DSC00655
nych kompleksów w tworzeniu mikrofibryl (rys. 2531 Terminalne kompleksy pow*wją * * okresie inicjowania (etany komórkowej, w wyniku leczenia w raedy pojedyncaycb narensh obecnych na plazmolemie. Takie kompleksy ustawiają sic równolcgk względem nów > * *"•« tworzenia mi kro fi brył przesuwają sic ruchem śrubowym ku biegunom komórki Po osiągów** bieguna zmieniają one kierunek na prostopadły do poprzedniego i rozpoczyna sic tworze*** nowych mikrofibryl (etany
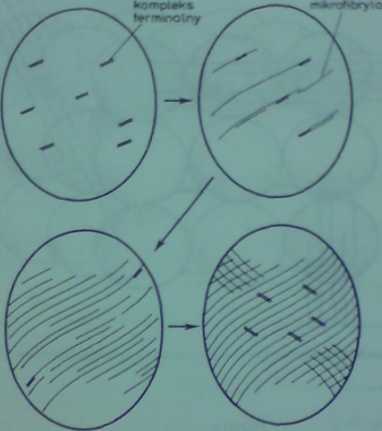
Rys, 25.5, Odkładanie mikrofibryl celulozowych na powierzchni jednokomórkowego glonu Oocystli t udzialetn pleksów terminalnych (wg Montezinosa i Browna, 1976). Aby zachować przejrzystość rysunku pominlcto tnllyunbłyls oraz kompleksy terminalne na biegunach komórek
Zgodny kierunek przebiegu mikrotubul i mikrofibryl celulozowych w okresie tworze ols i rozbudowy ściany wskazuje na udział tych struktur w depozycji mikrofibryli Istnieje dobra udokumentowany pogląd, że mikrotubule przyścienne łącząc się poprzez plazmolemę / kompl#* ksem syntazy celulozy wpływają na kierunek, w którym kompleksy syntazy celulozy przesuwni się w płaszczyźnie plazmolemy, w czasie tworzenia mikrofibryl.
Mikrofibryle celulozowe wydłużają sic drogą wzrostu szczytowego z jednej lub dwu stroft
Do wyjątków należą rośliny, u których celuloza nie występuje w ścianach komórkowy® U niektórych glonów morskich (Bryopsis, Caulerpa) materiałem szkieletowym ściany są ksyUay połączone wiązaniami f) (1—»3). Występują one w postaci mikrofibryl o kształcie spirali ukrfi*1 nych w sposób podobny jak celuloza.
W skład ścian komórek grzybów z wyjątkiem Oomycetes, których ściany zbudowane M z celulozy, wchodzi chityna. Składa sic ona z reszt acctyloglukozoaminy połączonych wiązanhdjf
Wyszukiwarka
Podobne podstrony:
(-Tworzenie Usług Rys. 3. Tworzenie usług - Pięć aspektów
IMG2 TworzenieiMOftaif 71 Rys. 1.7. Powstawanie wiązań siania się
5WW5b pcb ukł form ssb + 4v +12v(kluczo#.) przy SSB mikrofon Ry
100 rtys 71. Zasada zgrzewania garbowego: a, b, c - poszczególne elapy tworzenia zgrzeiny Rys, 72. P
Wybór sekwencji i zakończenie wyboru (alternatywa) Podczas tworzenia alternatywy (rys. 16): należy
Symulacja komputerowa mechanizmu tworzenia się rys w dźwigarze 100 Punkt pomiarowy - U2 Badani a lab
Symulacja komputerowa mechanizmu tworzenia się rys w dźwigarze 101 Sfc (kN) 0
Symulacja komputerowa mechanizmu tworzenia się rys w dźwigarze 78 S = ^Si, gdzie >7 -> co,
Symulacja komputerowa mechanizmu tworzenia się rys w dźwigarze 84 powierzchni i co z tym się wiąże b
Symulacja komputerowa mechanizmu tworzenia się rys w dźwigarze 85 Za pracą Majewskiego [2003] wartoś
Symulacja komputerowa mechanizmu tworzenia się rys w dźwigarze 90 podczas rozciągania nie jest trakt
Symulacja komputerowa mechanizmu tworzenia się rys w dźwigarze 99 zespolonych. Obraz uszkodzeń otrzy
DSC00691 nPMUV*-Mir tvf rcK Rys. 26.18. Mechanizmy witania CO2 u roślin typu O (rośliny lypu CU mogą
DSC00698 nych małe wakuole (prowakuole) biorą swój początek z siateczki śródplazmatycznej. Mogą one
DSC00699 (7) no£ć jest z kolei przekładana na terminy teoratyjfb ne. Dla socjologicznego zrozumienia
10572 IMG 94 bmp Tłumik Dl 20 dBl+HH (podwójny) ■o § Generat mikrofalo Rys. 1. Schemat stanowiska do
Rys historyczny Termin: „ZIELONA CHEMIA” został użyty po raz pierwszy przez P.T. Anastasa w powołany
więcej podobnych podstron