165. Jaki zjawiska cyklu komórkowego zapewniają, że przechodzenie komórek przez stadia cyklu jest jednokierunkowe i nieodwracalne. Jakie mechanizmy leżą u podstaw tej strategii?
Przejście przez kolejne fazy cyklu kontrolują kinazy cyklinozależne (CDK). Enzymy te łączą się z odpowiednimi cyklinami (stają się wtedy aktywne) fosforylując białka, w tym czynniki transkrypcyjne, co prowadzi do zmiany ekspresji genów dając w rezultacie przejście do następnej fazy cyklu komórkowego. Fosforyluja również inhibitory kinaz cyklinozależnych charakterystycznych dla następnych faz, co powoduje degradację tych inhibitorów. Z kolei kinazy faz następnych promują degradację swoich poprzedniczek. Czyli mamy powiedzmy kompleksy cyklina/CDK z fazy G1 i cyklina/CDK z S, ale ta druga jest hamowana. G1 powodują degradacje inhibitorów S, a S degradację cyklin/CDK G1. Czynniki powodujące pojawienie się nowych cyklin G1 czy inhibitorów aktywne są tylko na początku G1, więc cykl nie może się cofnąć, bo proteolizy nie da się odwrócić.
166. Właściwości komórek znajdujących się w cyklu podziałowym.
Komórki znajdujące się w cyklu podziałowym, w przeciwieństwie do komórek w spoczynku, ciągle przechodzą kolejno przez fazy G1, S, G2 i M cyklu. Charakterystyczne dla nich jest znaczny wzrost syntezy białek potrzebnych do replikacji DNA w fazie G1, podwojenie DNA w komórce w fazie S, synteza białek potrzebnych do samej mitozy w fazie G2 i w końcu podział w fazie M. Charakterystyczne są też cykliczne zmiany w poziomach enzymów kontrolujących cykl komórkowy - kinaz cyklinozależnych oraz ich aktywatorów - cyklin. We wczesnej fazie G1 rośnie poziom cyklin G1, które wraz ze swoimi CDK (cyclin dependent kinases) inicjują ekspresję genów potrzebnych do wzrostu komórki i replikacji DNA. W tej fazie pojawiają się też cykliny S, ale są one inhibowane. Dopiero gdy cykliny G1 się zakumulują, ich CDK fosforylują inhibitory cyklin S degradując je. Wtedy cykliny i CDK S powodują przejście do fazy S i replikację na powstałych wcześniej kompleksach. Jednocześnie hamują powstawanie nowych kompleksów replikacyjnych, dzięki czemu chromosomy podwajane są tylko raz na podział. Akumulacja cyklin i CDK 2 powoduje uaktywnienie cyklin i CDK G2, które aktywują geny potrzebne do mitozy i samą mitozę. Przy końcu mitozy uaktywnia się APC - anaphase promoting complex - który rozkłada mitotyczne cykliny i powoduje syntezę cyklin G1.
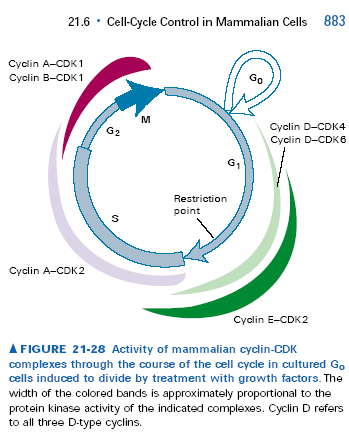
167. Właściwości komórek w fazie G1/G0.
W fazie G1 komórki na skutek działania kompleksów cyklina D-CDK4/6 oraz cyklina E-CDK2 intensywnie syntetyzują białka i rosną. Białka te potrzebne są do późniejszej replikacji DNA w fazie S. Jednak komórki ssacze wymagają czynników wzrostowych i mitogenów aby przejść cykl komórkowy. Jeśli po zakończonej mitozie nie będzie aktywności mitogenów z zewnątrz komórki, to nie będzie aktywności wspomnianych cyklin/CDK. Komórki wchodzą wtedy w fazę G0, czyli opuszczają cykl komórkowy i przestają się dzielić. Wyjście takie może być nieodwracalne lub odwracalne mitogenami. Komórki w fazie G0 nie syntetyzują DNA, a synteza białek i wzrost są zwykle słabsze niż w komórkach dzielących się, a także bardziej wyspecjalizowane, gdyż komórki w fazie G0 są zwykle zróżnicowane, tzn. przystosowane do pełnienia konkretnych funkcji. Przykładem komórek w permanentnej fazie G0 są np. neurony i miotuby. Przejściowo w fazę G0 wchodzą np. nieaktywne fibroblasty.
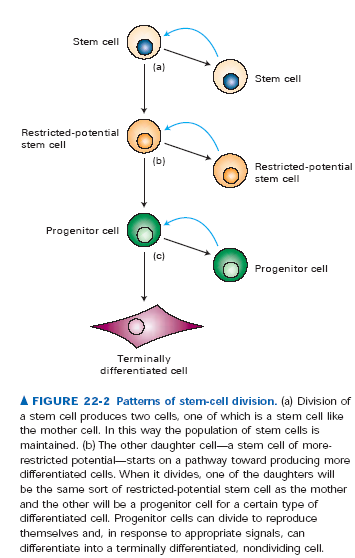
168. Komórki macierzyste.
Komórki macierzyste to niezróżnicowane komórki zachowujące potencjał podziałowy i różnicowania teoretycznie w nieskończoność. Cechą, która odróżnia je od wszystkich innych to zdolność samoodnawiania. Zawsze przynajmniej jedna komórka po podziale komórki macierzystej będzie z nią identyczna. Występują w zarodkach a także w dorosłych organizmach np. w szpiku kostnym. Te drugie, czyli tzw. somatyczne komórki macierzyste służą odbudowywaniu zużytych i zniszczonych tkanek. Komórki macierzyste mogą dzielić się symetryczni - powstają wtedy dwie identyczne komórki macierzyste. Mogą też dzielić się asymetrycznie - powstaje wtedy jedna komórka macierzysta i jedna bardziej wyspecjalizowana i z mniejszym potencjałem proliferacyjnym. Komorki macierzyste mogą być totipotencjalne, czyli mieć potencjał zmiany we wszystkie typy komórek organizmu (prawdziwie totipotencjalne są tylko komórki linii zarodkowej i pierwsze 8 komórek zarodka), pluripotencjalne, czyli mieć możliwość przejścia w kilka typów komórek (np. niektóre komórki szpiku we wszystkie rodzaje krwinek). Mogą być wreszcie unipotencjalne, czyli wprawdzie mogą się dzielić w nieskończoność, ale z ich specjalizacji powstaje tylko jeden typ komórek.
Komorki progenitorowe to wstępnie zróżnicowane komórki macierzyste które mają przejść różnicowanie a nie odtwarzać kolejne pokolenia. Nie potrafią się same odnawiać, ich potomstwo jest zawsze inne od nich samych.
Jeszcze tylko przykłady i będzie to i następne zagadnienie zrobione:
-totipotencjalne: pierwszych 8 komórek zarodka, komórki linii zarodkowej, tzn te, które dają jaja/plemniki (są totipotencjalne, ale teoretycznie nie są komórkami macierzystymi, bo nie odnawiają same siebie)
-pluripotencjalne: komórki zarodka po fazie 8 komórek, komórki macierzyste szpiku kostnego.
-unipotencjalne: komórki macierzyste naskórka i nabłonka jelita cienkiego.
169. Zrobione razem z 168.
Wyszukiwarka
Podobne podstrony:
(165 169) 10 Retoryka A Pedagogika
Ustawa z dnia 25 06 1999 r o świadcz pien z ubezp społ w razie choroby i macierz
macierz BCG
macierze 2
04 Analiza kinematyczna manipulatorów robotów metodą macierz
macierze i wyznaczniki lista nr Nieznany
macierze 1
Macierz przykrycia testów akceptacyjnych Jasiek
MACIERZE
macierze moje i rzad id 275988 Nieznany
ćw 15 Rachunek macierzowy
Lab Wypełnianie macierzy dendro meteo
165 167 607 pol ed01 2007
Potencjał węglowodorowy skał macierzystych i geneza gazu zie, geologia, AGH, SzM, GEOLOGIA
Macierze i wyznaczniki, Politechnika Poznańska, Elektrotechnika, Matematyka, semestr 2
Laboratorium 2 Macierze
Opis macierzy
Zadania macierze
więcej podobnych podstron