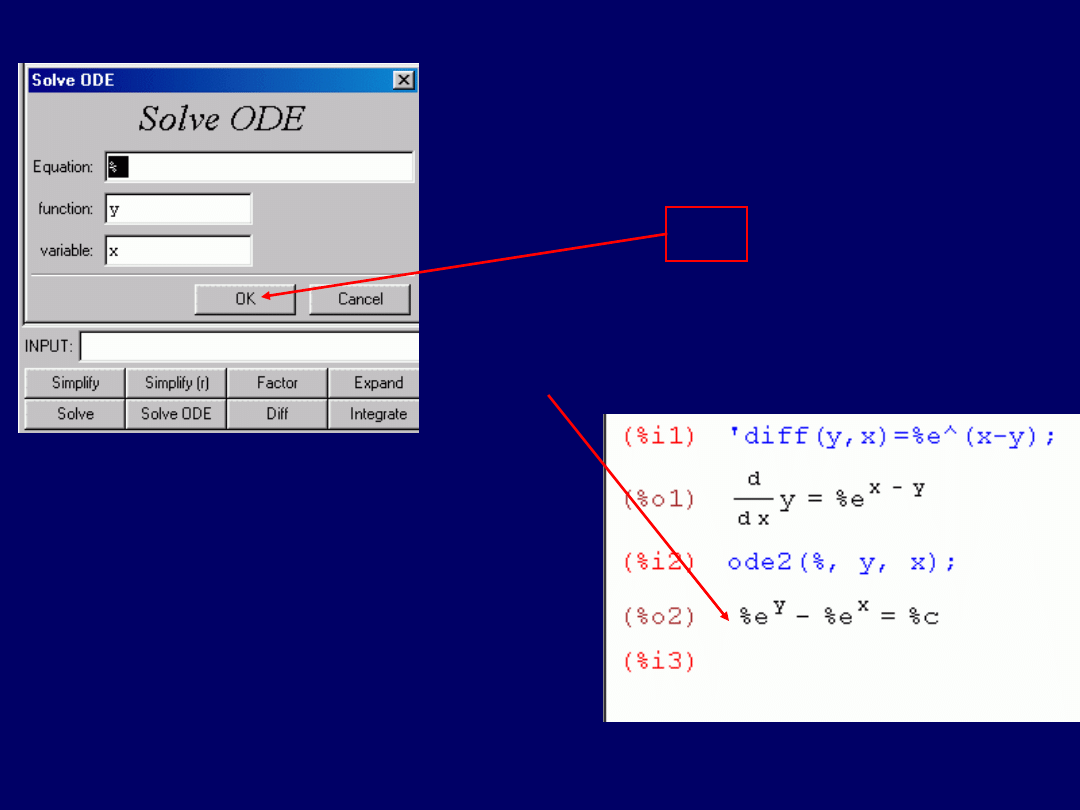
Okno dialogowe rozwiązywania równań różniczkowych
:
Akceptujemy wszystkie
opcje domyślne
i klikamy w przycisk
OK
. Znak „%” w
w polu „Equa-
Klik
ion” oznacza
wzięcie do obliczeń poprzedniego wyni-
ku.
W następstwie tego, uzyskujemy
rozwią-
zanie
:
Nie jest to jednak rozwiązanie typu:
y = f(x)
, o jakie nam chodzi.
Aby uzyskać tego typu rozwiązanie,
należy uruchomić w wxMaxima
komendę: „
Solve(y)
”. Można ją wpro-
wadzić bezpośrednio do pola:
„INPUT:”. Można też kliknąć w przy-
cisk „
Solve
”, a po ukazaniu się okna
dialogowego rozwiązywania równań,
zamienić x na y w polu „for variable(s):”
(następne przeźrocze).
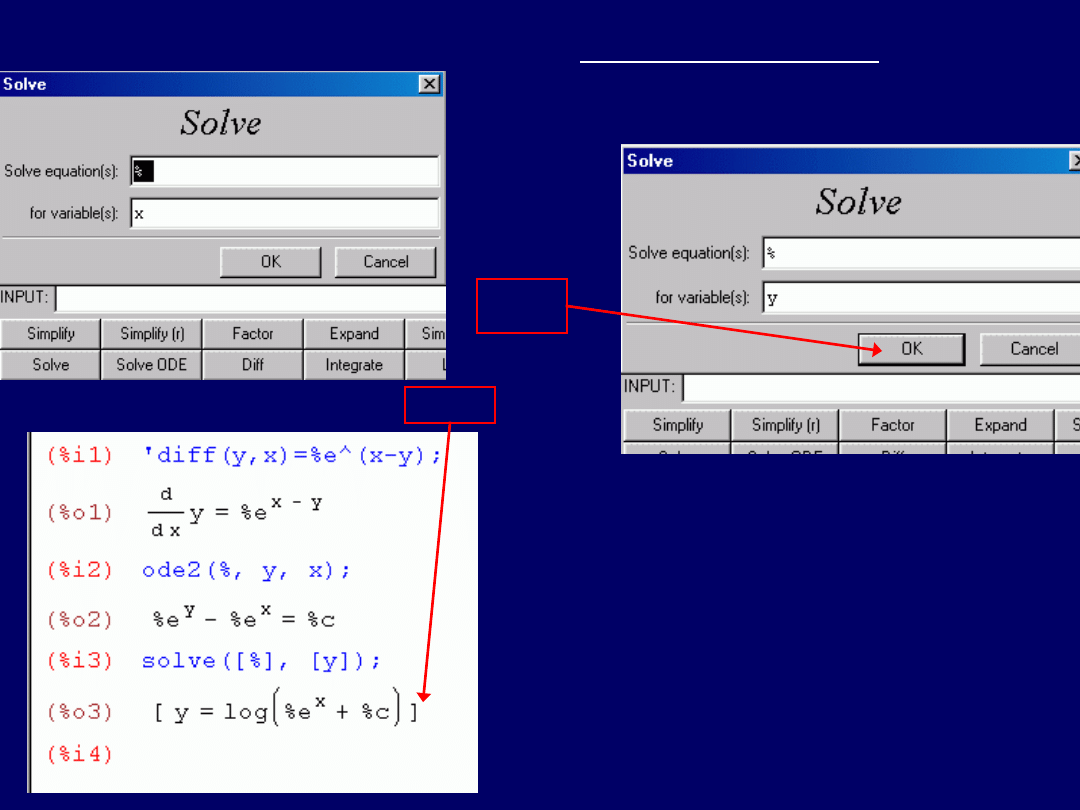
Okno dialogowe rozwiązywania równań
(nie różniczkowych!!!!!):
Zamieniamy:
x y
, uzyskując:
Klik
Ostatecznie, uzyskujemy wynik:
Wynik ten, należy odczytać jako:
y = ln(e
x
+ C)
.
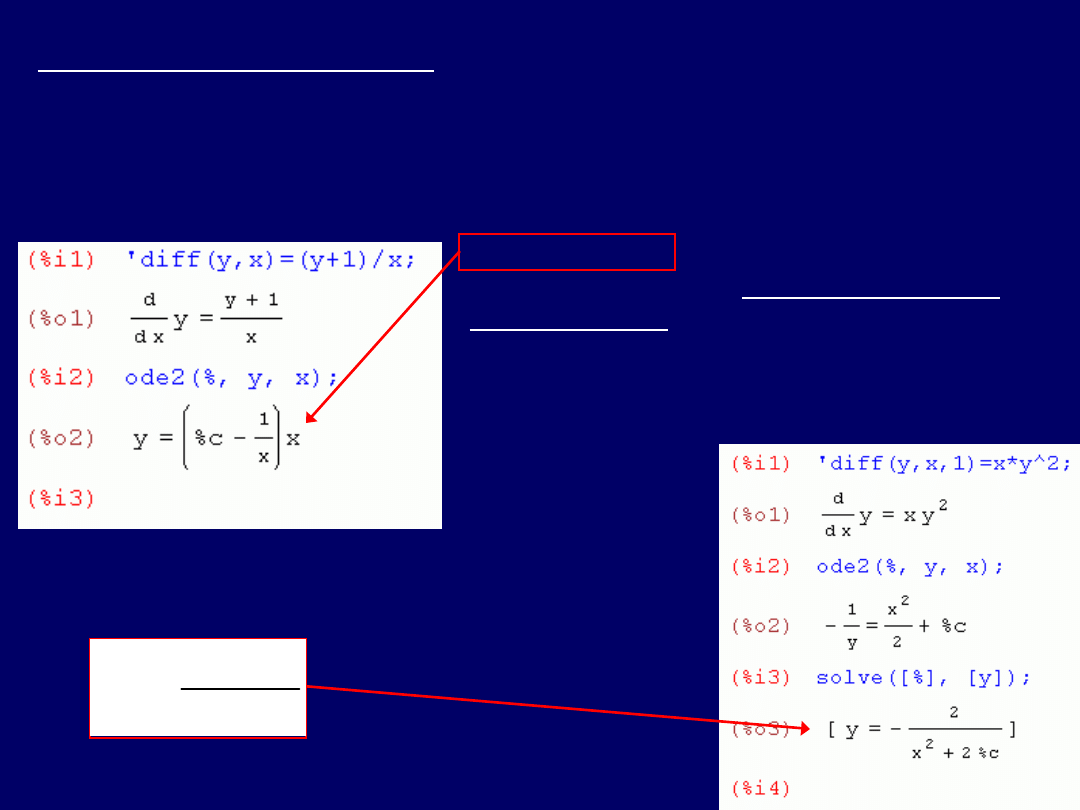
Drugie równanie różniczkowe:
dy/dx = (y+1)/x
; wszystkie czynności
przeprowadzamy
analogicznie, jak przy rozwiązywaniu równania
poprzedniego
(poprawne wprowadzenie:
‘diff(y,x)=(y+1)/x
) – z tym, że
pomijamy ostatni etap „solve(y)”
(3 ostatnie zrzuty ekranu dla
równania poprzedniego), uzyskując od razu
właściwe rozwiąza-nie
:
Rozwiązanie to odczytujemy:
y = (C – 1/x)x
.
Przy rozwiązywaniu trzeciego równania
różniczkowego:
dy/dx = xy
2
(r-nie kwad-
ratowe; popr. wprowadzenie:
‘diff(y,x)=x*y^2
), postępujemy analo-
gicznie, jak w przypadku r-nia I-go:
Rozwiązanie to, należy odczytać:
2
2
2
y
x
C

Czwarte równanie różniczkowe:
dy/dx = –ay
n
; czynności rozwiązywa-nia
ogólnie wykonujemy tak, jak dla równań poprzednich
(poprawne
wprowadzenie:
‘diff(y,x)=–a*y^n
).
Jednakże po urucho-
mieniu „
Solve ODE
”, po-
jawia się pytanie: „
Is
n – 1 zero or nonzero?
”
Odpowiadamy w polu:
„INPUT:” „
nonzero
” I
wciskamy <Enter>. Po-
jawia się kolejne pyta-
nie: „
Is n an integer?
”
(„Czy n jest liczbą cał-
kowitą?). Odpowiadamy
: „
yes
” <Enter>.
Po tym pojawia
się
ostateczne rozwią-
zanie
, które odczytuje-
my:
1
1 n
y (anx ax Can Ca)
-
=
-
+
-
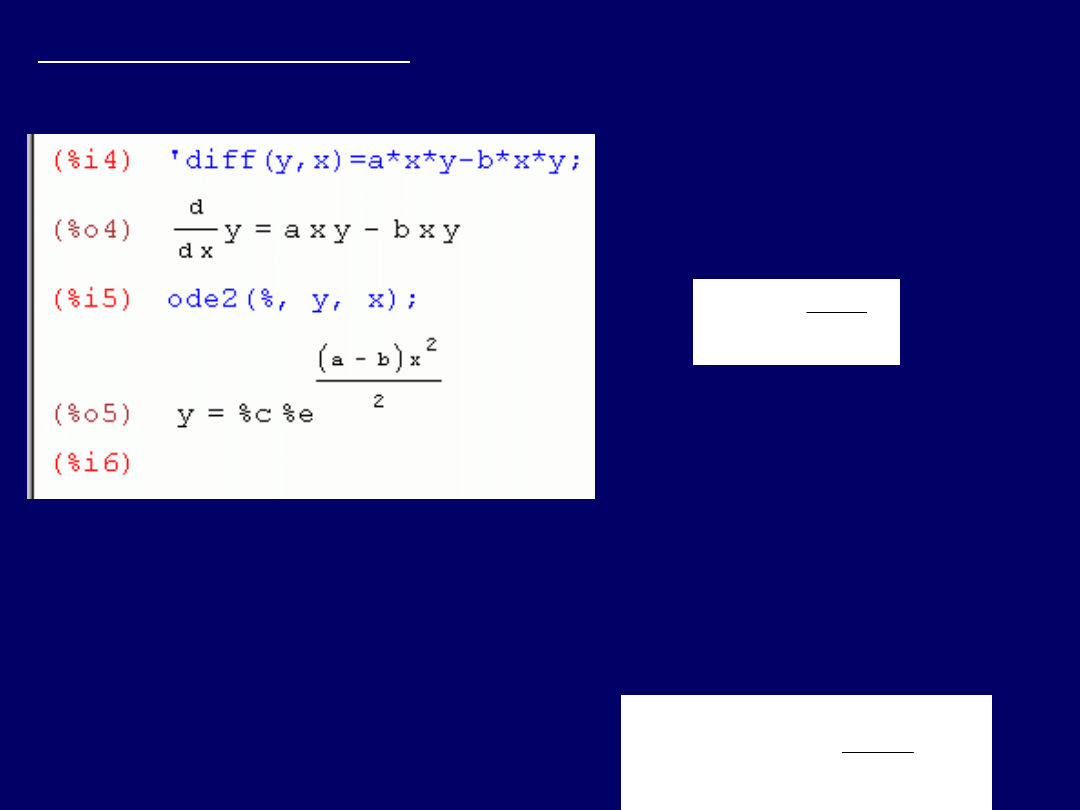
Piąte równanie różniczkowe:
dy/dx = axy – bxy
;
powtarzamy czynno-ści
rozwiązywania
, jak w przypadku równań poprzednich. Poprawne
wprowadzenie:
‘diff(y,x) = a*x*y – b*x*y
. Nie jest konieczne korzys-
tanie z opcji „Solve(y)”.
Ostateczne rozwiązanie
:
Biologiczne zastosowanie
po-
wyższego równania różnicz-
kowego:
modelowanie pro-
cesów ewolucji
(specjacja,
wymieranie..) dowolnego taksonu organizmów żywych (zwierzęta
lub rośliny) – przedstawione
na początku części teoretycznej
. Zna-czenie
symboli (zmiana):
y N
(liczba gatunków w czasie t),
C N
0
(stała C:
liczba gatunków w czasie t
0
= 0),
a c
[prawdopodobień-stwo (częstość)
specjacji],
b e
[prawdopodobieństwo (częstość) wymierania],
x t
(czas).
Po zamianie
:
.
2
(a b)
x
2
y Ce
-
=
2
0
c e
N N exp
t
2
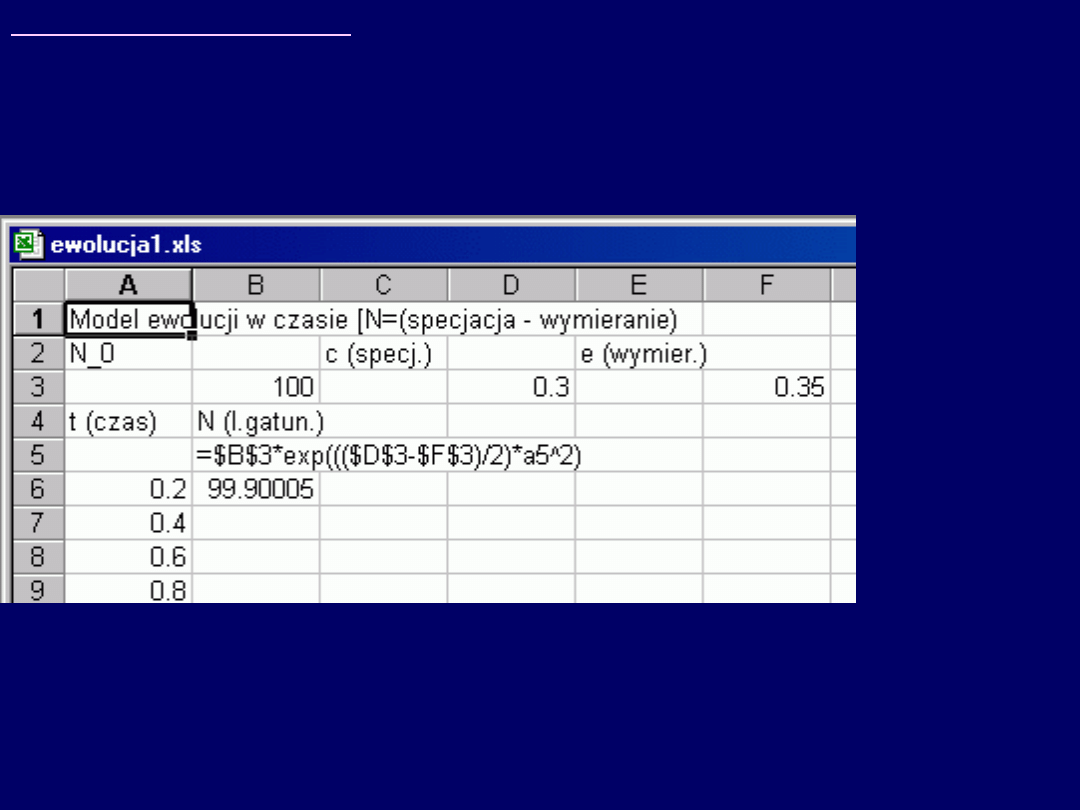
Wskazówki do zadania 3:
Model procesu ewolucji:
N = N
0
exp[((c – e)/2)t
2
],
został przedstawiony
(włącznie z podaniem opisów zmiennych i parametrów)
na przeźroczu
poprzednim
. Plik „ewolucja1.xls”, dostępny ze stron internetowych ćwiczeń,
umożliwia
automatyczne zmiany podstawowych parametrów równania funkcji
N(t): N
0
, c i e
.
Po pobraniu i zapisie pliku, dokończeniu obliczeń i sporządzeniu wykre-su
punktowego (XY), możliwe są łatwe
zmiany parametrów równania i
obserwacje wynikłych stąd zmian krzywych N(t)
. Najważniejszą rolę
odgrywają tu
zmiany wartości c i e – a w szczególności – wzajemna proporcja
c do e
. Przykładowe zrzuty ekranów – patrz dalej.
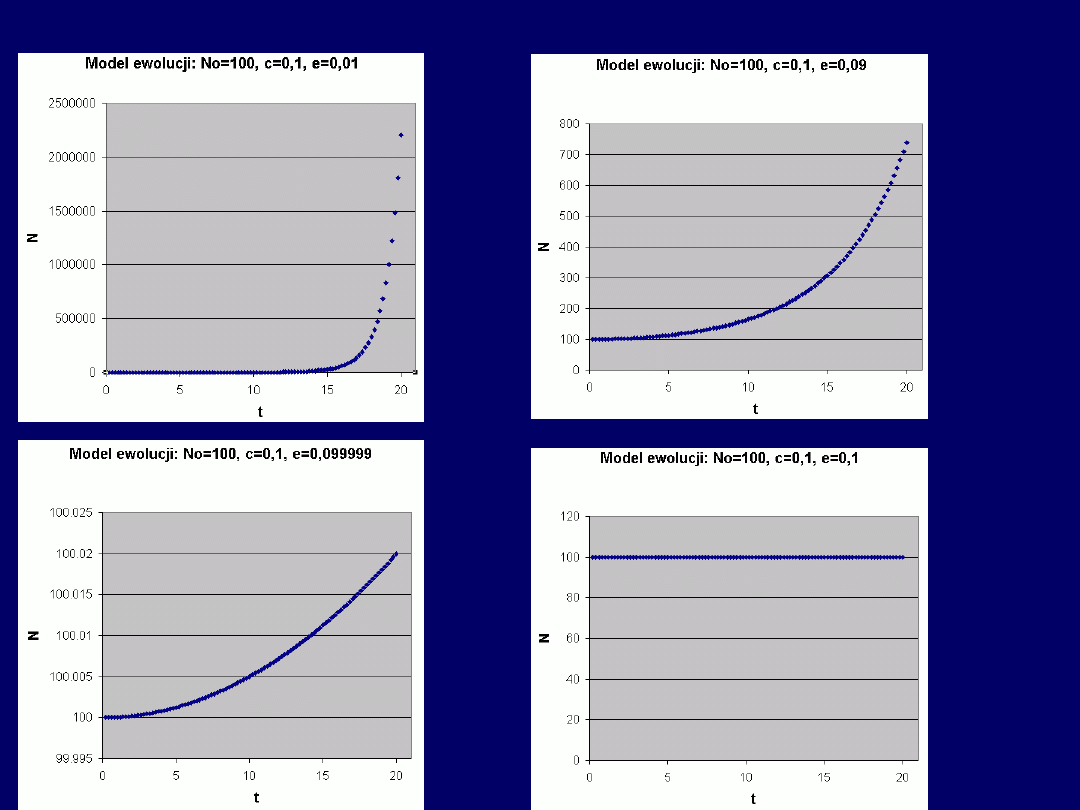
Przykłady –
funkcja
rosnąca
(1, 2 i 3) i
stała
(4)
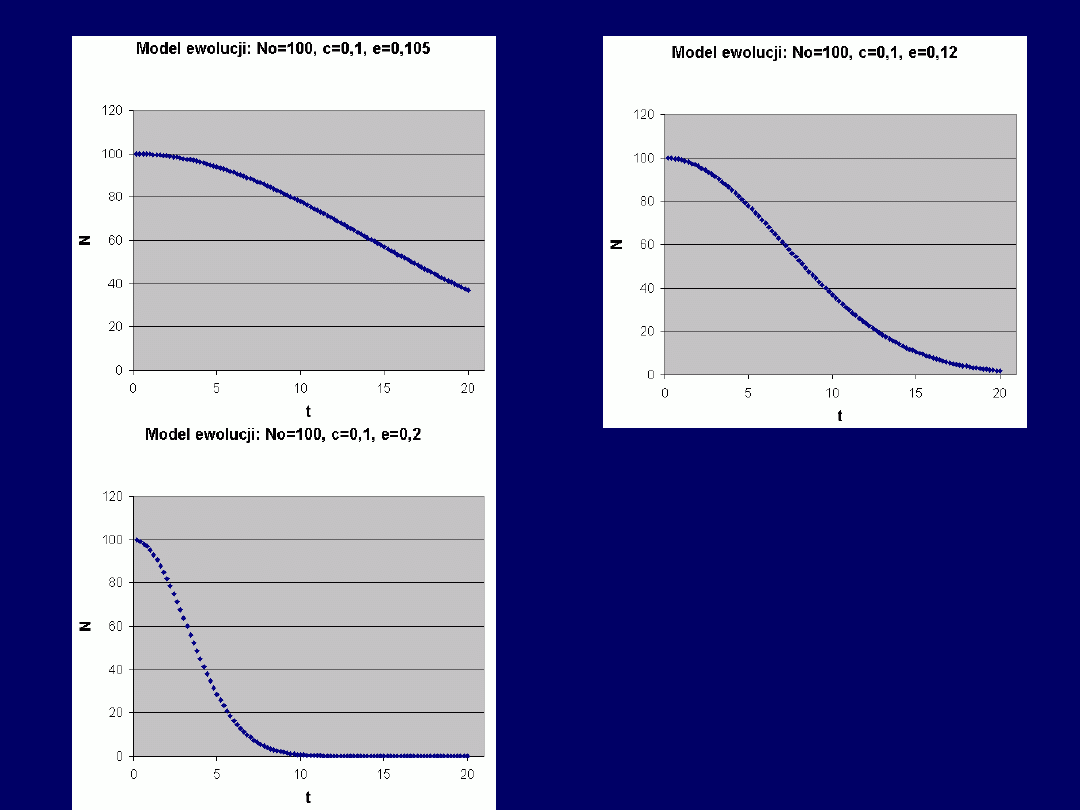
Przykłady, cd. –
funkcja
malejąca
:
Przy
stałym N
0
, parametry równa-
nia mają następujący
wpływ na
przebieg krzywej
N(t):
c > e: funkcja rosnąca
c = e: funkcja stała (N = N
0
)
c < e: funkcja malejąca
.

Wskazówki do zadania 4:
Logistyczne równanie wzrostu:
N(t) = K/[1+C*exp(–a
2
t)]
, zostało przed-
stawione (włącznie z podaniem opisów zmiennych i parametrów)
w
podstawowej instrukcji do ćwiczenia na WWW
. Plik „logist1.xls”,
dostępny ze stron internetowych ćwiczeń, umożliwia
automatyczne
zmiany podstawowych parametrów równania funkcji N(t): K, C i a
2
.
Po pobraniu i zapisie pliku, dokończeniu obliczeń i sporządzeniu wykre-
su punktowego (XY), możliwe są łatwe
zmiany parametrów równania
i obserwacje wynikłych stąd zmian krzywych N(t)
. Najważniejszą rolę
odgrywają tu
zmiany wartości C i a
2
– a w szczególności – wzajemna
proporcja C do a
2
. Przykładowe zrzuty ekranów – patrz dalej.
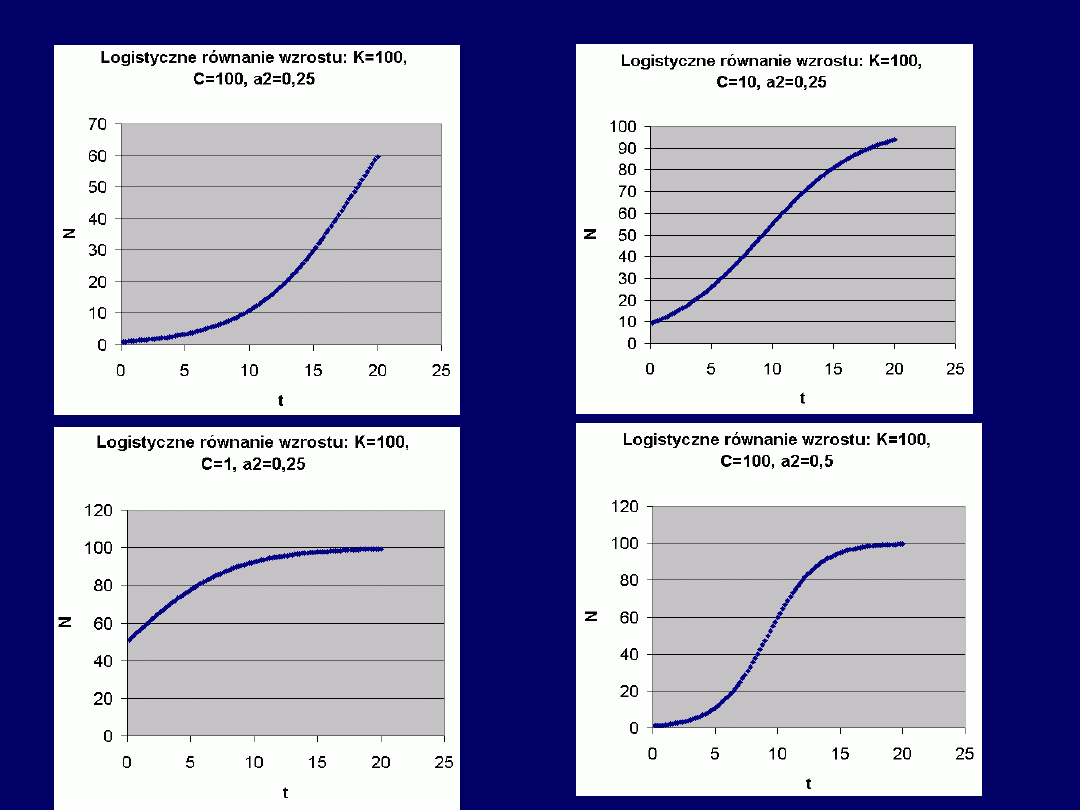
Przykłady (
1-4
):
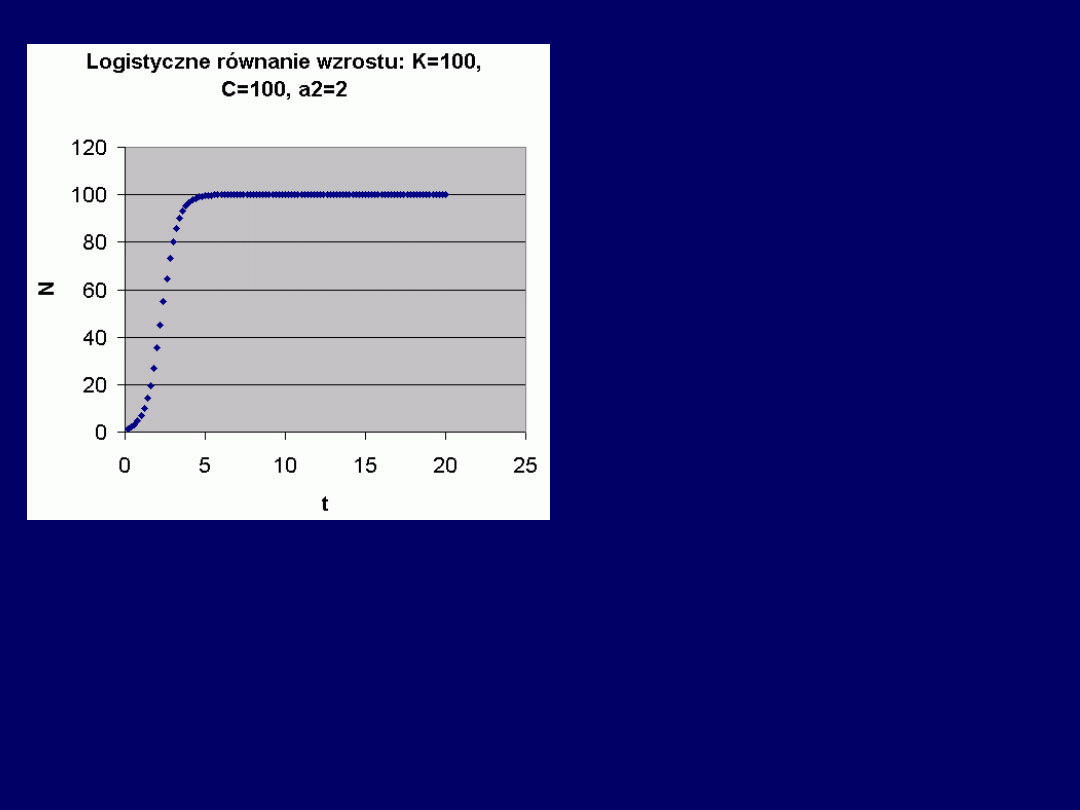
Przykład
5
(ostatni):
Wpływ parametrów równania lo-
gistycznego na kształt krzywej
(z wyjątkiem K)
:
C – z 1 strony decyduje o tym,
„która część krzywej” będzie
widoczna na wykresie
(jeśli wyso-
kie – to początkowa; jeśli niskie
– końcowa),
a z II-giej – o wartoś-
ci początkowej liczby organiz-
mów (N
0
)
: im niższe – tym wyższa.
Wynika to z zależności:
C = K/N
0
–1
.
Stała
a
2
decyduje o tym, jak szy-
bko krzywa dąży do swej wartości
maksymalnej
(asymptotycznie do K): im wyższa, tym szybciej (~
współ-
czynnik kierunkowy
krzywej logistycznej).

Wskazówki do zadania 5:
Model opisujący podawania glukozy do krwioobiegu w zależności od czasu
G(t) = c/a + [G(0) – c/a]exp(–at)
, zostało przedstawione (włącznie z
podaniem opisów zmiennych i parametrów)
w podstawowej instrukcji do
ćwiczenia na WWW
. Plik „glukoza1.xls”, dostępny ze stron internetowych
ćwiczeń, umożliwia
automatyczne zmiany podstawowych parametrów
równania funkcji G(t): G(0), c i a
.
Po pobraniu i zapisie pliku, dokończeniu obliczeń i sporządzeniu wykre-
su punktowego (XY), możliwe są łatwe
zmiany parametrów równania
i obserwacje wynikłych stąd zmian krzywych G(t)
. Najważniejszą rolę
odgrywają tu
zmiany wartości c i a – a w szczególności – wzajemna
proporcja c do a
. Przykładowe zrzuty ekranów – patrz dalej.
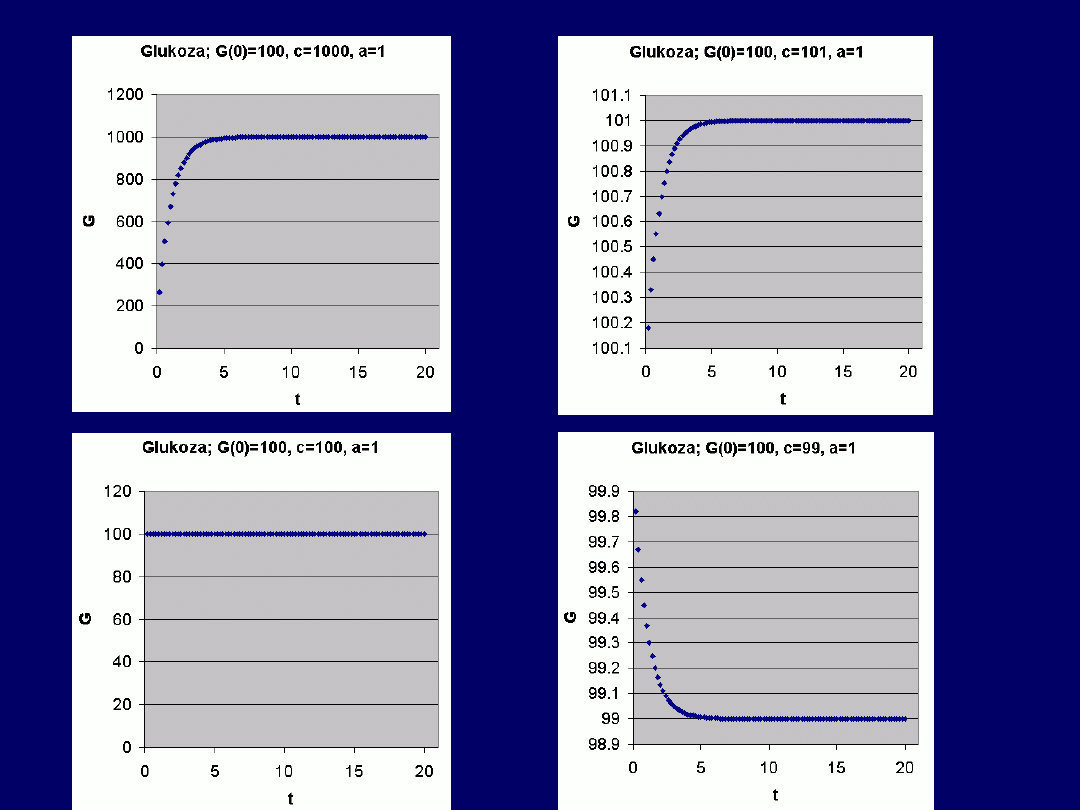
Przykłady –
funkcja
rosnąca
(1, 2),
stała
(3) i
malejąca
(4)
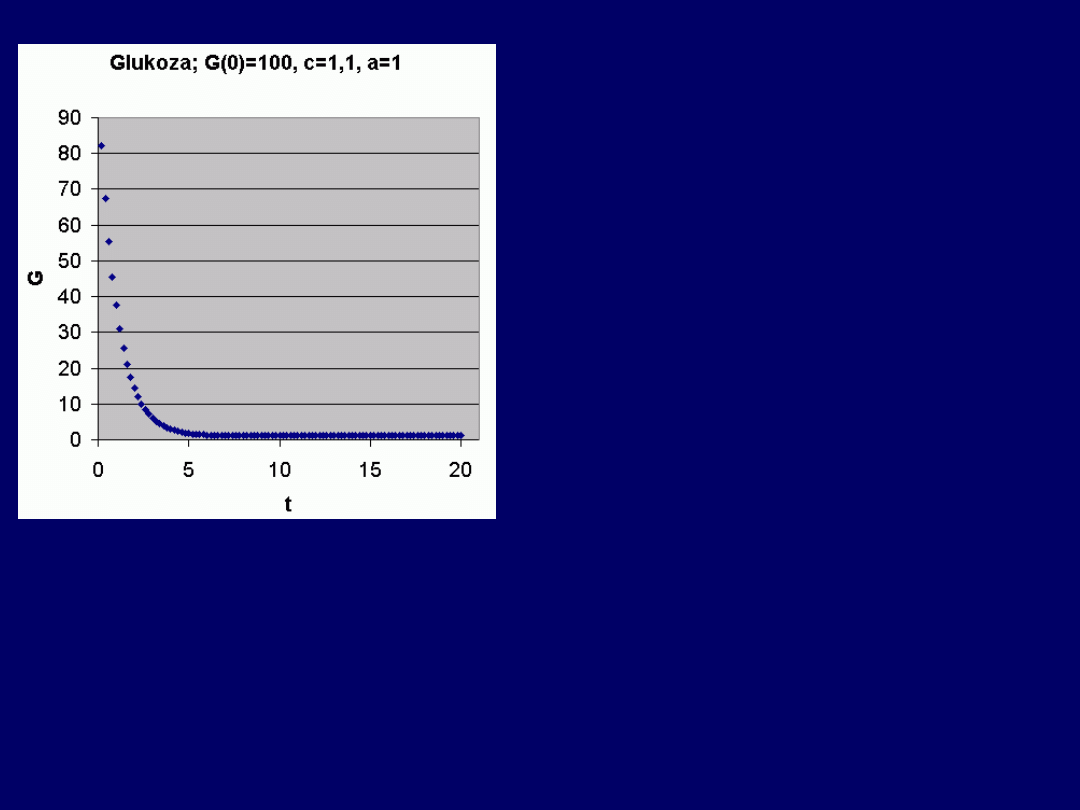
Przykład 5 –
funkcja
malejąca
:
Wpływ parametrów równania
na kształt krzywej:
parametry równania mają następują-
cy
wpływ na przebieg krzywej
G(t):
G(0) > c: funkcja malejąca
G(0) = c: funkcja stała [G = G(0)]
G(0) < c: funkcja rosnąca
Bez względu na wartości G(0),
funkcja
asymptotycznie dąży
do wartości równej:
G(t) = c/a
(dla wysokich wartości
t)

Wskazówki do zadania 6:
Model metapopulacji, opisujący zmiany w częstości zasiedlonych skrawków
ekosystemuw zależności od czasu:
p(t)=(1–(v/))/[(1–exp((–v)C)*exp((v–)t))]
, został przedstawiony (włącznie z
podaniem opisów zmiennych i parametrów)
w podstawowej instrukcji do
ćwiczenia na WWW
. Plik „metapop1.xls”, dostępny ze stron internetowych
ćwiczeń, umożliwia
automatyczne zmiany podstawowych parametrów
równania funkcji p(t): C, i v
.
Po pobraniu i zapisie pliku, dokończeniu obliczeń i sporządzeniu wykre-
su punktowego (XY), możliwe są łatwe
zmiany parametrów równania
i obserwacje wynikłych stąd zmian krzywych p(t)
. Najważniejszą rolę
odgrywają tu
zmiany wartości
i
v
– a w szczególności – wzajemna
proporcja
do
v
. Przykładowe zrzuty ekranów – patrz dalej.
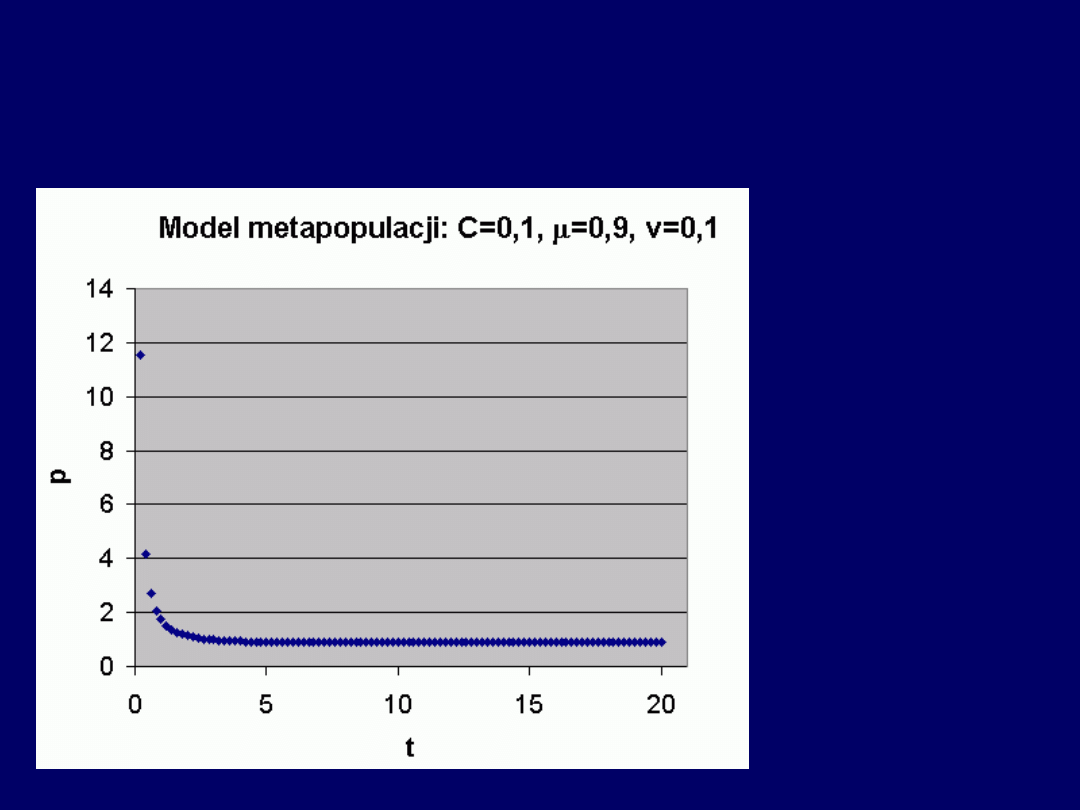
Przykład a) wysokie i niskie v
:
Przy wysokim i niskim v – populacja
nie wymiera całkowicie
, lecz
dąży do stanu równowagi
:
p(t) = 1 – v/
. W konkretnym przypadku, stan
równowagi jest osiągany dla:
p(t) = 1 – 0,1/0,9 = 1 – 0,1111 = 0,8889
.
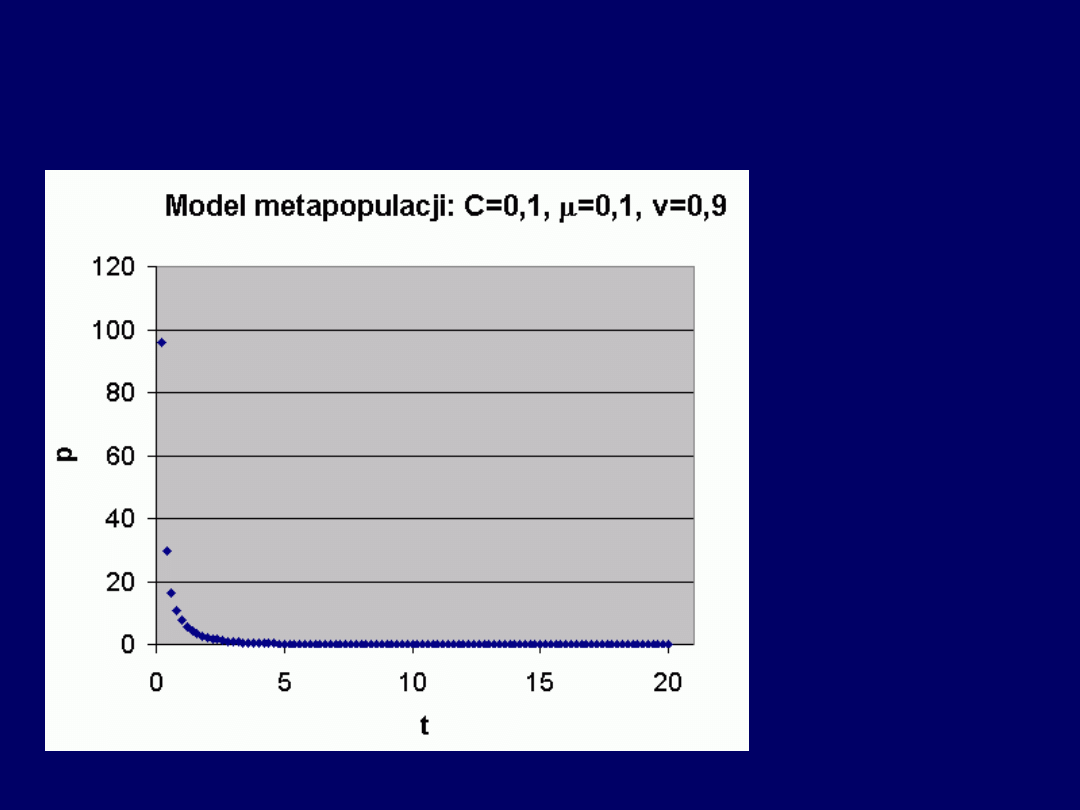
Przykład b) niskie i wysokie v
:
Przy niskim i wysokim v – populacja
wymiera całkowicie
, po odpo-wiednio
długim czasie
(w niniejszym przykładzie, model osiąga
warto-ści skrajnie
niskie
dla względnie
wysokich wartości czasu
).

Dziękuję
za uwagę ;-)
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
Wyszukiwarka
Podobne podstrony:
Wyklad2 mat
Mat 10 Ceramika
Mat dla stud 2
Wyklad7 mat
mat skale pomiarowe
logika mat
Magn mat
7Komunikacja org mat
mat bud 006 (Kopiowanie) (Kopiowanie)
Materialy do seminarium inz mat 09 10 czesc III
mat bud 102 (Kopiowanie) (Kopiowanie)
mat 2013 k11
Mat 3
MB2 mat pom 1 id 289843 Nieznany
mat dp14 mat dp14
mat bud 067 (Kopiowanie) (Kopiowanie)
więcej podobnych podstron