Cechy ilościowe
Wprowadzenie do zagadnienia

Cechy ilościowe - definicja
• Cechy dziedziczne, których
wartości są mierzone w skali
ciągłej (
albo: wykazują zmienność
ciągłą)
.
Indywidualne pomiary danej cechy
u wszystkich osobników pozwalają
na uszeregowanie ich w klasy
prowadzące do uzyskania rozkładu
normalnego

Cechy ilościowe
(charakterystyka) (1a)
Bydło:
-
Wydajność: mleka, tłuszczu,
białka [kg]
- Zawartość (%)
tłuszczu, białka, laktozy
- zawartość suchej masy
-
Wytrwałość w mleczności
- Długość laktacji
- Masa ciała cielęcia przy urodzeniu
- Masa ciała cielęcia przy
odsadzeniu
- Przyrosty dzienne od urodzenia
do
odsadzenia

Cechy ilościowe
(charakterystyka) (1b)
Bydło:
-
Przyrosty dzienne od urodzenia
do
odsadzenia
- Zużycie paszy na jednostkę
przyrostu
- Wydajność rzeźna (%)
- Jakość tuszy
- Powierzchnia „oka” polędwicy

Cechy ilościowe
(charakterystyka) (2a)
Owce
-
Wielkość miotu (liczba jagniąt
urodzonych)
- Masa ciała przy urodzeniu
- Masa ciała przy odsadzeniu
- Wydajność wełny potnej
- Wydajność wełny czystej
- Wysadność wełny
- Grubość wełny
- Karbikowanie wełny

Cechy ilościowe
(charakterystyka) (3a)
Świnie
-
Wielkość miotu
(liczba prosiąt urodzonych)
- Masa ciała przy urodzeniu
(prosię
i miot)
- Masa ciała przy odsadzeniu
(
prosię i miot)
- Przyrosty dzienne od odsadzenia
do
masy c. 96 kg
-Efektywność przyrostów
- Wiek przy osiągnięciu m.c 96 kg

Cechy ilościowe
(charakterystyka) (3b)
Świnie
-
Długość tuszy
- Grubość słoniny na grzbiecie
- Wielkość i kształt szynki
- Powierzchnia „oka” polędwicy
- Wydajność rzeźna
- Mięsność
- i... wiele innych

Cechy ilościowe
(charakterystyka) (4a)
Kury – typ nieśny
-
Wiek dojrzewania
- Nieśność początkowa
- Masa jaja
- Kształt jaja
- Masa ciała kury nioski

Cechy ilościowe
(charakterystyka) (4b)
Kury – typ mięsny
- Masa ciała w wieku 4 tygodni
- Masa ciała w wieku 8 tygodni
- Kąt piersiowy
- Długość mostka
- Długość skoków
- Zużycie paszy na jednostkę
przyrostu
- Cechy użytkowości mięsnej
(podobnie jak
u innych gatunków)

Cechy ilościowe
Model dziedziczenia
Cechy ilościowe (grubość
słoniny)
Linia o cienkiej
słoninie
aabb
(2 cm)
Linia o grubej
słoninie
AABB
[(2 +(4x0,5)] = 4 cm
aabb x
AABB
F
1
AaBb
2 + (2 x 0,5) = 3
cm
Krzyżowanie osobników F
1
między sobą
AaBb
X
AaBb

Rozkład grubości słoniny w
pokoleniu F
2
(w nawiasie l-ba genów
dominujących)
♀ / ♂
AB
Ab
aB
ab
AB
AABB
(4)
AABb
(3)
AaBB
(3)
AaBb
(2)
Ab
AABb
(3)
AAbb
(2)
AaBb
(2)
Aabb
(1)
aB
AaBB
(3)
AaBb
(2)
aaBB
(2)
aaBb
(1)
ab
AaBb
(2)
Aabb
(1)
aaBb
(1)
aabb
(0)

Współczynniki rozwiniętego
dwumianu
(a + b )
n
(trójkąt Pascala)
1
1
1
2
1
1
3
3
1
1
4
6
4
1
1
5
1
0
1
0
5
1
1
6
1
5
2
0
1
5
6
1

Zastosowanie dwumianu do
określania frekwencji i wartości
genotypów cech kumulatywnych
(a + b)
4
= 1a
4
+ 4a
3
b + 6a
2
b
2
+ 4 ab
3
+ 1b
4
Genotypy:
Liczebność
1/16 4/16 6/16 4/16 1/16
Wartości (cm)
4 3,5 3,0 2,5 2,0
Znaczenie wykładników:
- przy symbolu „a” liczba genów dominujących,
- przy symbolu „b” liczba genów recesywnych
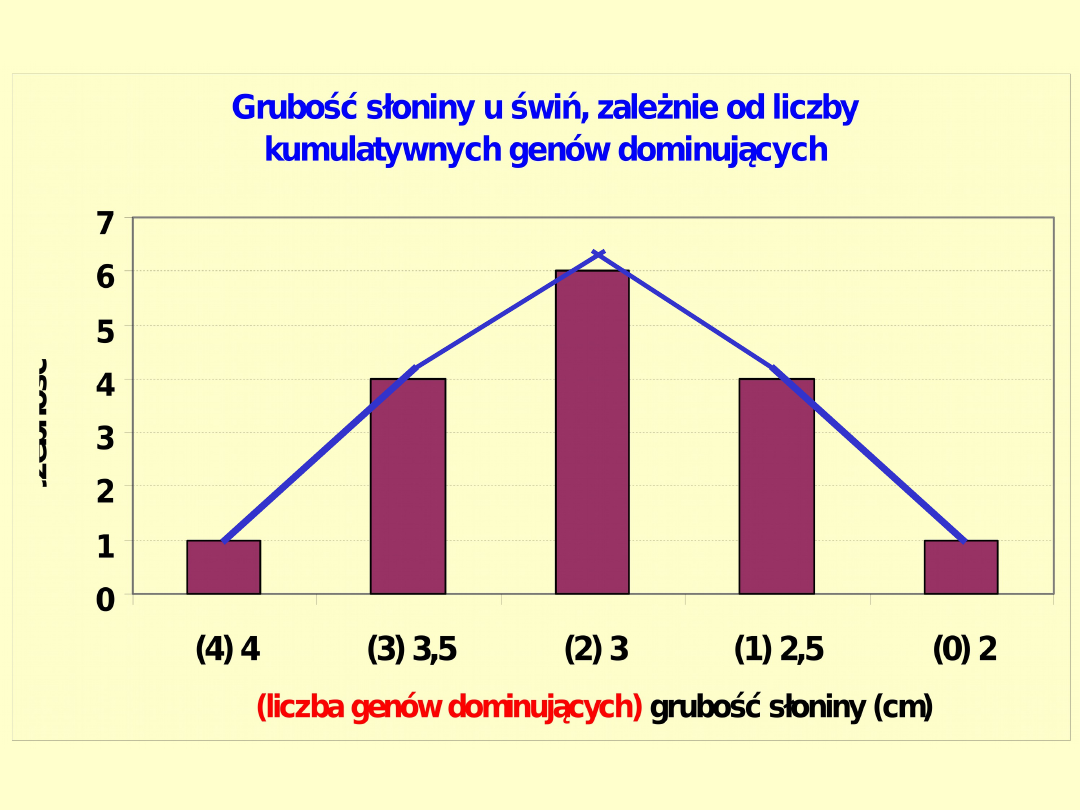
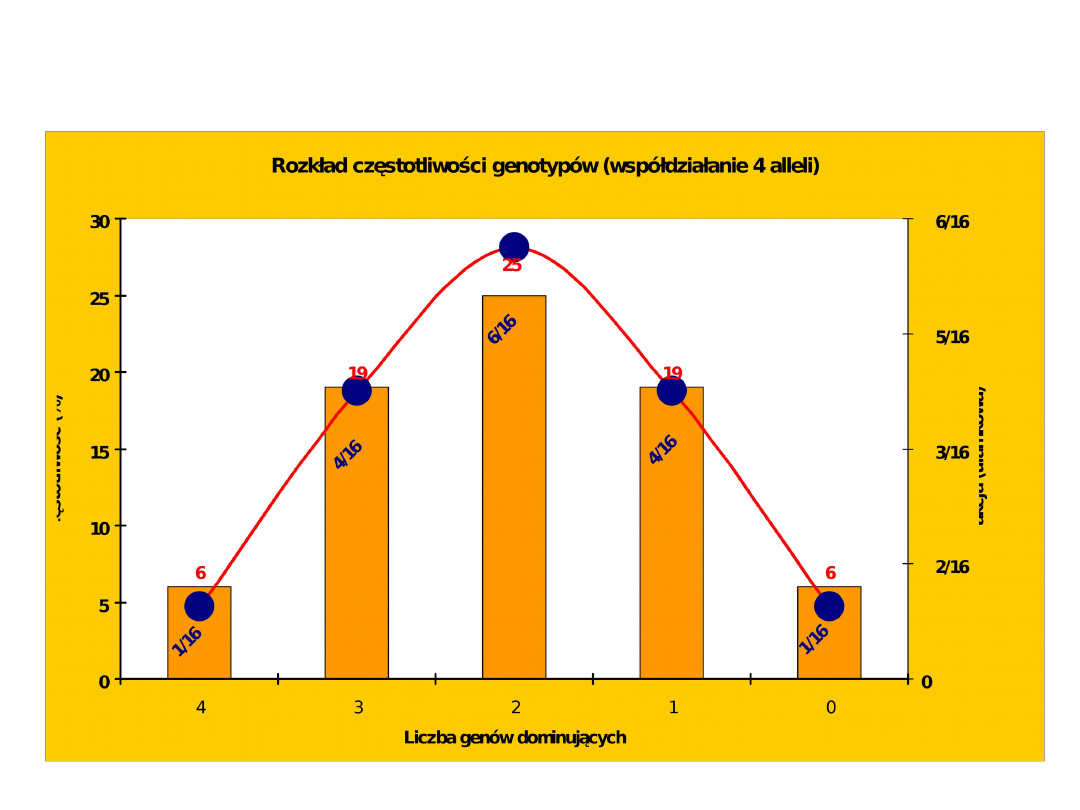
Grubość słoniny u świń, zależnie od liczby kumulatywnych genów
dominujących (1)
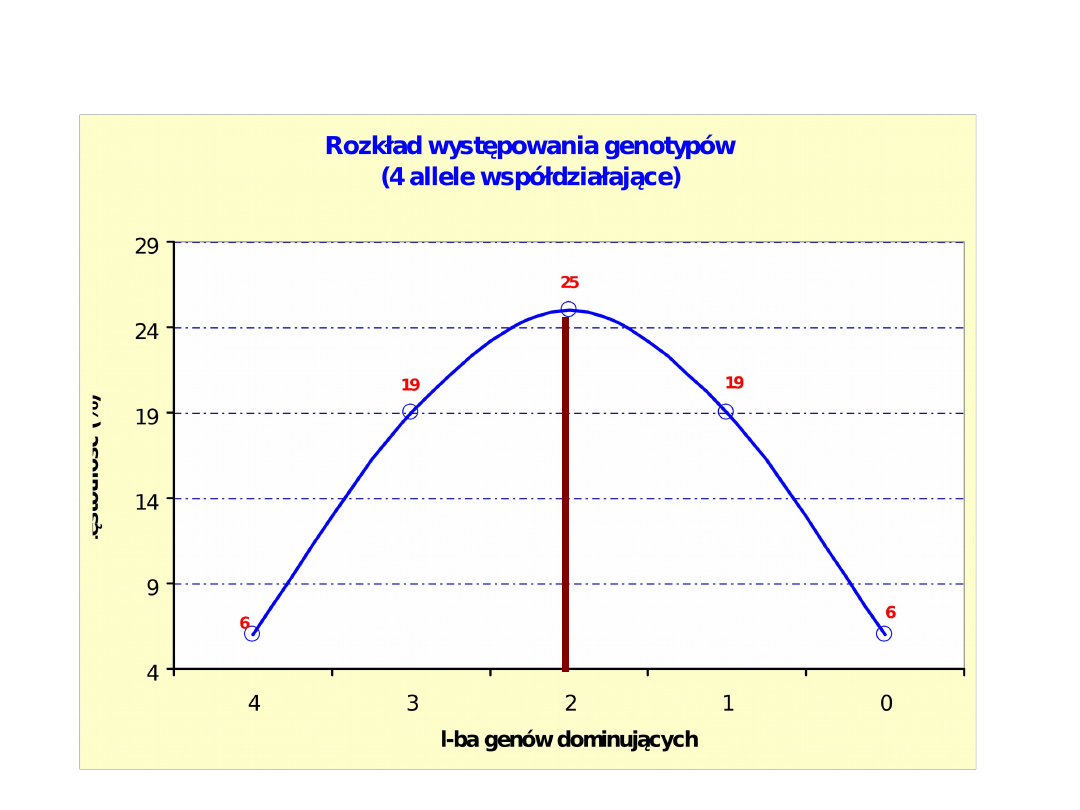
Grubość słoniny u świń, zależnie od liczby kumulatywnych genów
dominujących (2)

Przykład:
dziedziczenia
wzrostu u krów (3 pary genów
kumulatywnych
)
AABBCC
170
cm (maksymalny wzrost)
aabbcc
140
cm (minimalny wzrost)
• Różnica między efektami genów
recesywnych a dominujących
= 170 – 140
= 30 cm
• Przyjmując, że efekty wszystkich genów
dominujących są identyczne, wtedy efekt
pojedynczego genu dominującego wyniesie:
30 : 6 =
5
cm,

1)
Z krzyżowania odmiennych potrójnych
homozygot:
AABBCC
x
aabbcc
uzyska się potrójne heterozygoty
:
A
a
B
b
C
c
u których spodziewana wysokość w kłębie
wyniesie:
140
+ (
3
*
5
cm) =
140
+
15
=
155
cm.
2)
Kojarzenie losowe potrójnych heterozygot
między sobą:
A
a
B
b
C
c x
A
a
B
b
C
c
Liczba możliwych rodzajów gamet
= 2
n
Gdzie:
n = l-ba par alleli w stanie heterozygotycznym
Liczba możliwych rodzajów gamet
= 2
3
=
8
Liczba możliwych kombinacji 8 x 8 = 64
Z tego genotypy z 0, 1, 2, 3, 4, 5, 6 genami
dominującymi:

Przykład dziedziczenia wzrostu u
krów (3 pary genów
kumulatywnych
)
(a + b)
6
=
1
a
6
+ 6a
5
b + 15a
4
b
2
+ 20a
3
b
3
+ 15a
2
b
4
+ 6ab
5
+ 1b
6
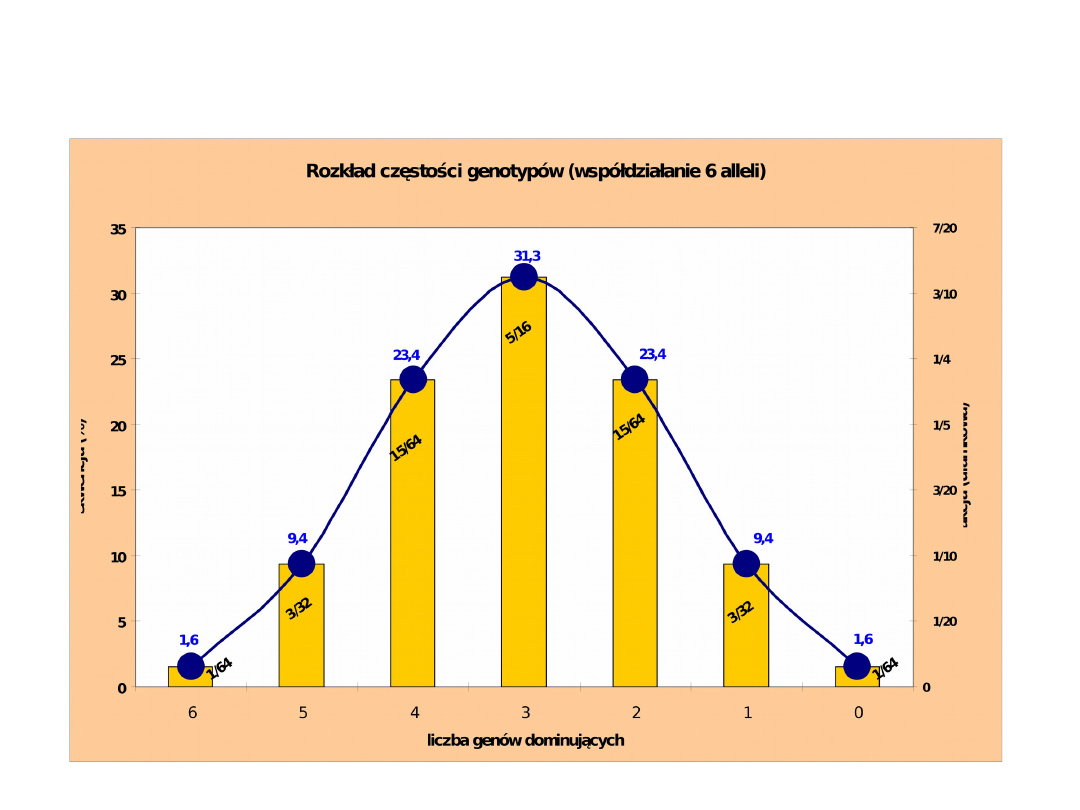
Genotypy:
Częstotliwość:
1/64 6/64 15/64 20/64 15/64 6/64
1/64
Wartości
170 165 160 155 150 145
140
(wzrost w cm)
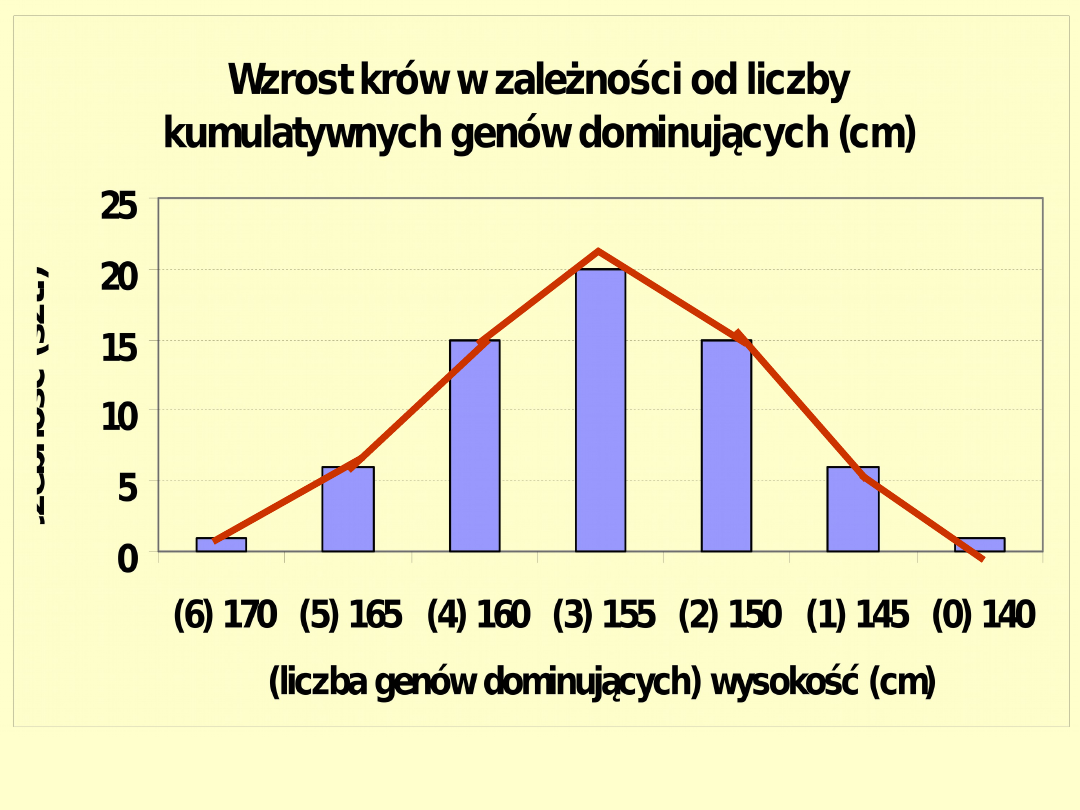
Wzrost krów w modelu z 6 kumulatywnymi genami (1)
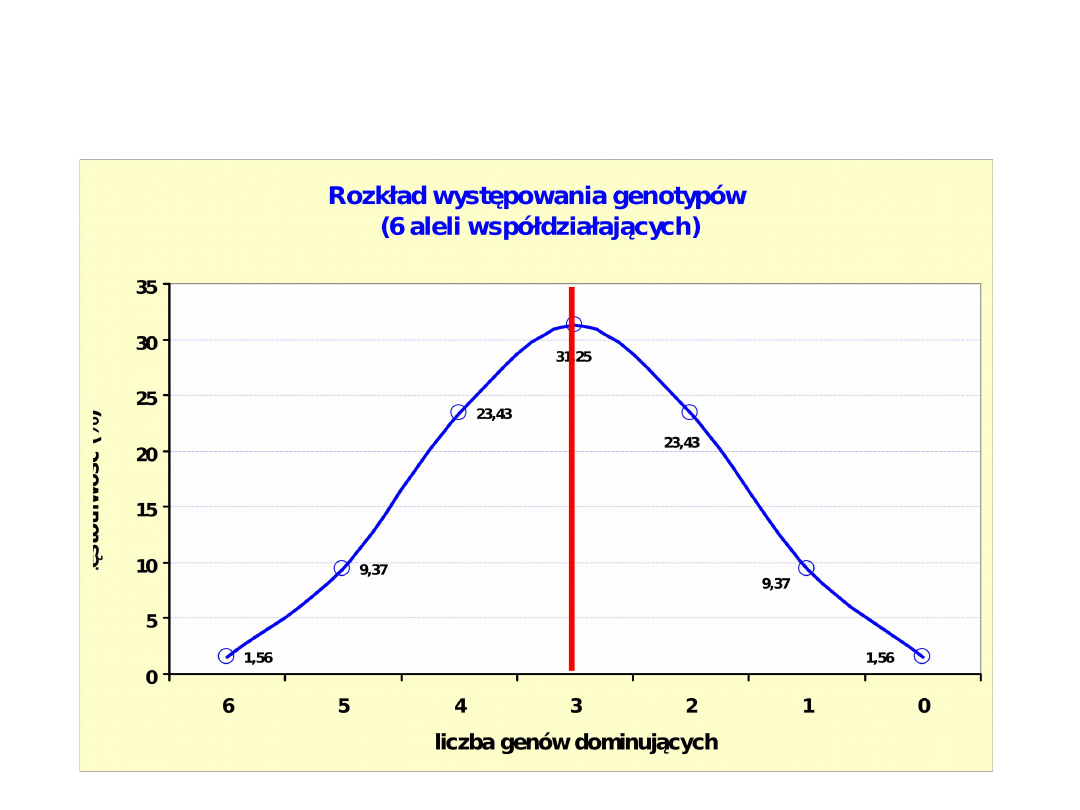
Wzrost krów w modelu z 6 kumulatywnymi genami
(2)
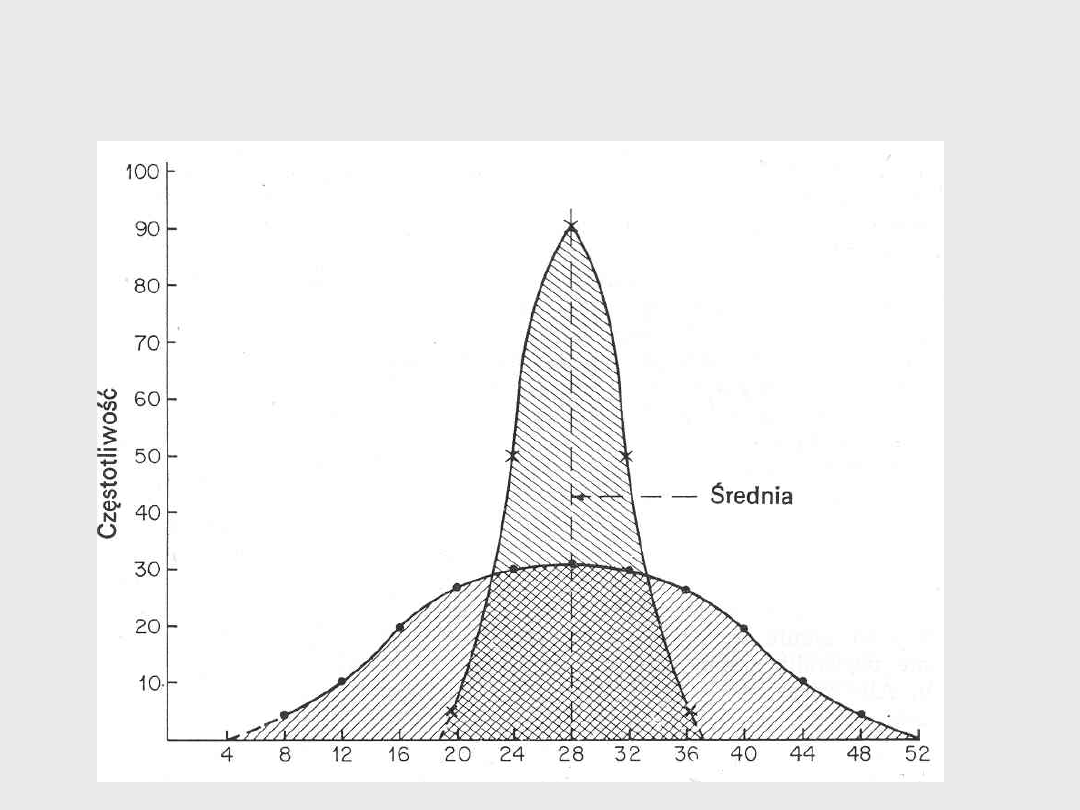
Rozkład wartości cech
kumulatywnych
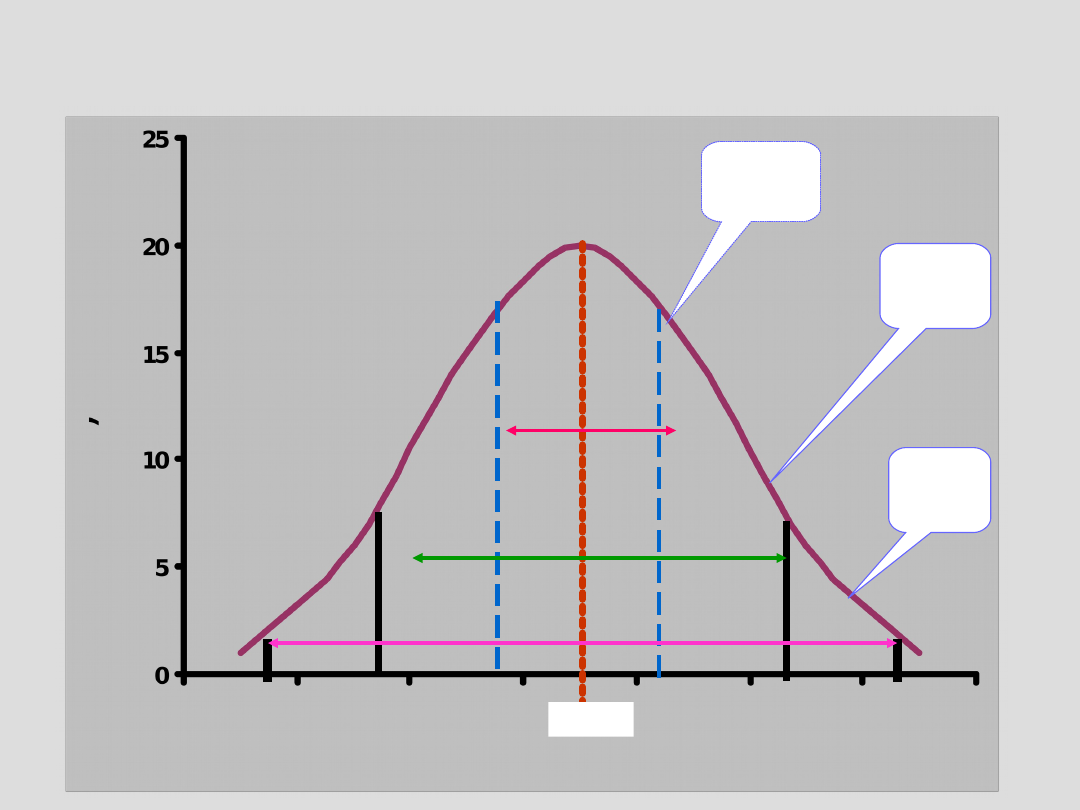
Rozkład wartości wokół średniej
x -
1∂
x +
1∂
x
+2∂
x
+3∂
x -
2∂
x -
3∂
± 1∂
68%
± 2∂
95%
± 3∂
99%
x

Właściwości populacji w
odniesieniu do cech
ilościowych
• średnia
• wariancja
• Odchylenie standardowe
• kowariancja
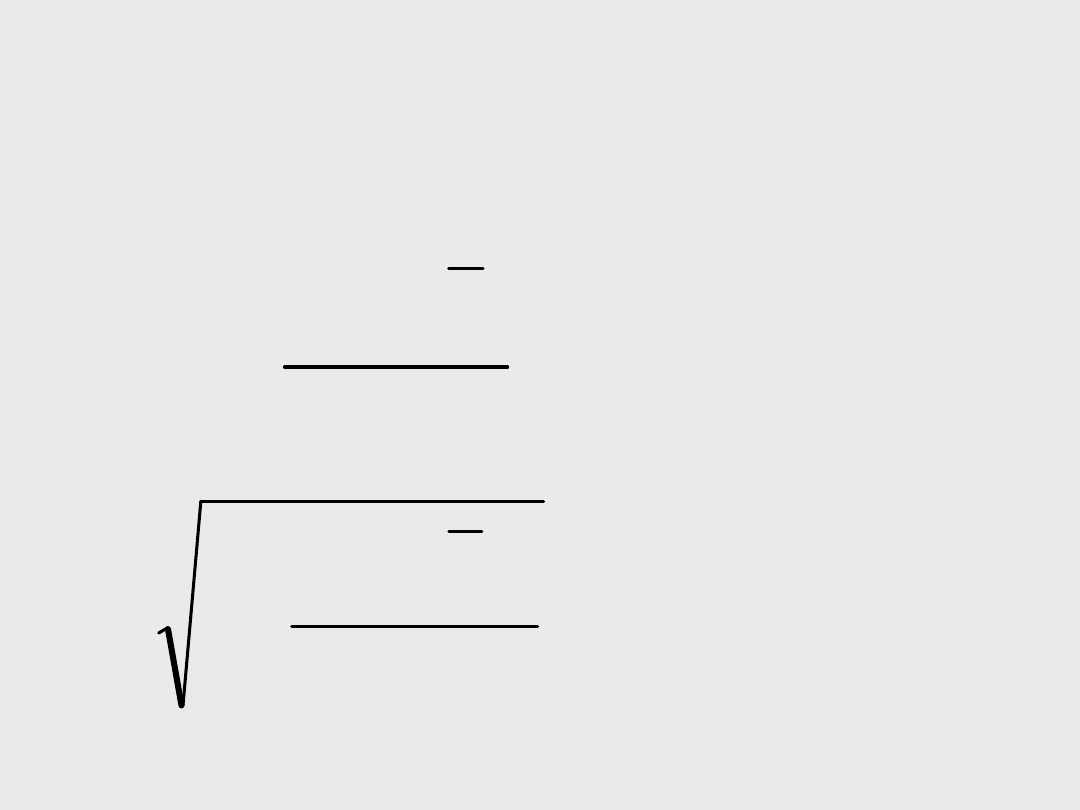
Miary rozkładu
• wariancja: średni
kwadrat odchyleń
od średniej
• odchylenie
standardowe:
pierwiastek
średniego kwadratu
odchyleń od średniej
2
2
1
(
n
x
X
1
(
2
n
x
X
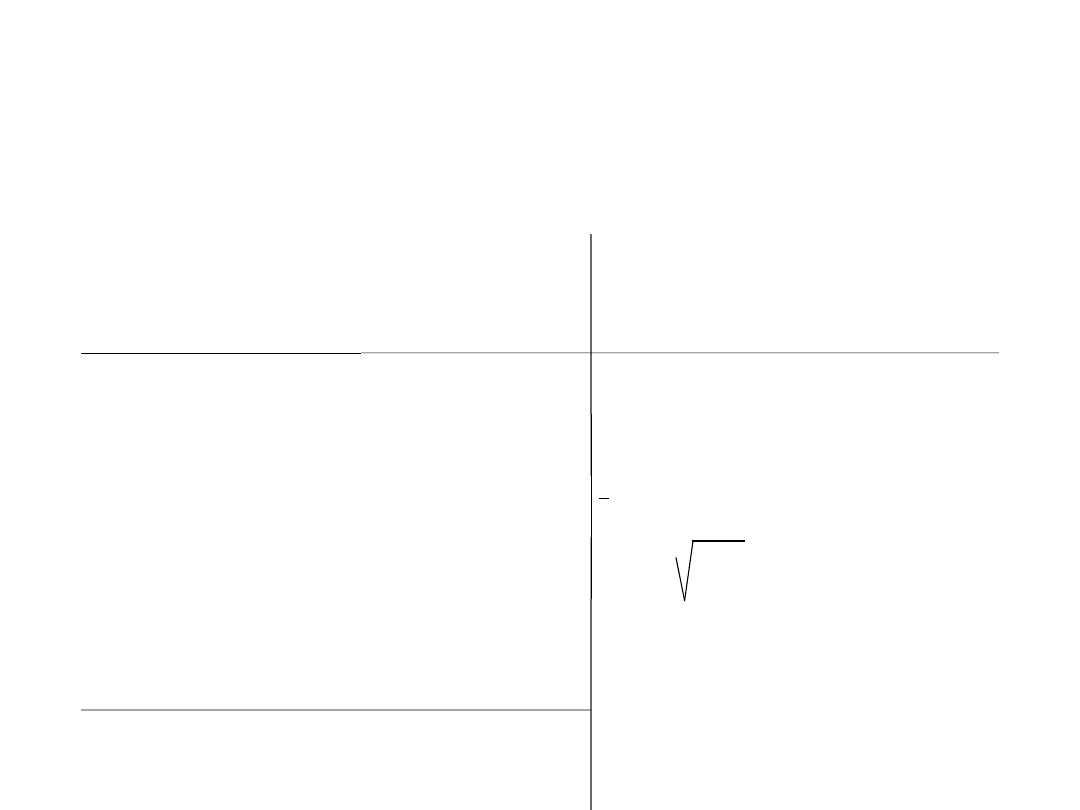
Wariancja
(
∂
2)
i odchylenie
standardowe
(
∂
) -
przykłady
X
i
(X
i
–
x)
(X
i
– x)
2
5
0
0
∂
2
=
10
: 4
=
2,5
6
+1
1
4
- 1
1
3
- 2
4
∂
=
2,5
=
1,58
7
+2
4
x = 5
0
Σ(X
i
– x)
2
=
10
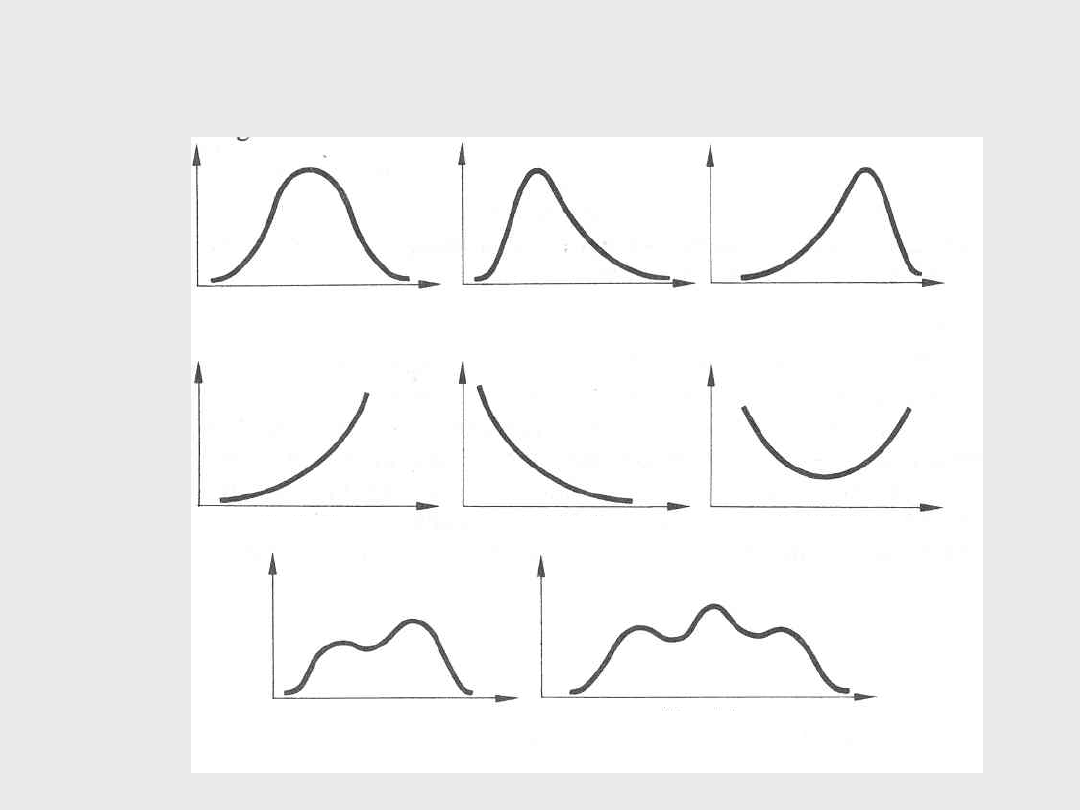
Kształty krzywych rozkładu
Kształt dzwonu
lewoskośna
prawoskośna
J - kształtna
i - kształtna
u - kształtna
dwuszczytowa
wieloszczytowa
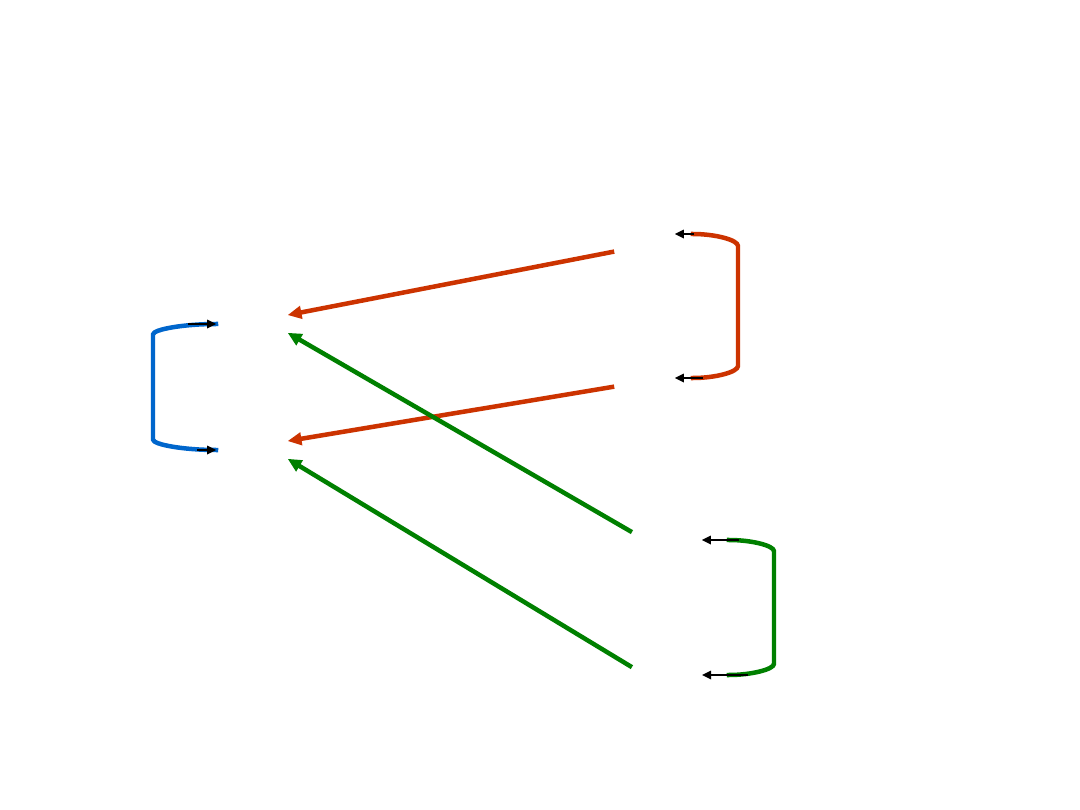
Korelacje genetyczne i
fenotypowe
G
1
P
1
G
2
P
2
E
1
E
2
r
p
r
G
r
E
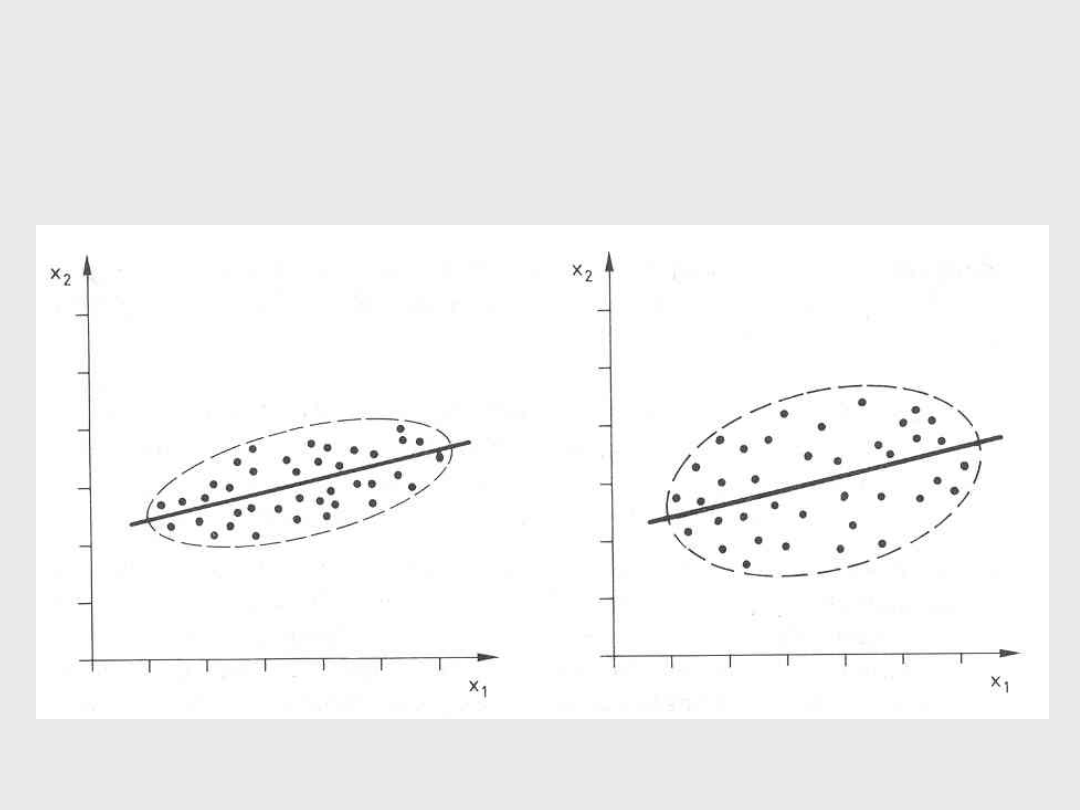
Korelacje (1)
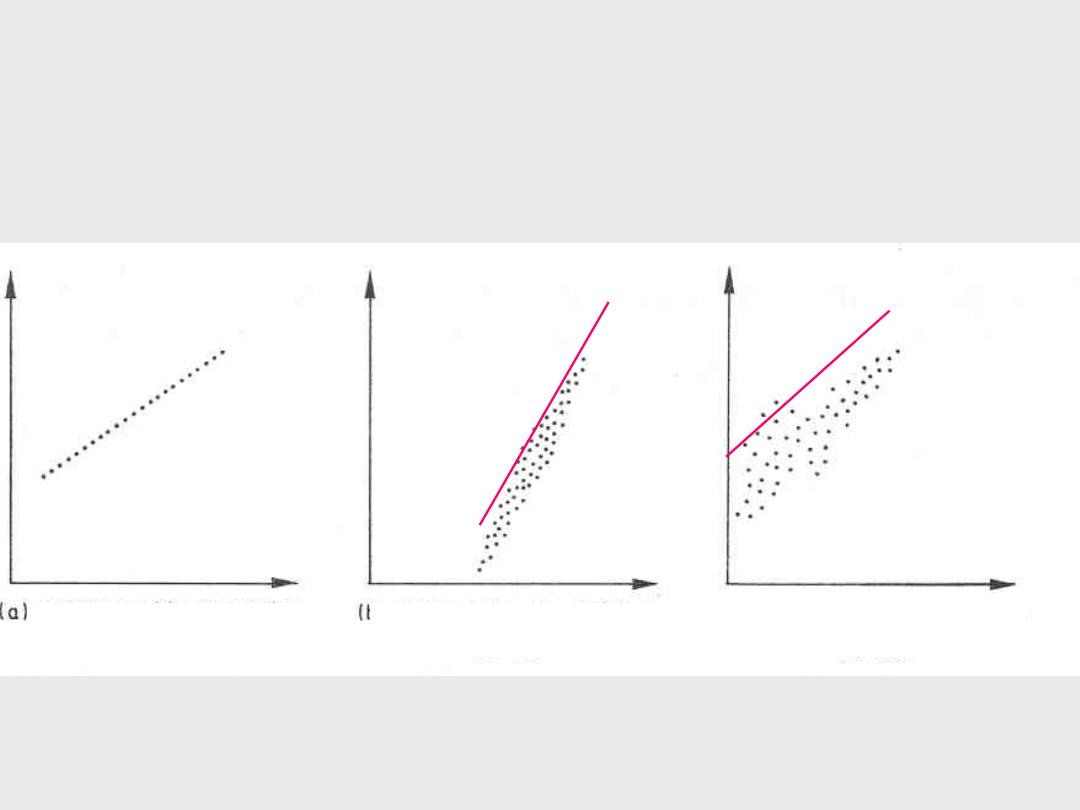
Korelacje (2)
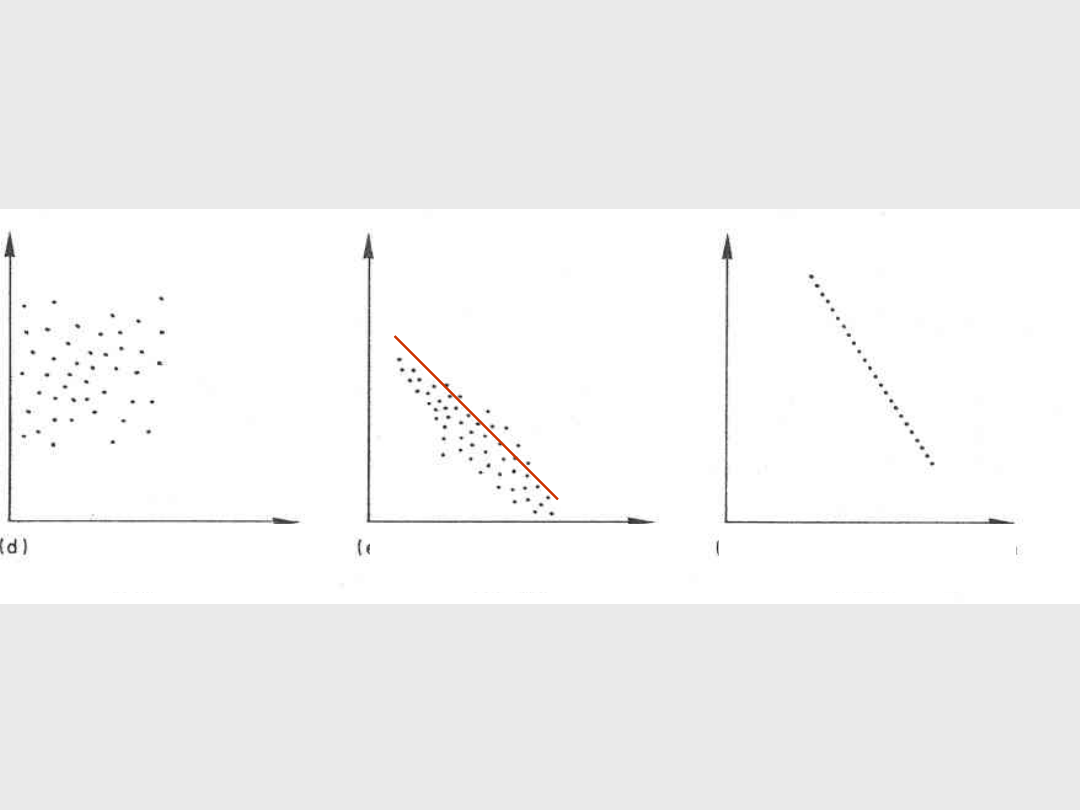
a) Całkowita korelacja dodatnia
r = 1
b) Silna korelacja dodatnia
r = + 0,7
c) Słaba korelacja dodatnia
r = + 0,5
Korelacje (3)
d) Korelacja bliska zeru
r = 0
e) Korelacja ujemna
r = - 0,6
e) Całkowita korelacja ujemna
r = - 1
Korelacje (4)
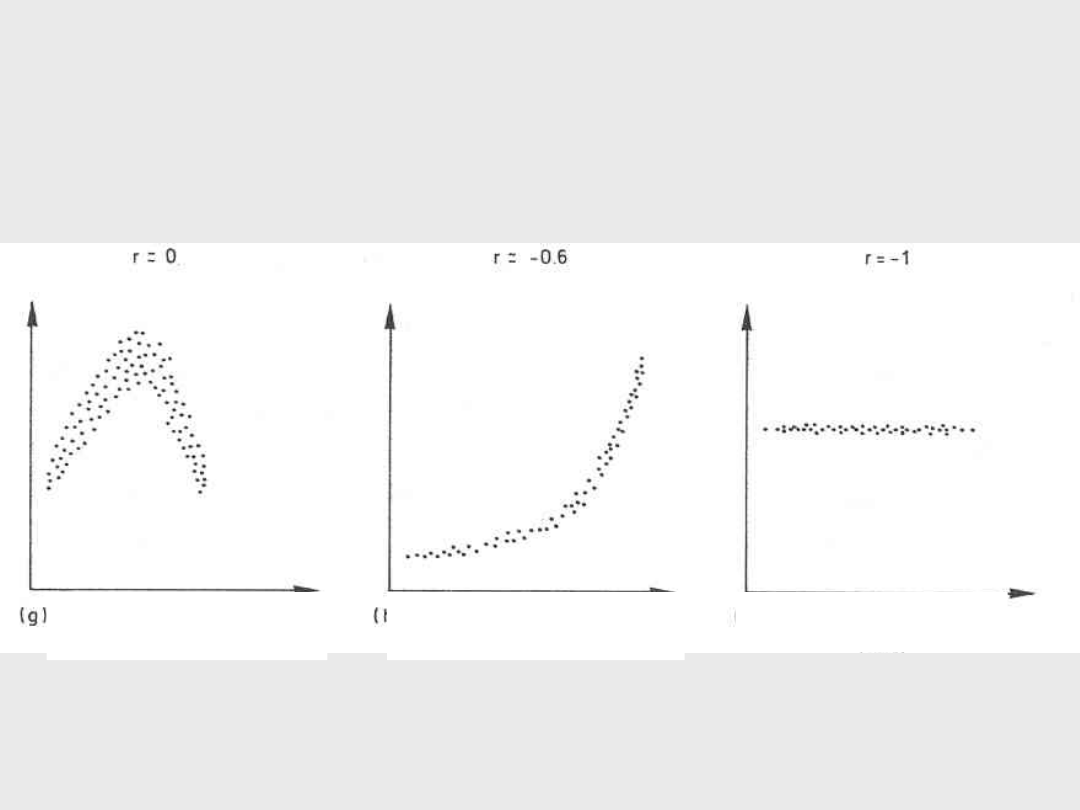
g) Korelacja nieliniowa
r = 0
h) Silna korelacja nieliniowa
r = + 0,5
i)
Brak korelacji
r = 0

Korelacje (5)
j) Współczynnik korelacji
niezdefiniowany
k) Brak korelacji
r = 0
l) Współczynnik korelacji
niezdefiniowany

K o n i e c w y k ł a d u
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
Wyszukiwarka
Podobne podstrony:
Cechy ilosciowe sciaga
Metody ilosciowe wyklad 1
Cechy uwarunkowane wieloczynnikowo cechy ilościowe,8 05
studenci cechy ilościowe
Cechy ilościowe
Wykład 9 ILOŚCIOWE PODEJŚCIE DO ZARZĄDZANIA
rozpiska analityczna wykłady, farmacja, II sem, chemia ilościowa
egzamin, 07 - Omów cechy wspólne metod jakościowych oraz cechy wspólne metod ilościowych i dokonaj i
Prelekcja 11 Cechy poligenowe ilościowe
1d Wykład CECHY RYNKOWE NIERUCHOMOŚCIid 18890 ppt
Metody ilościowej i jakościowej oceny ryzyka wykład
Szczególne cechy gosp. rol. i war. jej rozw wykład I, Ekonomika
chemia an wykład 16.10 , farmacja, II sem, chemia ilościowa
więcej podobnych podstron