
NEUROFIZJOLOGIA
UKŁADU RUCHU
KLINIKA REHABILITACJI

Za sterowanie aktywnością motoryczną, a tym
samym za wytwarzanie przez mięśnie odpowiednich sił
i momentów obrotowych odpowiedzialny jest układ
nerwowy
w pewnych krytycznych sytuacjach układ ruchowy pracuje na
granicy swojej wydolności
reakcje ruchowe
odruchy
ruchy dowolne

reakcje ruchowe
Odruchy - są to proste, schematyczne reakcje ruchowe
realizowane bez udziału naszej świadomości, ich
podstawowym zadaniem jest szybkie reagowanie na
bodźce istotne ze względu na bezpieczeństwo organizmu
Łuk odruchowy - schemat przepływu informacji
część dośrodkowa (aferentna) przekazująca informacje od
receptora, do określonego obszaru układu nerwowego.
w odpowiedzi na ten bodziec uruchamiany jest gotowy, zazwyczaj
zdefiniowany przez samą strukturę połączeń nerwowych, program
ruchowy
program ten przekazywany jest do mięśniowych elementów
wykonawczych drogą odśrodkową, czyli eferentną – stąd bierze się
ich duża szybkość
i skuteczność realizacji

Analizując neuronalną kontrolę ruchu, można mówić o
podwójnym sterowaniu aktywnością mięśni: centralnym i
obwodowym
w motoneuronach rogów przednich rdzenia kręgowego
następuje integracja kontroli obwodowej z ośrodkową, której
wynikiem jest tonus mięśniowy oraz aktywny skurcz
•odruchy obronne
•korekcje
dostosowawcze
•uczenie się
•tworzenie programów
ruchowych
•rozpoczęcie i zakończenie
ruchu
•korekcje programu

centralny generator wzorca lokomocyjnego
(ang. central pattern generator, CPG)
generator uruchamiany jest tonicznymi sygnałami
pochodzącymi
z najwyższych pięter mózgu: kory asocjacyjnej i jąder
podstawnych
na poziomie kory mózgowej wyznaczany jest cel ruchu i wybierany jest
sposób jego realizacji
w momencie zrealizowania celu ośrodkowy system motywacyjny wyłącza
generator rdzeniowy
skuteczna realizacja zadania ruchowego wymaga strategii, o
której decydują najwyższe piętra układu nerwowego, lecz
także pewnych działań taktycznych
ośrodki kontroli taktycznej, przystosowują zgrubny wzorzec
aktywności mięśniowej do warunków otoczenia
kora ruchowa i móżdżek
obwodowa kontrola ruchu, pozwala np. na dostosowywanie położenia stopy
do nierównomierności podłoża, obwodowe sygnały sterujące powodują
nawet znaczne korekcje wzorców aktywności mięśniowej bez zakłócania
rytmu lokomocji

Kontrola biomechanicznych parametrów
mięśni na poziomie rdzenia kręgowego
somatyczny układ ruchowy - mięśnie szkieletowe wraz z
częścią układu nerwowego, która je kontroluje
dwa poziomy sterowania mięśniami:
•
rdzeniowy - na poziomie rdzenia generowane są programy
skoordynowanego skurczu mięśni
•
ośrodkowy - wykonanie oraz dostosowanie programów do
zmieniających się warunków
•
motoneuron (neuron ruchowy) - komórki nerwowe sterujące
aktywnością mięśni szkieletowych, to jedyne neurony, które
bezpośrednio łączą się
z komórkami mięśniowymi i są odpowiedzialne za skurcz
mięśni

Kontrola biomechanicznych parametrów
mięśni na poziomie rdzenia kręgowego
układ nerwowy kontroluje parametry wykonywanego ruchu przez:
•
zmianę siły skurczu właściwej grupy mięśni (ang. prime movers)
•
modulowanie skurczu przed jednoczesny współskurcz mięśni
antagonistycznych, współpobudzenie (ang. coactivation) mięśni
antagonistycznych
•
mięśniowe odruchy rdzeniowe
neurony ruchowe: motoneurony alfa, beta i gamma
•
alfa - elementy wyjściowe sterowania motorycznego rozmieszczone
w rogach przednich rdzenia kręgowego, w nich następuje integracja
sterowania ośrodkowego i obwodowego zapewniająca właściwą realizację
ruchu, w wyniku czasowo-przestrzennego sumowania
•
beta - unerwiają włókna mięśniowe oraz włókna śródwrzecionowe -rola
tych komórek nerwowych nie została jeszcze do końca poznana
•
gamma - pobudzenie wywołuje skurcz włókien ruchowych
wyspecjalizowanych receptorów - wrzeciona mięśniowe

motoneuron alfa
motoneuron alfa - może kontrolować skurcz od kilku do
kilkuset włókien mięśniowych
zespół funkcjonalny, składający się z komórki ruchowej alfa
oraz unerwianych przez nią włókien mięśniowych, został
nazwany
Charlesa Sherringtona jednostką ruchową
w polskim piśmiennictwie stosuje się wymiennie nazwę jednostka
motoryczna

wiązki wypustek aksonalnych motoneuronów opuszczają
rdzeń kręgowy, tworząc korzeń przedni (brzuszny), który
następnie po połączeniu
z przewodzącym sygnały czuciowe korzeniem tylnym
(grzbietowym) tworzy wspólnie nerw rdzeniowy
nerwy przez obustronne wcięcia kręgowe wydostają się poza
kręgosłup
i przez rozgałęziającą się sieć nerwów obwodowych docierają
do poszczególnych mięśni
•
nerwy szyjne (C1-C8)
•
nerwy piersiowe (T1-T12)
•
lędźwiowe (L1-L5)
•
krzyżowe (S1-S5)

Typy jednostek ruchowych
włókna czerwone - zawierające dużo mitochondriów i enzymów,
które wyspecjalizowane są w metabolizmie tlenowym
•
włókna te tworzą wolne jednostki ruchowe
•
czas narastania skurczu jest bardzo długi i sięga nawet 100 ms
•
są w stanie utrzymać napięcie przez dłuższy czas
•
włókna odporne na zmęczenie
•
liczba wolnych włókien mięśniowych tworzących pojedynczą jednostkę ruchową zmienia się
w granicach 10-180
•
przez wolne jednostki ruchowe układ nerwowy jest wstanie precyzyjnie kontrolować siłę
skurczu mięśnia
•
białe włókna - zawierają mniej mitochondriów, a ich aktywność
opiera się na metabolizmie beztlenowym
•
kurczą się znacznie szybciej
•
czas narastania skurczu wynosi około 50 ms
•
na każdą jednostkę ruchową przypada 300-800 włókien mięśniowych
•
szybkie jednostki ruchowe mogą wytwarzać znaczną siłę skurczu, jednakże gradacja tej siły
jest znacznie gorsza
•
jednostki te nie są w stanie utrzymać napięcia przez dłuższy czas

Typy jednostek ruchowych
w skład pojedynczej jednostki ruchowej wchodzi tylko jeden typ
włókien
obydwa typy włókien występują w różnych proporcjach w
poszczególnych mięśniach
•
proporcje oraz bezwzględna liczba włókien określają parametry biomechaniczne
danego mięśnia
motoneurony unerwiające poszczególne typy włókien mięśniowych
różnią się między sobą
•
ciała komórkowe oraz średnice aksonów są znacznie większe w neuronach
ruchowych kontrolujących szybkie włókna mięśniowe - motoneurony alfa szybkich
jednostek ruchowych generują krótkie salwy impulsów o częstotliwości 30-60 Hz
•
neurony ruchowe kontrolujące wolne włókna mięśniowe generują stałe ciągi
potencjałów czynnościowych z częstodiwością 10-20 impulsów na sekundę
średnica włókien nerwowych - aksony o większej średnicy
unerwiają więcej włókien mięśniowych, czyli wchodzą w skład
większych jednostek ruchowych
•
średnica aksonu decyduje o szybkości przewodzenia potencjałów czynnościowych

Kontrola siły skurczu mięśnia
płytka końcowa lub złącze nerwowo-mięśniowe - połączenie
aksonu
z włóknem mięśniowym
•
nerwowy potencjał czynnościowy docierający do płytki końcowej
powoduje wydzielanie neuroprzekaźnika- acetylocholiny (ACh)
•
pojedynczy potencjał czynnościowy motoneuronu powoduje wyrzut
ACh, we włóknie mięśniowym wytwarzany jest potencjał
postsynaptyczny pobudzający (ang. EPSP), że wystarcza do
wygenerowania potencjału czynnościowego włókna
•
w wyniku działania pojedynczego potencjału czynnościowego we
włóknie mięśniowym generowany jest krótkotrwały skurcz
jednostkowy (ang. twitch)
•
skurcz taki obserwowany jest w postaci szybkiej dwufazowej sekwencji
skrócenia i relaksacji włókna
•
skurcz jednostkowy jest źródłem niewielkiej siły, jednak dzięki
sumowaniu czasowo-przestrzennemu poszczególnych skurczów
jednostkowych we włóknie mięśniowym, a dokładniej w całej
jednostce ruchowej, generowana jest wymagana siła skurczu

Kontrola siły skurczu mięśnia
sumowanie czasowo-przestrzenne
integracja pojedynczych skurczów w ramach tej samej jednostki ruchowej
•
wraz ze wzrostem częstotliwości pobudzenia motoneuronu alfa skraca się czas między
kolejnymi skurczami jednostkowymi włókien mięśniowych, jeśli przerwa między kolejnymi
pobudzeniami będzie na tyle krótka, że nie nastąpi pełna relaksacja jednostki ruchowej,
wystąpi efekt sumowania skurczów
•
siła generowana w czasie takiego skurczu zależy od wielkości jednostki, czyli od liczby
włókien mięśniowych przypadających na tę jednostkę oraz od częstotliwości impulsów
nerwowych
•
powyżej pewnej częstotliwości pobudzenia, przekraczającej w przypadku wolnych
jednostek ruchowych 30 Hz, siła skurczu osiąga maksymalną wartość i dalsze zwiększenie
siły mięśniowej może nastąpić tylko przez rekrutację kolejnych jednostek ruchowych
•
maksymalną częstotliwość pobudzenia motoneuronów ogranicza tzw. hamowanie
autogenne przez niewielkie interneurony - komórki Renshawa
•
liczba aktywnych jednostek ruchowych - zasada rekrutacji jednostek
ruchowych w mięśniu
•
małe jednostki unerwiane przez niewielkie neurony ruchowe są rekrutowane jako pierwsze
•
następnie włączane są coraz to większe jednostki ruchowe
•
tak uporządkowana kolejność włączania się jednostek ruchowych ma wpływ dla
stopniowania, czyli gradacji siły skurczu

Organizacja sterowania pracą mięśni
praca mięśni jest bezpośrednio kontrolowana przez
motoneurony alfa
komórki nerwowe zostały nazwane przez Sheringtona wspólną drogą
końcową układu ruchowego (ang. final common path)
na nich zbiegają się wszystkie pobudzenia wynikające zarówno ze
sterowania obwodowego, jak i centralnego
Istnieją trzy bezpośrednie wejścia synaptyczne na
motoneurony alfa:
•
(1) z wrzecion mięśniowych
•
(2) z kory ruchowej (droga piramidowa)
•
(3) z neuronów wstawkowych (interneuronów) rdzenia kręgowego

Organizacja sterowania pracą mięśni
Kontrola obwodowa
mechanoreceptory - wyspecjalizowane czujniki bodźców mechanicznych
•
rozmieszczone są w mięśniach, w stawach oraz na powierzchni skóry
•
przekazują do mózgu za pośrednictwem nerwów obwodowych informacje o położeniu, ruchu
oraz siłach pojawiających się w poszczególnych elementach łańcuchów kinematycznych
w czasie aktywności ruchowej
•
każdy program ruchowy niezależnie od jego złożoności reprezentowany jest
w mózgu w postaci odpowiadających mu wzorców aktywności sensorycznej
•
wzorce umożliwiają zapoczątkowanie ruchu oraz biorą udział w kontroli jego realizacji
•
mózg, ażeby skutecznie zrealizować określony program ruchowy, musi znać aktualną pozycję
i ruch poszczególnych części ciała względem siebie, położenie całego ciała w przestrzeni
rozbudowane układy sensoryczne tworzą różnorodne pętle sprzężeń
zwrotnych
•
odgrywają największą rolę w czasie nabywania (uczenia się) określonej reakcji ruchowej

receptory mięśniowe, stawowe i skórne
wrzeciona mięśniowe - występują wewnątrz wszystkich
mięśni szkieletowych, przekazują do rdzenia kręgowego
informacje o jego aktualnej długości oraz szybkości
rozciągania mięśnia
•
długość wrzecion mięśniowych wynosi 0,5-13 mm (typowo 2-4 mm)
•
znajdują się we wszystkich mięśniach, jednak są liczniej reprezentowane w
dalszym umięśnieniu kończyn, a szczególnie w krótkich mięśniach dłoni i
stóp
•
receptor wrzecionowy zbudowany jest z kilku (3-10) krótkich - długości kilku
milimetrów - komórek mięśniowych zamkniętych w torebce łącznotkankowej
•
komórki mające zdolność wykonywania aktywnego skurczu, nazywane są
włóknami wewnątrzwrzecionowymi i mają własne unerwienie ruchowe
•
skurczem tych włókien sterują motoneurony gamma
•
w połowie długości kurczliwych włókien rozmieszczone są zakończenia
nerwowych włókien czuciowych Ia - należą do najgrubszych
zmielinizowanych aksonów = 120 m/s
•
włókna la wnikają do rdzenia poprzez korzenie grzbietowe

receptory mięśniowe, stawowe i skórne
wrzeciona mięśniowe
•
w rdzeniu włókna la się rozgałęziają, tworząc pobudzające połączenia synaptyczne
z motoneuronami alfa własnego mięśnia oraz rozbudowaną sieć połączeń z
neuronami wstawkowymi, dzięki czemu zyskują wpływ na aktywność innych mięśni
zarówno synergistycznych, jak i antagonistycznych
rozciąganiu mięśnia towarzyszy wydłużenie wrzeciona mięśniowego
powodując generowanie w zakończeniach włókien czuciowych la impulsów
czynnościowych
•
ich częstotliwość zależy od szybkości i amplitudy rozciągania
•
następuje pobudzenie własnego mięśnia, który próbuje się skurczyć,
przeciwdziałając rozciąganiu
•
wrzeciona mięśniowe wbudowane są w pętlę sprzężenia zwrotnego tworzącego
monosynaptyczny łuk odruchowy - odruch na rozciąganie
mięśnie o przebiegających powierzchniowo ścięgnach są szczególnie przydatne w
ocenie:
•
mięsień trójgłowy łydki
•
czworogłowy uda (odruch kolanowy)

receptory mięśniowe, stawowe i skórne
receptory ścięgnowe
Receptory ścięgnowe, nazywane również narządami
ścięgnowymi Golgiego, bodźcem specyficznym dla receptora
ścięgnowego jest naprężenie wywołane skurczem pęczka
włókien mięśniowych, hamują aktywność motoneuronów alfa
własnego mięśnia
•
w obszarze złącza mięśniowo-ścięgnowego zakończenia sąsiednich włókien
mięśniowych łączą się ze sobą, tworząc pęczki liczące 10-15 włókien
•
w tym rejonie znajdują się zakończenia czuciowe receptora ścięgnowego
•
generowane potencjały czynnościowe przewodzone są do rdzenia grubymi
włóknami nerwowymi typu Ib, których średnica wynosi 12-20 um
•
włókna te wnikają do rdzenia kręgowego korzeniami grzbietowymi
•
w rdzeniu włókna Ib silnie się rozgałęziają, przekazując sygnały czuciowi do
neuronów wstawkowych rogów przednich
•
sygnały obwodowe z narządów Golgiego przekazywane są również do
móżdżku
•
hamująca aktywność własnych neuronów ruchowych alfa, tworzy obwód
reakcji odruchowej nazywanej odruchem ścięgnistym

receptory mięśniowe, stawowe i skórne
receptory stawowe
Receptory stawowe - przekazują do układu nerwowego
informację
o położeniu stawu, o szybkości ruchu oraz o wielkości i
lokalizacji sił działających na staw
•
znajduje się w torebkach i więzadłach stawowych
•
generują spoczynkowe potencjały czynnościowe z częstotliwością kilku
impulsów na sekundę
•
deformacja struktur stawowych w czasie ruchu powoduje zwiększone
mechaniczne pobudzenie receptorów
•
receptory sygnalizujące położenie stawu wykazują dość znaczną
selektywność kątową
•
wzmożoną aktywność wykazują receptory stawowe w skrajnych położeniach
stawu

receptory mięśniowe, stawowe i skórne
mechanoreceptory skórne
Zidentyfikowano dwa typy mechanoreceptorów wrażliwych
na przemieszczenie oraz na prędkość odkształceń skóry, należą
one do grupy wolno adaptujących się receptorów typu I i II
Typ I – charakteryzują się małymi polami recepcyjnymi,
(ciała dotykowe Meissnera i łąkotki dotykowe Merkela)
•
pod nieobecność bodźców mechanicznych komórki te nie wykazują żadnej
aktywności spontanicznej
•
są bardzo wrażliwe na drobne przemieszczenia skóry
•
receptory te są związane z percepcją kształtu i rodzaju powierzchni
•
największe zagęszczenia receptorów typu I występują na powierzchni dłoni,
stóp, wargach
•
najsłabiej receptory te reprezentowane są na powierzchni tułowia
•
sygnały z ciałek Meissnera mają istotne znaczenie w regulacji siły uścisku
palców lub dłoni
•
ciałka Meissnera są czułymi detektorami szybkości ruchu

receptory mięśniowe, stawowe i skórne
mechanoreceptory skórne
mechanoreceptory skórne Typu II identyfikowane są z
rozlokowanymi głęboko w skórze ciałkami Ruffniego i ciałkami
Paciniego
•
mają znacznie większe i słabiej rozgraniczone pola recepcyjne
•
wykazują aktywność spontaniczną
•
wrażliwe na niewielkie odkształcenia skóry leżącej bezpośrednio nad nimi
(czułości < 15 µm)
•
na rozciąganie skóry wskutek działania bodźców mechanicznych w
obszarach odległych od receptora
•
w odpowiedzi tych receptorów można wyróżnić dwie składowe:
statyczną zależną od siły bodźca mechanicznego
dynamiczną, czyli zależną od szybkości jego zmian

Kontrola ośrodkowa
podstawowy udział w kontroli aktywności neuronów ruchowych mają
ośrodkowe sygnały zstępujące
wykonanie każdego ruchu poprzedzają ośrodkowe procesy decyzyjne
oparte na sensorycznej ocenie otoczenia i warunków realizacji
•
kora somatosensoryczna i wzrokowa
uruchamiany jest odpowiedni program ruchowy za pomocą
decyzyjnych neuronów wstawkowych pola 7 kory ciemieniowej
•
neurony te działają na zasadzie wyłączników, których włączenie pozwala wybrać
w dodatkowej korze ruchowej optymalny - dostosowany do warunków otoczenia
program
•
program kierowany jest przez korę przedruchową do odpowiednich obszarów
pierwszorzędowej kory ruchowej -stąd wysłane są sygnały sterujące do
odpowiednich jednostek ruchowych
optymalne wykonanie ruchu dowolnego wymaga dostosowawczych
zmian postawy ciała, bieżącej kontroli ruchu oraz reagowania na
zakłócenia w jego wykonaniu
•
móżdżek w oparciu o informacje zwrotne z mięśni o skuteczności realizacji
wybranego programu wprowadza odpowiednie korekty

Kontrola ośrodkowa
wszelkie sygnały sterujące przekazywane są do rdzenia kręgowego
kilkoma specyficznymi kanałami informacyjnymi utworzonymi przez
boczne i brzuszno-przyśrodkowe drogi mózgowo-rdzeniowe
•
drogi boczne - wywodzące się z korowych ośrodków ruchowych
ukierunkowane są na aktywność dowolną mięśni dalszych (dystalnych)
kończyn
•
drogi brzuszno-przyśrodkowe, rozpoczynające się w pniu mózgu,
kontrolują przede wszystkim aktywność mięśni osiowych i obręczowych,
czyli mięśni zaangażowanych w utrzymanie postawy ciała i w ruchach
lokomocyjnych
•
drogi łączące ruchowe ośrodki mózgowe z rdzeniem kręgowym
drogi korowo-rdzeniowe
przedsionkowo – rdzeniowe
siatkowo-rdzeniowe (drogi rozpoczynające się w jądrach mostu oraz drogi
rozpoczynające się w rdzeniu przedłużonym)

Boczne drogi mózgowo-rdzeniowe
Zstępująca bocznie droga korowo-rdzeniowa (KR),
nazywana także drogą piramidową, jest najdłuższą i jedną z
największych w układzie nerwowym
•
składa się na nią prawie milion włókien nerwowych
•
ponad 60% tych włókien to aksony neuronów rozmieszczonych w polu 4 i 6
płata czołowego (kora ruchowa)
•
pozostałe włókna pochodzą z kory czuciowej i ciemieniowej
•
włókna zstępujące z obu półkul mózgowych przechodzą na drugą stronę w
skrzyżowaniu piramid na poziomie rdzenia przedłużonego
•
prawa połowa mózgu kontroluje mięśnie lewej strony ciała i na odwrót - kora
ruchowa lewej półkuli kontroluje mięśnie po prawej stronie ciała
•
włókna szlaku korowo-rdzeniowego łączą się z neuronami ruchowymi i
wstawkowymi leżącymi w tylno-bocznych rejonach rogów przednich rdzenia
•
neurony te kontrolują ruchy dystalnych części kończyn, przy czym
sterowanie korowe ma znacznie większy wpływ na mięśnie zginające

Boczne drogi mózgowo-rdzeniowe
Szlakami bocznymi rdzenia kręgowego zstępują również włókna drogi
czerwienno-rdzeniowej (CzR). Tworzą ją włókna neuronów skupionych
w jądrze czerwiennym śródmózgowia.
•
na poziomie mostu aksony neuronów jądra czerwiennego przechodzą na przeciwległą
stronę i następnie w rdzeniu zstępują wspólnie z włóknami drogi korowo-rdzeniowej
•
jądro czerwienne otrzymuje sygnały wejściowe z tych samych obszarów kory, w których
rozpoczyna się droga korowo-rdzeniowa
•
w trakcie rozwoju ewolucyjnego rola połączeń czerwienno-rdzeniowych została
zredukowana przez zastąpienie ich bezpośrednimi połączeniami korowo-rdzeniowymi
uszkodzenie bezpośrednich i pośrednich połączeń korowo-rdzeniowych
powoduje niedowład przejawiający się znacznym upośledzeniem oraz
spowolnieniem ruchów dowolnych
•
pozbawia możliwości wykonywania niezależnych ruchów w poszczególnych stawach, tzw.
ruchów cząstkowych kończyny górnej
ostry deficyt ruchowy w postaci niedowładu
w wyniku prowadzenia długotrwałej rehabilitacji wiele funkcji ruchowych może powrócić

Zstępujące drogi brzuszno-przyśrodkowe
grupa sygnałów ruchowych dociera do motoneuronów rdzenia
kręgowego drogami brzuszno-przyśrodkowymi
cztery drogi rozpoczynające się w pniu mózgu - ich włókna tworzą połączenia
synaptyczne
z neuronami wstawkowymi rdzenia, które sterują aktywnością neuronów ruchowych
kontrolujących zespoły mięśni osiowych i obręczowych
połączenia:
przedsionkowo-rdzeniowe
pokrywowo-rdzeniowe
dwie drogi siatkowo-rdzeniowe: mostowa oraz rozpoczynająca się w rdzeniu
przedłużonym opuszkowa
Podział funkcjonalny
•
drogi przedsionkowo-rdzeniowe
•
kontrolują położenie głowy w warunkach statycznych i dynamicznych,
•
pozwalają utrzymać stabilną pozycję głowy w czasie lokomocji
•
sygnalizują pozycję oraz ruchy głowy względem linii sił pola grawitacyjnego
•
jądra przedsionkowe integrują sygnały zstępujące z wyższych ośrodków ruchowych
z informacją z mechanoreceptorów narządu przedsionkowego oraz mięśni szyi

Zstępujące drogi brzuszno-przyśrodkowe
•
drogi pokrywowo-rdzeniowe
•
rozpoczyna ją się we wzgórkach górnych śródmózgowia
•
struktury otrzymują bezpośrednią projekcję z siatkówki oka oraz kory wzrokowej
•
otrzymują informacje słuchowe oraz somatosensoryczne
•
na podstawie tych sygnałów we wzgórkach górnych tworzona jest mapa naszego środowiska
•
odpowiedzialne są za reakcję orientacyjną, czyli nakierowanie głowy i oczu na określony cel
•
drogi siatkowo-rdzeniowe - rozpoczynają się w moście
•
kontrolują aktywność posturalną mięśni tułowia i kończyn
•
wzmacniają aktywność antygrawitacyjnych odruchów rdzeniowych
•
wzmaga aktywność mięśni prostowników nóg oraz zginaczy kończyn górnych
•
droga opuszkowo-rdzeniowa
•
sygnały zmniejszają udział kontroli odruchowej w sterowania mięśniami antygrawitacyjnymi
Interakcja aktywności obydwu dróg pozwala na sprawną koordynację
ruchowo-posturalną, zapewniając tym samym stabilność dynamiczną
postawy w czasie wykonywania ruchu

Sygnały przekazywane drogami siatkowo-rdzeniowymi
zaangażowane są
w regulację odpowiedzi na bodźce bólowe
•
odruchowe cofnięcie kończyny w odpowiedzi na bodziec bólowy
•
jednocześnie przez komórki Renshawa sygnały z tworu siatkowego ustalają
wzajemne relacje pobudzenia mięśni antagonistycznych
•
komórki Renshawa hamują zarówno aktywność motoneuronów alfa mięśni
antagonistycznych, jak i neuronów wstawkowych la
•
dzięki temu aktywność neuronów siatkowo-rdzeniowych może modulować
hamowanie wzajemne mięśni antagonistycznych
•
w przypadku uszkodzenia drogi siatkowo-rdzeniowej następuje wzrost
wrażliwości na pobudzenie motoneuronów mięśni antagonistycznych,
którego objawem jest ciągłe pobudzenie tych mięśni

Rola ośrodków korowych mózgu
w kontroli motorycznej
poszczególne części ciała mają swoją reprezentację w obszarze
zakrętu przedśrodkowego (pole 4 według klasyfikacji Brodmanna)
•
reprezentacja jest tym rozleglejsza, im bardziej złożone i precyzyjne ruchy
wykonuje dana część ciała
•
w obszarze kory zespoły neuronów reprezentujące poszczególne części ciała
zorganizowane są w kolumny
•
neurony sterujące dystalną częścią kończyny znajdują się w ich centralnej
części, wokół której warstwowo ułożone są komórki kontrolujące mięśnie bliższe
Poszczególne mięśnie mają zazwyczaj kilka reprezentacji w wielu
różnych kolumnach
•
oprócz reprezentacji somatotopowej w korze ruchowej znajdują się kolumny
reprezentujące różne synergię, - korą ruchową pierwszorzędową (M1)
•
Obszar pola 6 - kora przedruchowa (PMA)
PMA daje silną projekcję do neuronów połączeń korowo-rdzeniowych
kontrolujących mięśnie bliższe i osiowe
•
położony przyśrodkowo w stosunku do PMA obszar - dodatkowa kora
ruchową (SMA)
SMA kontroluje aktywność dalszych mięśni kończyn

Rola ośrodków korowych mózgu
w kontroli motorycznej
obydwa obszary kory przedruchowej są zaangażowane w
planowaniu
i koordynacji obustronnych ruchów dowolnych, które następnie
przez PMA przekazywane są pierwszorzędowej korze ruchowej
jądra postawne oraz decyzyjne neurony wstawkowe
(ang. command interneurons) pola 7 kory ciemieniowej
•
są odpowiedzialne za wybór właściwych programów przy wyborze sterowanie
złożonymi ruchami wymagającymi współdziałania wielu mięśni
•
aktywność jąder podstawowych umożliwia wybór w dodatkowej korze ruchowej
(SMA) odpowiednich podprogramów, które składane są w optymalny program
ruchu w korze przedruchowej (PMA)
•
zestawiony program wysyłany jest do realizacji przez pierwszorzędową korę
ruchową
•
pętlę sprzężeń zwrotnych tworzą kora ruchowa z jądrami podstawnymi i
móżdżkiem, zapewniając płynne przejścia między poszczególnymi programami

Rola ośrodków korowych mózgu
w kontroli motorycznej
•
pierwszorzędowa kora ruchowa jest miejscem integracji programów z kory
przedruchowej
z sygnałami pochodzącymi z jąder wzgórza
•
przez jądro brzuszne boczne wzgórza przekazywane są do kory ruchowej
instrukcje móżdżkowe
•
przez jądro brzuszne tylne przesyłane są informacje proprioceptywne
•
Sygnały te pozwalają na wykonanie całej sekwencji programu
ruchowego oraz wprowadzają ewentualne korekcje błędów
wykonania
•
w górnych neuronach ruchowych pierwotnej kory motorycznej tworzone są
rozkazy ruchowe przekazywane drogą korowo-rdzeniową do motoneuronów
rdzenia kręgowego
•
część sygnałów wyjściowych z kory M1 przesyłana jest również zwrotnie do
wzgórza (jądro brzuszne boczne) i do móżdżku

Planowanie ruchu
Planowanie ruchu jest złożonym procesem nerwowym, w którym
znaczący udział mają jądra podstawne:
•
prążkowie - podzielone przez torebkę wewnętrzną na jądro ogoniaste i łupinę
•
gałka blada
•
jądro niskowzgórzowe
•
istota czarna
•
jądra zaangażowane są także w procesy pamięciowe, poznawcze i motywacyjne
•
połączenia jąder podstawnych tworzą pętle motoryczne, które odgrywają
kluczową rolę w planowaniu i rozpoczęciu ruchu
•
prążkowie otrzymuje sygnały pobudzające (glutaminergiczne) prawie z całej kory
mózgu
•
na tej podstawie wysyłane są do gałki bladej i istoty czarnej sygnały hamujące
•
połączenia GABAergiczne z tych dwóch struktur hamują czynność jąder brzusznych
wzgórza
•
wychodzące z jąder wzgórza połączenia glutaminergiczne pobudzają korę ruchową w
polu 6
•
obydwa obszary kory PMA i SMA otrzymują informacje niezbędne w planowaniu
aktywności ruchowej, wyboru w zależności od motywacji i warunków wykonania ruchu

Koordynacja ruchów
Proces czasowo-przestrzennego składania ruchów prostych w jedną
zharmonizowaną sekwencję nosi nazwę koordynacji ruchowej
•
móżdżek odgrywa kluczową rolę w procesie koordynacji ruchów
•
zawiera ona ponad 50% wszystkich neuronów tworzących ośrodkowy układ nerwowy
Funkcjonalnie móżdżek można podzielić na trzy części:
•
przedsionkowy
•
bierze udział w kontroli postawy i równowagi, koordynacji posturalno-ruchowej, oraz
koordynuje ruch głowy i gałek ocznych
•
oprócz sygnałów z narządów przedsionkowych, docierają również informacje z
siatkówki oka
rdzeniowy
•
uczestniczy w kontroli napięcia mięśni tułowia i proksymalnych części kończyn
•
kontroluje funkcje dystalnych mięśni kończyn
nowy, mostowy
•
jest czynnościowo związany z korą mózgu
•
połączenia korowo-móżdżkowe tworzą pętlę ruchową
•
Informacje z kory mózgu docierają do móżdżku przez dwa neurony: korowo-mostowy,
mostowo-móżdżkowy

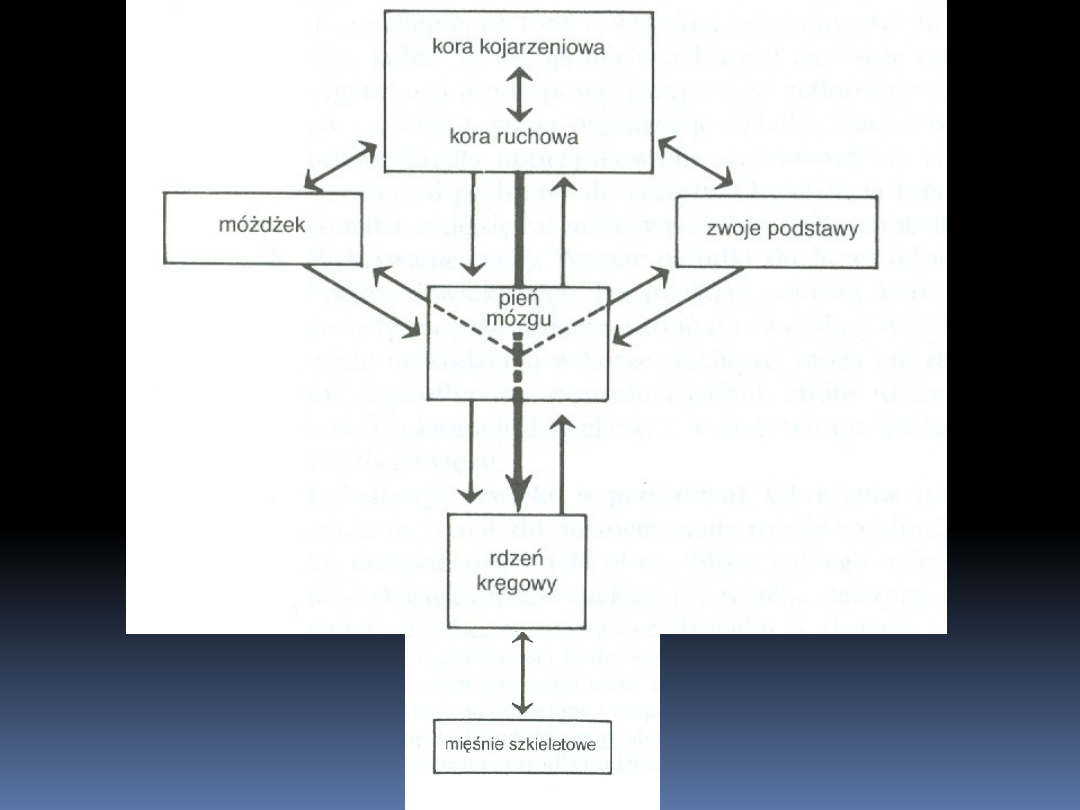
Układ kontroli ruchu
Kora mózgowa jest odpowiedzialna za
planowanie ruchów dowolnych i wysyłanie
poleceń ruchowych inicjujących ich
wykonanie

Układ kontroli ruchu
Ośrodki podkorowe są odpowiedzialne za modulację i koordynację
tych pleceń, aby zadania ruchowe zostały wykonane prawidłowo
Zwoje podstawy dostarczają wzorców ruchowych niezbędnych do
utrzymania postawy ciała koniecznej do prawidłowego
wykonywania poleceń ruchowych.
Móżdżek otrzymuje z kory mózgowej informacje o naturze
zamierzonego ruchu, natomiast z rdzenia kręgowego o tym jak
dalece prawidłowo jest on wykonywany. Informacja ta jest
wykorzystywana w celu odpowiedniego dostosowania poleceń
ruchowych, tak aby zamierzony ruch był wykonywany prawidłowo
Pień mózgu jest wielką stacją przekażnikową dla wszystkich
poleceń ruchowych, z wyjątkiem tych, które wymagają największej
precyzji (są przekazywane bezpośrednio do rdzenia kręgowego).
Pień mózgu jest odpowiedzialny również za utrzymanie normalnej
postawy ciała podczas aktywności ruchowej

Układ kontroli ruchu
Rdzeń kręgowy zawiera wspólne
końcowe drogi, poprzez które ruch
jest wykonywany. Uczestniczy we
właściwym wykonaniu zadania
ruchowego przez wybór właściwych
motoneuronów do poszczególnych
zadań i przez odruchowe
dostosowywanie stopnia ich
aktywności

Układ kontroli ruchu
Receptory zapewniają czuciowe sprzężenie
zwrotne z c.u.n., które może być użyte do
dostosowywania komend ruchowych w czasie
wykonywania ruchu
Informacja nieświadoma o pozycji ciała w
przestrzeni i napięciu wytwarzanym przez
kurczące się mięśnie szkieletowe jest
dostarczana z więzadeł, mięśni i narządów
przedsionkowych
Świadoma informacja o pozycji ciała i kończyn
w przestrzeni jest dostarczana przede
wszystkim przez wzrok i skórne narządy
czuciowe

Układ kontroli ruchu
Jednostka ruchowa jest modułem
czynnościowym wykorzystywanym przez
układ kontroli ruchu do wykonania ruchu
Jednostka ruchowa składa się z
motoneuronu i wszystkich włókien
mięśniowych, które on unerwia.
Włókna mięśniowe w jednostce motorycznej
dzielą się w zależności od ich
charakterystyki histochemicznej i zdolności
kurczenia się

Układ kontroli ruchu
Miocyty białe kurczą się szybko, męczą
łatwo (mała liczba mitochondriów-nie mogą
korzystać z przemiany tlenowej), słabe
unaczynienie
Miocyty czerwone kurczą się wolno i
właściwie się nie męczą, bogate
unaczynienie
Miocyty czerwone szybkie mają cechy
charakterystyczne miocytów białych i
czerwonych

Układ kontroli ruchu
Miocyty białe kurczą się szybko, męczą
łatwo (mała liczba mitochondriów-nie mogą
korzystać z przemiany tlenowej), słabe
unaczynienie
Miocyty czerwone kurczą się wolno i
właściwie się nie męczą, bogate
unaczynienie
Miocyty czerwone szybkie mają cechy
charakterystyczne miocytów białych i
czerwonych

Układ kontroli ruchu
Odruchy skórne
Najważniejszy spośród nich jest odruch cofania
(zgięcia, odruch bólowy), powodujący
odsunięcie części ciała z zasięgu działania
bodżca bólowego.
Receptory tego odruchu są nocyreceptorami
Efektorami są mięśnie szkieletowe, powodujące
cofanie kończyny. Chociaż nazywa się je
zginaczami, są one zginaczami z znaczeniu
fizjologicznym, nie anatomicznym

Układ kontroli ruchu
Odruchy mięśniowe
Odruch na rozciąganie powoduje odruchowy
skurcz rozciąganego mięśnia (odruch kolanowy)
Receptor – wrzecionko mięśniowe; efektory –
zginacze i prostowniki
Odwrócony odruch na rozciąganie
Receptor – narządy ścięgniste Golgiego;
efektory – zginacze i prostowniki
Rola – odruch ochronny, odgrywa istotną rolę w
regulacji napięcia podczas normalnej czynności
mięśniowej.

Kontrola korowa ruchu i
świadomości
Kora mózgowa zawiera obwody neuronalne
odpowiedzialne za koncepcje, planowanie i
tworzenie poleceń ruchowych.
W korze mózgowej zaczynają się dwa
równoległe układy dróg zstępujących:
układ piramidowy (unerwia mięśnie
wykonujące ruchy precyzyjne np. mięśnie
poruszające palcami i ręką oraz mięśnie
związane z mową) i pozapiramidowy
(utrzymuje napięcie posturalne i kieruje
ruchami dowolnymi).

Róg tylny
Sznur boczny
Róg przedni


Odruchy
Reakcje organizmu na specyficzny bodziec,
pobudzający receptory.
Anatomiczne podłoże odruchu – łuk
odruchowy (receptor, droga dośrodkowa,
ośrodek odruchu, droga odśrodkowa,
efektor)
Odruchy monosynaptyczne (odruch na
rozciąganie) i polisynaptyczne (odruch
zginania)


Odruch na rozciąganie jest
wywoływany przez pobudzenie
wrzecion mięśniowych z
których biegną włókna
czuciowe typu Ia bezpośrednio
pobudzające neurony ruchowe.
Odruch na rozciąganie polega
na skurczu rozciąganego
mięśnia

Odruch zginania jest
wywoływany przez pobudzenie
różnych receptorów
znajdujących się w skórze.
Odruch zginania polega na
obronnym zginaniu
podrażnionej kończyny i
jednoczesnym prostowaniu
kończyny przeciwnej

Ośrodkowy Układ Nerwowy ma
szczególne znaczenie w procesie
treningu, gdyż steruje skurczami
mięśni szkieletowych, umożliwiając
realizację ruchów, a także integruje
czynność innych układów,
dostosowując ich pracę do poziomu
realizowanego wysiłku fizycznego.

Document Outline
- Slide 1
- Slide 2
- reakcje ruchowe
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- motoneuron alfa
- Slide 9
- Typy jednostek ruchowych
- Typy jednostek ruchowych
- Kontrola siły skurczu mięśnia
- Kontrola siły skurczu mięśnia sumowanie czasowo-przestrzenne
- Organizacja sterowania pracą mięśni
- Organizacja sterowania pracą mięśni Kontrola obwodowa
- receptory mięśniowe, stawowe i skórne
- receptory mięśniowe, stawowe i skórne wrzeciona mięśniowe
- receptory mięśniowe, stawowe i skórne receptory ścięgnowe
- receptory mięśniowe, stawowe i skórne receptory stawowe
- receptory mięśniowe, stawowe i skórne mechanoreceptory skórne
- receptory mięśniowe, stawowe i skórne mechanoreceptory skórne
- Kontrola ośrodkowa
- Kontrola ośrodkowa
- Boczne drogi mózgowo-rdzeniowe
- Boczne drogi mózgowo-rdzeniowe
- Zstępujące drogi brzuszno-przyśrodkowe
- Zstępujące drogi brzuszno-przyśrodkowe
- Slide 28
- Rola ośrodków korowych mózgu w kontroli motorycznej
- Rola ośrodków korowych mózgu w kontroli motorycznej
- Rola ośrodków korowych mózgu w kontroli motorycznej
- Planowanie ruchu
- Koordynacja ruchów
- Układ kontroli ruchu
- Układ kontroli ruchu
- Układ kontroli ruchu
- Układ kontroli ruchu
- Układ kontroli ruchu
- Układ kontroli ruchu
- Układ kontroli ruchu
- Układ kontroli ruchu
- Układ kontroli ruchu
- Slide 43
- Kontrola korowa ruchu i świadomości
- Slide 45
- Slide 46
- Odruchy
- Slide 48
- Slide 49
- Slide 50
- Slide 51
- Slide 52
Wyszukiwarka
Podobne podstrony:
neurofizjologia ukladu ruchu id Nieznany
NEUROFIZJOLOGIA UKLADU RUCHU, podstawy kinezjologii
neurofizjologia układu ruchu
Podstawy wytrzymałości tkanek układu ruchu człowieka, Biomechanika, biomechanika calosc
WADY WRODZONE UKŁADU RUCHU, fizjo, ortopedia
15 Badanie układu ruchu i nerwowego
Badanie układu ruchu i nerwowego
Choroby układu ruchu
4. Urazy układu ruchu, Anatomia, Chirurgia, chirurgia1
Choroby zawodowe układu ruchu
Choroby układu ruchu wykład
badanie układu ruchu
ćw 1 fizjologia układu ruchu z higieną
SCHORZENIA UKLADU RUCHU, pedagogika
Biomchanika, Biomechanika spr.z wyskku, Możliwości dynamiczne układu ruchu człowieka mogą być ocenia
Biomchanika, nie wiem, Możliwości dynamiczne układu ruchu człowieka mogą być oceniane przez rozpatry
Anatomia układu ruchu, Anatomia
Urazy układu ruchu
Choroby układu ruchu, Prace zaliczeniowe-kosmetyka
więcej podobnych podstron