
1
Oznaczanie aktywności proteolitycznej trypsyny
Cel ćwiczenia
Celem ćwiczenia jest zapoznanie się z metodą oznaczania aktywności endopeptydaz na
przykładzie trypsyny.
Wprowadzenie
Peptydazy (hydrolazy peptydowe EC 3.4.) to enzymy, które rozrywają, przy udziale
cząsteczki wody, wiązania peptydowe łączące aminokwasy w peptydach i białkach. Reakcja
hydrolizy wiązania peptydowego prowadzi do odtworzenia grupy α-karboksylowej jednego, i
grupy α-aminowej drugiego aminokwasu. Peptydazy charakteryzują się niską specyficznością
wobec hydrolizowanego substratu, wykazują natomiast specyficzność wobec położenia
rozrywanego wiązania. Na tej podstawie wyróżnia się endopeptydazy (proteinazy), które
rozszczepiają wiązania peptydowe położone w pewnej odległości od końca łańcucha oraz
egzopeptydazy hydrolizujące wiązania peptydowe położone skrajnie. Egzopeptydazy
działające przy C-końcu peptydu nazywane są karboksypeptydazami, zaś te, które
odszczepiają aminokwas N-końcowy – aminopeptydazami. Znane są także omega peptydazy
rozrywające wiązania peptydowe położone zarówno przy N-, jak i C- końcu peptydu.
Endopeptydazy podzielono na podpodklasy na podstawie mechanizmu katalizy
powiązanego z budową centrum aktywnego tych enzymów na: serynowe (EC 3.4.21),
cysteinowe (EC 3.4.22), aspartylowe (EC 3.4.23), treoninowe (EC 3.4.25), metaloproteinazy
(EC 3.4.24) oraz proteinazy o nieznanym mechanizmie katalizy (EC 3.4.99). Wiedza
dotycząca sekwencji aminokwasowej peptydaz, wtórnej struktury oraz preferencji wobec
miejsca cięcia w substratach jest stale aktualizowana na stronie http://merops.sanger.ac.uk/.
Ze względu na miejsce działania, peptydazy dzieli się na wewnątrzkomórkowe
(lizosomalne/wakuolarne i cytozolowe) oraz pozakomórkowe (peptydazy wydzielane przez
komórki bakteryjne oraz zwierzęce peptydazy trawienne wydzielane do przewodu
pokarmowego). Do grupy peptydaz działających pozakomórkowo należy zaliczyć także
peptydazy syntetyzowane podczas kiełkowania w liścieniach i komórkach warstwy
aleuronowej ziarniaków zbóż, a następnie transportowane do bielma skrobiowego.
Peptydazy wewnątrzkomórkowe są odpowiedzialne przede wszystkim za regulację
wielkości puli białek w komórce, usuwanie białek nieprawidłowych i uszkodzonych, a także
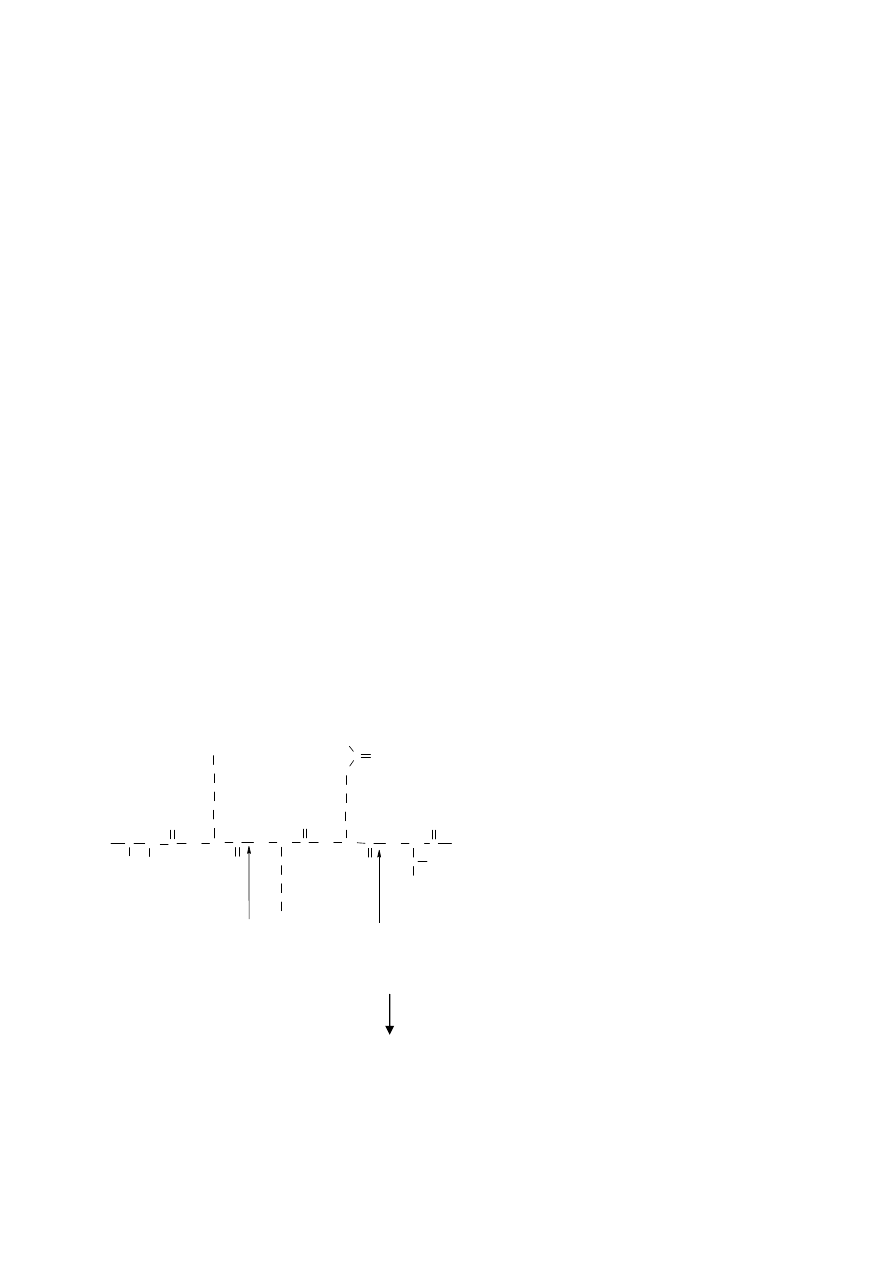
2
za inicjację procesu apoptozy. Natomiast biologiczna rola peptydaz pozakomórkowych
działających w przewodzie pokarmowym zwierząt polega na wstępnej hydrolizie białek
pokarmowych. Do tej grupy enzymów zalicza się: pepsynę, podpuszczkę (syn. chymozyna,
rennina), trypsynę, chymotrypsynę, karboksypeptydazę A i elastazę. Z kolei roślinne
peptydazy syntetyzowane podczas kiełkowania nasion (głównie proteinazy cysteinowe oraz
karboksypeptydazy) hydrolizują białka zapasowe do peptydów i ostatecznie do
aminokwasów, transportowanych następnie do rosnącej siewki.
Endopeptydazy charakteryzują się niską specyficznością substratową, wykazują jednak
preferencje wobec charakteru łańcuchów bocznych aminokwasów tworzących rozkładane wiązanie
peptydowe. Przykładowo chymotrypsyna hydrolizuje wiązania peptydowe powstałe z udziałem grupy
karboksylowej aminokwasów aromatycznych (fenyloalanina, tyrozyna, tryptofan) lub aminokwasów o
dużych, hydrofobowych łańcuchach bocznych np. metionina czy leucyna. Elastaza natomiast
rozszczepia wiązania peptydowe utworzone przez aminokwasy o niewielkich, pozbawionych ładunku
łańcuchach bocznych. Z kolei pepsyna wykazuje aktywność wobec wiązań peptydowych utworzonych
z udziałem grupy aminowej aminokwasów aromatycznych i kwaśnych oraz między waliną i leucyną
(Bańkowski 2006). Peptydazy trawienne syntetyzowane są w postaci nieaktywnych prekursorów
ulegających aktywacji proteolitycznej najczęściej inicjowanej przez trypsynę.
Trypsyna
(EC 3.4.21.4). Enzym ten, podobnie jak chymotrypsyna i elastaza, jest
zaliczany do podpodklasy proteinaz serynowych, których centrum aktywne tworzą łańcuchy
boczne seryny, kwasu asparaginowego i histydyny. Trypsyna katalizuje hydrolizę wiązań
peptydowych, w których grupy karbonylowe należą do argininy lub lizyny (rys. 1A).
C
O
CH
NH
C
H
OH
CH
3
C
O
N
H
3
+
C
CH
NH
CH
2
CH
2
CH
2
N
H
NH
C
O
CH
NH
CH
2
CH
2
S
CH
3
C
O
CH
NH
3
+
NH
CH
2
CH
2
CH
2
CH
2
C
O
CH
N
CH
2
OH
H
Thr
Ser
Lys
Arg
er
trypsyna
trypsyna
Met
Ser – Ala – Asp – Gly – Lys – Met – Trp – Gly – Arg – Gly – Ala – Lys – Cys – Gly
Ser – Ala – Asp – Gly – Lys
Met – Trp – Gly – Arg
Gly – Ala – Lys – Cys – Gly
Rys. 1 Działanie trypsyny na białko: A) wiązania peptydowe hydrolizowane przez
trypsynę, B) uwalnianie peptydów o różnej długości.
A)
B)
trypsyna

3
Produktem działania trypsyny na białko są peptydy o różnej długości łańcucha z resztą
lizyny lub argininy na C-końcu (Rys. 1B).
Ze względu na specyficzność działania, enzym ten jest stosowany do rozszczepiania
białek na peptydy przed ich sekwencjonowaniem metodą Edmana – metoda ta pozwala na
poprawne ustalenie sekwencji peptydów zbudowanych maksymalnie z 50 aminokwasów.
Trypsyna jest syntetyzowana przez komórki trzustki w postaci nieaktywnego
prekursora (proenzymu) – trypsynogenu (pojedynczy łańcuch polipetydowy zawierający 247
reszt aminokwasowych), który ulega w jelicie cienkim aktywacji pod wpływem enterokinazy
jelitowej
(syn.
enteropeptydaza).
Proces
aktywacji
polega
na
odłączeniu
sześcioaminokwasowego odcinka prekursorowego, co powoduje zmiany konformacyjne
prowadzące do uzyskania przez cząsteczkę właściwości katalitycznych. Zaktywowane w ten
sposób cząsteczki trypsyny aktywują kolejne cząsteczki trypsynogenu oraz prekursory innych
peptydaz wydzielanych przez komórki trzustki do światła jelita cienkiego. Optimum pH
działania trypsyny zawiera się w granicach 7–9.
Metody oznaczania aktywności endopeptydaz
bazują na fotometrycznym pomiarze
ilości peptydów (produktów reakcji) uwolnionych z substratu białkowego przez enzym.
Pomiaru tego dokonuje się po usunięciu, przez wirowanie lub sączenie, z mieszaniny
poreakcyjnej pozostałości substratu:
bezpośrednio:
- przy 280 nm - pomiar pochłaniania promieniowania elektromagnetycznego przez
wiązania peptydowe oraz pierścienie aromatyczne łańcuchów bocznych aminokwasów
- przy 340 nm dla produktów hydrolizy barwnych białek azowanych (białka poddane
reakcji sprzęgania z diazowanym kwasem sulfanilowym podstawianym do reszt histydyny i
tyrozyny)
pośrednio, po przeprowadzeniu reakcji zachodzącej pomiędzy łańcuchami bocznymi
aminokwasów zawartych w uwolnionych peptydach a odczynnikami dającymi barwny
produkt.
Zasada
metody Ansona. Metoda ta polega na pomiarze ilości tyrozyny i tryptofanu
zawartych w peptydach uwolnionych przez enzym z substratu (kazeina). Peptydy te, w
odróżnieniu od białek, są rozpuszczalne w 3-proc. kwasie trichlorooctowym. Wytrącone
kwasem białka usuwa się z mieszaniny przez wirowanie lub sączenie.
Enzymatyczną hydrolizę białka (substratu) prowadzi się w pH 8,6 i temp. 37°C.
Oznaczenie zawartości tyrozyny i tryptofanu w uwolnionych przez enzym peptydach

4
wykonuje się przy użyciu odczynnika Folina. Końcowa barwa, której natężenie jest wprost
proporcjonalne do ilości tyrozyny i tryptofanu w próbie, jest wynikiem redukcji odczynnika
fosforomolibdenofosforowolframowego (odczynnik Folina) w środowisku zasadowym do
błękitu fosforomolibdenowego, zachodzącej pod wpływem wymienionych aminokwasów.
Stężenie barwnego produktu w próbie oznacza się mierząc pochłanianie (absorbancję)
promieniowania świetlnego o długości fali 670 nm.
Miarą aktywności trypsyny będzie ilość µmoli tyrozyny zawarta w uwolnionych
peptydach.
Odczynniki
1.
0,1-molowy bufor Tris–HCl o pH 8,6 (500 ml 0,2-molowego roztworu Tris-
(hydroksymetylo)aminometanu doprowadzić do pH 8,6 za pomocą 1-molowego kwasu
solnego i uzupełnić wodą destylowaną do 1 litra)
2.
1% (w/v) roztwór kazeiny w 0,1 molowym buforze Tris-HCl o pH 8,6 (rozpuszczać na
mieszadle magnetycznym w czasie 45 minut, zmierzyć pH)
3.
Odczynnik Folina (Ciocalteu) rozcieńczony czterokrotnie wodą.
4.
0,5-molowy wodorotlenek sodowy.
5.
7,5-proc. roztwór kwasu trichlorooctowego (przechowywać w lodówce).
Wykonanie
Oznaczenie
1.
Hydroliza enzymatyczna kazeiny.
Otrzymany w kolbie miarowej na 10 ml roztwór trypsyny dopełnić do kreski 0,1-
molowym buforem Tris-HCl (1) i dobrze wymieszać. Do trzech enzymatycznie czystych
probówek (dwie próby pełne i jedna kontrolna) odmierzyć po 2 ml roztworu kazeiny (2) i
umieścić je w łaźni wodnej w temp. 37°C. Po około 5 minutach do dwóch probówek z
kazeiną (próby pełne) dodać po 1 ml enzymu z kolby miarowej na 10 ml, szybko
wymieszać i inkubować wszystkie trzy próby w temp. 37°C dokładnie przez 15 minut.
Następnie do wszystkich trzech prób dodać po 2 ml kwasu trichlorooctowego (5), wyjąć
probówki z łaźni wodnej i do próby kontrolnej dodać 1 ml enzymu z kolby miarowej na 10

5
ml. Wszystkie trzy próby wymieszać i pozostawić przez 30 minut w temperaturze
pokojowej w celu całkowitego wytrącenia osadu białka. Próby przesączyć do czystych
probówek przez sączki bibułowe.
2.
Fotometryczne oznaczanie tyrozyny w produktach hydrolizy enzymatycznej
Do kolejnych trzech czystych probówek odmierzyć po 1 ml klarownego przesączu z
dwóch prób pełnych i z próby kontrolnej. Do wszystkich trzech probówek dodać po 2 ml
0,5-molowego wodorotlenku sodowego (4) oraz po 0,6 ml odczynnika Folina (3) -
zawartość probówek dobrze wymieszać. Po 10 minutach odczytać absorbancję prób pełnych
w fotometrze przy długości fali 670 nm ustawionym na zero wobec próby kontrolnej.
Obliczenie aktywności trypsyny
1.
Obliczyć średnią wartości absorbancji prób pełnych.
2.
Posługując się obliczoną różnicą absorbancji odczytać z krzywej wzorcowej ilość µmoli
tyrozyny zawartej w 1 ml przesączu.
3.
Obliczyć aktywność enzymu w µmolach tyrozyny zawartej w peptydach uwolnionych w
warunkach metody w ciągu 1 min przez 1 mg trypsyny [µmole tyrozyny × min.
-1
×
mg
trypsyny
-1
] uwzględniając:
a.
schemat rozcieńczeń trypsyny podany przez prowadzącego zajęcia
b.
czas trwania enzymatycznej hydrolizy substratu .
Przykładowe obliczenia:
Absorbancja
próby pełnej
nr 1
Absorbancja
próby pełnej
nr 2
Ś
rednia
absorbancja
Ilość µmoli tyrozyny odczytana z krzywej
wzorcowej dla średniej absorbancji
0,182
0,188
0,185
0,2
Ś
rednia absorbancja prób pełnych wynosi 0,185, czyli w 1 ml przesączu znajduje się 0,2
µmola tyrozyny.
Ponieważ do reakcji barwnej pobrano 1 ml z 5 ml przesączu, ilość tyrozyny zawartej w
peptydach uwolnionych z substratu przez 1 ml trypsyny wynosi 1 µmol. Aktywność trypsyny,
zawartej w 10 ml roztworu, jest więc równa 10 µmolom tyrozyny. Wiedząc, że przed
uzupełnieniem do objętości 10 ml kolbka zawierała 2 ml roztworu trypsyny przygotowanego

6
przez rozpuszczenie 7 mg tego enzymu w 40 ml buforu możemy obliczyć aktywność trypsyny
w następujący sposób:
•
aktywność 2 ml wyjściowego roztworu trypsyny wynosi 10 µmoli tyrozyny.
•
aktywność 7 mg trypsyny jest równa 200 µmoli tyrozyny
•
aktywność 1 mg trypsyny wynosi 28,57 µmoli tyrozyny, a po uwzględnieniu czasu
trwania reakcji enzymatycznej wynoszącego15 minut, aktywność ta będzie równa 1,91
µmola tyrozyny × min.
-1
× mg trypsyny
-1
.
Pytania
1.
Jakie kryteria podziału stosuje się wobec peptydaz?
2.
Czym różnią się karboksypeptydazy od aminopeptydaz? Przedstaw wzór dowolnego
tripeptydu i wskaż wiązanie peptydowe, które może rozerwać aminopeptydaza.
3.
Wymień peptydazy działające w przewodzie pokarmowym i krótko je scharakteryzuj.
4.
Podaj miejsce syntezy trypsynogenu i omów jego aktywację.
5.
Jaką rolę w aktywacji trypsynogenu pełni enterokinaza?
6.
Jakie praktyczne zastosowanie znalazła trypsyna w badaniach biochemicznych?
7.
Omów metodę oznaczania aktywności trypsyny na ćwiczeniu. Jaką role spełnia w tej
metodzie kwas trichlorooctowy (TCA)?
8.
Przedstaw, używając trzyliterowych skrótów nazw aminokwasów, fragment łańcucha
polipeptydowego i wskaż miejsce działania trypsyny.
Literatura
1.
Anson, M.L. (1938) J. Gen. Physiol. 22, 79-89.
2.
Chen J-M., Kurko Z., Le Marechal C., Toth C., Tsakiris L., Raguenes O., Ferec C.,
Sahin-Toth M. (2003) Mol. Biol. Evol. 20(11), 1767-1777
3.
Bańkowski E., Biochemia, MedPharm Polska, Wrocław 2006
Wyszukiwarka
Podobne podstrony:
11) Oznaczanie aktywności proteolitycznej trypsyny
Oznaczanie aktywności proteolitycznej trypsyny
9) Oznaczanie aktywności enzymów amylolitycznych
Oznaczanie aktywności enzymów lipolitycznych
Kraking katalityczny – oznaczanie aktywności katalizatorów metodą UOP
Biochemia, Oznaczanie aktywnościi amylazy metodą Noeltinga i Bernfelda w ziarnie pszenicy
cwiczenie 5 amylaza oznaczanie aktywnosci enzymu metoda kolorymetryczna 05 05 2014
Spektrofotometryczna metoda oznaczania aktywności peroksydazy
Ćwiczenie 6 Oznaczanie aktywności katalazy
IG.6 - Oznaczanie aktywności enzymatycznej metaloproteinaz komórkowych, Genetyka, Inżynieria genetyc
Ćwiczenie 6 Oznaczanie aktywności katalazy
oznaczanie aktywności enzymów, Biotechnologia, laborki
Oznaczanie aktywności katalazy i wyznaczanie szybkości początkowej reakcji enzymatycznejx
Oznaczenie aktywnosci czastek p Nieznany
Oznaczanie aktywności lipazy trzustkowej
Aktywność inhibitora trypsyny w plazmie nasienia lisa polarnego w sezonie rozrodczym
więcej podobnych podstron