
FIZJOLOGIA
UKŁADU
KRĄŻENIA
FIZJOLOGIA
UKŁADU
KRĄŻENIA
Liana Puchalska, Stanisław Kowalewski
Liana Puchalska, Stanisław Kowalewski
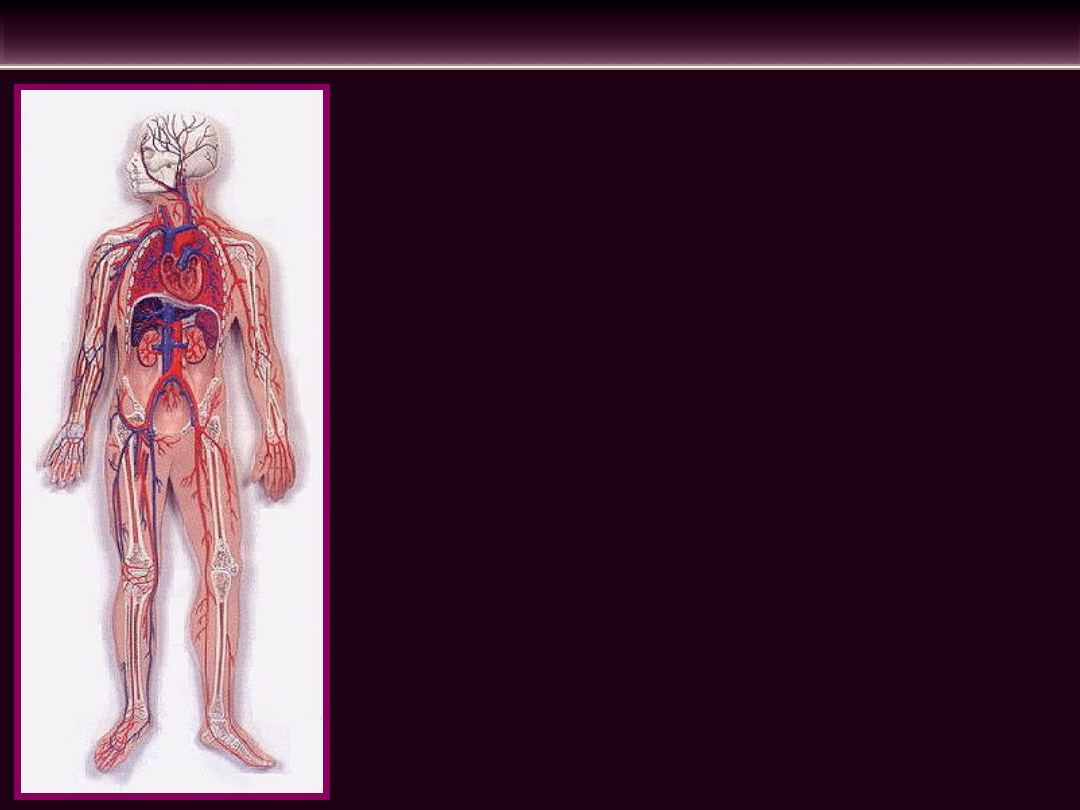
Układ sercowo-naczyniowy ze względu
na jego czynność dzieli się na:
• Serce składające się z dwóch przedsionków
(prawego i lewego) i dwóch komór (prawej i
lewej)
• Tętnice i żyły krążenia dużego, które tworzą
dwa zbiorniki: zbiornik tętniczy duży i
zbiornik żylny duży
• Tętnice i żyły krążenia małego (płucnego),
które tworzą dwa zbiorniki: zbiornik tętniczy
płucny i zbiornik żylny płucny
• Dwie sieci naczyń włosowatych
- pomiędzy zbiornikiem tętniczym dużym i
zbiornikiem żylnym dużym
- pomiędzy zbiornikiem tętniczym płucnym i
zbiornikiem żylnym płucnym
Układ sercowo-naczyniowy ze względu
na jego czynność dzieli się na:
• Serce składające się z dwóch przedsionków
(prawego i lewego) i dwóch komór (prawej i
lewej)
• Tętnice i żyły krążenia dużego, które tworzą
dwa zbiorniki: zbiornik tętniczy duży i
zbiornik żylny duży
• Tętnice i żyły krążenia małego (płucnego),
które tworzą dwa zbiorniki: zbiornik tętniczy
płucny i zbiornik żylny płucny
• Dwie sieci naczyń włosowatych
- pomiędzy zbiornikiem tętniczym dużym i
zbiornikiem żylnym dużym
- pomiędzy zbiornikiem tętniczym płucnym i
zbiornikiem żylnym płucnym
ANATOMIA UKŁADU KRĄŻENIA
ANATOMIA UKŁADU KRĄŻENIA

UNERWIENIE UKŁADU KRĄŻENIA
UNERWIENIE UKŁADU KRĄŻENIA
UNERWIENIE SERCA
1. Unerwienie współczulne
• Lewy pień współczulny unerwia komory
• Prawy
pień
współczulny
unerwia
przedsionki
i
węzeł
zatokowo-
przedsionkowy
2.
Unerwienie przywspółczulne
– nerwy
błędne
• Unerwienie
węzła
zatokowo
-przedsionkowego,
przedsionkowo
-
komorowego oraz mięśniówki roboczej
przedsionków
UNERWIENIE SERCA
1. Unerwienie współczulne
• Lewy pień współczulny unerwia komory
• Prawy
pień
współczulny
unerwia
przedsionki
i
węzeł
zatokowo-
przedsionkowy
2.
Unerwienie przywspółczulne
– nerwy
błędne
• Unerwienie
węzła
zatokowo
-przedsionkowego,
przedsionkowo
-
komorowego oraz mięśniówki roboczej
przedsionków

UNERWIENIE UKŁADU KRĄŻENIA
UNERWIENIE UKŁADU KRĄŻENIA
RECEPTORY ADRENERGICZNE SERCA
•Receptory β
1
i β
2
– oba sprzężone z CA przez białko G
s
w przedsionkach znajduje się ok.
70
% receptorów β
1
, w
komorach – ok.
80
%
Noradrenalina (NA) ma powinowactwo zarówno do
receptorów β
1
jak β
2
Andrenalina wiąże się z receptorem β
2
Efekt pobudzenia: dodatni efekt ino-, dromo- i
chronotropowy
•Receptory
α
1
–
szlak PLC (DAG i IP
3
)
Efekt pobudzenia: słaby efekt inotropowy dodatni
RECEPTORY CHOLINERCICZNE SERCA
•Receptory M
2
–
sprzężone z CA przez białko G
i
Efekt pobudzenia: ujemny efekt chrono- i
dromotro-powy, w przedsionkach słaby ujemny
efekt inotropowy
RECEPTORY ADRENERGICZNE SERCA
•Receptory β
1
i β
2
– oba sprzężone z CA przez białko G
s
w przedsionkach znajduje się ok.
70
% receptorów β
1
, w
komorach – ok.
80
%
Noradrenalina (NA) ma powinowactwo zarówno do
receptorów β
1
jak β
2
Andrenalina wiąże się z receptorem β
2
Efekt pobudzenia: dodatni efekt ino-, dromo- i
chronotropowy
•Receptory
α
1
–
szlak PLC (DAG i IP
3
)
Efekt pobudzenia: słaby efekt inotropowy dodatni
RECEPTORY CHOLINERCICZNE SERCA
•Receptory M
2
–
sprzężone z CA przez białko G
i
Efekt pobudzenia: ujemny efekt chrono- i
dromotro-powy, w przedsionkach słaby ujemny
efekt inotropowy

UNERWIENIE UKŁADU KRĄŻENIA
UNERWIENIE UKŁADU KRĄŻENIA
RECEPTORY NIEADRENERGICZNE SERCA
•Receptory sprzężone z CA przez białko G
s
- 5-HT
4
efekt inotropowy dodatni w przedsionkach
- H
2
– efekt inotropowy dodatni w przedsionkach i
komo-rach
- VIP – efekt dodatni inotropowy w komorach
•Receptory sprzężone z CA przez białko G
i
-
A
1
(receptor dla adenozyny) - efekt pobudzenia:
ujemny
efekt
chrono-
i
dromotro-powy,
w
przedsionkach słaby ujemny efekt inotropowy
- Receptor dla somatostatyny – efekt inotropwy
ujemny w przedsion-kach
•Receptory sprzężone z PLC
- AT
1
, AT
2
dla angiotenzyny II
- ET
A
i ET
B
dla endoteliny
RECEPTORY NIEADRENERGICZNE SERCA
•Receptory sprzężone z CA przez białko G
s
- 5-HT
4
efekt inotropowy dodatni w przedsionkach
- H
2
– efekt inotropowy dodatni w przedsionkach i
komo-rach
- VIP – efekt dodatni inotropowy w komorach
•Receptory sprzężone z CA przez białko G
i
-
A
1
(receptor dla adenozyny) - efekt pobudzenia:
ujemny
efekt
chrono-
i
dromotro-powy,
w
przedsionkach słaby ujemny efekt inotropowy
- Receptor dla somatostatyny – efekt inotropwy
ujemny w przedsion-kach
•Receptory sprzężone z PLC
- AT
1
, AT
2
dla angiotenzyny II
- ET
A
i ET
B
dla endoteliny

NEUROGENNE NAPIĘCIE NACZYŃ KRWIONOŚNYCH
NEUROGENNE NAPIĘCIE NACZYŃ KRWIONOŚNYCH
•Unerwienie przez zwojowe włókna współczulne
zwężające naczynia dotyczy wszystkich naczyń za
wyjątkiem mikrokrążenia i naczyń łożyska
•Obficie są unerwione zespolenia tętniczo-żylne w
skórze. Bardzo słabo unerwione są naczynia mózgu
i naczynia wieńcowe
•Najlepiej unerwione są tętniczki, słabiej tętnice i
żyłki. Żyły są słabo unerwione
•Neurogenne rozszerzenie naczyń krwionośnych
odbywa się głównie przez zahamowanie tonicznej
aktywności
współczulnej.
Rzadko
naczynia
rozszerzane są w sposób czynny przez współczulne
lub przywspółczulne włókna rozszerzające i
nie
jest to wpływ toniczny
•Unerwienie przez zwojowe włókna współczulne
zwężające naczynia dotyczy wszystkich naczyń za
wyjątkiem mikrokrążenia i naczyń łożyska
•Obficie są unerwione zespolenia tętniczo-żylne w
skórze. Bardzo słabo unerwione są naczynia mózgu
i naczynia wieńcowe
•Najlepiej unerwione są tętniczki, słabiej tętnice i
żyłki. Żyły są słabo unerwione
•Neurogenne rozszerzenie naczyń krwionośnych
odbywa się głównie przez zahamowanie tonicznej
aktywności
współczulnej.
Rzadko
naczynia
rozszerzane są w sposób czynny przez współczulne
lub przywspółczulne włókna rozszerzające i
nie
jest to wpływ toniczny

UNERWIENIE NACZYŃ KRWIONOŚNYCH
UNERWIENIE NACZYŃ KRWIONOŚNYCH
•Współczulne
włókna
naczyniorozszerza-jące
nieadrenerciczne uwalniają:
- ACh
- Histaminę
- Dopaminę
•Przywspółczulne
włókna
naczyniorozsze-rzające uwalniają:
- ACh (naczynia opon mózgowych i mózgu)
-
VIP (naczynia ślinianek)
- ATP za pośrednictwem NO (naczynia
narządów płciowych zewnętrznych)
•Współczulne
włókna
naczyniorozszerza-jące
nieadrenerciczne uwalniają:
- ACh
- Histaminę
- Dopaminę
•Przywspółczulne
włókna
naczyniorozsze-rzające uwalniają:
- ACh (naczynia opon mózgowych i mózgu)
-
VIP (naczynia ślinianek)
- ATP za pośrednictwem NO (naczynia
narządów płciowych zewnętrznych)

UNERWIENIE NACZYŃ KRWIONOŚNYCH
UNERWIENIE NACZYŃ KRWIONOŚNYCH
NARZĄD
TRANSMITER
RECEPTO
R
DZIAŁANIE
MECHANIZM
Tętnice
wieńcowe
NA
β
2
rozszerzanie
↑cAMP
NA
α
2
zwężenie
↓cAMP
NPY
Y
1
zwężenie
↑IP
3
, DAG
Tętnice
trzewne
NA
α
1
zwężenie
↑IP
3
, DAG
Dopamina
D
1
rozszerzenie
↑cAMP
ATP
P
2x
zwężenie
→Ca
2+
P
2y
rozszerzenie
→NO
Tętnice
nerkowe
NA
α
1
zwężenie
↑IP
3
, DAG
NA
β
2
rozszerzenie
↑cAMP
Dopamina
D
1
rozszerzenie
↑cAMP
Tętnice
płciowe
NA
α
1
zwężenie
↑IP
3
, DAG
NA
β
2
rozszerzenie
↑cAMP
Żyły
NA
α
1
zwężenie
↑IP
3
, DAG
NA
β
2
rozszerzenie
↑cAMP
FIZJOLOGIA
SERCA
FIZJOLOGIA
SERCA
Opracowano na podstawie D.E. Mohrman, L.J. Heller. Cardiovascular Physiology. McGraw-Hill,
1997)
Opracowano na podstawie D.E. Mohrman, L.J. Heller. Cardiovascular Physiology. McGraw-Hill,
1997)
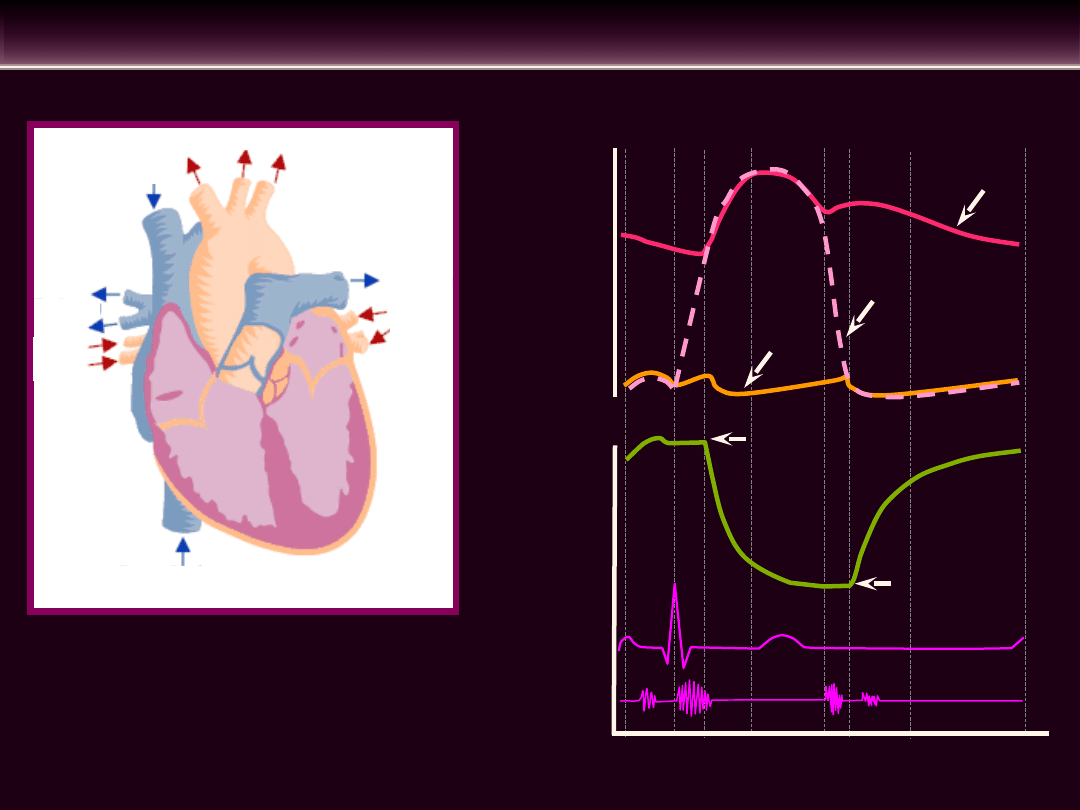
CYKL PRACY SERCA
CYKL PRACY SERCA
do
krążenia
płucnego
z krążenia dużego
z krążenia
płucnego
do krążenia dużego
z krążenia dużego
do
krążenia
płucnego
z krążenia
płucnego
1
1
2
2
3
3
4
4
5
5
6
6
7
7
0
0
0.4
0.4
0.8
0.8
12
0
12
0
80
80
40
40
0
0
12
0
12
0
80
80
40
40
EKG
EKG
Tony
serc
a
Tony
serc
a
O
b
ję
to
ś
ć
le
w
e
j
k
o
m
o
ry
(m
l)
O
b
ję
to
ś
ć
le
w
e
j
k
o
m
o
ry
(m
l)
C
iś
n
ie
n
ie
(
m
m
H
g
)
C
iś
n
ie
n
ie
(
m
m
H
g
)
Czas (sek)
Czas (sek)
Objętość
późnoskurczowa
Objętość
późnoskurczowa
Objętość
późnorozkurczow
a
Objętość
późnorozkurczow
a
Ciśnienie w
aorcie
Ciśnienie w
aorcie
Ciśnienie
w
lewej komorze
Ciśnienie
w
lewej komorze
Ciśnienie w
lewym
przedsionku
Ciśnienie w
lewym
przedsionku
a
a
c
c
v
v
Faza
Faza
faza 1 skurcz przedsionków
faza 2 skurcz komór izowolumetryczny
faza 3 szybki wyrzut
faza 4 zredukowany wyrzut
faza 5 rozkurcz izowolumetryczny
faza 6 szybkie wypełnienie komór
faza 7 zredukowane wypełnianie komór
faza 1 skurcz przedsionków
faza 2 skurcz komór izowolumetryczny
faza 3 szybki wyrzut
faza 4 zredukowany wyrzut
faza 5 rozkurcz izowolumetryczny
faza 6 szybkie wypełnienie komór
faza 7 zredukowane wypełnianie komór
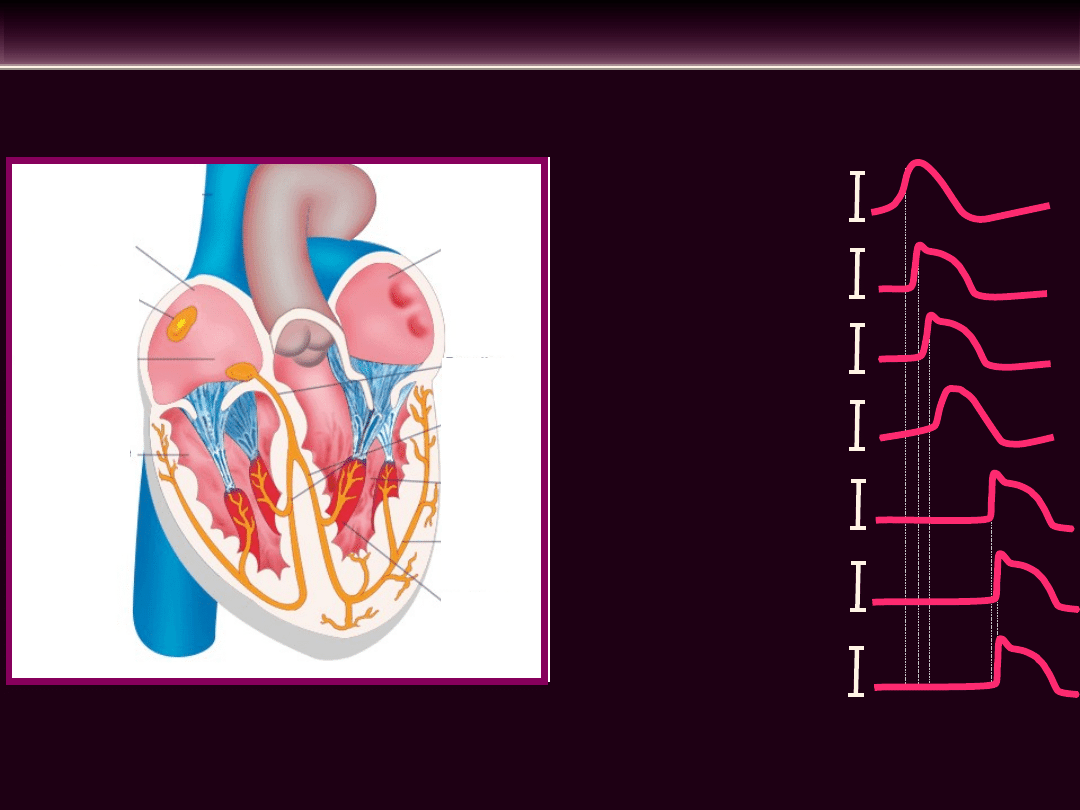
UKŁAD BODŹCOPRZEWODZĄCY SERCA
UKŁAD BODŹCOPRZEWODZĄCY SERCA
Lewy
przedsione
k
Lewa
komora
Mięśnie
brodawkowe
Włókna
Purkinje’go
Węzeł zatokowo-
przedsionkowy
Prawy
przedsione
k
Węzeł
przedsionkowo
-komorowy
Prawa
komora
Odnogi
pęczka
Pęczek
Hisa
-100
-100
0
0
-100
-100
0
0
-100
-100
0
0
-100
-100
0
0
-100
-100
0
0
-100
-100
0
0
-100
-100
0
0
Węzeł zatokowy
Węzeł zatokowy
Mięsień prawego
przedsionka
Mięsień prawego
przedsionka
Mięsień lewego przedsionka
Mięsień lewego przedsionka
Włókna Purkinje’go
Włókna Purkinje’go
Mięsień komory lewej
Mięsień komory lewej
Węzeł przedsionkowo-
komorowy
Węzeł przedsionkowo-
komorowy
Mięsień komory prawej
Mięsień komory prawej
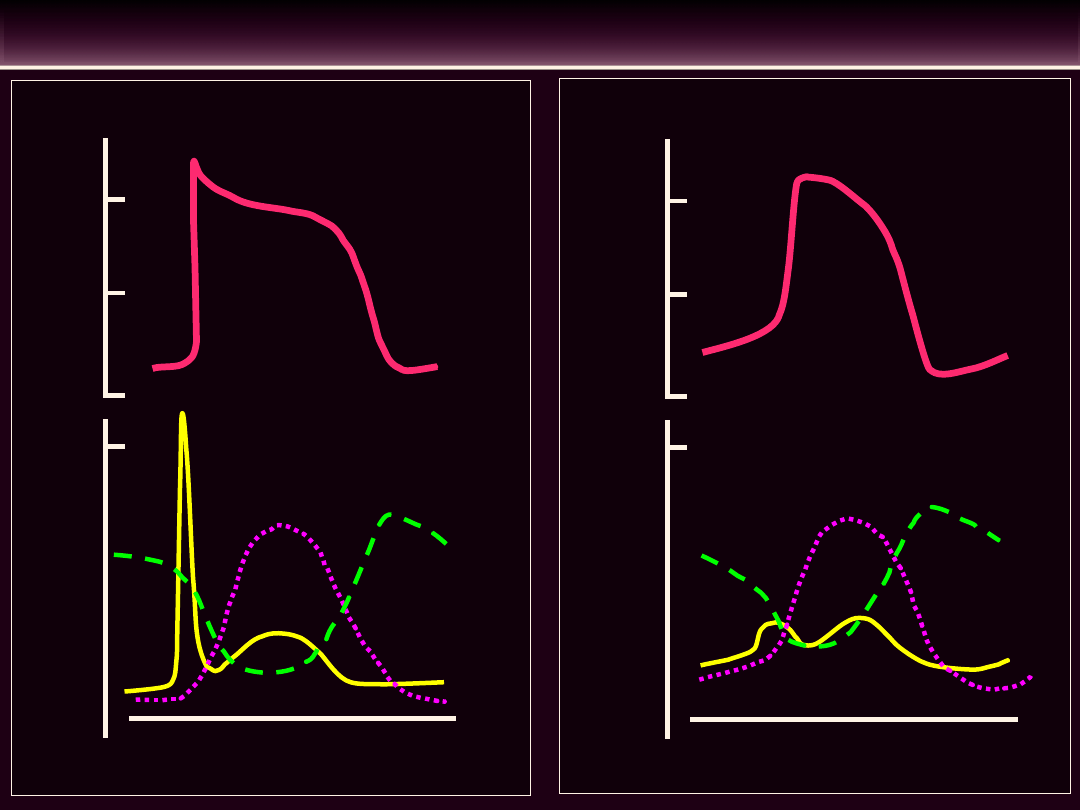
Faza 4
Faza 4
F
a
za
0
F
a
za
0
Faza 1
Faza 1
Faza 2
Faza 2
F
a
za
3
F
a
za
3
Szybko
narastający
potencjał
czynnościowy
Szybko
narastający
potencjał
czynnościowy
K+
K+
Na+
Na+
Ca2+
Ca2+
Czas (sek)
Czas (sek)
0,15
0,15
0
0
0,30
0,30
Wolno narastający potencjał czynnościowy
Wolno narastający potencjał czynnościowy
Faza 4
Faza 4
Na+
Na+
K+
K+
Ca2+
Ca2+
0
0
0,15
0,15
0,30
0,30
-100
-100
0
0
-50
-50
K
o
m
ó
rk
o
w
y
p
o
te
n
c
ja
ł
cz
y
n
n
o
ś
c
io
w
y
(
m
V
)
K
o
m
ó
rk
o
w
y
p
o
te
n
c
ja
ł
cz
y
n
n
o
ś
c
io
w
y
(
m
V
)
W
z
g
lę
d
n
a
p
rz
e
p
u
s
z
c
z
a
ln
o
ś
ć
b
ło
n
y
k
o
m
ó
rk
o
w
e
j
W
z
g
lę
d
n
a
p
rz
e
p
u
s
zc
z
a
ln
o
ś
ć
b
ło
n
y
k
o
m
ó
rk
o
w
e
j
10,0
10,0
1,0
1,0
0,1
0,1
-80
Czas (sek)
Czas (sek)
POTENCJAL CZYNNOŚCIOWY W KOMÓRKACH MIĘŚNIOWYCH
POTENCJAL CZYNNOŚCIOWY W KOMÓRKACH MIĘŚNIOWYCH
-100
-100
0
0
-50
-50
K
o
m
ó
rk
o
w
y
p
o
te
n
c
ja
ł
c
z
y
n
n
o
ś
c
io
w
y
(
m
V
)
K
o
m
ó
rk
o
w
y
p
o
te
n
c
ja
ł
c
z
y
n
n
o
ś
c
io
w
y
(
m
V
)
W
z
g
lę
d
n
a
p
rz
e
p
u
s
z
c
za
ln
o
ś
ć
b
ło
n
y
k
o
m
ó
rk
o
w
e
j
W
z
g
lę
d
n
a
p
rz
e
p
u
s
z
c
z
a
ln
o
ś
ć
b
ło
n
y
k
o
m
ó
rk
o
w
e
j
10,0
10,0
1,0
1,0
0,1
0,1
-80

SKURCZ MIĘŚNIA SZKIELETOWEGO
SKURCZ MIĘŚNIA SZKIELETOWEGO
Podział
skurczów
względem
zmiany
długości mięśnia i generowania siły
Podział
skurczów
względem
zmiany
długości mięśnia i generowania siły
Skurcz izotoniczny
Występuje
wtedy,
gdy
mięsień może się skracać
ale nie generuje napięcia,
bo nie są rozciągane
elementy sprężyste
Skurcz izotoniczny
Występuje
wtedy,
gdy
mięsień może się skracać
ale nie generuje napięcia,
bo nie są rozciągane
elementy sprężyste
Skurcz izometryczny
Występuje
wtedy,
gdy
mięsień nie może się
skracać.
Generuje
napięcie,
ponieważ
sarkomery skra-cają się
kosztem
rozcią-gania
elementów
sprężys-tych
ułożonych szeregowo.
Skurcz izometryczny
Występuje
wtedy,
gdy
mięsień nie może się
skracać.
Generuje
napięcie,
ponieważ
sarkomery skra-cają się
kosztem
rozcią-gania
elementów
sprężys-tych
ułożonych szeregowo.
Czysty skurcz izotoniczny nie występuje nigdy, a czysty
skurcz izometryczny rzadko
Najczęściej występują skurcze dwufazowe - auksotoniczne
Czysty skurcz izotoniczny nie występuje nigdy, a czysty
skurcz izometryczny rzadko
Najczęściej występują skurcze dwufazowe - auksotoniczne
SKURCZ MIĘŚNIA SZKIELETOWEGO
SKURCZ MIĘŚNIA SZKIELETOWEGO
spoczynek skurcz izotoniczny
spoczynek skurcz izotoniczny
Skurcz izotoniczny
Skurcz izotoniczny
spoczynek skurcz izometryczny
spoczynek skurcz izometryczny
Skurcz izometryczny
Skurcz izometryczny
REGULACJA SIŁY
SKURCZ MIĘŚNIA SZKIELETOWEGO I
SERCOWEGO
REGULACJA SIŁY
SKURCZ MIĘŚNIA SZKIELETOWEGO I
SERCOWEGO
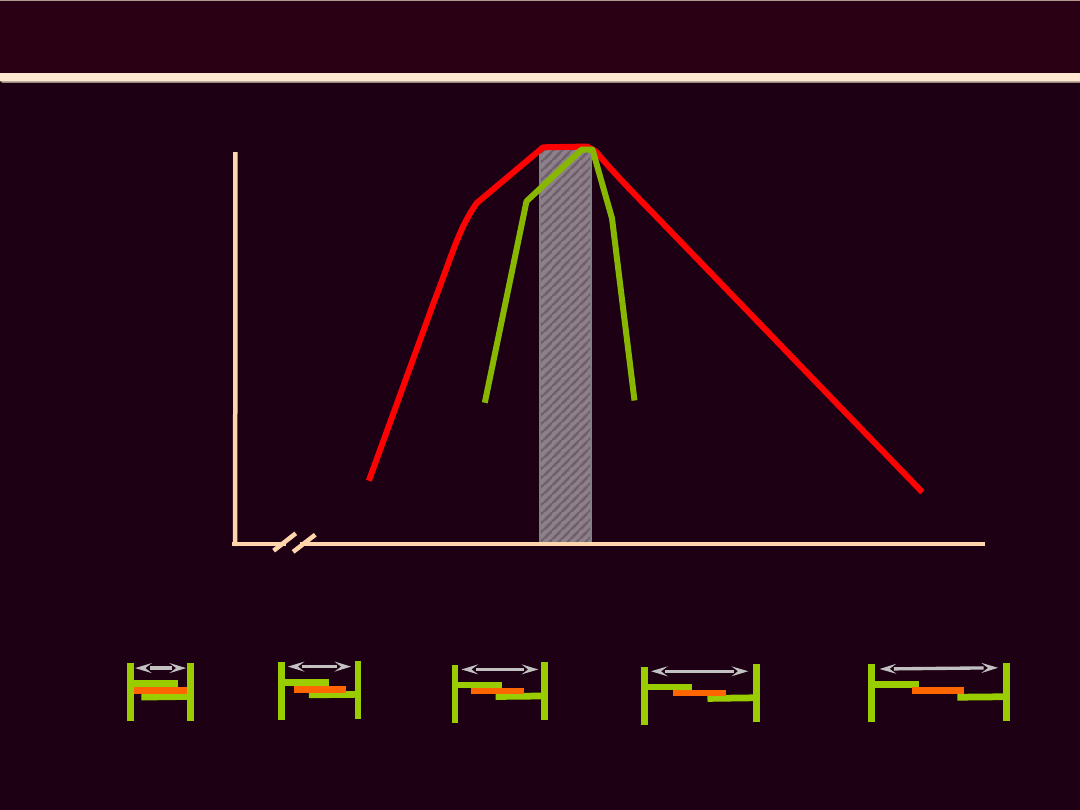
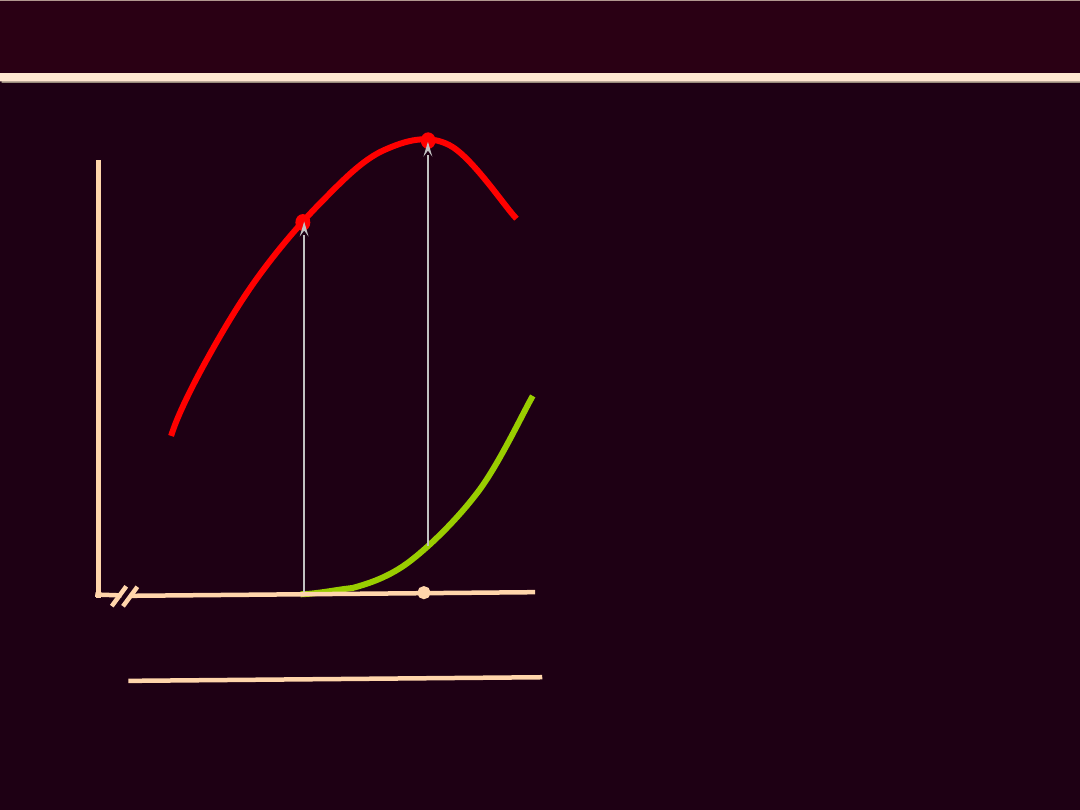
1.2 1.4 1.6 1.8 2.0 2.2 2.4 2.6 2.8 3.0 3.2 3.4 3.6
1.2 1.4 1.6 1.8 2.0 2.2 2.4 2.6 2.8 3.0 3.2 3.4 3.6
100%
100%
40%
40%
0%
0%
długość sarkomeru (μm)
długość sarkomeru (μm)
80%
80%
20%
20%
60%
60%
N
a
p
ię
c
ie
i
zo
m
e
tr
y
c
zn
e
(%
m
a
k
s
im
u
m
)
N
a
p
ię
c
ie
i
zo
m
e
tr
y
c
zn
e
(%
m
a
k
s
im
u
m
)
Z
a
k
re
s
m
a
k
s
y
m
a
ln
e
g
o
n
a
p
ię
c
ia
Z
a
k
re
s
m
a
k
s
y
m
a
ln
e
g
o
n
a
p
ię
c
ia
1.65
1.65
1.90
1.90
2.05
2.05
2.20
2.20
3.65
3.65
Mięsień
szkieletowy
Mięsień
szkieletowy
Mięsień
sercowy
Mięsień
sercowy
REGULACJA SIŁY
SKURCZ MIĘŚNIA SZKIELETOWEGO I
SERCOWEGO
REGULACJA SIŁY
SKURCZ MIĘŚNIA SZKIELETOWEGO I
SERCOWEGO
80 90 100
80 90 100
100%
100%
40%
40%
0%
0%
względna długość mięśnia
(100% = długość przy sile maksymalnej)
względna długość mięśnia
(100% = długość przy sile maksymalnej)
80%
80%
20%
20%
60%
60%
S
ił
a
m
ię
ś
n
ia
(%
o
d
m
a
k
s
im
u
m
w
s
k
u
rc
z
u
i
z
o
m
e
tr
y
c
z
n
y
m
)
S
ił
a
m
ię
ś
n
ia
(%
o
d
m
a
k
s
im
u
m
w
s
k
u
rc
z
u
i
z
o
m
e
tr
y
c
z
n
y
m
)
napięcie całkowite
napięcie całkowite
napięcie bierne
napięcie bierne
1.8 2.0 2.2
1.8 2.0 2.2
długość sarkomeru (μm)
długość sarkomeru (μm)
n
a
p
ię
c
ie
c
zy
n
n
e
n
a
p
ię
c
ie
c
zy
n
n
e
Wypadkowa napięcia
całkowitego jest sumą
napięcia
biernego
i
czynnego
Sprężystość mięśnia
zale-ży
od
elementów
sprężys-tych położonych
szeregowo
(elementy
tkanki łącznej, odcinki
szyjkowe
miozyny)
i
równolegle (błona włókna
mięśniowego,
tkanka
łącz-na) oraz elementów
kurczli-wych. Rozciąganie
tych
ele-mentów
jest
przyczyną
na-pięcia
biernego
Napięcie czynne jest
określone
przez
ilość
most-ków
aktynowo-
miozyno-wych i dlatego
zmienia
się
wraz
z
długością mięśnia
Wypadkowa napięcia
całkowitego jest sumą
napięcia
biernego
i
czynnego
Sprężystość mięśnia
zale-ży
od
elementów
sprężys-tych położonych
szeregowo
(elementy
tkanki łącznej, odcinki
szyjkowe
miozyny)
i
równolegle (błona włókna
mięśniowego,
tkanka
łącz-na) oraz elementów
kurczli-wych. Rozciąganie
tych
ele-mentów
jest
przyczyną
na-pięcia
biernego
Napięcie czynne jest
określone
przez
ilość
most-ków
aktynowo-
miozyno-wych i dlatego
zmienia
się
wraz
z
długością mięśnia
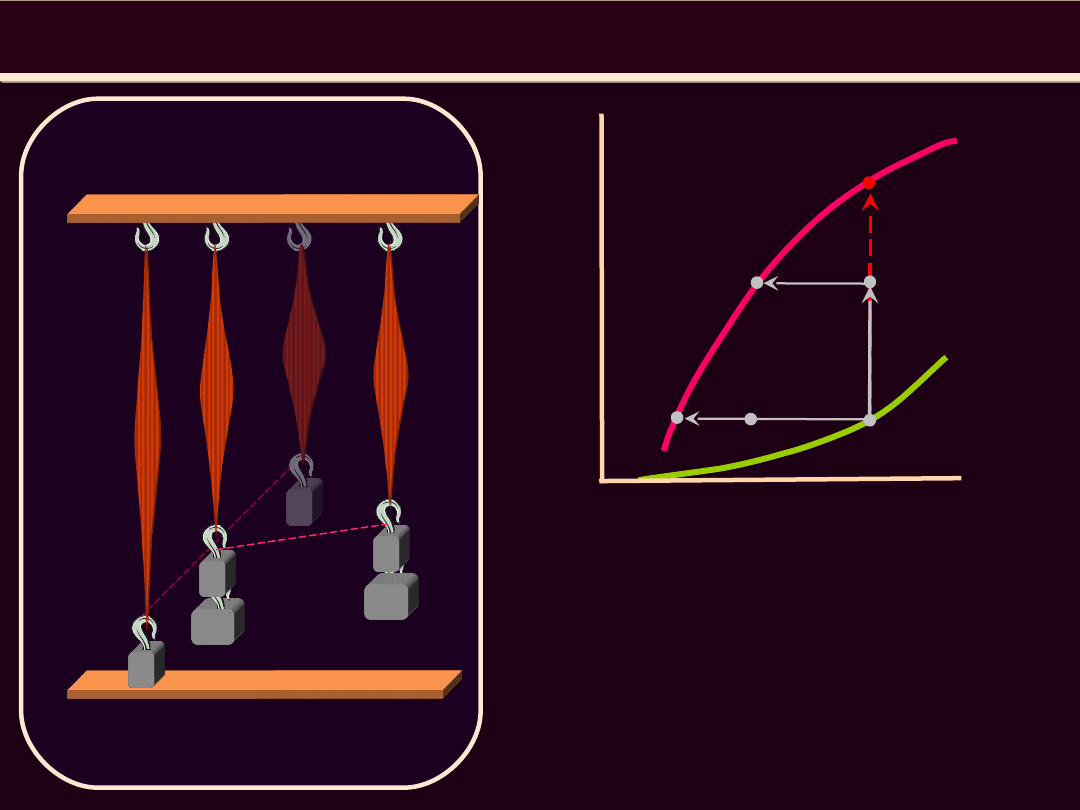
1
g
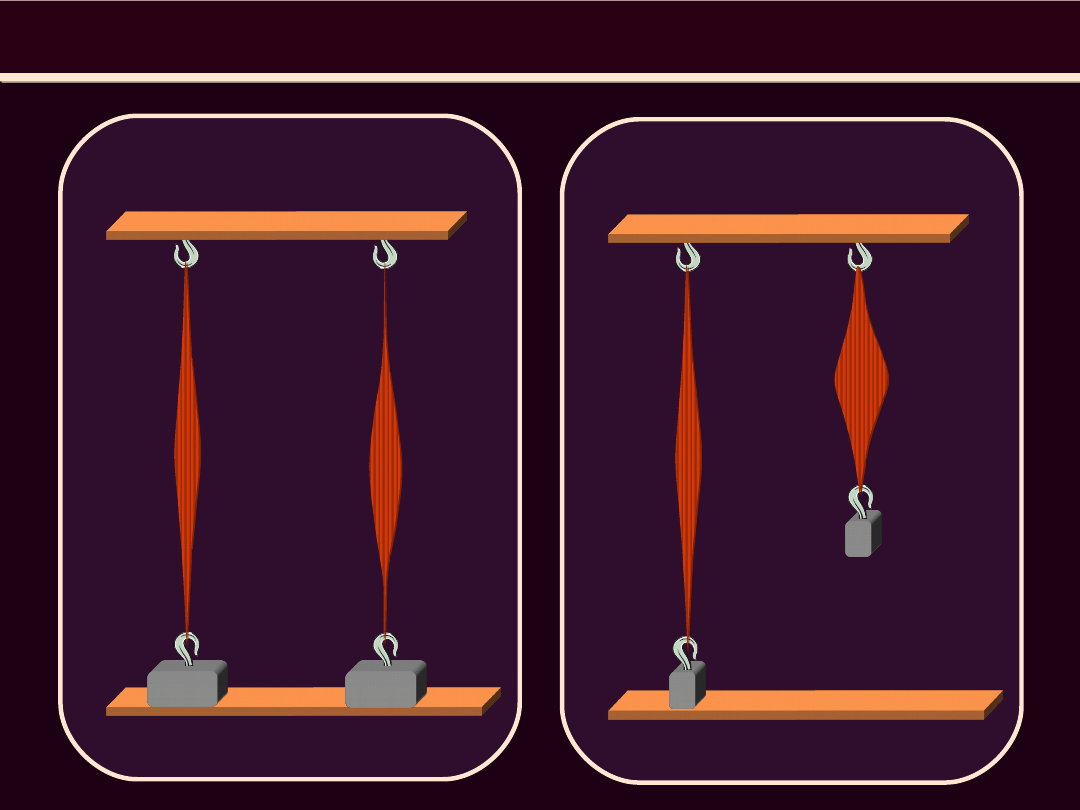
SKURCZ IZOTONICZNY MIĘŚNIA WTÓRNIE
OBCIĄŻONEGO
SKURCZ IZOTONICZNY MIĘŚNIA WTÓRNIE
OBCIĄŻONEGO
spoczynek skurcz izotoniczny mięśnia
wtórnie obciążonego
spoczynek skurcz izotoniczny mięśnia
wtórnie obciążonego
Skurcz izotoniczny
Skurcz izotoniczny
1
g
obciążeni
e wstępne
obciążeni
e wstępne
1
g
2
g
2
g
1
g
obciążeni
e
następcze
obciążeni
e
następcze
1
1
3
3
2
2
4
4
5
5
napięcie bierne
napięcie bierne
5
5
2
2
0
0
względna długość mięśnia
względna długość mięśnia
4
4
1
1
3
3
N
a
p
ię
c
ie
m
ię
ś
n
io
w
e
(
g
)
N
a
p
ię
c
ie
m
ię
ś
n
io
w
e
(
g
)
maksymalne
napięcie
izometryczne
maksymalne
napięcie
izometryczne
1
1
2
2
3
3
4
4
5
5
•
Tor
1,2,3
– skurcz izotoniczny w warunkach
obciążenia wstępnego, mniejszego niż potrzebne
do
wywołania
maksymalnego
napięcia
izometrycznego dla danej długości mięśnia
•
Tor
1,4,5
– na samym początku skurczu
izotonicznego mięsień został wtórnie obciążony.
Tor
1-4
– skurcz izometryczny, podczas którego
mięsień
rozwija
napięcie
niezbędne
do
zrównoważenia obciążenia następczego. Tor 4-5 –
skurcz izotoniczny, podczas którego mięsień się
skraca
•
Tor
1,2,3
– skurcz izotoniczny w warunkach
obciążenia wstępnego, mniejszego niż potrzebne
do
wywołania
maksymalnego
napięcia
izometrycznego dla danej długości mięśnia
•
Tor
1,4,5
– na samym początku skurczu
izotonicznego mięsień został wtórnie obciążony.
Tor
1-4
– skurcz izometryczny, podczas którego
mięsień
rozwija
napięcie
niezbędne
do
zrównoważenia obciążenia następczego. Tor 4-5 –
skurcz izotoniczny, podczas którego mięsień się
skraca
REGULACJA SIŁY SKURCZU MIĘŚNIA SERCOWEGO
REGULACJA SIŁY SKURCZU MIĘŚNIA SERCOWEGO
• ZALEŻNOŚĆ „SIŁA BODŹCA – SIŁA SKURCZU”
Siła skurczu mięśnia sercowego nie zależy od siły bodźca. W danych warunkach
pracy mięśnia sercowego każdy bodziec ponadprogowy wywoła skurcz o sile
maksymalnej – prawo „wszystko albo nic”
• ZALEŻNOŚĆ „CZĘSTOŚĆ BODŹCA – SIŁA SKURCZU”
W mięśniu sercowym ze względu na długi okres refrakcji roboczych komórek
mięśniowych nie występuje skurcz tężcowy. Obserwuje się wzrost siły skurczu
mięśnia sercowego w miarę wzrostu częstości skurczów serca (efekt Bowditcha).
Przyczyną tego efektu jest prawdopodobnie wzrost wewnątrzkomórkowego
stężenia jonów Ca
2+
• ZALEŻNOŚĆ „DŁUGOŚĆ (OBCIĄŻENIE WSTĘPNE) – SIŁA SKURCZU”
Siła skurczu mięśnia sercowego jest tym większa im większy jest poprzedzający ją
stopień rozciągnięcia włókien mięśniowych. Zależność siły skurczu mięśnia
sercowego od stopnia jego rozciągnięcia leży u podstaw prawa Franka-Starlinga,
które mówi, że:
Objętość wyrzutowa serca wzrasta w miarę wzrostu stopnia wypełnienia komór
(wzrost obciążenia wstępnego)
• ZALEŻNOŚĆ „SZYBKOŚĆ – SIŁA (OBCIĄŻENIE NATĘPCZE)”
Im większe jest obciążenie następcze mięśnia, tym mniejsza jest szybkość skurczu
mięśnia sercowego. Zależność szybkości skurczu od obciążenia opisuje prawo
Hilla:
• ZALEŻNOŚĆ „SIŁA BODŹCA – SIŁA SKURCZU”
Siła skurczu mięśnia sercowego nie zależy od siły bodźca. W danych warunkach
pracy mięśnia sercowego każdy bodziec ponadprogowy wywoła skurcz o sile
maksymalnej – prawo „wszystko albo nic”
• ZALEŻNOŚĆ „CZĘSTOŚĆ BODŹCA – SIŁA SKURCZU”
W mięśniu sercowym ze względu na długi okres refrakcji roboczych komórek
mięśniowych nie występuje skurcz tężcowy. Obserwuje się wzrost siły skurczu
mięśnia sercowego w miarę wzrostu częstości skurczów serca (efekt Bowditcha).
Przyczyną tego efektu jest prawdopodobnie wzrost wewnątrzkomórkowego
stężenia jonów Ca
2+
• ZALEŻNOŚĆ „DŁUGOŚĆ (OBCIĄŻENIE WSTĘPNE) – SIŁA SKURCZU”
Siła skurczu mięśnia sercowego jest tym większa im większy jest poprzedzający ją
stopień rozciągnięcia włókien mięśniowych. Zależność siły skurczu mięśnia
sercowego od stopnia jego rozciągnięcia leży u podstaw prawa Franka-Starlinga,
które mówi, że:
Objętość wyrzutowa serca wzrasta w miarę wzrostu stopnia wypełnienia komór
(wzrost obciążenia wstępnego)
• ZALEŻNOŚĆ „SZYBKOŚĆ – SIŁA (OBCIĄŻENIE NATĘPCZE)”
Im większe jest obciążenie następcze mięśnia, tym mniejsza jest szybkość skurczu
mięśnia sercowego. Zależność szybkości skurczu od obciążenia opisuje prawo
Hilla:
V = b •
V = b •
P
0
- P
P
0
- P
P+a
P+a
V – szybkość skurczu
a, b – stałe współczynniki ciepła i szybkości przejścia energii chemicznej w energię
mechaniczną
P
0
– maksymalnie możliwa siła skurczu
P – siła skurczu mięśnia uwarunkowana obciążeniem
V – szybkość skurczu
a, b – stałe współczynniki ciepła i szybkości przejścia energii chemicznej w energię
mechaniczną
P
0
– maksymalnie możliwa siła skurczu
P – siła skurczu mięśnia uwarunkowana obciążeniem

KURCZLIWOŚĆ MIĘŚNIA SERCOWEGO
KURCZLIWOŚĆ MIĘŚNIA SERCOWEGO
•Kurczliwość
mięśnia jest jego cechą wewnętrzną.
Kurczliwość mięśnia zależy od ilości elementów
kurczliwych we włóknach mięśniowych, stężenia
wewnątrzkomórkowego
jonów
Ca
2+
,
wrażliwości
elementów kurczliwych na jony Ca
2+
. O zmianie
kurczliwości świadczy taka zmiana siły i szybkości
skurczu, która nie jest związana ze zmianą wyjściowej
długości mięśnia
•Każdy bodziec zwiększający maksymalne napięcie
izometryczne mięśnia bez zmiany jego długości
zwiększa jego kurczliwość, czyli wywiera działanie
inotropowe dodatnie
•Odzwierciedleniem
stanu
kurczliwości
mięśnia
sercowego jest
frakcja wyrzutowa
(stosunek
objętości
póznorozkurczowej
serca
do
objętości
wyrzutowej)
•Kurczliwość
mięśnia jest jego cechą wewnętrzną.
Kurczliwość mięśnia zależy od ilości elementów
kurczliwych we włóknach mięśniowych, stężenia
wewnątrzkomórkowego
jonów
Ca
2+
,
wrażliwości
elementów kurczliwych na jony Ca
2+
. O zmianie
kurczliwości świadczy taka zmiana siły i szybkości
skurczu, która nie jest związana ze zmianą wyjściowej
długości mięśnia
•Każdy bodziec zwiększający maksymalne napięcie
izometryczne mięśnia bez zmiany jego długości
zwiększa jego kurczliwość, czyli wywiera działanie
inotropowe dodatnie
•Odzwierciedleniem
stanu
kurczliwości
mięśnia
sercowego jest
frakcja wyrzutowa
(stosunek
objętości
póznorozkurczowej
serca
do
objętości
wyrzutowej)
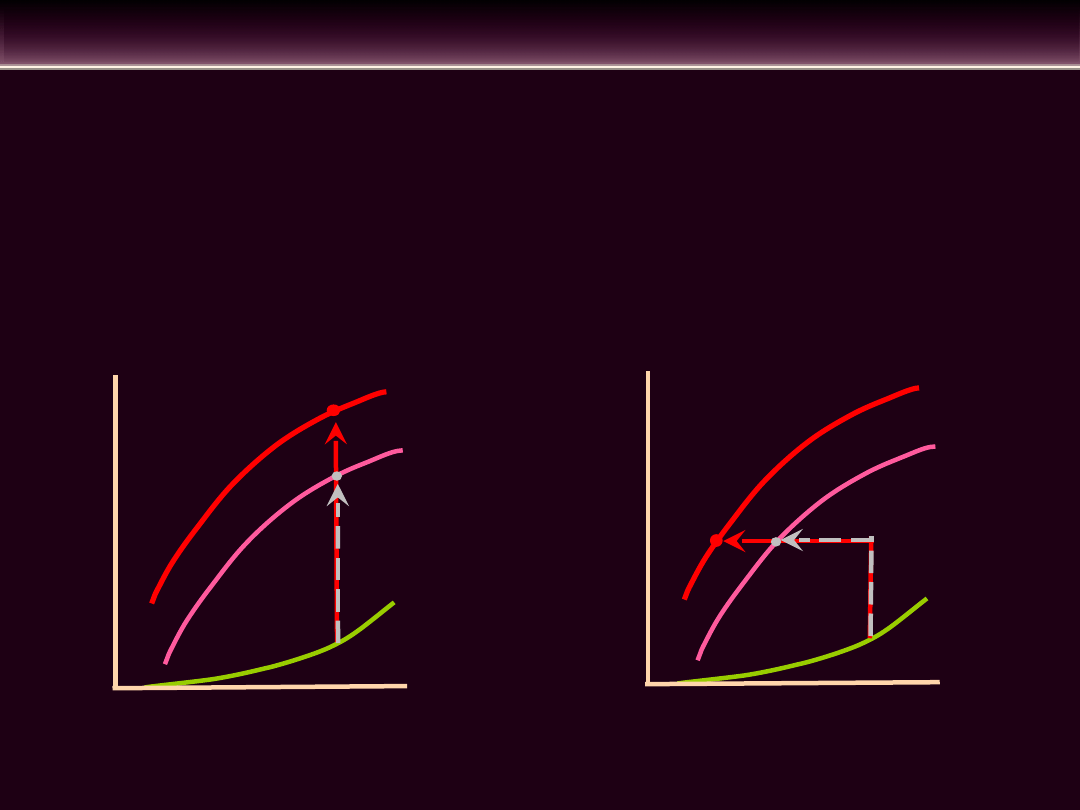
KURCZLIWOŚĆ MIĘŚNIA SERCOWEGO
KURCZLIWOŚĆ MIĘŚNIA SERCOWEGO
• Najważniejszym fizjologicznym regulatorem kurczliwości
mięśnia
sercowego
jest
noradrenalina (NA)
uwalniana z zakończeń nerwowych włókien współczulnych
• Najważniejszym fizjologicznym regulatorem kurczliwości
mięśnia
sercowego
jest
noradrenalina (NA)
uwalniana z zakończeń nerwowych włókien współczulnych
5
5
2
2
0
0
długość mięśnia
długość mięśnia
4
4
1
1
3
3
N
a
p
ię
c
ie
m
ię
ś
n
io
w
e
(
g
)
N
a
p
ię
c
ie
m
ię
ś
n
io
w
e
(
g
)
napięcie bierne
napięcie bierne
maksymalne
napięcie
izometryczne
maksymalne
napięcie
izometryczne
6
6
Skurcz izometryczny
Skurcz izometryczny
z
NA
be
z
NA
be
z
NA
z NA
lub b
ez NA
z NA
lub b
ez NA
5
5
2
2
0
0
długość mięśnia
długość mięśnia
4
4
1
1
3
3
N
a
p
ię
c
ie
m
ię
ś
n
io
w
e
(
g
)
N
a
p
ię
c
ie
m
ię
ś
n
io
w
e
(
g
)
napięcie bierne
napięcie bierne
maksymalne
napięcie
izometryczne
maksymalne
napięcie
izometryczne
6
6
Skurcz mięśnia wtórnie obciążonego
Skurcz mięśnia wtórnie obciążonego
z
NA
z
NA
be
z
NA
be
z
NA
z NA
lub b
ez NA
z NA
lub b
ez NA
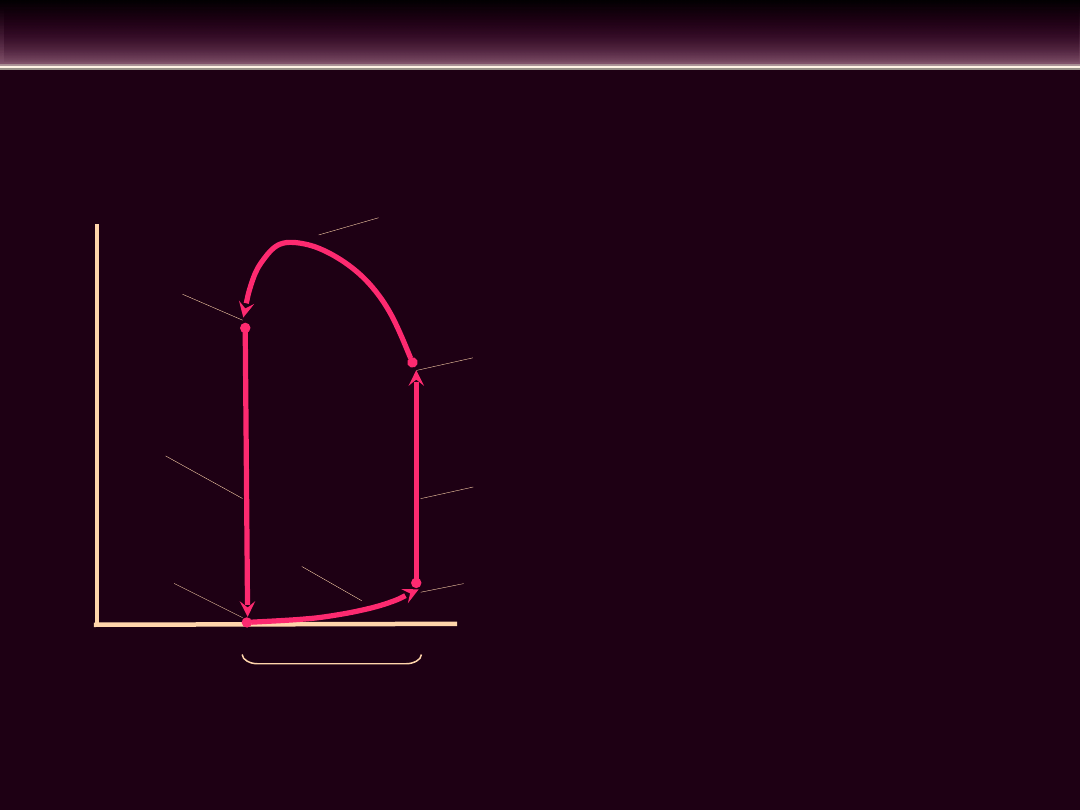
REGULACJA OBJĘTOŚCI WYRZUTOWEJ
REGULACJA OBJĘTOŚCI WYRZUTOWEJ
objętość
późnorozkurczowa
objętość
późnorozkurczowa
skurcz
izowolumetryczny
skurcz
izowolumetryczny
otwarcie zastawki
aortalnej
otwarcie zastawki
aortalnej
80
80
Objętość lewej komory (ml)
Objętość lewej komory (ml)
60
60
120
120
C
iś
n
ie
n
ie
w
l
e
w
e
j
k
o
m
o
rz
e
(
m
m
H
g
)
C
iś
n
ie
n
ie
w
l
e
w
e
j
k
o
m
o
rz
e
(
m
m
H
g
)
120
120
CYKL LEWEJ KOMORY
CYKL LEWEJ KOMORY
objętość wyrzutowa (SV)
objętość wyrzutowa (SV)
wyrzut
wyrzut
objętość
późnoskurczowa
objętość
późnoskurczowa
rozkurcz
izowolumetryczny
rozkurcz
izowolumetryczny
otwarcie zastawki
dwudzielnej
otwarcie zastawki
dwudzielnej
Wypełnienie komory
Wypełnienie komory
• Ciśnienie późnorozkurczo-
we w komorze określa sto-
pień rozciągnięcia włókien
mięśniowych w spoczynku
podczas rozkurczu i jest
nazywane
obciążeniem
wstępnym
• Średnie ciśnienie tętnicze
(MAP)
warunkuje
napięcie,
które
musi
rozwinąć mięsień sercowy
podczas
skurczu,
by
wyrzucić pewną objętość
krwi
do
aorty
jest
nazywane
obciążeniem
następ-czym
• Ciśnienie późnorozkurczo-
we w komorze określa sto-
pień rozciągnięcia włókien
mięśniowych w spoczynku
podczas rozkurczu i jest
nazywane
obciążeniem
wstępnym
• Średnie ciśnienie tętnicze
(MAP)
warunkuje
napięcie,
które
musi
rozwinąć mięsień sercowy
podczas
skurczu,
by
wyrzucić pewną objętość
krwi
do
aorty
jest
nazywane
obciążeniem
następ-czym

WPŁYW ZMIAN OBCIĄŻENIA WSTĘPNEGO NA
OBJĘTOŚĆ WYRZUTOWĄ
WPŁYW ZMIAN OBCIĄŻENIA WSTĘPNEGO NA
OBJĘTOŚĆ WYRZUTOWĄ
•PRAWO FRANKA STARLINGA: Objętość wyrzutowa
serca wzrasta w miarę wzrostu stopnia wypełnienia
komór
•Wzrost
obciążenia
wstępnego
prowadzi
do
zwiększenia objętości poźnorozkurczowej serca, a
zatem do wzrostu spoczynkowej długości włókien
mięśniowych
•Regulacja objętości wyrzutowej (SV) w zależności
od
obciążenia
wstępnego
jest
nazywana
autoregulacją heterometryczną
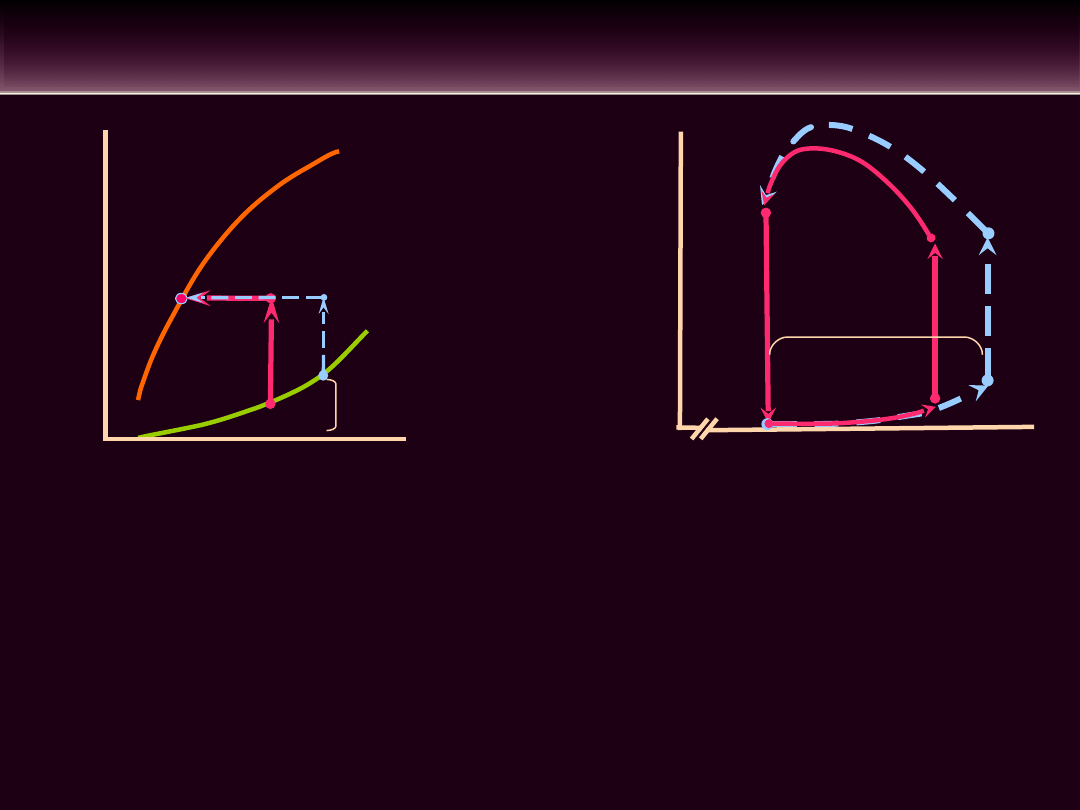
WPŁYW ZMIAN OBCIĄŻENIA WSTĘPNEGO NA
OBJĘTOŚĆ WYRZUTOWĄ
WPŁYW ZMIAN OBCIĄŻENIA WSTĘPNEGO NA
OBJĘTOŚĆ WYRZUTOWĄ
• Wzrost obciążenia wstępnego nie prowadzi do zmiany
objętości późnoskurczowej mięśnia sercowego. W wyniku
regulacji heterometrycznej objętość wyrzutowa wzrasta o
taką samą wartość jak objętość późnorozkurczowa
bez
jakichkolwiek zmian frakcji wyrzutowej
• Wzrost obciążenia wstępnego nie prowadzi do zmiany
objętości późnoskurczowej mięśnia sercowego. W wyniku
regulacji heterometrycznej objętość wyrzutowa wzrasta o
taką samą wartość jak objętość późnorozkurczowa
bez
jakichkolwiek zmian frakcji wyrzutowej
5
5
2
2
długość mięśnia
długość mięśnia
4
4
1
1
3
3
N
a
p
ię
c
ie
m
ię
śn
io
w
e
(
g
)
N
a
p
ię
c
ie
m
ię
śn
io
w
e
(
g
)
napięcie bierne
napięcie bierne
maksymalne napięcie
izometryczne
maksymalne napięcie
izometryczne
80
80
objętość lewej komory
(ml)
objętość lewej komory
(ml)
60
60
120
120
C
iś
n
ie
n
ie
w
l
e
w
e
j
k
o
m
o
rz
e
(m
m
H
g
)
C
iś
n
ie
n
ie
w
l
e
w
e
j
k
o
m
o
rz
e
(m
m
H
g
)
120
120
wzrost objętości
wyrzutowej
wzrost objętości
wyrzutowej
Wzrost siły
skurczu mięśnia
Wzrost siły
skurczu mięśnia
wzrost obciążenia
wstępnego
wzrost obciążenia
wstępnego
40
40
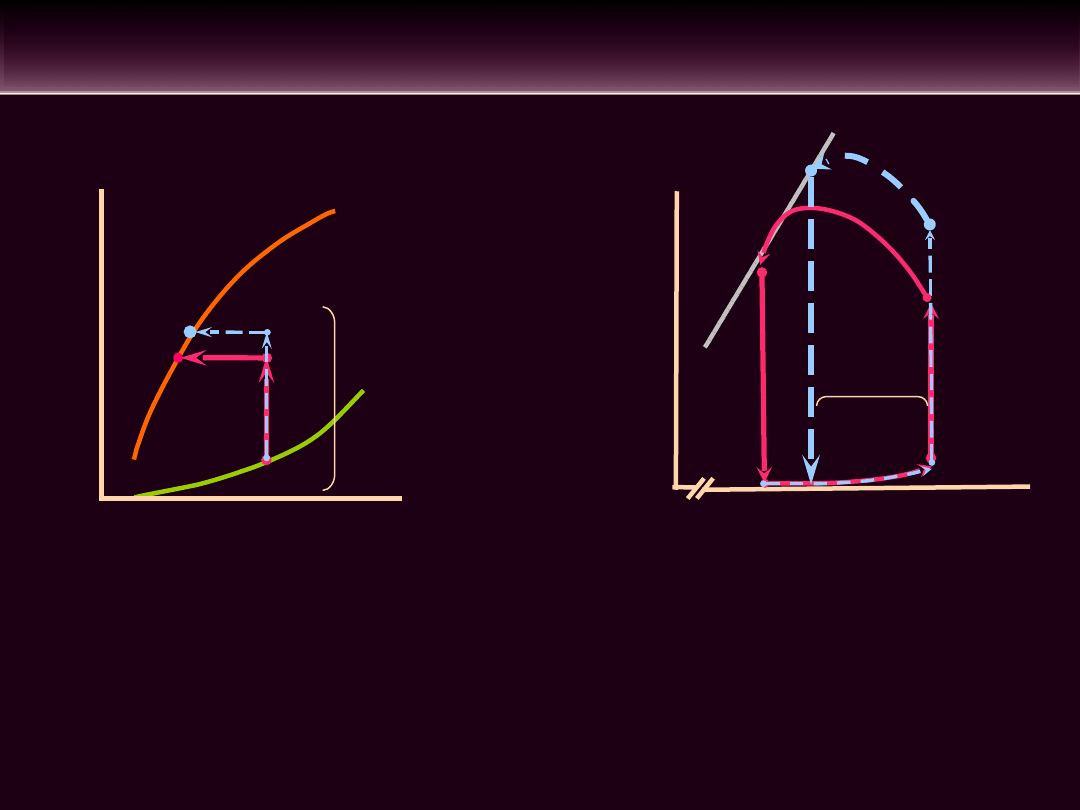
WPŁYW ZMIAN OBCIĄŻENIA NASTĘPCZEGO NA
OBJĘTOŚĆ WYRZUTOWĄ
WPŁYW ZMIAN OBCIĄŻENIA NASTĘPCZEGO NA
OBJĘTOŚĆ WYRZUTOWĄ
5
5
2
2
długość mięśnia
długość mięśnia
4
4
1
1
3
3
N
a
p
ię
c
ie
m
ię
śn
io
w
e
(
g
)
N
a
p
ię
c
ie
m
ię
śn
io
w
e
(
g
)
napięcie bierne
napięcie bierne
maksymalne napięcie
izometryczne
maksymalne napięcie
izometryczne
zmniejszenie siły
skurczu mięśnia
zmniejszenie siły
skurczu mięśnia
wzrost obciążenia
następczego
wzrost obciążenia
następczego
80
80
objętość lewej komory
(ml)
objętość lewej komory
(ml)
60
60
120
120
C
iś
n
ie
n
ie
w
l
e
w
e
j
k
o
m
o
rz
e
(m
m
H
g
)
C
iś
n
ie
n
ie
w
l
e
w
e
j
k
o
m
o
rz
e
(m
m
H
g
)
120
120
spadek objętości
wyrzutowej
spadek objętości
wyrzutowej
40
40
• Wzrost obciążenia następczego prowadzi do wydłużenia
czasu
trwania
skurczu
izowolumetrycznego
mięśnia
sercowego. Zmniejsza się czas trwania okresu wyrzutu.
Maleje objętość wyrzutowa, wzrasta natomiast objętość
późnoskurczowa
• Wzrost obciążenia następczego prowadzi do wydłużenia
czasu
trwania
skurczu
izowolumetrycznego
mięśnia
sercowego. Zmniejsza się czas trwania okresu wyrzutu.
Maleje objętość wyrzutowa, wzrasta natomiast objętość
późnoskurczowa
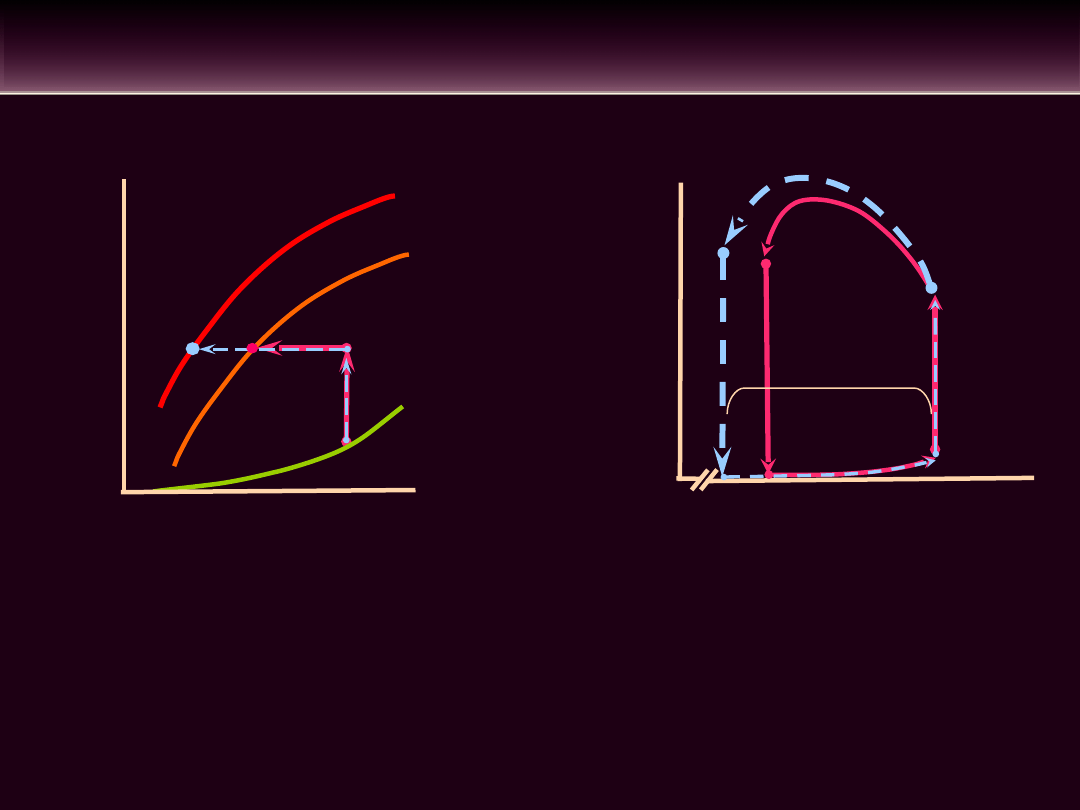
WPŁYW ZMIAN KURCZLIWOŚCI MIĘŚNIA
SERCOWEGO NA OBJĘTOŚĆ WYRZUTOWĄ
WPŁYW ZMIAN KURCZLIWOŚCI MIĘŚNIA
SERCOWEGO NA OBJĘTOŚĆ WYRZUTOWĄ
5
5
2
2
0
0
długość mięśnia
długość mięśnia
4
4
1
1
3
3
N
a
p
ię
c
ie
m
ię
śn
io
w
e
(
g
)
N
a
p
ię
c
ie
m
ię
śn
io
w
e
(
g
)
napięcie bierne
napięcie bierne
maksymalne
napięcie
izometryczne
maksymalne
napięcie
izometryczne
6
6
z N
A
z N
A
be
z N
A
be
z N
A
z NA
lub b
ez NA
z NA
lub b
ez NA
80
80
objętość lewej komory
(ml)
objętość lewej komory
(ml)
60
60
120
120
C
iś
n
ie
n
ie
w
l
e
w
e
j
k
o
m
o
rz
e
(m
m
H
g
)
C
iś
n
ie
n
ie
w
l
e
w
e
j
k
o
m
o
rz
e
(m
m
H
g
)
120
120
wzrost objętości
wyrzutowej
wzrost objętości
wyrzutowej
40
40
wzrost siły skurczu
wzrost siły skurczu
• Określenie kurczliwości mięśnia sercowego w sposób bezpośredni
nie jest możliwe. Wskaźnikiem kurczliwości mięśnia sercowego jest
maksymalna szybkość przyrostu ciśnienia w lewej komorze podczas
skurczu izowolumetrycznego dP/dt
max
mierzona po umiejscowieniu
cewnika w komorze. Metodą nieinwazyjną oceniającą między innymi
kurczliwość mięśnia sercowego jest USG serca
• Parametrem odzwierciedlającym kurczliwość mięśnia sercowego jest
frakcja wyrzutowa, której prawidłowa wartość w spoczynku wynosi
średnio ok. 65%
• Określenie kurczliwości mięśnia sercowego w sposób bezpośredni
nie jest możliwe. Wskaźnikiem kurczliwości mięśnia sercowego jest
maksymalna szybkość przyrostu ciśnienia w lewej komorze podczas
skurczu izowolumetrycznego dP/dt
max
mierzona po umiejscowieniu
cewnika w komorze. Metodą nieinwazyjną oceniającą między innymi
kurczliwość mięśnia sercowego jest USG serca
• Parametrem odzwierciedlającym kurczliwość mięśnia sercowego jest
frakcja wyrzutowa, której prawidłowa wartość w spoczynku wynosi
średnio ok. 65%

METABOLIZM MIĘŚNIA SERCOWEGO
METABOLIZM MIĘŚNIA SERCOWEGO
• Przemiana metaboliczna mięśnia sercowego ma charakter głównie
aerobowy
• Podstawowa przemiana materii mięśnia sercowego (bez uwzględnienia
procesów elektromechanicznych) stanowi ok. 25% zużycia tlenu w
stanie spoczynku
• Około 75% wydatku energetycznego jest związane ze skurczem włókien
mięśniowych
• Podczas skurczu izowolumetrycznego mięsień sercowy zużywa ok. 50%
całego tlenu, nie wykonując przy tym pracy użytecznej
• Obciążenie następcze jest głównym czynnikiem warunkującym wzrost
zużycia tlenu przez mięsień sercowy
• Podczas skurczu izotonicznego mięsień sercowy wykonuje pracę
zewnętrzną, użyteczną, wymagającą zużycia tlenu. Wzrost kurczliwości
mięśnia sercowego prowadzi do wzrostu pracy zewnętrznej i wzrostu
zużycia tlenu
• Ważnym czynnikiem minutowego zużycia tlenu przez mięsień sercowy
jest częstość skurczów serca
• Najskuteczniejszym ze względu na wydatek energetyczny sposobem
osiągnięcia określonej pojemności minutowej (CO) jest znaczny wzrost
objętości wyrzutowej (SV) przy niewielkim wzroście częstości skurczów
serca (HR)
• Przemiana metaboliczna mięśnia sercowego ma charakter głównie
aerobowy
• Podstawowa przemiana materii mięśnia sercowego (bez uwzględnienia
procesów elektromechanicznych) stanowi ok. 25% zużycia tlenu w
stanie spoczynku
• Około 75% wydatku energetycznego jest związane ze skurczem włókien
mięśniowych
• Podczas skurczu izowolumetrycznego mięsień sercowy zużywa ok. 50%
całego tlenu, nie wykonując przy tym pracy użytecznej
• Obciążenie następcze jest głównym czynnikiem warunkującym wzrost
zużycia tlenu przez mięsień sercowy
• Podczas skurczu izotonicznego mięsień sercowy wykonuje pracę
zewnętrzną, użyteczną, wymagającą zużycia tlenu. Wzrost kurczliwości
mięśnia sercowego prowadzi do wzrostu pracy zewnętrznej i wzrostu
zużycia tlenu
• Ważnym czynnikiem minutowego zużycia tlenu przez mięsień sercowy
jest częstość skurczów serca
• Najskuteczniejszym ze względu na wydatek energetyczny sposobem
osiągnięcia określonej pojemności minutowej (CO) jest znaczny wzrost
objętości wyrzutowej (SV) przy niewielkim wzroście częstości skurczów
serca (HR)

RÓŻNICE W CZYNNOŚCI SERCA W ZALEŻNOŚCI OD
WIEKU
RÓŻNICE W CZYNNOŚCI SERCA W ZALEŻNOŚCI OD
WIEKU
• Grubość mięśnia lewej i prawej komory serca u noworodka
jest podobna. Przewaga grubości lewej komory uwidocznia
się od 20-30 dna od momentu urodzenia
• Różnicowanie i dojrzewanie komórek roboczych i układu
bodźcoprzewodzącego serca trwa do 2 roku życia
• Mięsień sercowy noworodków i niemowląt jest mało podatny.
Podatność mięśnia sercowego zwiększa się po pierwszym
roku życia. W związku z tym po pierwszym roku życia wzrasta
rola regulacji heterometrycznej czynności serca (prawo
Franka-Starlinga)
• Wrażliwość receptorów adrenergicznyh serca noworodków
jest wyraźnie mniejsza niż u osób dorosłych. W miarę wzrostu
noworodka wrażliwość receptorów wzrasta, warunkując
stopniowe nasilenie wpływu układu współczulnego na
mięsień sercowy
• Wpływ toniczny nerwu błędnego na serce u noworodków jest
minimalny. W miarę wzrostu noworodka wpływ układu
przywspółczulnego na serce wzrasta. W skutek tego częstość
skurczów serca maleje z 140 sk/min u noworodka do 70
sk/min w wieku 14-16 lat
• Grubość mięśnia lewej i prawej komory serca u noworodka
jest podobna. Przewaga grubości lewej komory uwidocznia
się od 20-30 dna od momentu urodzenia
• Różnicowanie i dojrzewanie komórek roboczych i układu
bodźcoprzewodzącego serca trwa do 2 roku życia
• Mięsień sercowy noworodków i niemowląt jest mało podatny.
Podatność mięśnia sercowego zwiększa się po pierwszym
roku życia. W związku z tym po pierwszym roku życia wzrasta
rola regulacji heterometrycznej czynności serca (prawo
Franka-Starlinga)
• Wrażliwość receptorów adrenergicznyh serca noworodków
jest wyraźnie mniejsza niż u osób dorosłych. W miarę wzrostu
noworodka wrażliwość receptorów wzrasta, warunkując
stopniowe nasilenie wpływu układu współczulnego na
mięsień sercowy
• Wpływ toniczny nerwu błędnego na serce u noworodków jest
minimalny. W miarę wzrostu noworodka wpływ układu
przywspółczulnego na serce wzrasta. W skutek tego częstość
skurczów serca maleje z 140 sk/min u noworodka do 70
sk/min w wieku 14-16 lat
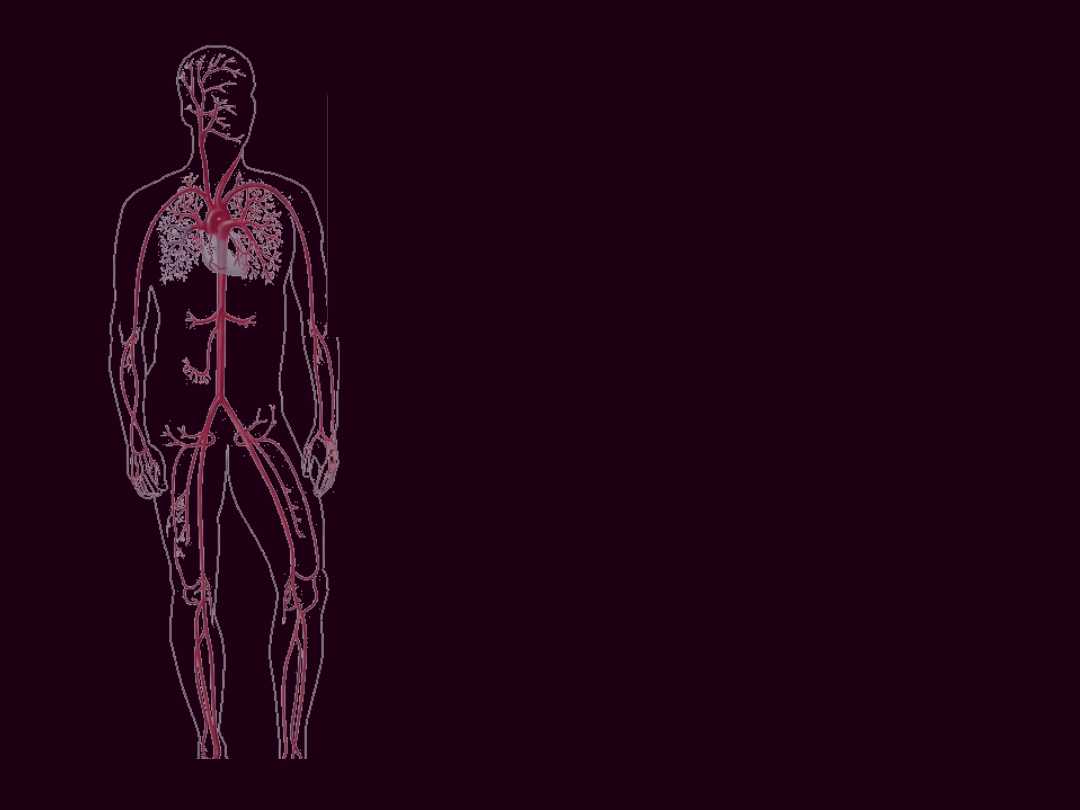
FIZJOLOGIA
NACZYŃ
KRWIONOŚNYC
H
FIZJOLOGIA
NACZYŃ
KRWIONOŚNYC
H

Tętnica i żyła obwodowa
Tętnica i żyła obwodowa
TĘTNICE i TĘTNICZKI
Aorta i tętnice o dużej średnicy zawierają dużą ilość tkanki łącznej sprężystej.
Drobne tętnice i tętniczki zawierają stosunkowo mniej elementów sprężystych,
natomiast o wiele więcej mięśni gładkich
TĘTNICE i TĘTNICZKI
Aorta i tętnice o dużej średnicy zawierają dużą ilość tkanki łącznej sprężystej.
Drobne tętnice i tętniczki zawierają stosunkowo mniej elementów sprężystych,
natomiast o wiele więcej mięśni gładkich
NACZYNIA WŁOSOWATE
Odchodzą od metarterioli (tętniczek końcowych). Znajdują się tu mięśnie
gładkie, tworzące zwieracze przedwłośniczkowe, które regulują dopływ krwi do
naczyń włosowatych. Same naczynia włosowate nie zawierają elementów
sprężystych ani mięśni gładkich. Ściany naczyń włosowatych zbudowane są z
pojedynczej
warstwy
komórek
śródbłonkowych,
połączonych
międzykomórkową substancją spajającą. W ścianach naczyń włosowatych
znajdują się pory, których wielkość zależy od rodzaju narządu. Przez pory te
mogą wraz z wodą swobodnie przenikać różne substancje rozpuszczone w
osoczu
NACZYNIA WŁOSOWATE
Odchodzą od metarterioli (tętniczek końcowych). Znajdują się tu mięśnie
gładkie, tworzące zwieracze przedwłośniczkowe, które regulują dopływ krwi do
naczyń włosowatych. Same naczynia włosowate nie zawierają elementów
sprężystych ani mięśni gładkich. Ściany naczyń włosowatych zbudowane są z
pojedynczej
warstwy
komórek
śródbłonkowych,
połączonych
międzykomórkową substancją spajającą. W ścianach naczyń włosowatych
znajdują się pory, których wielkość zależy od rodzaju narządu. Przez pory te
mogą wraz z wodą swobodnie przenikać różne substancje rozpuszczone w
osoczu
ŻYŁY i ŻYŁKI
Ściany żył i żyłek są cienkie i łatwo ulegają
rozciągnięciu.
Zawierają
stosunkowo
mało
mięśni gładkich. Błona wewnętrzna uwypukla
się tworząc w pewnych odstępach zastawki
żylne, których nie ma w drobnych żyłkach,
żyłach głównych ani w żyłach mózgu i trzewi
ŻYŁY i ŻYŁKI
Ściany żył i żyłek są cienkie i łatwo ulegają
rozciągnięciu.
Zawierają
stosunkowo
mało
mięśni gładkich. Błona wewnętrzna uwypukla
się tworząc w pewnych odstępach zastawki
żylne, których nie ma w drobnych żyłkach,
żyłach głównych ani w żyłach mózgu i trzewi
BUDOWA ŚCIANY NACZYŃ
BUDOWA ŚCIANY NACZYŃ

ŚRÓDBŁONEK
• Stanowi barierę pomiędzy krwią naczyń włosowatych a przestrzenią
zewnątrz-naczyniowa
• Spełnia funkcje metaboliczne, dezaktywując lub aktywując substancje
naczynio-aktywne znajdujące się w osoczu
• Uczestniczy w procesach koagulacji
• Wydzielają szereg substancji modulujących stan napięcia mięśniówki
gładkiej naczyń
WŁÓKNA ELASTYCZNE
• Są bardzo podatne na rozciąganie
• Tworzą gęstą sieć w warstwie głębokiej ściany naczyniowej
• Rozciągane
wytwarzają
napięcie
przeciwdziałające
ciśnieniu
krwi,
rozciągającemu naczynie
WŁÓKNA KOLAGENOWE
• Są mniej podatne na rozciąganie, stawiają większy opór rozciąganiu naczynia
niż włókna elastyczne
• Położone są głównie w warstwie środkowej i zewnętrznej ściany naczyniowej
MIĘŚNIE GŁADKIE
• Główną funkcją jest wytworzenie miogennego napięcia podstawowego ściany
naczyń oraz zmiana wielkości światła naczynia w zależności od
zapotrzebowania fizjologicznego
• Unerwione przez autonomiczny układ nerwowy
• Wrażliwe na wpływ lokalnych czynników modyfikujących wielkość światła
naczynia
ŚRÓDBŁONEK
• Stanowi barierę pomiędzy krwią naczyń włosowatych a przestrzenią
zewnątrz-naczyniowa
• Spełnia funkcje metaboliczne, dezaktywując lub aktywując substancje
naczynio-aktywne znajdujące się w osoczu
• Uczestniczy w procesach koagulacji
• Wydzielają szereg substancji modulujących stan napięcia mięśniówki
gładkiej naczyń
WŁÓKNA ELASTYCZNE
• Są bardzo podatne na rozciąganie
• Tworzą gęstą sieć w warstwie głębokiej ściany naczyniowej
• Rozciągane
wytwarzają
napięcie
przeciwdziałające
ciśnieniu
krwi,
rozciągającemu naczynie
WŁÓKNA KOLAGENOWE
• Są mniej podatne na rozciąganie, stawiają większy opór rozciąganiu naczynia
niż włókna elastyczne
• Położone są głównie w warstwie środkowej i zewnętrznej ściany naczyniowej
MIĘŚNIE GŁADKIE
• Główną funkcją jest wytworzenie miogennego napięcia podstawowego ściany
naczyń oraz zmiana wielkości światła naczynia w zależności od
zapotrzebowania fizjologicznego
• Unerwione przez autonomiczny układ nerwowy
• Wrażliwe na wpływ lokalnych czynników modyfikujących wielkość światła
naczynia
BUDOWA ŚCIANY NACZYŃ
BUDOWA ŚCIANY NACZYŃ

BUDOWA ŚCIANY NACZYŃ
BUDOWA ŚCIANY NACZYŃ
Ściana aorty
Ściana aorty
Tętnica oporowa
Tętnica oporowa
Żyła
Żyła
Duża
tętnica
1 - 2 cm
Duża
tętnica
1 - 2 cm
Tętnica
oporowa 0.1
- 1 cm
Tętnica
oporowa 0.1
- 1 cm
Tętniczka
20 – 200
μm
Tętniczka
20 – 200
μm
Naczynie
włosowate 5 – 7
μ
Naczynie
włosowate 5 – 7
μ
Duża żyła
1.5 – 3
cm
Duża żyła
1.5 – 3
cm
Żyła średnia
0.15 – 1.5
cm
Żyła średnia
0.15 – 1.5
cm
Żyłka
drobna
20 – 500
μm
Żyłka
drobna
20 – 500
μm
1/5
1/5
1/5
1/5
1/3
1/3
1/5
1/5
1/1
1/1
1/8
1/8
1/12
1/12
1/12
1/12
1/9
1/9
1/12
1/12
1/9
1/9
śródbłonek
śródbłonek
składowa elastyczna
składowa elastyczna
składowa mięśniowa
składowa mięśniowa
składowa kolagenowa
składowa kolagenowa
Struktura ściany naczyń oraz stosunek grubości ściany naczynia do jego promienia (h/r)
w stanie luźnym (po lewej) i podczas skurczu mięśni gładkich ściany naczyniowej (po
prawej)
Struktura ściany naczyń oraz stosunek grubości ściany naczynia do jego promienia (h/r)
w stanie luźnym (po lewej) i podczas skurczu mięśni gładkich ściany naczyniowej (po
prawej)
według: E. Neil, B. Folkow. Circulation. Oxford University Press, 1971)
według: E. Neil, B. Folkow. Circulation. Oxford University Press, 1971)

• Stosunek
h/r
drobnych tętnic i tętniczek (naczyń
oporowych) jest większy, niż dużych tętnic. Składowa
mięśniowa naczyń oporowych jest większa niż składowa
elastyczna. Są mniej podatne na rozciąganie. Regulacja
przepływu krwi przez takie naczynia odbywa się poprzez
zmianę wielkości ich promienia na drodze zmian napięcia
mięśni gładkich. Napięcie bierne, powstające na skutek
rozciągania elementów elastycznych w naczyniach oporowych
ma znacznie mniejszy wpływ na warunki przepływu niż
skurcz mięśni gładkich
• Duża
wartość
stosunku
h/r
warunkuje
większe
zmniejszenie światła naczyń, spowodowane skurczem
mięsni gładkich. Już
• Stosunek
h/r
drobnych tętnic i tętniczek (naczyń
oporowych) jest większy, niż dużych tętnic. Składowa
mięśniowa naczyń oporowych jest większa niż składowa
elastyczna. Są mniej podatne na rozciąganie. Regulacja
przepływu krwi przez takie naczynia odbywa się poprzez
zmianę wielkości ich promienia na drodze zmian napięcia
mięśni gładkich. Napięcie bierne, powstające na skutek
rozciągania elementów elastycznych w naczyniach oporowych
ma znacznie mniejszy wpływ na warunki przepływu niż
skurcz mięśni gładkich
• Duża
wartość
stosunku
h/r
warunkuje
większe
zmniejszenie światła naczyń, spowodowane skurczem
mięsni gładkich. Już
skurcz tylko zewnętrznych warstw mięśni
gładkich powoduje przesuniecie znacznej
ilości tkanki ściany naczynia w kierunku
jego światła, powodując szybkie i znaczne
zmniejszenie jego promienia
skurcz tylko zewnętrznych warstw mięśni
gładkich powoduje przesuniecie znacznej
ilości tkanki ściany naczynia w kierunku
jego światła, powodując szybkie i znaczne
zmniejszenie jego promienia
BUDOWA ŚCIANY NACZYŃ
BUDOWA ŚCIANY NACZYŃ

Płuca
10–12%
Płuca
10–12%
Serce
8–11%
Serce
8–11%
Część tętnicza
10–12%
Część tętnicza
10–12%
Naczynia
włosowate 4–5%
Naczynia
włosowate 4–5%
Część żylna 60 – 70 %
Duże żyły
Duże żyły
Drobne
żyły
i
żyłki
Drobne
żyły
i
żyłki
DYSTRYBUCJA KRWI W POSZCZEGÓLNYCH CZĘŚCIACH UKŁADU
KRĄŻENIA
DYSTRYBUCJA KRWI W POSZCZEGÓLNYCH CZĘŚCIACH UKŁADU
KRĄŻENIA

• Objętość całkowita krwi (Q) przepływającej przez układ
krążenia w ciągu minuty jest równa pojemności minutowej
serca (CO). Pojemność minutowa jest to ilość krwi tłoczonej
przez jedną z komór serca w czasie jednej minuty
• Objętość całkowita krwi (Q) przepływającej przez układ
krążenia w ciągu minuty jest równa pojemności minutowej
serca (CO). Pojemność minutowa jest to ilość krwi tłoczonej
przez jedną z komór serca w czasie jednej minuty
CO (L/min) = SV • HR
CO (L/min) = SV • HR
• Objętość wyrzutowa (SV; ml) jest to ilość krwi wtłaczanej
przez
każdą
komorę
w
ciągu
jednego
cyklu
hemodynamicznego serca do odpowiedniego zbiornika
tętniczego
• Częstość skurczów serca (HR; sk/min) jest to liczba cykli
hemodynamicznych serca w ciągu jednej minuty
• Objętość wyrzutowa (SV; ml) jest to ilość krwi wtłaczanej
przez
każdą
komorę
w
ciągu
jednego
cyklu
hemodynamicznego serca do odpowiedniego zbiornika
tętniczego
• Częstość skurczów serca (HR; sk/min) jest to liczba cykli
hemodynamicznych serca w ciągu jednej minuty
PODSTAWOWE POJĘCIA HEMODYNAMICZNE
PODSTAWOWE POJĘCIA HEMODYNAMICZNE
• Krew wprawiana jest w ruch siłą wytwarzaną przez ciśnienie
napędowe – różnicę ciśnień pomiędzy początkiem dużego
krążenia w aorcie (MAP ≈
95
mmHg ) a jego końcem w
prawym przedsionku (M
Art
P ≈
5
mmHg)
• Ciśnienie napędowe będzie zmniejszać się w miarę
przesuwania krwi w naczyniach w związku z pokonywaniem
oporu tarcia. Profil podłużny ciśnienia na charakter linii
spadającej
• Krew wprawiana jest w ruch siłą wytwarzaną przez ciśnienie
napędowe – różnicę ciśnień pomiędzy początkiem dużego
krążenia w aorcie (MAP ≈
95
mmHg ) a jego końcem w
prawym przedsionku (M
Art
P ≈
5
mmHg)
• Ciśnienie napędowe będzie zmniejszać się w miarę
przesuwania krwi w naczyniach w związku z pokonywaniem
oporu tarcia. Profil podłużny ciśnienia na charakter linii
spadającej
C
iś
n
ie
n
ie
k
rw
i
C
iś
n
ie
n
ie
k
rw
i
tętnic
e
tętnic
e
tętnicz
ki
tętnicz
ki
n. włosowate
n. włosowate
żyły
żyły
Rozszerzenie
tętniczek
Rozszerzenie
tętniczek
Zwężenie
tętniczek
Zwężenie
tętniczek
Norma
Norma
PODSTAWOWE POJĘCIA HEMODYNAMICZNE
PODSTAWOWE POJĘCIA HEMODYNAMICZNE

PODSTAWOWE POJĘCIA HEMODYNAMICZNE
PODSTAWOWE POJĘCIA HEMODYNAMICZNE
• Podstawowe prawo hemodynamiki jest analogiczne do prawa
Ohma
• Podstawowe prawo hemodynamiki jest analogiczne do prawa
Ohma
MAP - M
Atr
P = Q • R
MAP - M
Atr
P = Q • R
•
MAP – średnie ciśnienie tętnicze w aorcie, M
Atr
P – średnie ciśnienie w prawym przedsionku, Q = CO –
pojemność minutowa, R = TPR – całkowity opór obwodowy
MAP – średnie ciśnienie tętnicze w aorcie, M
Atr
P – średnie ciśnienie w prawym przedsionku, Q = CO –
pojemność minutowa, R = TPR – całkowity opór obwodowy
• Ciśnienie średnie mierzone na tętnicy ramiennej jest ok. 5
mmHg mniejsze, niż w aorcie. Ciśnienie w prawym
przedsionku jest równe ok. 5 mmHg. Biorąc poprawkę na te
wartości:
• Ciśnienie średnie mierzone na tętnicy ramiennej jest ok. 5
mmHg mniejsze, niż w aorcie. Ciśnienie w prawym
przedsionku jest równe ok. 5 mmHg. Biorąc poprawkę na te
wartości:
MAP = CO • TPR
lub
TPR =
MAP = CO • TPR
lub
TPR =
MAP
CO

PODSTAWOWE POJĘCIA HEMODYNAMICZNE
PODSTAWOWE POJĘCIA HEMODYNAMICZNE
• Najwyższą wartość, jaką osiąga ciśnienie tętnicze w cyklu
sercowym, określamy jako ciśnienie skurczowe (SP) równe
ok. 120 mmHg, a wartość najniższą jako ciśnienie
rozkurczowe (DP) – ok. 70 mmHg
• Wartość ciśnienia tętniczego waha się pomiędzy ciśnieniem
skurczowym i rozkurczowym, przy czym szybkość wzrostu
ciśnienia krwi w aorcie podczas wyrzutu jest znacznie
większa od szybkości jego spadku w okresie rozkurczu
mięśnia sercowego
• Ciśnienie średnie, panujące w części tętniczej układu
sercowo-naczyniowego jest równe:
• Najwyższą wartość, jaką osiąga ciśnienie tętnicze w cyklu
sercowym, określamy jako ciśnienie skurczowe (SP) równe
ok. 120 mmHg, a wartość najniższą jako ciśnienie
rozkurczowe (DP) – ok. 70 mmHg
• Wartość ciśnienia tętniczego waha się pomiędzy ciśnieniem
skurczowym i rozkurczowym, przy czym szybkość wzrostu
ciśnienia krwi w aorcie podczas wyrzutu jest znacznie
większa od szybkości jego spadku w okresie rozkurczu
mięśnia sercowego
• Ciśnienie średnie, panujące w części tętniczej układu
sercowo-naczyniowego jest równe:
MAP = TPR
•
CO
MAP = TPR
•
CO
MAP = DP +1/3(SP-
DP)
MAP = DP +1/3(SP-
DP)

PODSTAWOWE POJĘCIA HEMODYNAMICZNE
PODSTAWOWE POJĘCIA HEMODYNAMICZNE
• Bezpośrednią przyczyną ciśnienia tętniczego krwi są siły
sprężyste biernego napięcia sprężystego ścian tętnic
rozciąganych objętością krwi wyrzucanej przez komorę serca
• Zależność pomiędzy napięciem rozciąganej ściany naczynia a
wywołanym przez nie ciśnieniem wewnątrz naczynia określa
prawo Laplace’a. W oparciu o prawo Laplace’a Frank (1920)
w
celu
określenia
napięcia
wprowadził
poprawkę,
uwzględniającą grubość ściany naczynia
• Bezpośrednią przyczyną ciśnienia tętniczego krwi są siły
sprężyste biernego napięcia sprężystego ścian tętnic
rozciąganych objętością krwi wyrzucanej przez komorę serca
• Zależność pomiędzy napięciem rozciąganej ściany naczynia a
wywołanym przez nie ciśnieniem wewnątrz naczynia określa
prawo Laplace’a. W oparciu o prawo Laplace’a Frank (1920)
w
celu
określenia
napięcia
wprowadził
poprawkę,
uwzględniającą grubość ściany naczynia
T = P
t
• r
T = P
t
• r
T = P
t
•
T = P
t
•
r
r
h
h
T- napięcie sprężyste ściany, P
t
– ciśnienie transmuralne (różnica ciśnień wewnątrz naczynia a
ciśnieniem otaczającym, r – promień naczynia, h – grubość ściany naczynia
T- napięcie sprężyste ściany, P
t
– ciśnienie transmuralne (różnica ciśnień wewnątrz naczynia a
ciśnieniem otaczającym, r – promień naczynia, h – grubość ściany naczynia

PODSTAWOWE POJĘCIA HEMODYNAMICZNE
PODSTAWOWE POJĘCIA HEMODYNAMICZNE
• Krew przepływająca przez narząd musi pokonać opór
stawiany przez szeregowo połączone ze sobą tętnice,
tętniczki, naczynia włosowate, żyłki i żyły
Całkowity opór układu krążenia danego narządu zgodnie z
prawem Kirchoffa jest równy:
• Krew przepływająca przez narząd musi pokonać opór
stawiany przez szeregowo połączone ze sobą tętnice,
tętniczki, naczynia włosowate, żyłki i żyły
Całkowity opór układu krążenia danego narządu zgodnie z
prawem Kirchoffa jest równy:
R
narządu
= R
tetnic
+ R
tętniczek
+ R
n.wł
+ R
żyłek
+ R
żył
R
narządu
= R
tetnic
+ R
tętniczek
+ R
n.wł
+ R
żyłek
+ R
żył
• Aorta
rozgałęzia
się
na
tętnice,
które
zaopatrują
poszczególne narządy i części organizmu. Poszczególne
obszary krążeniowe są połączone są ze sobą równolegle.
Wobec tego zgodnie z prawem Kirchoffa:
• Aorta
rozgałęzia
się
na
tętnice,
które
zaopatrują
poszczególne narządy i części organizmu. Poszczególne
obszary krążeniowe są połączone są ze sobą równolegle.
Wobec tego zgodnie z prawem Kirchoffa:
= + +
+ ...
= + +
+ ...
1
1
TPR
TPR
1
1
R
1
R
1
1
1
R
2
R
2
1
1
R
3
R
3
1
1
R
n
R
n
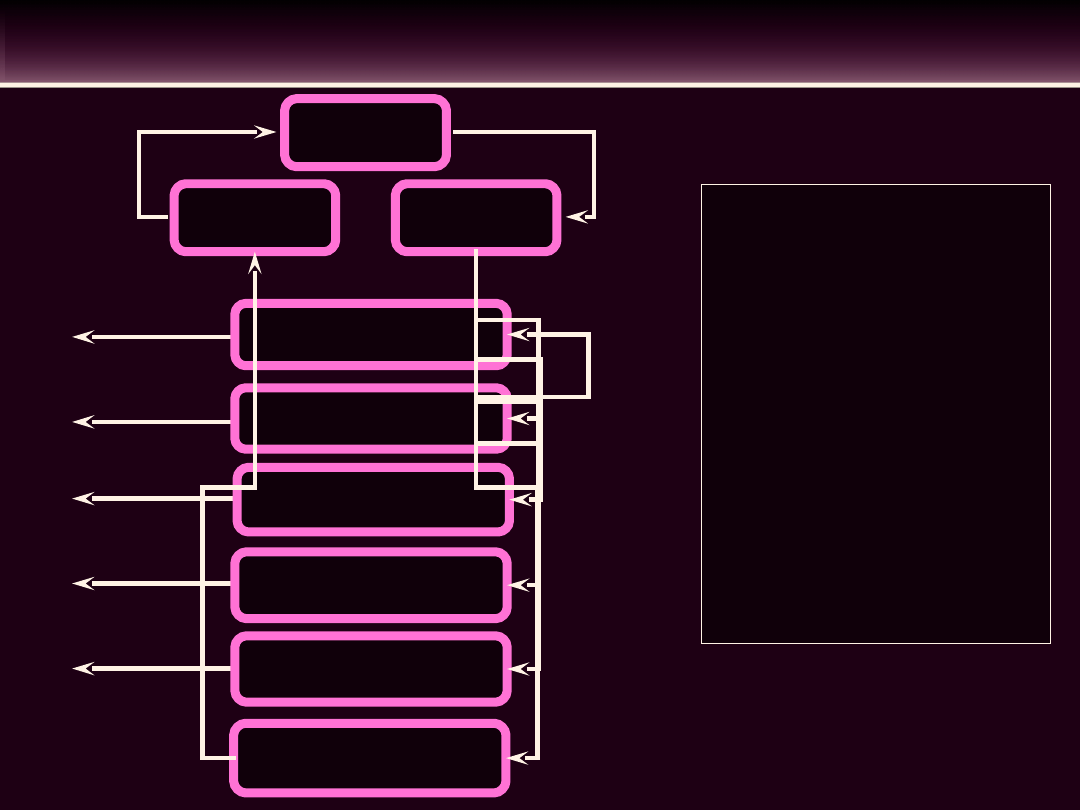
Prawa
połowa serca
Prawa
połowa serca
Lewa połowa
serca
Lewa połowa
serca
Płuca
Płuca
Naczynia
wieńcowe
Naczynia
wieńcowe
Mózg
Mózg
Mięśnie
Mięśnie
Trzewia
Trzewia
Nerki
Nerki
Skóra, kości i
inne tkanki
Skóra, kości i
inne tkanki
100
%
100
%
100
%
100
%
5 %
5 %
15 %
15 %
15 %
15 %
35 %
35 %
20 %
20 %
10 %
10 %
DYSTRYBUCJA KRWI W POSZCZEGÓLNYCH OBSZARACH
KRĄŻENIOWYCH
DYSTRYBUCJA KRWI W POSZCZEGÓLNYCH OBSZARACH
KRĄŻENIOWYCH
• Opór naczyniowy prze-
pływu
krwi
jest
znacznie
większy
w
pojedynczym
narządzie,
niż
całkowity
opór
obwodowy (TPR).
• Przy
prawie
takim
samym ciśnieniu krwi
jak w aorcie, przepływ
krwi
przez
poszczególne obszary
krążeniowe
sta-nowi
tylko
mały
procent
pojemności minutowej
(CO)
• Opór naczyniowy prze-
pływu
krwi
jest
znacznie
większy
w
pojedynczym
narządzie,
niż
całkowity
opór
obwodowy (TPR).
• Przy
prawie
takim
samym ciśnieniu krwi
jak w aorcie, przepływ
krwi
przez
poszczególne obszary
krążeniowe
sta-nowi
tylko
mały
procent
pojemności minutowej
(CO)

CZYNNIKI WPŁYWAJĄCE NA OPÓR PRZEPŁYWU KRWI W
NACZYNIACH
CZYNNIKI WPŁYWAJĄCE NA OPÓR PRZEPŁYWU KRWI W
NACZYNIACH
•Zgodnie z prawem Poiseuilla:
•Zgodnie z prawem Poiseuilla:
Q = ΔP •
Q = ΔP •
r
4
• π
8Lη
ΔP – różnica ciśnień podtrzymująca ruch cieczy, r – promień rurki , L – długość rurki, η – lepkość cieczy
ΔP – różnica ciśnień podtrzymująca ruch cieczy, r – promień rurki , L – długość rurki, η – lepkość cieczy
R =
R = r
4
• π
8Lη
Q =
Q =
R
ΔP
• Długość naczyń w układzie sercowo-naczyniowym oraz
lepkość krwi są to wartości w miarę stałe, nie ulegające
istotnym zmianą w krótkim czasie. W związku z tym wielkość
przepływu krwi przez narząd jest regulowana przez wielkość
promienia naczyń w tym narządzie
• Długość naczyń w układzie sercowo-naczyniowym oraz
lepkość krwi są to wartości w miarę stałe, nie ulegające
istotnym zmianą w krótkim czasie. W związku z tym wielkość
przepływu krwi przez narząd jest regulowana przez wielkość
promienia naczyń w tym narządzie
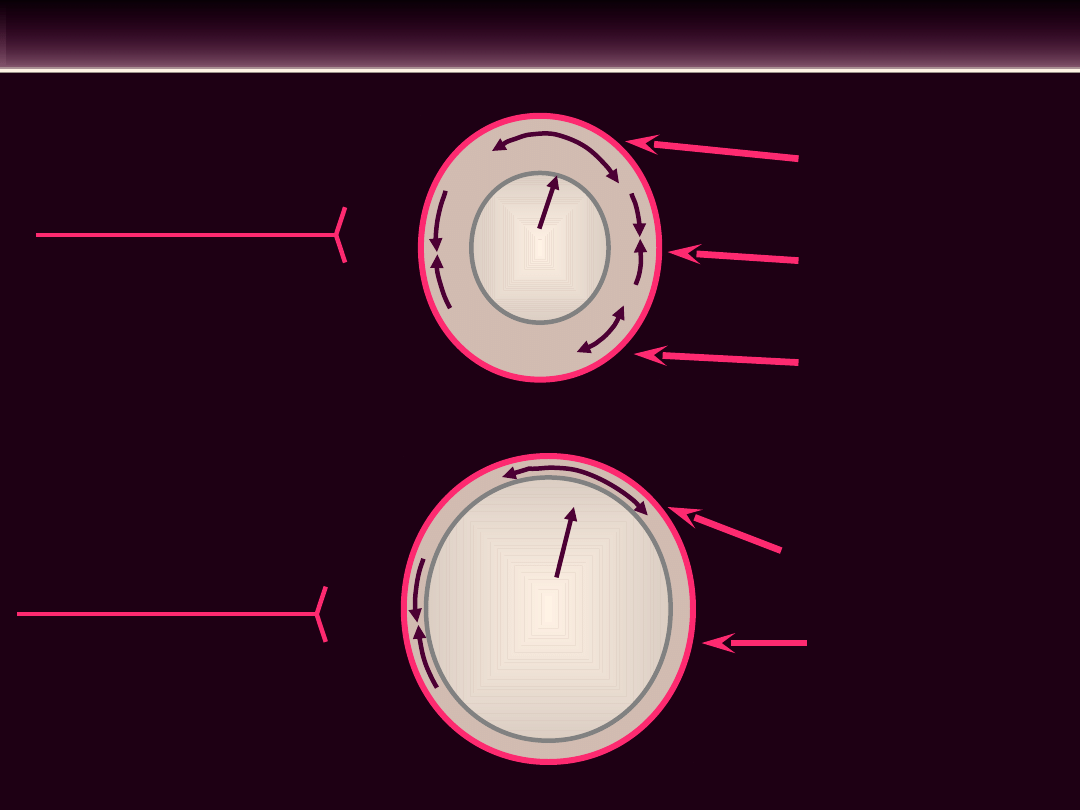
CZYNNIKI WARUNKUJĄCE NAPIĘCIE ŚCIANY
NACZYNIOWEJ
CZYNNIKI WARUNKUJĄCE NAPIĘCIE ŚCIANY
NACZYNIOWEJ
TĘTNICE
TĘTNICE
ŻYŁY
ŻYŁY
WPŁYW LOKALNY
WPŁYW LOKALNY
WPŁYW ODRUCHOWY
WPŁYW ODRUCHOWY
Napięcie okrężne
(składowa bierna)
Napięcie okrężne
(składowa bierna)
Napięcie
podstawo-we
miogenne (skła-
dowa czynna)
Napięcie
podstawo-we
miogenne (skła-
dowa czynna)
Rozkurczowe
dzia-łanie
metabolitów
na
mięśnie gładki
Rozkurczowe
dzia-łanie
metabolitów
na
mięśnie gładki
Napięcie okrężne
(składowa bierna)
Napięcie okrężne
(składowa bierna)
P
P
Ucisk zewnętrzny
Ucisk zewnętrzny
NA
→
α
1
NA
→
α
1
Włókna współczulne
Włókna współczulne
NA
→
α
1
NA
→
α
1
Włókna współczulne
Włókna współczulne
REGULACJA NAPIĘCIA ŚCIANY NACZYNIOWEJ W
POSZCZEGÓLNYCH NARZĄDACH
REGULACJA NAPIĘCIA ŚCIANY NACZYNIOWEJ W
POSZCZEGÓLNYCH NARZĄDACH
Maksymalna
aktywność
współczulna
Maksymalna
aktywność
współczulna
Brak
aktywności
współczulnej
Brak
aktywności
współczulnej
Maksymalny
metabolizm
Maksymalny
metabolizm
Tętnice
mózgowe, wieńcowe i mięśni
szkieletowych
cechuje duże napięcie podsta-
wowe pochodzenia miogennego. Wielkość
przepły-wu krwi przez te narządy jest niewiele
większa
od
ich
zapotrzebowania
metabolicznego.
Wzrost
aktywności
współczulnej nie prowadzi do znacznego
zwężenia światła tętnic z powodu gromadzenia
znacznej ilości metabolitów, wywierających
przeciwstawny wpływ na mięśnie gładkie. Z
powodu dużego napięcia podstawowego w tych
tętnicach spadek aktywności współczulnej nie
prowadzi do znacznego wzrostu przepływu krwi
Tętnice
mózgowe, wieńcowe i mięśni
szkieletowych
cechuje duże napięcie podsta-
wowe pochodzenia miogennego. Wielkość
przepły-wu krwi przez te narządy jest niewiele
większa
od
ich
zapotrzebowania
metabolicznego.
Wzrost
aktywności
współczulnej nie prowadzi do znacznego
zwężenia światła tętnic z powodu gromadzenia
znacznej ilości metabolitów, wywierających
przeciwstawny wpływ na mięśnie gładkie. Z
powodu dużego napięcia podstawowego w tych
tętnicach spadek aktywności współczulnej nie
prowadzi do znacznego wzrostu przepływu krwi
Maksymalna
aktywność
współczulna
Maksymalna
aktywność
współczulna
P
rz
e
p
ły
w
k
rw
i
p
rz
e
z
n
a
rz
ą
d
P
rz
e
p
ły
w
k
rw
i
p
rz
e
z
n
a
rz
ą
d
Maksymalnie możliwy przepływ
Maksymalnie możliwy przepływ
Przepływ
spoczynkowy
Przepływ
spoczynkowy
P
rz
e
p
ły
w
k
rw
i
p
rz
e
z
n
a
rz
ą
d
P
rz
e
p
ły
w
k
rw
i
p
rz
e
z
n
a
rz
ą
d
Maksymalnie możliwy przepływ
Maksymalnie możliwy przepływ
Przepływ
spoczynkowy
Przepływ
spoczynkowy
Brak
aktywności
współczulnej
Brak
aktywności
współczulnej
Maksymalny
metabolizm
Maksymalny
metabolizm
Tętnice
skórne, nerkowe i narządów jamy
brzusznej
cechuje niewielkie napięcie podsta-
wowe, pochodzenia miogennego. Wielkość
przepły-wu krwi przez te narządy jest znacznie
większa,
niż
ich
zapotrzebowanie
metaboliczne. Wzrost aktyw-ności współczulnej
prowadzi do znacznego zwęże-nia światła
tętnic i zmniejszenia przepływu krwi przez
narząd. Mimo zmniejszenia przepływu jest on i
tak wystarczający, by pokryć zapotrzebowanie
metaboliczne
tych
narządów.
Wzrost
zapotrzebowania
metabolicznego
też
nie
prowadzi do znacznych zmian przepływu
Tętnice
skórne, nerkowe i narządów jamy
brzusznej
cechuje niewielkie napięcie podsta-
wowe, pochodzenia miogennego. Wielkość
przepły-wu krwi przez te narządy jest znacznie
większa,
niż
ich
zapotrzebowanie
metaboliczne. Wzrost aktyw-ności współczulnej
prowadzi do znacznego zwęże-nia światła
tętnic i zmniejszenia przepływu krwi przez
narząd. Mimo zmniejszenia przepływu jest on i
tak wystarczający, by pokryć zapotrzebowanie
metaboliczne
tych
narządów.
Wzrost
zapotrzebowania
metabolicznego
też
nie
prowadzi do znacznych zmian przepływu
według: D.E. Mohrman, L.J. Heller. Cardiovascular Physiology. McGraw-Hill, 1997)
według: D.E. Mohrman, L.J. Heller. Cardiovascular Physiology. McGraw-Hill, 1997)
POWRÓT ŻYLNY
POWRÓT ŻYLNY
Z naczyń
włosowatych
P
pv
=
7
mmHg
P
pv
=
7
mmHg
Klatka piersiowa
Klatka piersiowa
P
kl
0
mmHg
P
kl
0
mmHg
według: D.E. Mohrman, L.J. Heller. Cardiovascular Physiology. McGraw-Hill,
1997)
według: D.E. Mohrman, L.J. Heller. Cardiovascular Physiology. McGraw-Hill,
1997)
Powrót żylny
P
cv
P
cv
Czynnościowa część
obwodowa zbiornika
żylnego
Czynnościowa część
obwodowa zbiornika
żylnego
Czynnościowa
część
centralna zbiornika żylnego
(składa
się
z
prawego
przedsionka i żył głównych
górnej i dolnej)
Czynnościowa
część
centralna zbiornika żylnego
(składa
się
z
prawego
przedsionka i żył głównych
górnej i dolnej)
Wielkość
przepływu
pomiędzy
zbior-nikiem żylnym obwodowym
a częścią centralną zbiornika
żylnego
określa
podstawowe
prawo hemodynamiki:
Wielkość
przepływu
pomiędzy
zbior-nikiem żylnym obwodowym
a częścią centralną zbiornika
żylnego
określa
podstawowe
prawo hemodynamiki:
Q =
Q =
ΔP
R
ΔP – różnica ciśnień pomiędzy
częścią
obwodową
zbiornika
żylne-go a jego częścią centralną
R – opór żył obwodowych
ΔP – różnica ciśnień pomiędzy
częścią
obwodową
zbiornika
żylne-go a jego częścią centralną
R – opór żył obwodowych
POWRÓT ŻYLNY
POWRÓT ŻYLNY
P
o
w
ró
t
ży
ln
y
(L
/m
in
)
P
o
w
ró
t
ży
ln
y
(L
/m
in
)
Ciśnienie centralnej części zbiornika żylnego
(mmHg)
Ciśnienie centralnej części zbiornika żylnego
(mmHg)
0 2 4 6 8
10
0 2 4 6 8
10
10
10
8
8
6
6
4
4
2
2
K
rz
yw
a k
on
tro
ln
a p
ow
ro
tu
ży
ln
eg
o
K
rz
yw
a k
on
tro
ln
a p
ow
ro
tu
ży
ln
eg
o
W
zr
os
t
ob
ję
to
śc
i
kr
w
i
lu
b
na
pię
cia
ży
ł
ob
w
od
ow
yc
h
W
zr
os
t
ob
ję
to
śc
i
kr
w
i
lu
b
na
pię
cia
ży
ł
ob
w
od
ow
yc
h
Sp
ad
ek
ob
ję
to
śc
i
kr
w
i
lu
b
na
pię
cia
ży
ł
ob
w
od
ow
yc
h
Sp
ad
ek
ob
ję
to
śc
i
kr
w
i
lu
b
na
pię
cia
ży
ł
ob
w
od
ow
yc
h
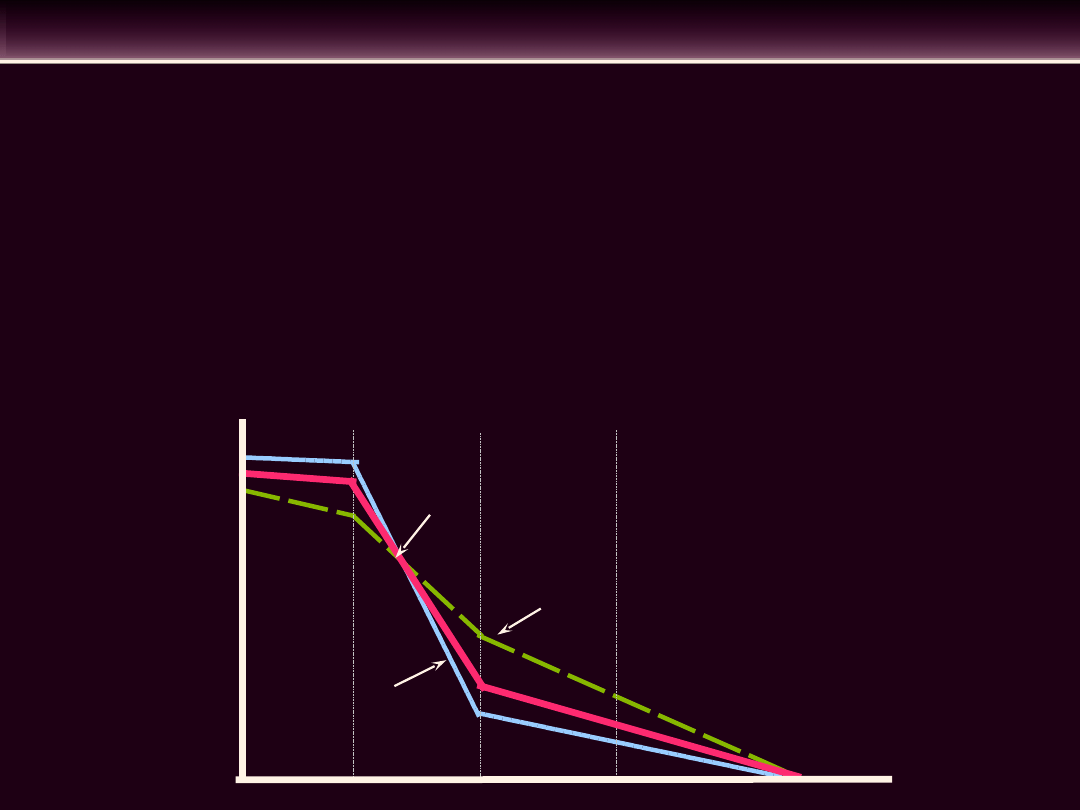
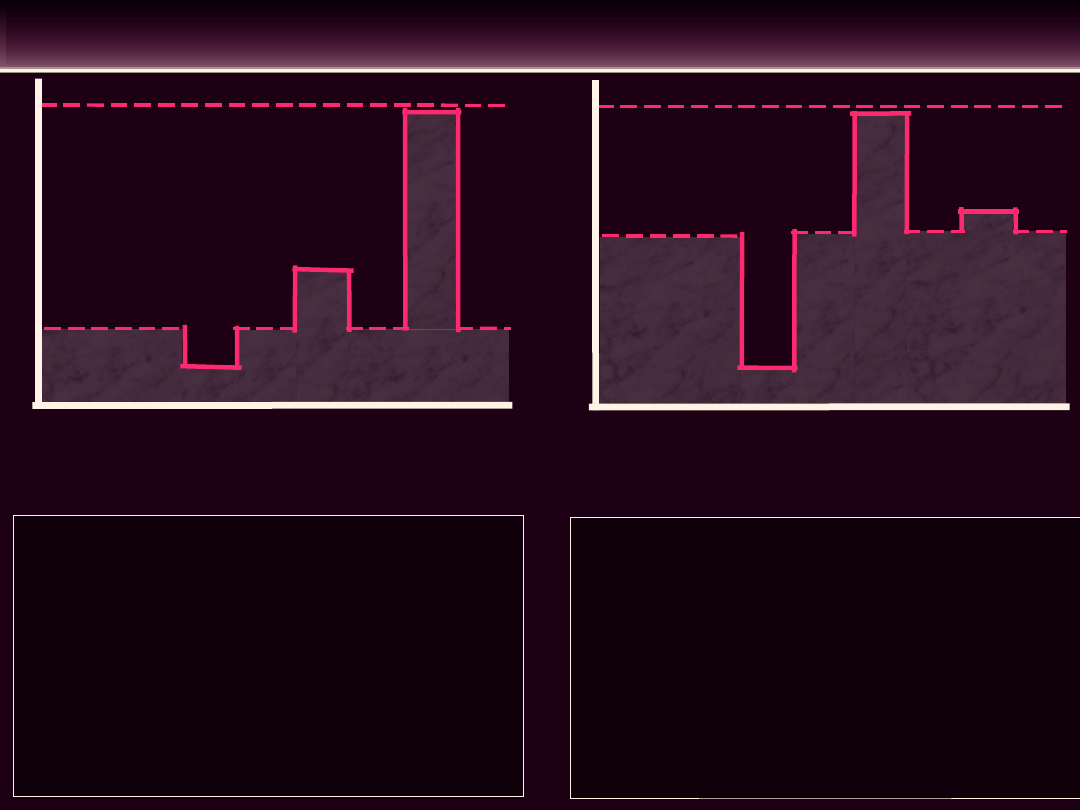
• Krzywa
powrotu
żylnego
demonstruje jaki wpływ wywiera
ciśnienie
centralnej
części
zbiornika żylnego na wielkość
powrotu żylnego pod warunkiem,
że pozostałe czynniki pozostaną
bez zmian
• Wzrost lub spadek ciśnienia w
obwodowym
zbiorniku
żylnym
również powoduje zmiany wielkości
powrotu żylnego
Wzrost ciśnienia w obwodowym
zbiorniku żylnym następuje, gdy:
- wzrasta objętość krwi w zbiorniku
żylnym
-
wzrasta
napięcie
ścian
żył
(niebieska krzywa)
Spadek ciśnienia w obwodowym
zbiorniku żylnym następuje, gdy:
- maleje objętość krwi w zbiorniku
żylnym
- maleje napięcie ścian żył (zielona
krzywa)
• Krzywa
powrotu
żylnego
demonstruje jaki wpływ wywiera
ciśnienie
centralnej
części
zbiornika żylnego na wielkość
powrotu żylnego pod warunkiem,
że pozostałe czynniki pozostaną
bez zmian
• Wzrost lub spadek ciśnienia w
obwodowym
zbiorniku
żylnym
również powoduje zmiany wielkości
powrotu żylnego
Wzrost ciśnienia w obwodowym
zbiorniku żylnym następuje, gdy:
- wzrasta objętość krwi w zbiorniku
żylnym
-
wzrasta
napięcie
ścian
żył
(niebieska krzywa)
Spadek ciśnienia w obwodowym
zbiorniku żylnym następuje, gdy:
- maleje objętość krwi w zbiorniku
żylnym
- maleje napięcie ścian żył (zielona
krzywa)
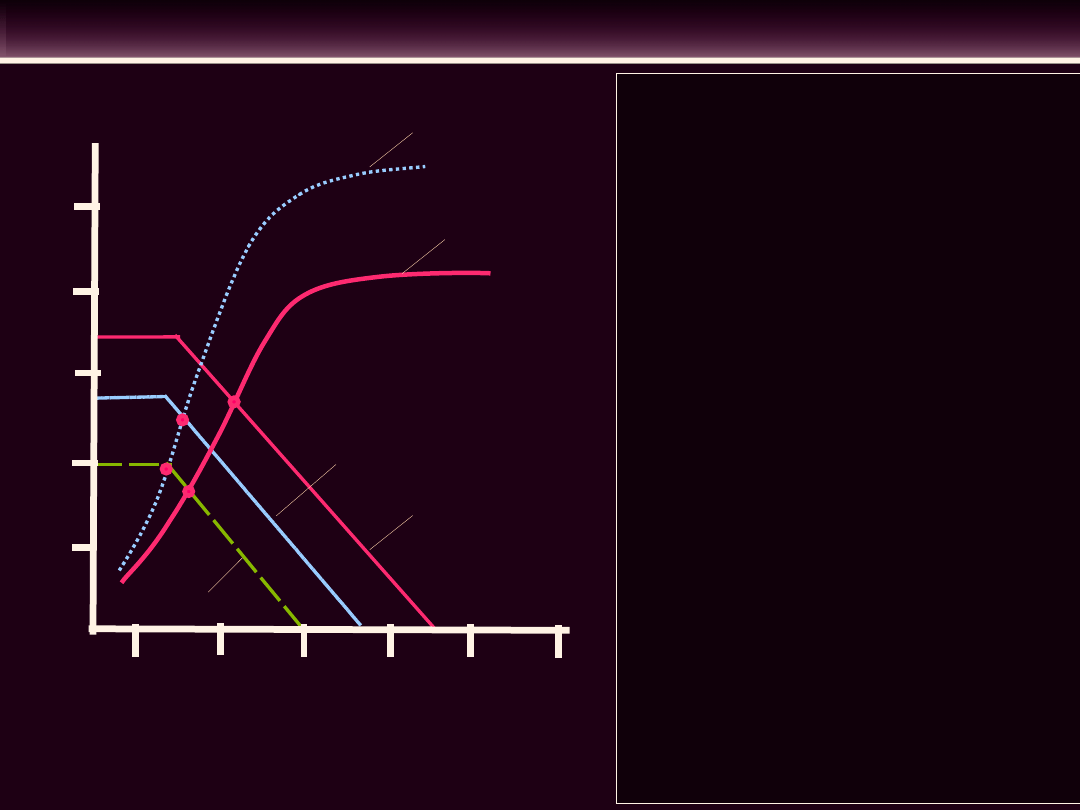
POJEMNOŚĆ MINUTOWA
POJEMNOŚĆ MINUTOWA
P
o
je
m
n
o
ść
m
in
u
to
w
a
lu
b
p
o
w
ró
t
ży
ln
y
(L
/m
in
)
P
o
je
m
n
o
ść
m
in
u
to
w
a
lu
b
p
o
w
ró
t
ży
ln
y
(L
/m
in
)
Ciśnienie centralnej części zbiornika żylnego
(mmHg)
Ciśnienie centralnej części zbiornika żylnego
(mmHg)
10
10
8
8
6
6
4
4
2
2
Wzrost aktywności współczulnej,
wzrost HR i kurczliwości mięśnia
sercowego
Wzrost aktywności współczulnej,
wzrost HR i kurczliwości mięśnia
sercowego
Spoczynkowy
powrót
żylny
Spoczynkowy
powrót
żylny
Spoczynkowa pojemność
minutowa
Spoczynkowa pojemność
minutowa
0 2 4 6 8
10
0 2 4 6 8
10
1
1
2
2
3
3
4
4
Tuż po krwotoku
Tuż po krwotoku
Wzrost aktywności układu
współczulnego,
skurcz
mięśni gładkich żył
Wzrost aktywności układu
współczulnego,
skurcz
mięśni gładkich żył
• W stanie stabilnym powrót żylny
jest równy pojemności minutowej
przy pewnej wartości ciśnienia
centralnej części zbiornika żylnego
– punkt
1
• Krwotok prowadzi do zmniejszenia
krwi krążącej, spadku powrotu
żylnego i obniżenia ciśnienia w
centralnej części zbiornika żylnego
a
zatem
spadku
pojemności
minutowej serca – punkt
2
• Wzrost aktywności współczulnej
prowadzi
do
przyspieszenia
czynności
serca
i
wzrostu
kurczliwości mięśnia sercowego.
Pojemność
minutowa
serca
nieznacznie
wzrasta
przy
jednoczesnym spadku ciśnienia w
centralnej części zbiornika żylnego
– punkt
3
• Równolegle
wzrost
aktywności
współczulnej prowadzi do skurczu
mięsni gładkich żył. W skutek tego
wzrasta ciśnienie żylne obwodowe i
powrót żylny. Te zmiany prowadza
do wzrostu ciśnienia w centralnej
części zbiornika żylnego a za tym
pojemności minutowej serca –
punkt
4
• W stanie stabilnym powrót żylny
jest równy pojemności minutowej
przy pewnej wartości ciśnienia
centralnej części zbiornika żylnego
–
punkt
1
• Krwotok prowadzi do zmniejszenia
krwi krążącej, spadku powrotu
żylnego i obniżenia ciśnienia w
centralnej części zbiornika żylnego
a
zatem
spadku
pojemności
minutowej serca –
punkt
2
• Wzrost aktywności współczulnej
prowadzi
do
przyspieszenia
czynności
serca
i
wzrostu
kurczliwości mięśnia sercowego.
Pojemność
minutowa
serca
nieznacznie
wzrasta
przy
jednoczesnym spadku ciśnienia w
centralnej części zbiornika żylnego
–
punkt
3
• Równolegle
wzrost
aktywności
współczulnej prowadzi do skurczu
mięsni gładkich żył. W skutek tego
wzrasta ciśnienie żylne obwodowe i
powrót żylny. Te zmiany prowadza
do wzrostu ciśnienia w centralnej
części zbiornika żylnego a za tym
pojemności minutowej serca –
punkt
4
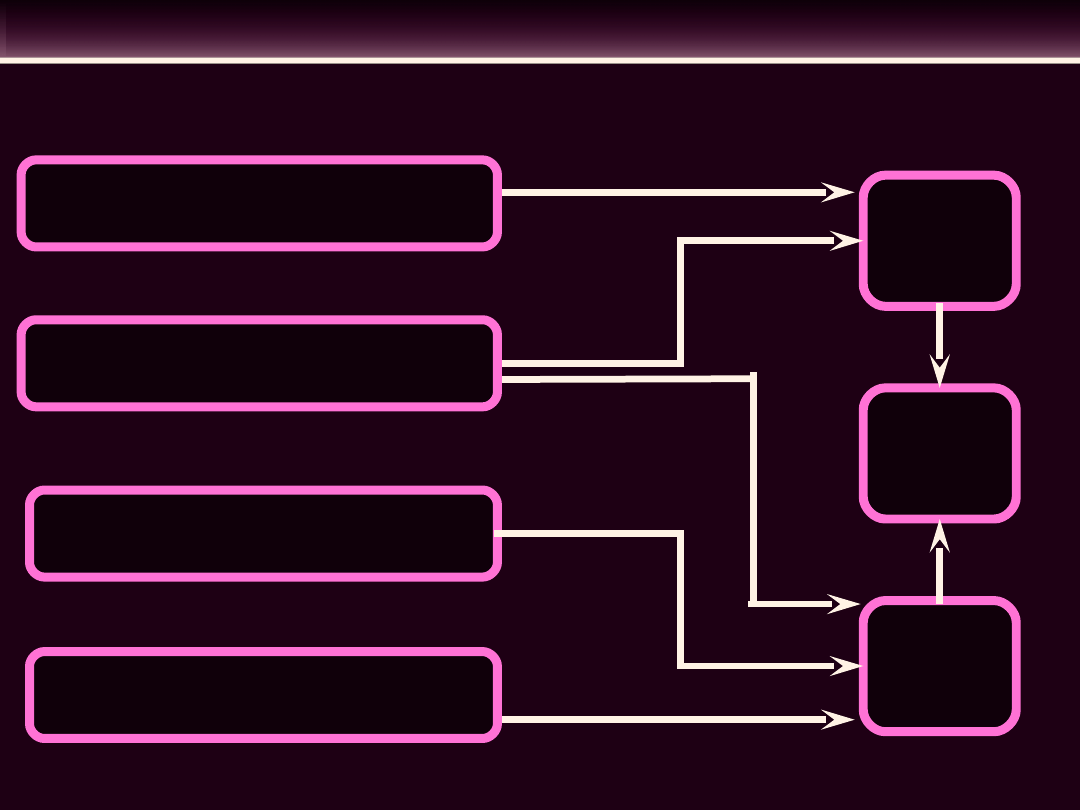
CO
CO
HR
HR
SV
SV
Ciśnienie tętnicze krwi
Ciśnienie tętnicze krwi
Wzrost aktywności
układu
przywspółczulnego
Wzrost aktywności
układu
przywspółczulnego
-
+
Wzrost aktywności
układu współczulnego
Wzrost aktywności
układu współczulnego
+
+
-
wpływ chronotropowy ujemny
wpływ chronotropowy ujemny
wpływ chronotropowy dodatni
wpływ chronotropowy dodatni
wpływ inotropowy dodatni
wpływ inotropowy dodatni
obciążenie następcze
obciążenie następcze
obciążenie wstępne
obciążenie wstępne
Ciśnienie w części
centralnej zbiornika żylnego
Ciśnienie w części
centralnej zbiornika żylnego
CZYNNIKI WARUNKUJĄCE WIELKOŚĆ POJEMNOŚCI MINUTOWEJ
CZYNNIKI WARUNKUJĄCE WIELKOŚĆ POJEMNOŚCI MINUTOWEJ

WSZYSTKIEGO
WSZYSTKIEGO
NAJLEPSZEGO Z OKAZJI
NAJLEPSZEGO Z OKAZJI
ŚWIĄT BOŻEGO
ŚWIĄT BOŻEGO
NARODZENIA
NARODZENIA
I
I
SZCZĘŚLIWEGO NOWEKO
SZCZĘŚLIWEGO NOWEKO
ROKU
ROKU
REGULACJA PRZEPŁYWU
WIEŃCOWEGO
REGULACJA PRZEPŁYWU
WIEŃCOWEGO
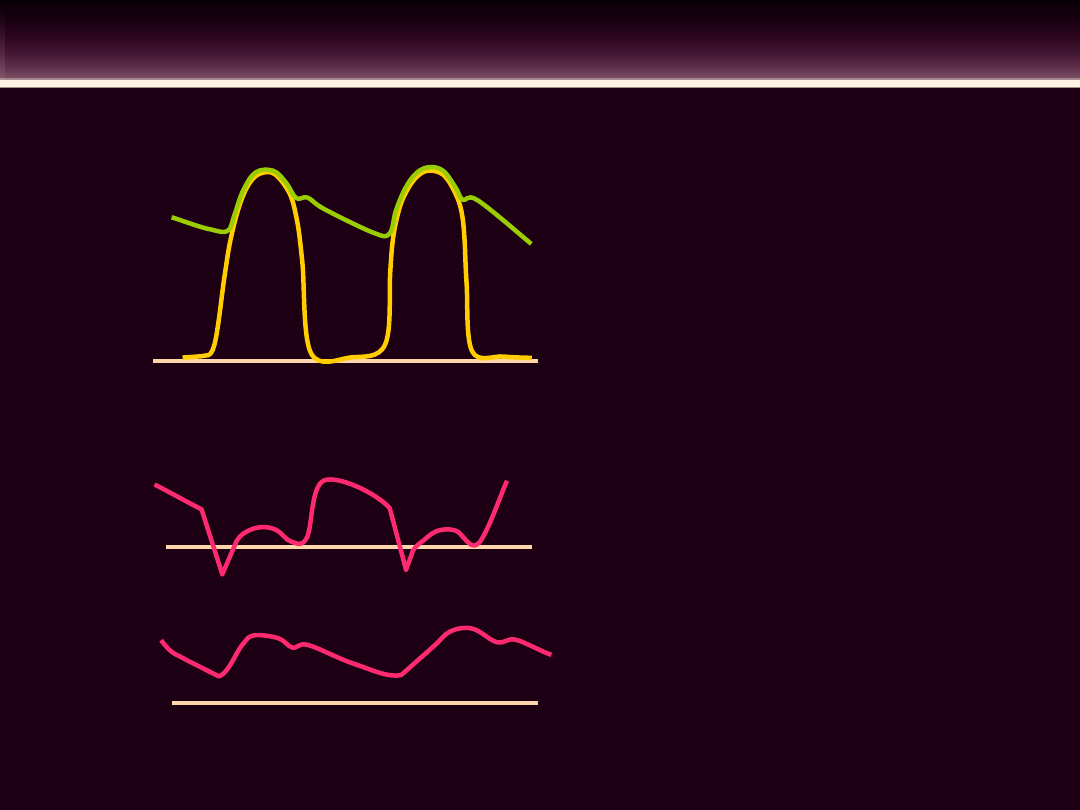
CYKL PRACY SERCA A PRZEPŁYW W NACZYNIACH WIEŃCOWYCH
CYKL PRACY SERCA A PRZEPŁYW W NACZYNIACH WIEŃCOWYCH
Ciśnienie w aorcie
Ciśnienie w aorcie
Ciśnienie w lewej
komorze
Ciśnienie w lewej
komorze
Przepływ krwi w lewej
tętnice wieńcowej
Przepływ krwi w lewej
tętnice wieńcowej
Przepływ
krwi
w
prawej
tętnice
wieńcowej
Przepływ
krwi
w
prawej
tętnice
wieńcowej
• Podczas
skurczu
w
skutek
mocnego ucisku zewnętrznego na
naczynia
wieńcowe
gwałtownie
wzrasta w nich opór przepływu
krwi. Mimo wyższego ciśnienia
krwi w tętnicach wieńco-wych
przepływ przez naczynia wień-
cowe
jest
znacznie
mniejszy
podczas
skurczu
niż
podczas
rozkurczu
• Różnice przepływu krwi przez lewą
a prawą tętnicę wieńcową uwarun-
kowane różną grubością mięśnia
lewej i prawej komory, różnym
ciśnie-niem
generowanym
w
komorach ser-ca podczas skurczu
a za tym różnym stopniem ucisku
na naczynia wieńco-we
• Podczas
rozkurczu
wielkość
przepły-wu krwi przez naczynia
wieńcowe
wzrasta
ponad
zapotrzebowanie
me-taboliczne
(przekrwienie czynnoś-ciowe)
• Podczas
skurczu
w
skutek
mocnego ucisku zewnętrznego na
naczynia
wieńcowe
gwałtownie
wzrasta w nich opór przepływu
krwi. Mimo wyższego ciśnienia
krwi w tętnicach wieńco-wych
przepływ przez naczynia wień-
cowe
jest
znacznie
mniejszy
podczas
skurczu
niż
podczas
rozkurczu
• Różnice przepływu krwi przez lewą
a prawą tętnicę wieńcową uwarun-
kowane różną grubością mięśnia
lewej i prawej komory, różnym
ciśnie-niem
generowanym
w
komorach ser-ca podczas skurczu
a za tym różnym stopniem ucisku
na naczynia wieńco-we
• Podczas
rozkurczu
wielkość
przepły-wu krwi przez naczynia
wieńcowe
wzrasta
ponad
zapotrzebowanie
me-taboliczne
(przekrwienie czynnoś-ciowe)

CYKL PRACY SERCA
CYKL PRACY SERCA
HR =
75
sk/min
HR =
200
sk/min
Czas trwania każdego
cyklu pracy serca
0.80
0.30
Czas trwania skurczu
0.27
0.16
Czas trwania rozkurczu
0.53
0.14
Czas trwania skurczu i rozkurczu mięśnia sercowego w zależności od
częstości skurczów serca (HR)
Czas trwania skurczu i rozkurczu mięśnia sercowego w zależności od
częstości skurczów serca (HR)
•Wzrost częstości skurczów serca powoduje skrócenie czasu
trwania cyklu pracy serca. Znacznie większemu skróceniu
ulega czas trwania rozkurczu. Skraca się okres wypełnienia
komór krwią oraz okres przepływu krwi przez naczynia
wieńcowe
•Krótki czas wypełnienia komór skutkuje spadkiem objętości
wyrzutowej
•Wzrost
częstości
skurczów
serca
powoduje
wzrost
zapotrzebowania mięśnia sercowego na tlen, które przy
znacznym wzroście HR nie może być pokryte
•Wzrost częstości skurczów serca powoduje skrócenie czasu
trwania cyklu pracy serca. Znacznie większemu skróceniu
ulega czas trwania rozkurczu. Skraca się okres wypełnienia
komór krwią oraz okres przepływu krwi przez naczynia
wieńcowe
•Krótki czas wypełnienia komór skutkuje spadkiem objętości
wyrzutowej
•Wzrost
częstości
skurczów
serca
powoduje
wzrost
zapotrzebowania mięśnia sercowego na tlen, które przy
znacznym wzroście HR nie może być pokryte

SPOCZYNKOWY METABOLIZM MIĘŚNIA SERCOWEGO i NIEKTÓRYCH
NARZĄDÓW
SPOCZYNKOWY METABOLIZM MIĘŚNIA SERCOWEGO i NIEKTÓRYCH
NARZĄDÓW
Przepływ krwi
Różnica
tętniczo-
żylna
zawartości
tlenu mL/L
Pochłanianie tlenu
mL/min
(% CO)
mL/
100g na
min
mL/min
(%CO)
mL/100g
/min
Mięsień
sercowy
250
(4.7%
)
84.0
114
29
(11.9
%)
9.7
Mózgowi
e
750
(13.9
%)
54.0
62
46
(18.4
%)
3.3
Skóra
462
(8.6%
)
12.8
25
12
(4.8%
)
0.3
Nerki
1260
(23.3
%)
420.0
14
18
(7.2%
)
6.0
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
- Slide 38
- Slide 39
- Slide 40
- Slide 41
- Slide 42
- Slide 43
- Slide 44
- Slide 45
- Slide 46
- Slide 47
- Slide 48
- Slide 49
- Slide 50
- Slide 51
- Slide 52
- Slide 53
Wyszukiwarka
Podobne podstrony:
Fizjologia krążenia zagadnienia (II kolokwium)
Kolokwium fizjologia krążenie 2009, II rok, fizjologia
FIZJOLOGIA KRAŻENIA KRWI
fizjologia ćwiczenie 2, II rok, II rok CM UMK, Giełdy, 2 rok od Pawła, fizjologia, 6. krazenie
poprawa fizjologia krążenia test
fizjologia krazenie
fizjologia krążenia, Fizjoterapia, fizjologia
fizjologia krazenia krwi, weterynaria, I semestr, Anatomia i fizjologia
Fizjologia krążenia mózgowego i wieńcowego
fizjologia krążenie
Fizjologia krążenie pytania seminarium
fizjologia krążenia
Fizjologia krążenie
Fizjologia krążenia zagadnienia (II kolokwium)
FIZJOLOGIA KRĄŻENIA
ŚCIĄGA Z FIZJOLOGII krążenie c d
więcej podobnych podstron