Katarzyna Spytkowska
Paweł Popecki
Układ bodźco-
przewodzący serca.
Przebieg
potencjałów
czynnościow
ych i
kierunki
prądów
jonowych
charakteryst
ycz-
nych dla
komórek
układu
bodźco-
przewodzące
go i
roboczych
serca
.

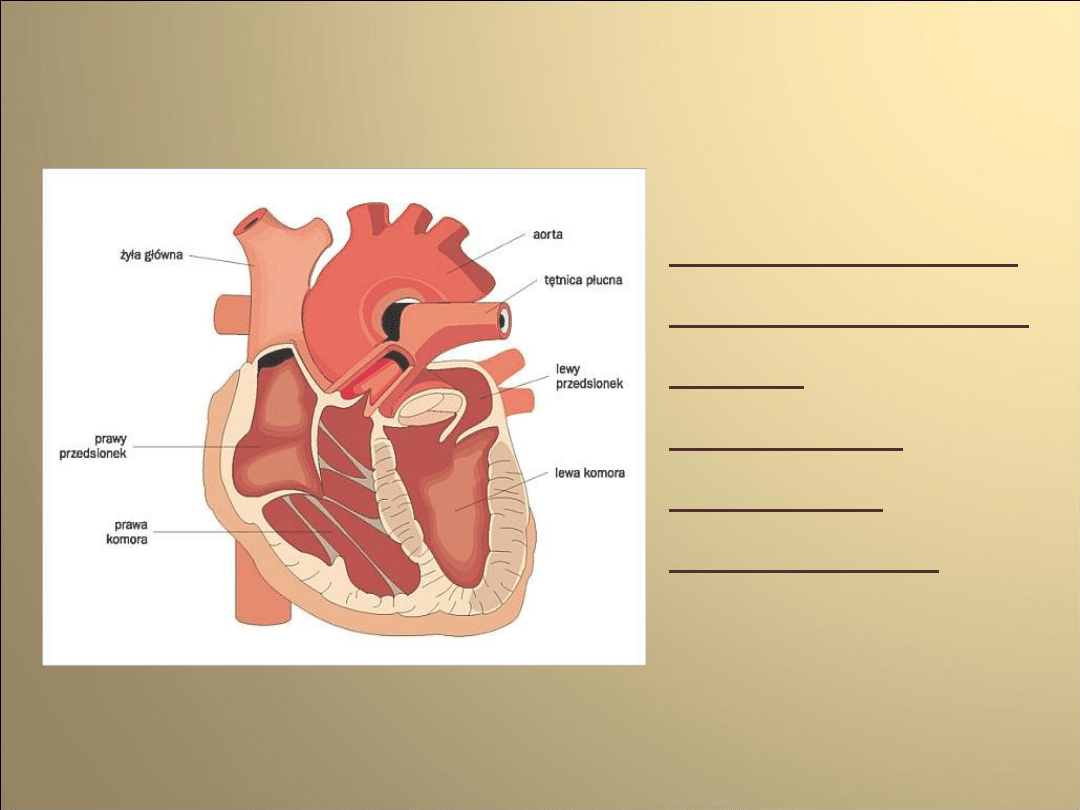
Układ bodźco-przewodzący
serca
• Zapewnia odpowiedni rytm i synchronizację skurczów
poszczególnych części serca.
• Składa się z komórek mięśniowych wyspecjalizowanych
do przewodzenia bodźców i odpowiednio
komunikujących się ze sobą
(za pomocą połączeń tworzonych przez koneksony,
nie powodujących praktycznie żadnego opóźnienia
przenoszonego impulsu)
• Cechą charakterystyczną komórek tego układu są
autonomiczność i automatyzm

Budowa układu bodźcowo-
przewodzącego serca
Komórki mięśniowe nazywane węzłami i
pęczkami:
• węzeł zatokowo- przedsionkowy
• węzeł przedsionkowo- komorowy
• pęczek przedsionkowo-
komorowy(Hisa)

Węzeł zatokowo-
przedsionkowy
• Położony w pobliżu ujścia żyły głównej
górnej do prawego przedsionka
• Generowane są w nim impulsy
elektryczne decydujące o częstotliwości
skurczów serca
• Powstające w węźle potencjały
czynnościowe(około 78 impulsów na
minutę) inicjują skurcz przedsionków i
za pośrednictwem pęczków
międzywęzłowych docierają do węzła
przedsionkowo- komorowego

Węzeł przedsionkowo-
komorowy
• Położony w dolnej części przegrody
międzyprzedsionkowej
• Posiada małą prędkość rozchodzenia się
potencjału czynnościowego, co wynika z
małej prędkości narastania potencjału
czynnościowego w komórkach
• Komórki są zdolne do samoistnego
generowania potencjałów czynnościowych
jednakże częstotliwość generowanych
impulsów jest mniejsza niż w węźle
zatokowo-przedsionkowym

Pęczek przedsionkowo-
komorowy (Hisa)
• Wychodzi z węzła przedsionkowo-komorowego w
postaci jednego pnia i następnie rozdziela się na
dwie odnogi (lewą i prawą)
Odnogi biegną w kierunku koniuszka i w jego
pobliżu zakręcają w stronę podstawy serca
• Pęczek przedsionkowo-komorowy zbudowany
jest z komórek przewodzących nazwanych
włóknami Purkinjego.
• Pęczek ma zdolność do samoistnego generowania
potencjałów czynnościowych o częstotliwości 30/min
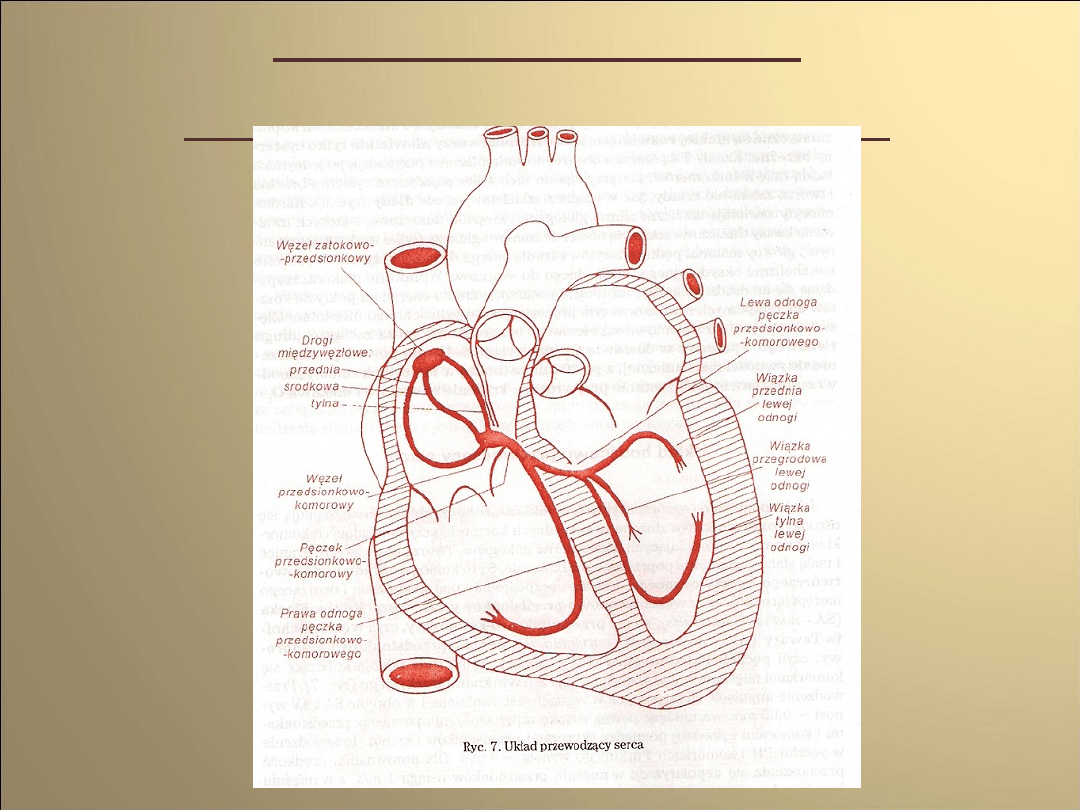
Układ bodźco-
przewodzący serca

Samoistna generacja
potencjału czynnościowego
• W komórkach układu przewodzącego serca
zachodzi zjawisko powolnej spoczynkowej
depolaryzacji. Dzięki temu komórki te mają
zdolność do samoistnego, periodycznego
generowania potencjału czynnościowego
• Najważniejszymi komórkami generującymi
impulsy są komórki P węzła zatokowo-
przedsionkowego.
(Zdolność tę mają również inne komórki układu
przewodzącego, jednakże z uwagi na dużo wolniejsze
tempo zachodzenia tego procesu nie mają tak
istotnego znaczenia-są pobudzane przez komórki P)
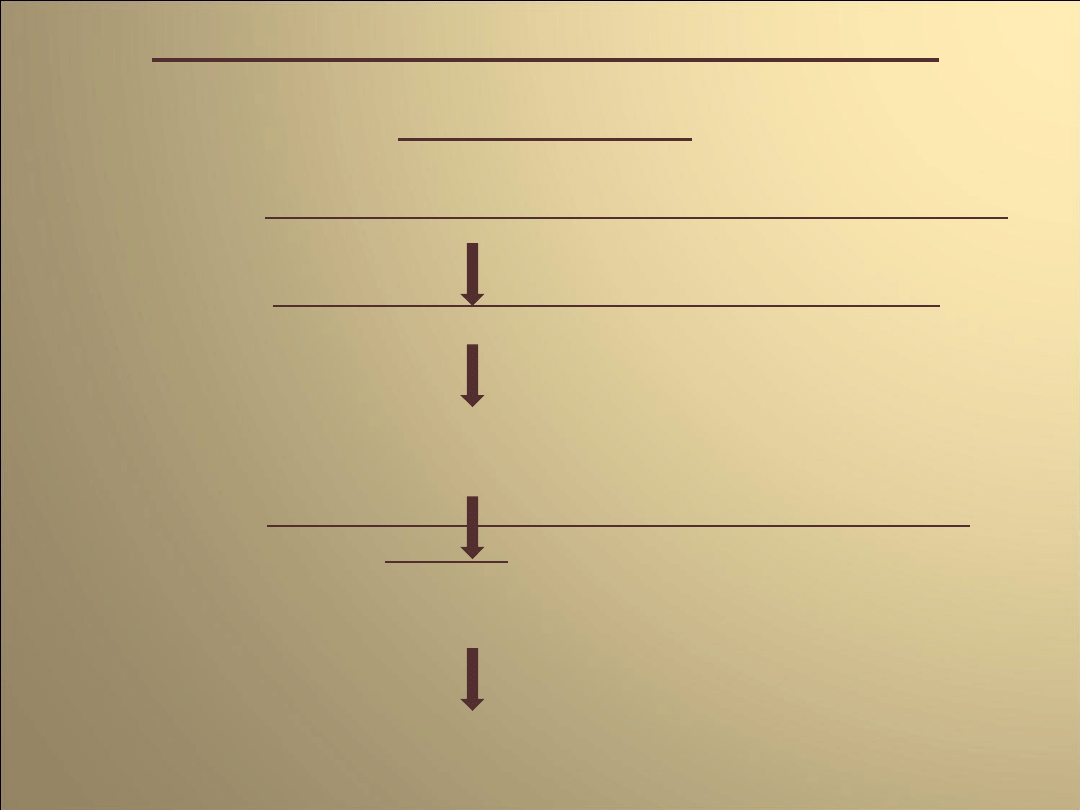
Mechanizm generacji
impulsu
Repolaryzacja poprzedniego potencjału czynnościowego,
ok. -75mV(opóźnione prostownicze kanały potasowe)
otwarcie nieselektywnych kanałów kationowych
powolna depolaryzacja błony
dezaktywacja kanałów potasowych
aktywacja kanałów wapniowych typu T oraz później
także L, ok. -50mV
generacja
potencjału czynnościowego

• Faza depolaryzacji w komórkach
rozrusznikowych jest znacznie
wolniejsza, bowiem kanały wapniowe
(typu T, a zwłaszcza typu L) otwierają się
znacznie wolniej od kanałów sodowych.
• Długi średni czas otwarcia kanałów
wapniowych typu L oraz bardzo powolna
aktywacja kanałów potasowych (około
tysiąc razy wolniejsza niż w komórkach
nerwowych) są odpowiedzialne za czas
trwania fazy repolaryzacji.

Potencjał czynnościowy komórek
układu przewodzącego serca
powstaje dzięki przepływowi
prądu wapniowego
(w odróżnieniu od komórek
nerwowych, gdzie największą
rolę pełni prąd sodowy)
Działanie komórek, w których występuje
wapniowy mechanizm depolaryzacji w potencjale
czynnościowym, może być modulowane przez
niektóre leki oraz trucizny
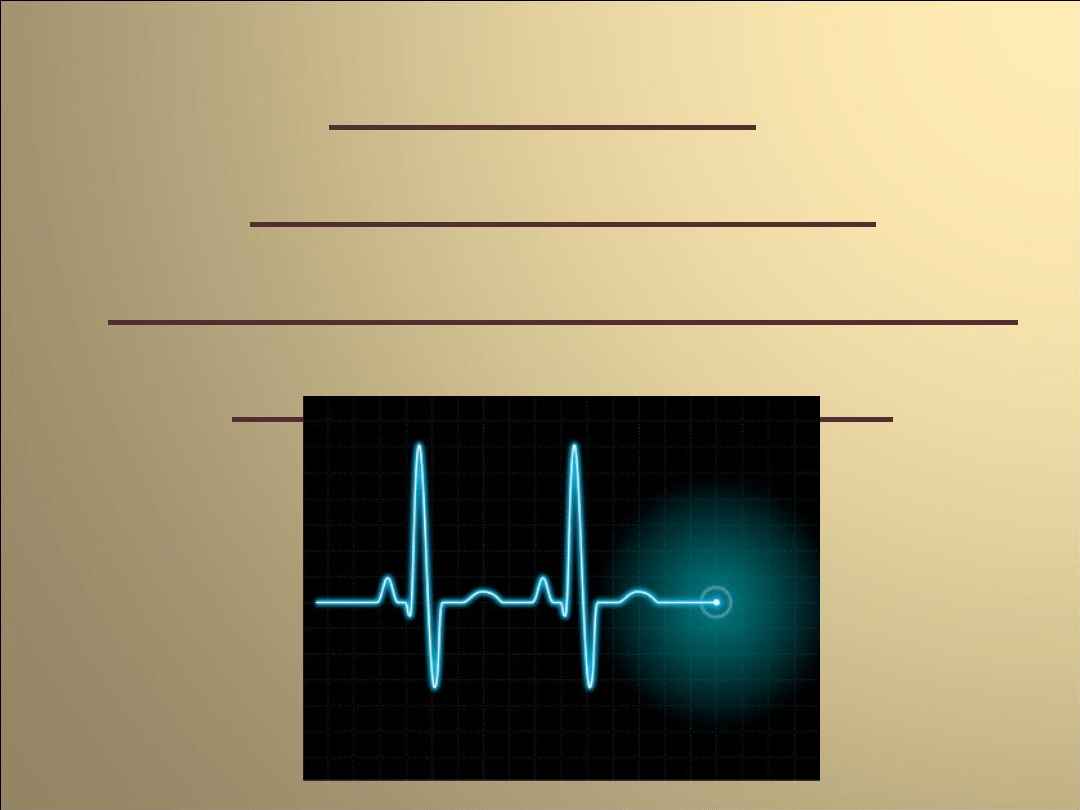
Potencjał
czynnościowy
mięśniowej komórki
roboczej serca
MIĘSIEŃ POPRZECZNIE PRĄŻKOWANY SERCOWY
•jest pobudliwy
• kurczy się rytmicznie i automatycznie
• wykonuje pracę bez przerwy, przez całe życie
• wypoczywa tylko między kolejnymi skurczami
• jest wrażliwy na niedotlenienie i niedokrwienie

Potencjał czynnościowy mięśniowy
komórki roboczej serca
różni się istotnie od potencjału
czynnościowego neuronu lub komórki mięśnia
szkieletowego.
W jego przebiegu wyróżnia sie kilka faz które
związane są z działaniem wielu typów kanałów
jonowych
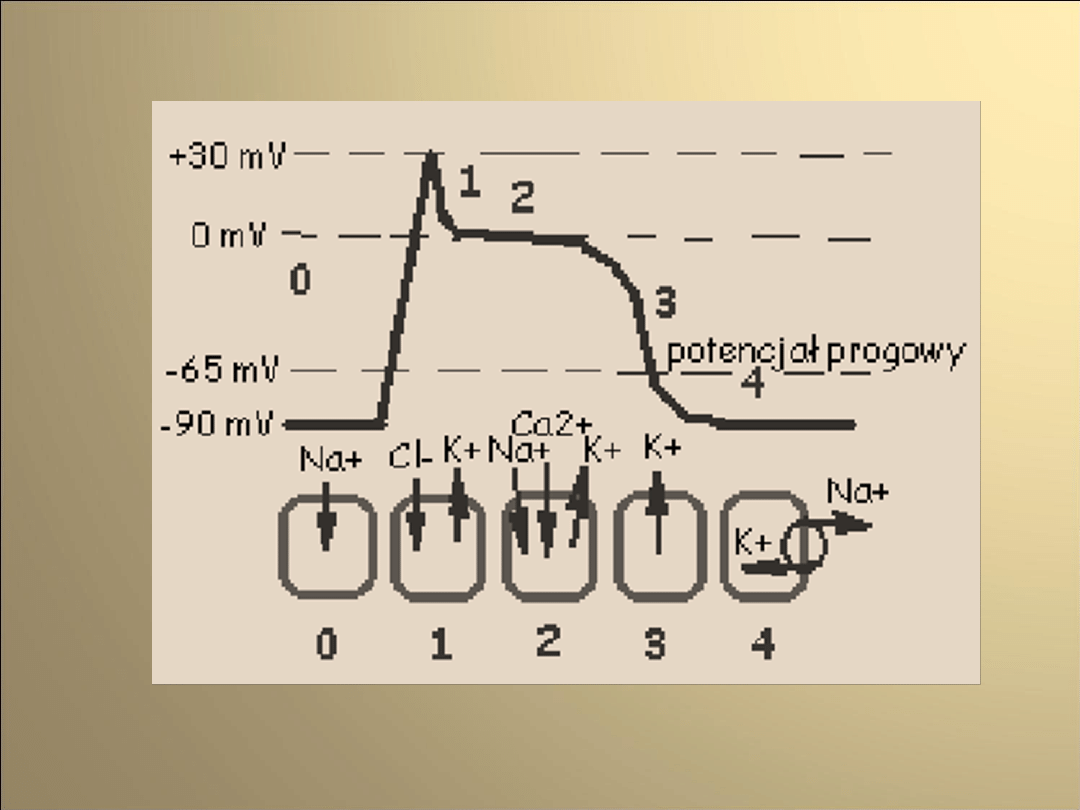
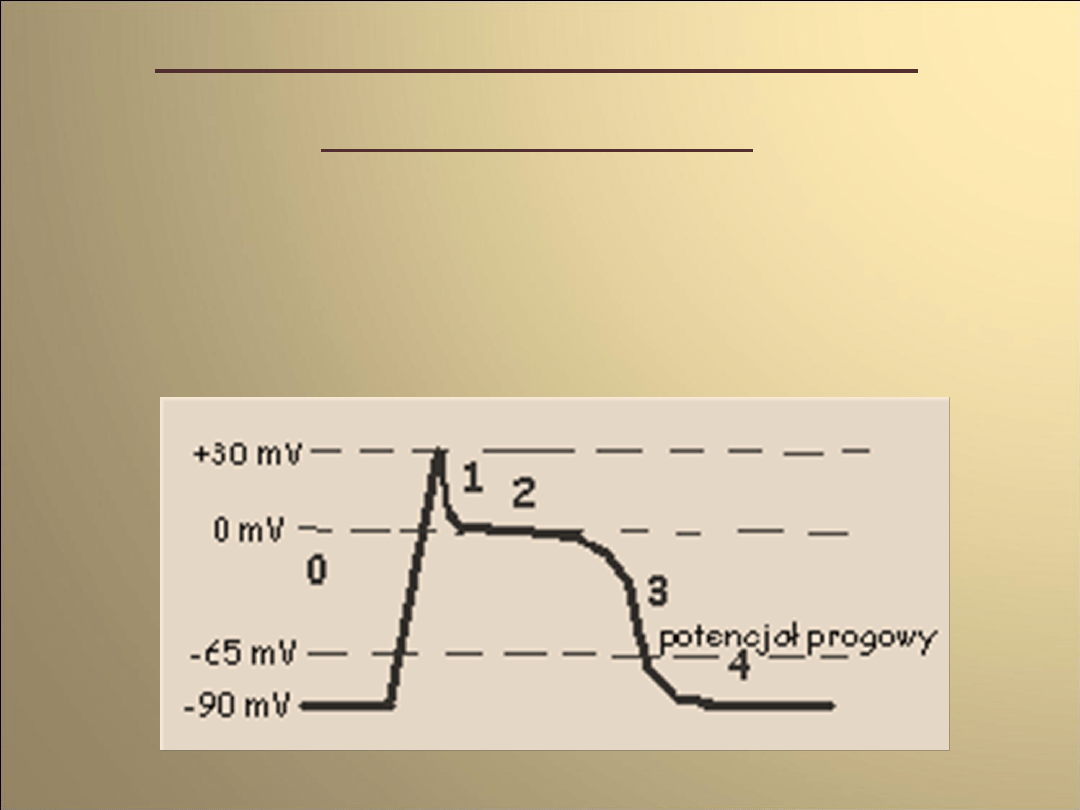
Faza 0
pobudzenie i przekroczenie przez potencjał błony
wartości progowej
aktywacja kanałów sodowych
(napływ jonów sodu do wnętrza komórki)
gwałtowna depolaryzację błony (+25mV)
napływ jonów chlorkowych do wnętrza komórki
(w odpowiedzi na zmianę potencjału błonowego)
Faza 1
spadek potencjału błonowego
(przez prąd chlorkowy oraz zmniejszenie
prądu sodowego)
przeciwdziałanie spadkowi potencjału
ok.-35mV.
(dokomórkowy prąd wapniowy-
aktywowane napięciem kanały
wapniowe typu L)
Faza 2
równoważenie repolaryzujących prądów
chlorkowego oraz potasowego przez
depolaryzujący prąd wapniowy
(przez około 150ms potencjał błonowy
pozostaje na stałym poziomie = faza
plateau)
inaktywacja kanałów wapniowych
pod koniec tej fazy
Faza 3
repolaryzacja błony
(rosnąca przewaga prądów
repolaryzujących)
zmniejszanie wartości prądów potasowego
(przewodzony przez opóźnione kanały
prostownicze) oraz chlorkowego
(w miarę zbliżania się potencjału błony do
wartości spoczynkowej )
Faza 4
Osiągnięcie przez błonę
potencjału spoczynkowego
0
2
3
Reakcja na ponowne
pobudzenie
Refrakcja
bezwzględna-
niezdolność komórki
do osiągnięcia
potencjału
czynnościowego
( faza 0,1,2, część3)
Refrakcja względna
obejmuje koniec fazy
3 i początek fazy 4 -
komórka stopniowo
odzyskuje
pobudliwość; można
ją wtedy pobudzić

Oszczędność energii
Stosunek czasu spędzanego w stanie
depolaryzacji jest znacznie dłuższy niż
komórka spędza w czasie spoczynku
( w porównaniu z komórką nerwową).
Dlatego komórki serca wypracowały
mechanizm oszczędności energii:
•Mała gęstość kanałów w sarkolemie,
•Prąd potasowy w komórce roboczej jest większy w
stanie spoczynku niż w stanie zdepolaryzowanym
(odwrotnie niż w neuronie)
- jest to możliwe dzięki prostowniczym
dokomórkowym kanałom potasowym, które
zamykają się w czasie depolaryzacji, a otwierają w
czasie hiperpolaryzacji (tzw. prostowanie
„anormalne”)

Regulacja siły skurczu
Siła skurczu
zależy od:
•ilości utworzonych wiązań aktyna-
miozyna
(a więc od ilości jonów wapniowych
w sarkoplazmie)
•czynników inotropowych
(substancji wpływających na siłę
skurczu dodatnio lub ujemnie)
Regulacja
częstotliwości pracy
serca
PRZYSPIESZE
NIE
ZWOLNIENIE
NORADRENALINA
+
Wydzielana przez neurony
układu współczulnego.
Zwiększa przepuszczalność
błony dla jonów:
*K⁺ - skraca fazę
repolaryzacji, a w jej
końcowej części błona
osiąga niższy potencjał
(poniżej – 75mV)
*Ca²⁺ - przyspiesza
depolaryzację i skraca czas
trwania potencjału
czynnościowego
ACETYLOCHOLINA
-
Wydzielana przez układ
przywspółczulny
Powoduje zmniejszenie
stopnia ufosforylowania
białek kanałowych –
wzrasta prąd potasowy i
zmniejsza się prąd
wapniowy = zmniejszenie
częstotliwości potencjałów
czynnościowych komórek
rozrusznikowych.
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
Wyszukiwarka
Podobne podstrony:
seminarium 36 podpunkt 4wer1 ppt
seminarium 36 podpunkt 5wer1 ppt
seminarium 36 podpunkt 3wer2 ppt
seminarium 36 podpunkt 2wer1 ppt
seminarium 36 podpunkt 1glikozydy ppt
seminarium 36 podpunkt 1ukladbodzcowoprzewodzacy ppt
seminarium 36 podpunkt 1wer2bezglikozydow ppt
36 NAUKA I NAUCZYCIELE W SEMINARIUM
Higiena seminaria, Kosmetologia 9 Higiena psychiczna
Seminarium IIIR do kopiowania
Prezentacja na seminarium
SEMINARIUM IMMUNOLOGIA Prezentacja
Seminarium3 Inne zaburzenia genetyczne
3 Seminarium Patofizjologia chorób rozrostowych
Seminarium 6 Immunologia transplantacyjna farmacja 2
Biochemia z biofizyką Seminarium 2
seminarium2 2
więcej podobnych podstron